
Does Temporal and Spatial Diet Alteration Lead to Successful Adaptation of the Eastern Imperial Eagle, a Top Predator?

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
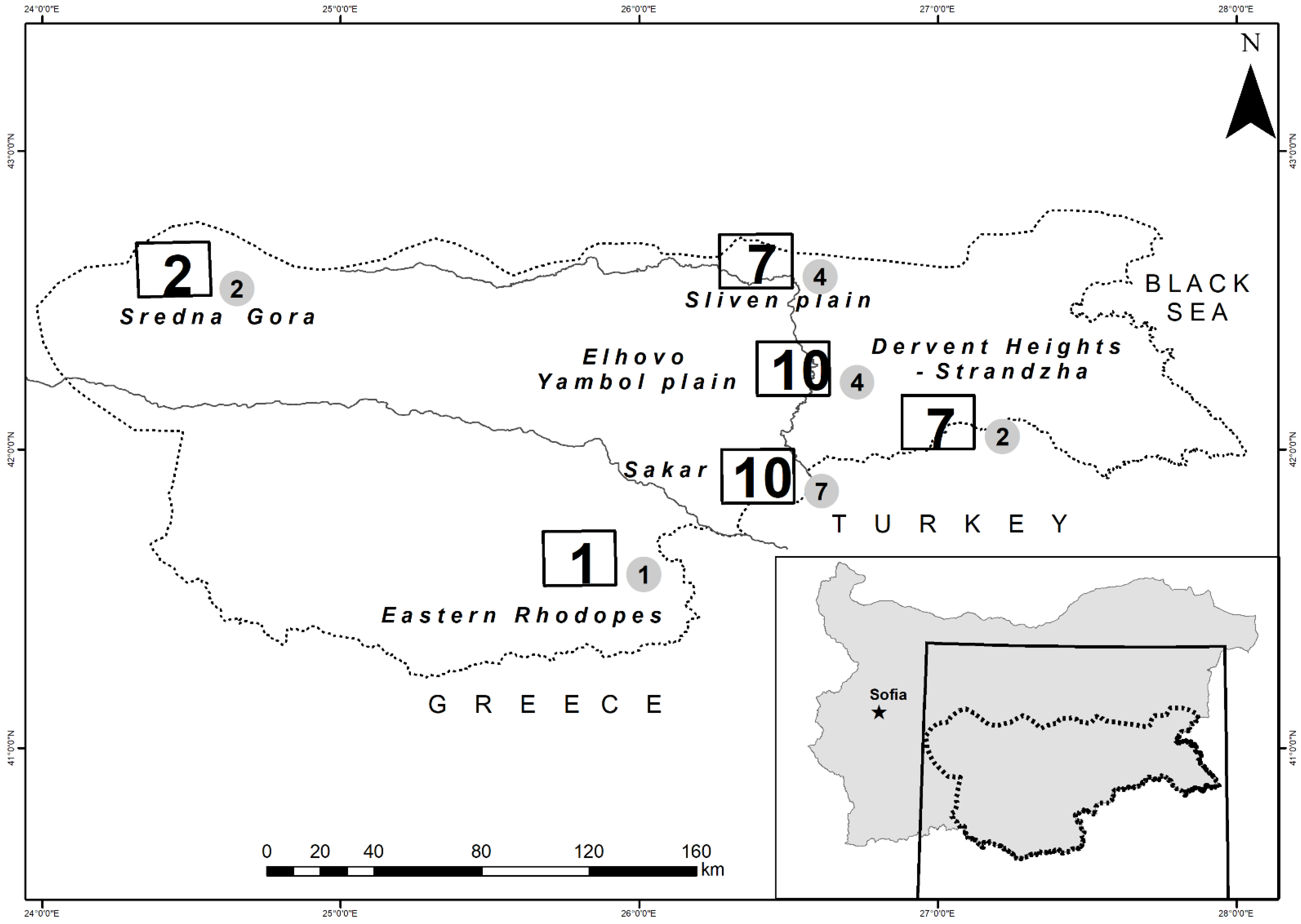
2.1. Study Area
2.2. Data Collection
2.3. Profitable Prey Abundance
2.4. Data Analyses
3. Results
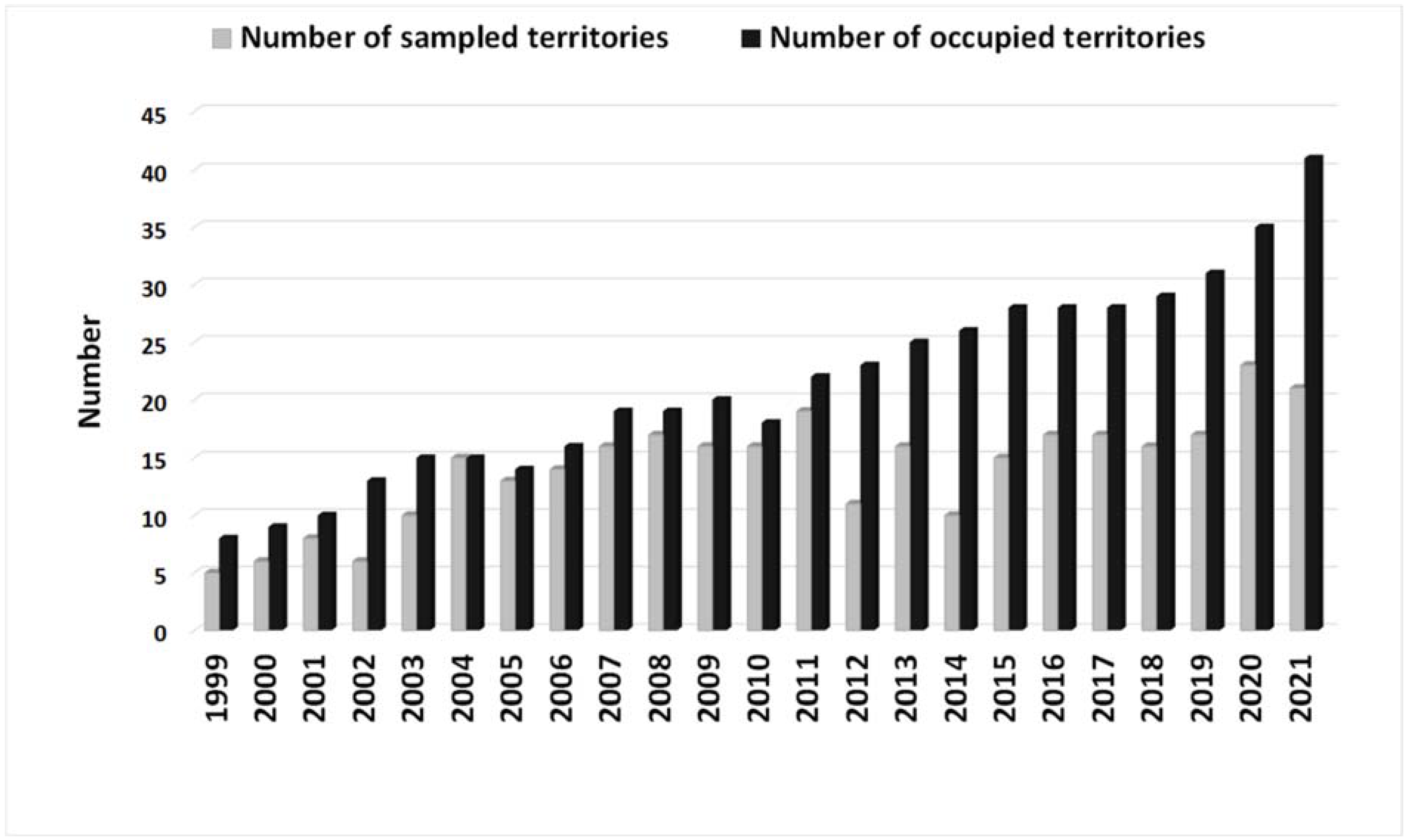
3.1. General Pattern of Main Prey Contribution and EIE Population
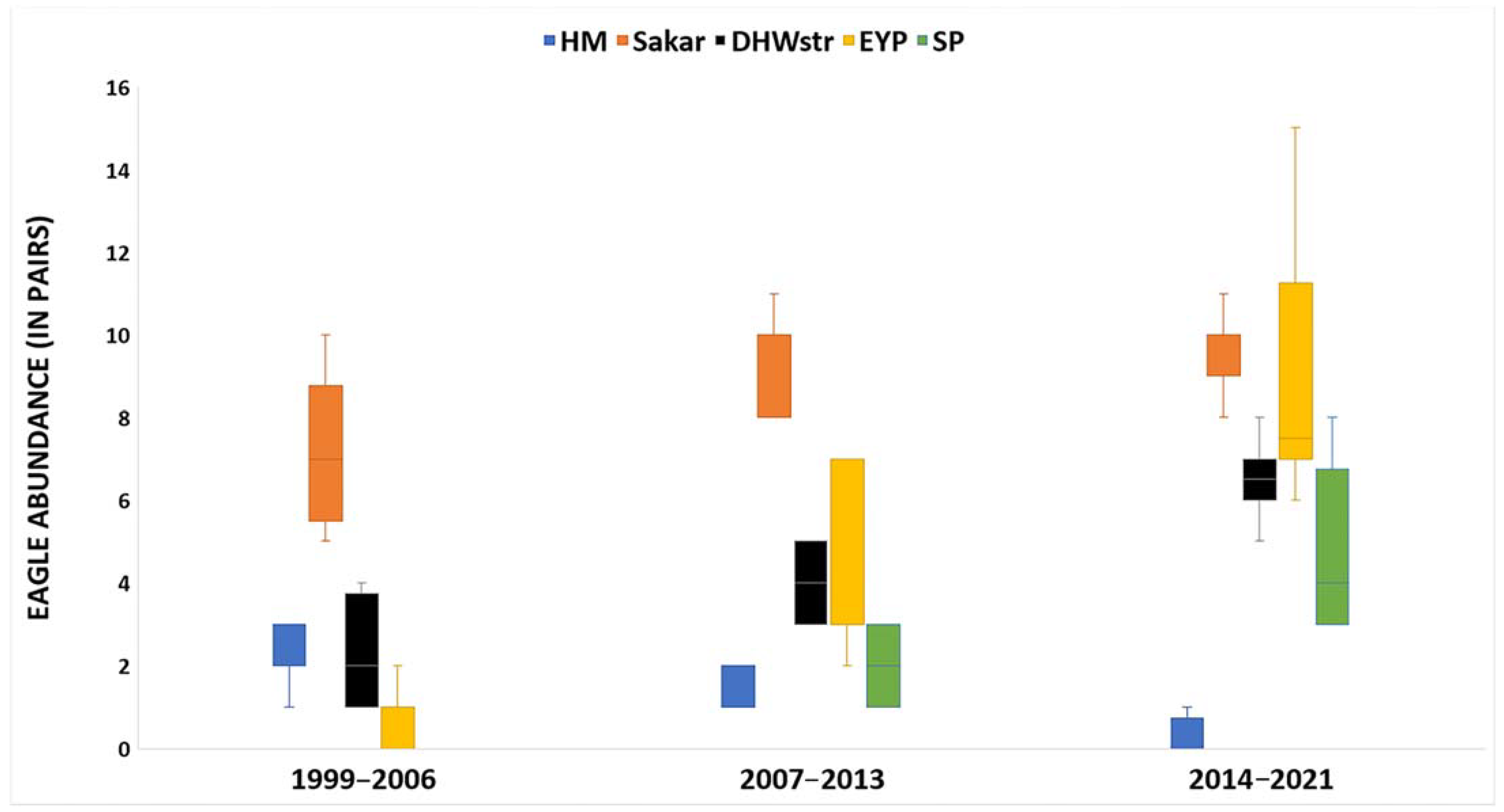
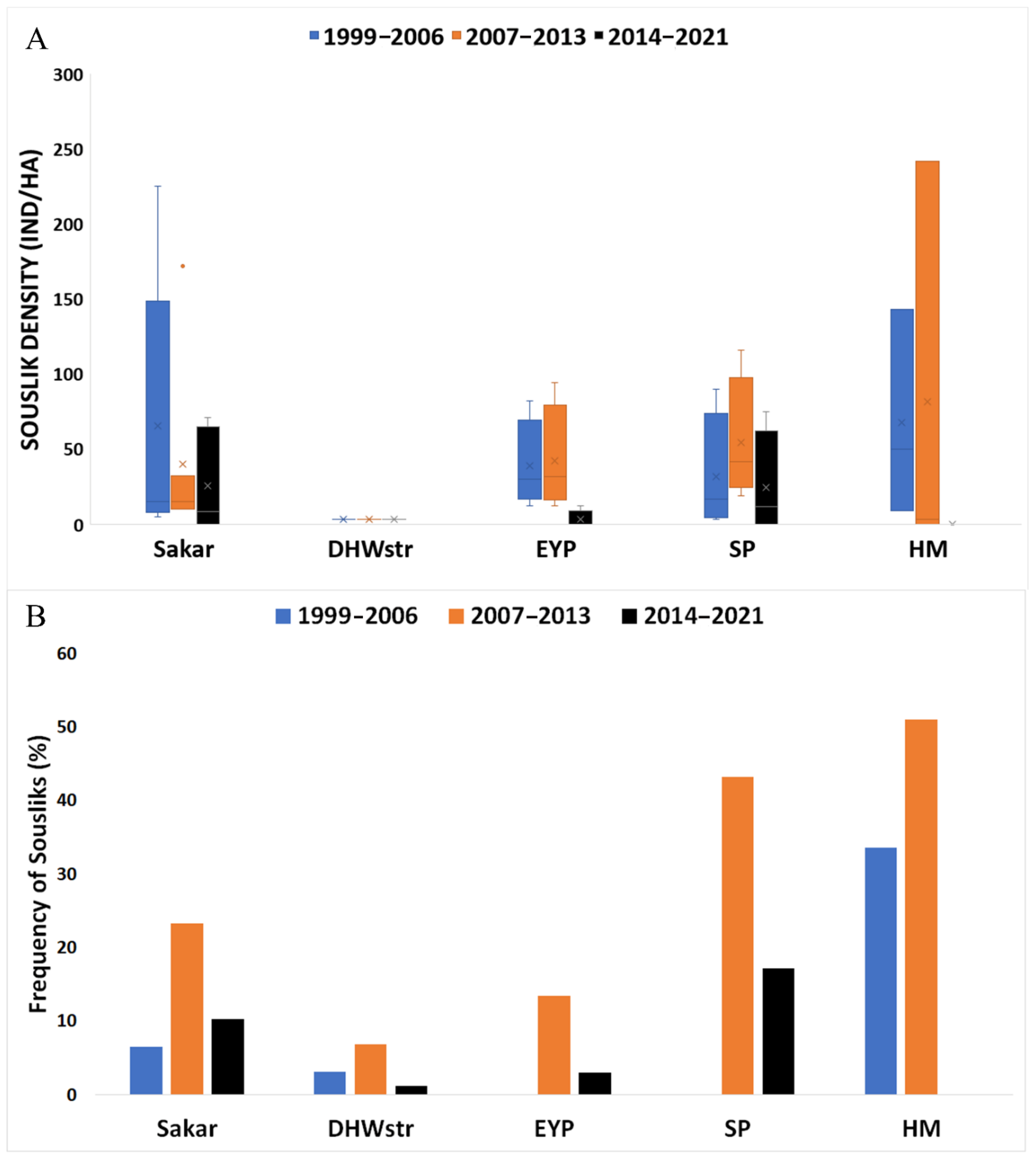
3.2. Temporal and Spatial Comparison of Eagle Abundance, Profitable Prey Abundance, and Diet Composition in the Studied Periods
4. Discussion
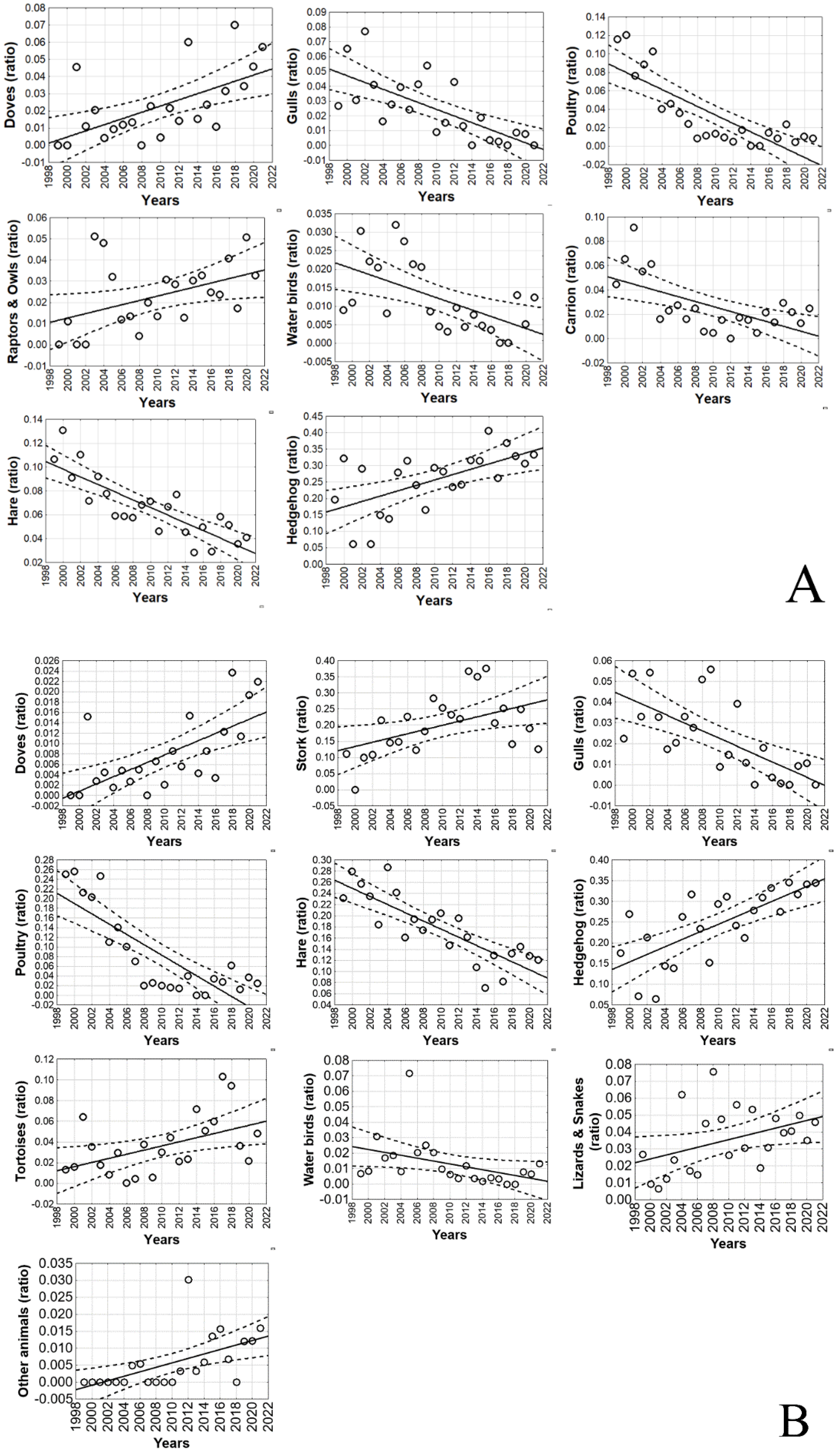
4.1. Long-Term and Large-Scale Changes in the Diet Pattern
4.2. Profitable Prey Abundance Changes and Adaptive Response of EIE
4.3. Effects of Diet Alteration and Conservation Suggestions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton, I. Population Limitation in Birds; Academic Press: Cambridge, MA, USA, 1998; p. 597. [Google Scholar]
- Reynolds, R.T.; Lambert, J.S.; Kay, S.L.; Sanderlin, J.S.; Bird, B.J. Factors affecting lifetime reproduction, long-term territory-specific reproduction, and estimation of habitat quality in northern goshawks. PLoS ONE 2019, 14, e0215841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Väli, Ü. Factors limiting reproductive performance and nestling sex ratio in the Lesser Spotted Eagle Aquila pomarina at the northern limit of its range: The impact of weather and prey abundance. Acta Ornithol. 2012, 47, 157–168. [Google Scholar] [CrossRef]
- Lourenço, R.; Delgado, M.D.M.; Campioni, L.; Korpimäki, E.; Penteriani, V. Evaluating the influence of diet-related variables on breeding performance and home range behaviour of a top predator. Popul. Ecol. 2015, 57, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Senior, A.M.; Nakagawa, S.; Lihoreau, M.; Simpson, S.J.; Raubenheimer, D. An overlooked consequence of dietary mixing: A varied diet reduces interindividual variance of fitness. Am. Nat. 2015, 186, 649–659. [Google Scholar] [CrossRef] [Green Version]
- Fatuyma, D.J.; Moreno, G. The evolution of ecological specialization. Ann. Rev. Ecol. Syst. 1988, 19, 207–233. [Google Scholar] [CrossRef]
