1. Introduction
The species
Cottus quadricornis Linnaeus, 1758 (common name: fourhorn sculpin), was described from the Baltic Sea. Later, this species was included in the list of fishes of the White Sea [
1]. In the faunistic summary of the fishes of the Russian Empire, the fourhorn sculpin is designated as
Myoxocephalus qufadricornis [
2]. The main morphological feature that distinguishes this species from other species of the genus
Myoxocephalus is the presence of four large bone outgrowths (mounds) on the head. A comparative analysis of the morphological characters, conducted by Leo Berg (1923), revealed differences between the White Sea form of the fourhorn sculpin and the Baltic Sea one [
3].
Leo Berg (1923, 1949) estimated these differences at the level of various subspecies of
Myoxocephalus quadricornis [
3,
4]. The form from the Baltic Sea retained the name of the nominal subspecies
M.
quadricornis quadricornis, and the fourhorn sculpin from the White Sea was
M. quadricornis labradoricus. The species with the name
Acanthocottus labradoricus was described by C. F. Girard (1850) from Yankee Harbor. The text of the description was published in an article by H. R. Storer (1850) [
5]. Specimens of
Acanthocottus labradoricus from the region of Labrador had two pairs of postorbital outgrowths on the head [
6]. This morphological feature occurs in the species
Myoxocephalus scorpius and is never seen in
M. quadricornis. This fact formed the basis to consider
A. labradoricus a synonym of
M. scorpius [
6].
Studies of the lateral line system in the Baltic and White Sea forms of the fourhorn sculpin revealed significant differences in its pattern with other species of the genus
Myoxocephalus [
7]. The main differences were in the morphology of canaliculi extending from the sensory canals. In the Baltic and White Sea specimens, single canaliculus has one pore [
7]. Due to this morphological feature, each sensory canal has a small total number of pores