- Whitfield, D.P.; Reid, R.; Haworth, P.F.; Madders, M.; Marquiss, M.; Tingay, R.; Fielding, A.H. Diet specificity is not associated with increased reproductive performance of golden eagles Aquila chrysaetos in western Scotland. Ibis 2009, 151, 255–264. [Google Scholar] [CrossRef]
- Bolnick, D.I.; Svanbäck, R.; Fordyce, J.A.; Yang, L.H.; Davis, J.M.; Hulsey, C.D.; Forister, M.L. The ecology of individuals: Incidence and implications of individual specialization. Am. Nat. 2003, 161, 1–28. [Google Scholar] [CrossRef]
- Murgatroyd, M.; Avery, G.; Underhill, L.G.; Amar, A. Adaptability of a specialist predator: The effects of land use on diet diversification and breeding performance of Verreaux’s eagles. J. Avian Biol. 2016, 47, 001–012. [Google Scholar] [CrossRef]
- Moleón, M.; Sánchez-Zapata, J.A.; Gil-Sánchez, J.M.; Ballesteros-Duperón, E.; Barea-Azcón, J.M.; Virgós, E. Predator-prey relationships in a Mediterranean vertebrate system: Bonelli’s eagles, rabbits and partridges. Oecologia 2012, 168, 679–689. [Google Scholar] [CrossRef]
- Pires, M.M.; Guimarães, P.R., Jr.; Araújo, M.S.; Giaretta, A.A.; Costa, J.C.L.; dos Reis, S.F. The nested assembly of individual-resource networks. J. Anim. Ecol. 2011, 80, 896–903. [Google Scholar] [CrossRef]
- Svanbäck, R.; Bolnick, D.I. Intraspecific competition affects the strength of individual specialization: An optimal diet theory method. Evol. Ecol. Res. 2005, 7, 993–1012. [Google Scholar]
- Levin, S.A. The problem of pattern and scale in ecology. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Hanski, I.; Henttonen, H.; Korpimäki, E.; Oksanen, L.; Turchin, P. Small-rodent dynamics and predation. Ecology 2001, 82, 1505–1520. [Google Scholar] [CrossRef]
- Koenig, W.D.; Liebhold, A.M. Effects of periodical cicada emergences on abundance and synchrony of avian populations. Ecology 2005, 86, 1873–1882. [Google Scholar] [CrossRef] [Green Version]
- Milchev, B. Diet shifting of tortoise-eating Golden Eagles (Aquila chrysaetos) in southeastern Bulgaria. Ornis Fenn. 2022, 99, 60–70. [Google Scholar] [CrossRef]
- Moleón, M.; Sánchez-Zapata, J.A.; Real, J.; Garćia-Charton, J.A.; Gil-Sánchez, J.M.; Palma, L.; Bautista, J.; Bayle, P. Large-scale spatio-temporal shifts in the diet of a predator mediated by an emerging infectious disease of its main prey. J. Biogeogr. 2009, 36, 1502–1515. [Google Scholar] [CrossRef]
- Angelstam, P.; Lindström, E.; Widen, P. Synchronous short-term population fluctuations of some birds and mammals of Fennoscandia—Occurrence and distribution. Holarct. Ecol. 1985, 8, 285–298. [Google Scholar] [CrossRef]
- Moleón, M.; Almaraz, P.; Sánchez-Zapata, J.A. An emerging infectious disease triggering large-scale hyper predation. PLoS ONE 2008, 3, e2307. [Google Scholar] [CrossRef]
- Del Hoyo, J.; Elliott, A.; Sargatal, J. Handbook of the Birds of the World: New World Vultures to Guineafowl; Lynx Edicions: Barcelona, Spain, 2007; Volume 2. [Google Scholar]
- Chavko, J.; Danko, Š.; Obuch, J.; Mihók, J. The Food of the Imperial Eagle (Aquila heliaca) in Slovakia. Slov. Raptor J. 2007, 1, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Demerdzhiev, D.; Boev, Z.; Dobrev, D.; Terziev, N.; Nedyalkov, N.; Stoychev, S.; Petrov, T. Diet of Eastern Imperial Eagle (Aquila heliaca) in Bulgaria: Composition, distribution and variation. Biodivers. Data J. 2022, 10, e77746. [Google Scholar] [CrossRef]
- Demerdzhiev, D.; Dobrev, D.; Isfendiyaroğlu, S.; Boev, Z.; Stoychev, S.; Terziev, N.; Spasov, S. Distribution, abundance, breeding parameters, threats, and prey preferences of the eastern imperial eagle (Aquila heliaca) in European Turkey. Slov. Raptor J. 2014, 8, 17–25. [Google Scholar] [CrossRef]
- Horváth, M.; Solti, B.; Fatér, I.; Juhász, T.; Haraszthy, L.; Szitta, T.; Ballók, Z.; Pásztory-Kovács, S. Temporal changes in the diet composition of the Eastern Imperial Eagle (Aquila heliaca) in Hungary. Ornis Hung. 2018, 1, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Horváth, M.; Szitta, T.; Firmánszky, G.; Solti, B.; Kovács, A.; Moskát, C. Spatial variation in prey composition and its possible effect on reproductive success in an expanding Eastern Imperial Eagle (Aquila heliaca) population. Acta Zool. Acad Sci. Hung. 2010, 56, 187–200. [Google Scholar]
- Katzner, T.; Bragin, E.; Knick, S.; Smith, A. Spatial structure in the diet of Eastern Imperial Eagle Aquila heliaca in Kazakhstan. J. Avian Biol. 2006, 37, 594–600. [Google Scholar] [CrossRef]
- Karyakin, I.; Kovalenko, A.; Levin, A.; Pazhenkov, A. Eagles of the Aral–Caspian Region, Kazakhstan. Raptors Conserv. 2011, 22, 92–152. [Google Scholar]
- Karyakin, I.; Nikolenko, E.; Levin, A.; Kovalenko, A. Imperial Eagle in Russia and Kazakhstan: Population status and trends. Raptors Conserv. 2008, 14, 18–27. [Google Scholar]
- Abuladze, A. Ecology of the Imperial Eagle Aquila heliaca in Georgia; Meyburg, B.-U., Chancellor, R.D., Eds.; Eagle Studies; WWGBP: Berlin, Germany; London, UK; Paris, France, 1996; pp. 447–457. [Google Scholar]
- Demerdzhiev, D. Eastern Imperial Eagle (Aquila heliaca heliaca Savigny, 1809) (Accipitridae–Aves) in Bulgaria—Distribution, Biology, Ecology, Numbers, and Conservation Measures; Synopsis of the Ph.D. Thesis; BAS-NMNH: Sofia, Bulgaria, 2011; (In Bulgarian, with English summary). [Google Scholar]
- Katzner, T.; Bragin, E.; Knick, S.; Smith, A. Relationship between demographics and diet specificity of Eastern Imperial Eagle Aquila heliaca in Kazakhstan. Ibis 2005, 147, 576–586. [Google Scholar] [CrossRef]
- Aulagner, S.; Haffner, R.; Mitchell-Jones, A.; Moutou, F.; Zima, J. Mammals of Europe, North Africa and the Middle East; Christopher Helm: London, UK, 2009. [Google Scholar]
- Boev, Z. Comparative Morphological Studies of Herons (Family Ardeidae–Aves) from Bulgaria. Ph.D. Thesis, Sofia University St. Kliment Ohridski, Sofia, Bulgaria, 1986; 193p. (In Bulgarian). [Google Scholar]
- Böhme, W. (Ed.) Askulapnatter (E. longisima Laurenti 1768). In Handbuch der Reptilien und Amphibien Europas; Aula Verlag: Weisbaden, Germany, 1993; pp. 331–372. [Google Scholar]
- Dunning, J. (Ed.) Handbook of Avian Body Masses; CRC Press: Abingdon, UK, 2008. [Google Scholar]
- Kunstmüller, I. The food ecology and the diet composition of Eagle Owl (Bubo bubo) at the breeding sites in the Bohemian-Moravian highlands, Czech Republic. Crex 2000, 16, 50–59. [Google Scholar]
- Petrov, P. Variability in the body mass of European Hare (Lepus europaeus, Pall.) in Bulgaria. Forest Sci. 1964, 1, 73–81. (In Bulgarian) [Google Scholar]
- Simeonov, S.; Michev, T.; Nankinov, D. Fauna of Bulgaria: Aves, 1; BAS: Sofia, Bulgaria, 1990; Volume 20. (In Bulgarian) [Google Scholar]
- Simeonov, S.; Petrov, T.S. Studying the diet of Eastern Imperial Eagle (Aquila heliaca Savigny), Common Buzzard (Buteo buteo L.), and Rough-legged Buzzard (Buteo lagopus Brünnich) in Bulgaria. Ecology 1980, 7, 22–30. (In Bulgarian) [Google Scholar]
- Koshev, Y. Distribution and status of the European Ground Squirrel (Spermophilus citellus) in Bulgaria. Lynx 2008, 39, 251–261. [Google Scholar]
- Demerdzhiev, D.; Dobrev, D.; Popgeorgiev, G.; Stoychev, S. Landscape alteration affects the demography of an endangered avian predator by reducing the habitat quality. Avian Res. 2022, 13, 100030. [Google Scholar] [CrossRef]
- Shapiro, S.; Wilk, M.; Chen, H. A comparative study of various tests of normality. J. Am. Stat. Assoc. 1968, 63, 1343–1372. [Google Scholar] [CrossRef]
- StatSoft Inc. STATISTICA (Data Analysis Software System), Version 12. 2013. Available online: www.statsoft.com (accessed on 12 March 2019).
- Marin, S.; Ivanov, I.; Georgiev, D.; Boev, Z. On the Food of the Imperial Eagle Aquila heliaca on Sakar Mountain and Dervent Heights, Bulgaria; Chancellor, R., Meyburg, B.-U., Eds.; Raptors Worldwide; WWGBP/MME: Towcester, UK, 2004; pp. 589–592. [Google Scholar]
- Zhelev, C.H.; Ninov, N.; Mihaylov, H.; Gruychev, G.; Stoyanov, S.; Mirchev, R. Density of Brown Hare (Lepus europaeus Pallas, 1778) in the plain habitats of Bulgaria. In Proceedings of the 2nd International Symposium on Hunting, Modern Aspects of Sustainable Management of Game Populations, Novi Sad, Serbia, 17–20 October 2013. [Google Scholar]
- Godfroid, J.; Cloeckaert, A.; Liautard, J.-P.; Kohler, S.; Fretin, D.; Walravens, K.; Garin-Bastuji, B.; Letesson, J.-J. From the discovery of the Malta fever’s agent to the discovery of a marine mammal reservoir, brucellosis has continuously been a re-emerging zoonosis. Vet. Res. 2005, 36, 313–326. [Google Scholar] [CrossRef] [Green Version]
- Wibbelt, G.; Frölich, K. Infectious diseases in European Brown Hare (Lepus europaeus). Wildl. Biol. Pract. 2005, 1, 86–93. [Google Scholar] [CrossRef]
- Margalida, A.; Moleón, M. Toward carrion-free ecosystems? Front. Ecol. Environ. 2016, 14, 183–184. [Google Scholar]
- Pyke, G.H.; Pulliam, R.; Charnov, E.L. Optimal foraging: A selective review of theory and tests. Q. Rev. Biol. 1977, 52, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Stephens, D.W.; Krebs, J.R. Foraging Theory; Princeton University Press: Princeton, NJ, USA, 1986. [Google Scholar]
- Bekmansurov, R.H.; Karyakin, I.V. Results of Monitoring of the Imperial Eagle Population in the Republic of Tatarstan in 2011–2012, Russia. Raptors Conserv. J. 2013, 26, 84–108. [Google Scholar]
- Sokolov, A.Y.; Sarychev, V.S.; Vlasov, A.A. Species of Aquila and Haliaeetus in the Breeding Fauna of the Central Black Earth Region: Current Status and Prospects for Existence. Raptors Conserv. J. 2019, 38, 109–126. [Google Scholar] [CrossRef]
- Angelstam, P.; Lindström, E.; Widen, P. Role of predation in short-term population fluctuations of some birds and mammals in Fennoscandia. Oecologia 1984, 62, 199–208. [Google Scholar] [CrossRef]
- Lazarova, I.; Dobrev, D.; Gradev, G.; Petrov, R.; Stoychev, S.; Klisurov, I.; Demerdzhiev, D. Main mortality factors for the Eastern Imperial Eagle (Aquila heliaca Savigny, 1809) in Bulgaria. Ornis Hung. 2020, 28, 120–134. [Google Scholar] [CrossRef]
- Demerdzhiev, D. Birds in Besaparski Ridove special protection area (Natura 2000), Southern Bulgaria: Conservation status and dynamics. Acta Zool. Bulg. 2014, 66 (Suppl. 5), 171–189. [Google Scholar]
- Demerdzhiev, D. Breeding parameters and factors influencing the reproduction of an expanding Long-legged Buzzard (Buteo rufnus) population under high breeding density conditions. J. Ornithol. 2022, 163, 405–415. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regions | Number of Sampled Occupied Territories | Number of Sampled Prey Specimen |
|---|---|---|
| HM (Sredna Gora Mnt. and Eastern Rhodope Mnt.) | 3 | 456 |
| Sakar Mnt. | 10 | 2090 |
| DHWstr (Dervent Heights-Western foothills of Strandzha Mnt.) | 7 | 1239 |
| EYP (Elhovo-Yambol plain) | 10 | 916 |
| SP (Sliven plain) | 7 | 614 |
| TOTAL | 37 | 5315 |
| Prey Categories | Frequency | Biomass | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Adjusted R2 | F (1.21) | β2 | LCL/UCL | p | Adjusted R2 | F (1.21) | β2 | LCL/UCL | p | |
| Lizards and snakes | 0.05 | 2.05 | 0.30 | −0.14/0.73 | 0.17 | 0.14 | 4.51 | 0.42 | 0.009/0.83 | 0.046 |
| Tortoises | 0.12 | 4.11 | 0.40 | −0.01/0.82 | 0.055 | 0.20 | 6.62 | 0.49 | 0.09/0.89 | 0.02 |
| Water birds | 0.29 | 10.16 | −0.57 | −0.94/−0.20 | 0.004 | 0.14 | 4.46 | −0.42 | −0.83/−0.006 | 0.047 |
| Poultry | 0.64 | 40.19 | −0.81 | −1.08/−0.54 | <0.001 | 0.65 | 42.36 | −0.82 | −1.08/−0.56 | <0.001 |
| Phasianids | 0.01 | 0.70 | 0.18 | −0.27/0.63 | 0.41 | 0.01 | 1.16 | 0.23 | −0.21/0.67 | 0.29 |
| Gulls | 0.48 | 21.57 | −0.71 | −1.03/−0.39 | <0.001 | 0.44 | 18.08 | −0.68 | −1.01/−0.35 | <0.001 |
| Doves | 0.33 | 11.91 | 0.60 | 0.24/0.96 | 0.002 | 0.42 | 16.61 | 0.66 | 0.33/1.00 | <0.001 |
| Songbirds | 0.05 | 0.03 | −0.04 | −0.49/0.41 | 0.86 | 0.05 | 0.02 | 0.03 | −0.42/0.48 | 0.89 |
| Corvids | 0.02 | 0.51 | 0.15 | −0.29/0.60 | 0.48 | 0.04 | 0.18 | 0.09 | −0.36/0.54 | 0.68 |
| Stork | 0.09 | 3.23 | 0.36 | −0.06/0.79 | 0.09 | 0.20 | 6.48 | 0.49 | 0.09/0.88 | 0.02 |
| Raptors and owls | 0.16 | 5.13 | 0.44 | 0.04/0.85 | 0.03 | 0.07 | 2.73 | 0.34 | −0.09/0.77 | 0.11 |
| Hedgehog | 0.34 | 12.41 | 0.61 | 0.25/0.97 | 0.002 | 0.50 | 22.95 | 0.72 | 0.41/1.04 | <0.001 |
| Hare | 0.67 | 44.79 | −0.83 | −1.08/−0.57 | <0.001 | 0.67 | 46.19 | −0.83 | −1.08/−0.58 | <0.001 |
| Souslik | 0.03 | 0.41 | −0.14 | −0.59/0.31 | 0.53 | 0.04 | 0.20 | −0.10 | −0.55/0.35 | 0.66 |
| Rodentia (excluding souslik) | 0.03 | 0.45 | −0.14 | −0.59/0.30 | 0.51 | 0.05 | 0.01 | −0.005 | −0.46/0.45 | 0.98 |
| Carnivores | 0.01 | 1.16 | −0.23 | −0.67/0.21 | 0.29 | 0.05 | 0.01 | 0.02 | −0.43/0.47 | 0.92 |
| Carrion | 0.35 | 12.70 | −0.61 | −0.97/−0.26 | 0.002 | NA | NA | NA | NA | NA |
| Other animals | 0.11 | 3.64 | 0.38 | −0.035/0.80 | 0.07 | 0.30 | 10.47 | 0.58 | 0.21/0.95 | 0.004 |
| Categories | Effect | Region | β2 | Std. Err | LCL/UCL | t | p |
|---|---|---|---|---|---|---|---|
| Souslik abundance | Random | Sakar Mnt. | 0.46 | 0.20 | −0.07/0.86 | 2.38 | 0.02 |
| Random | SP | 0.38 | 0.18 | −0.01/0.74 | 2.09 | 0.04 | |
| Random | EYP | 0.30 | 0.18 | −0.06/0.66 | 1.66 | 0.10 | |
| Random | HM | 0.24 | 0.16 | −0.07/0.56 | 1.55 | 0.13 | |
| Random | DHWstr | no tolerance | no tolerance | no tolerance | no tolerance | no tolerance | |
| EIE abundance | Random | Sakar Mnt. | 0.66 | 0.08 | 0.51/0.81 | 8.73 | <0.001 |
| Random | SP | no tolerance | no tolerance | no tolerance | no tolerance | no tolerance | |
| Random | EYP | 0.26 | 0.08 | 0.10/0.41 | 3.36 | 0.001 | |
| Random | HM | −0.11 | 0.08 | −0.26/0.04 | −1.44 | 0.15 | |
| Random | DHWstr | 0.32 | 0.08 | 0.17/0.47 | 4.23 | <0.001 |
| Prey Categories | 1999–2006 | 2007–2013 | 2014–2021 | Total (1999–2021) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | % N | % Biomass | Number | % N | % Biomass | Number | % N | % Biomass | Number | % N | % Biomass | |
| Lizards and snakes | 63 | 4.88 | 2.47 | 154 | 7.82 | 4.83 | 133 | 6.47 | 3.96 | 350 | 6.59 | 3.91 |
| Tortoises | 17 | 1.32 | 2.04 | 28 | 1.42 | 2.20 | 73 | 3.55 | 6.01 | 118 | 2.22 | 3.64 |
| Water birds | 24 | 1.86 | 2.25 | 21 | 1.07 | 1.16 | 11 | 0.54 | 0.47 | 56 | 1.05 | 1.16 |
| Poultry | 77 | 5.97 | 16.53 | 26 | 1.32 | 3.10 | 18 | 0.88 | 2.52 | 121 | 2.28 | 6.22 |
| Phasianids | 24 | 1.86 | 0.87 | 30 | 1.52 | 0.80 | 47 | 2.29 | 1.12 | 101 | 1.90 | 0.94 |
| Gulls | 42 | 3.26 | 2.98 | 57 | 2.89 | 2.99 | 11 | 0.54 | 0.55 | 110 | 2.07 | 2.04 |
| Doves | 12 | 0.93 | 0.29 | 38 | 1.93 | 0.65 | 74 | 3.60 | 1.30 | 124 | 2.33 | 0.81 |
| Songbirds | 50 | 3.88 | 0.30 | 70 | 3.55 | 0.23 | 75 | 3.65 | 0.30 | 195 | 3.67 | 0.27 |
| Corvids | 36 | 2.79 | 1.30 | 64 | 3.25 | 1.50 | 83 | 4.04 | 1.76 | 183 | 3.44 | 1.55 |
| Stork | 59 | 4.57 | 13.88 | 143 | 7.26 | 23.82 | 144 | 7.01 | 23.21 | 346 | 6.51 | 21.11 |
| Raptors and owls | 28 | 2.17 | 1.43 | 35 | 1.78 | 1.14 | 66 | 3.21 | 1.69 | 129 | 2.43 | 1.43 |
| Hedgehog | 237 | 18.37 | 18.46 | 493 | 25.03 | 24.88 | 653 | 31.78 | 31.72 | 1383 | 26.02 | 25.93 |
| Hare | 103 | 7.98 | 22.38 | 123 | 6.24 | 18.03 | 83 | 4.04 | 11.28 | 309 | 5.81 | 16.50 |
| Souslik | 146 | 11.32 | 3.90 | 431 | 21.88 | 7.92 | 141 | 6.86 | 2.44 | 718 | 13.51 | 4.80 |
| Rodents | 262 | 20.31 | 1.07 | 147 | 7.46 | 0.60 | 295 | 14.36 | 0.99 | 704 | 13.25 | 0.87 |
| Carnivores | 55 | 4.26 | 9.58 | 45 | 2.28 | 5.64 | 69 | 3.36 | 9.56 | 169 | 3.18 | 8.14 |
| Carrion | 45 | 3.49 | NA | 24 | 1.22 | NA | 34 | 1.65 | NA | 103 | 1.94 | NA |
| Other animals | 10 | 0.77 | 0.28 | 41 | 2.08 | 0.50 | 45 | 2.19 | 1.13 | 96 | 1.81 | 0.69 |
| Total | 1290 | 100.00 | 100.00 | 1970 | 100.00 | 100.00 | 2055 | 100.00 | 100.00 | 5315 | 100.00 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demerdzhiev, D.; Boev, Z.; Dobrev, D.; Nedyalkov, N.; Petrov, T. Does Temporal and Spatial Diet Alteration Lead to Successful Adaptation of the Eastern Imperial Eagle, a Top Predator? Diversity 2022, 14, 1000. https://doi.org/10.3390/d14111000
Demerdzhiev D, Boev Z, Dobrev D, Nedyalkov N, Petrov T. Does Temporal and Spatial Diet Alteration Lead to Successful Adaptation of the Eastern Imperial Eagle, a Top Predator? Diversity. 2022; 14(11):1000. https://doi.org/10.3390/d14111000
Chicago/Turabian StyleDemerdzhiev, Dimitar, Zlatozar Boev, Dobromir Dobrev, Nedko Nedyalkov, and Tseno Petrov. 2022. "Does Temporal and Spatial Diet Alteration Lead to Successful Adaptation of the Eastern Imperial Eagle, a Top Predator?" Diversity 14, no. 11: 1000. https://doi.org/10.3390/d14111000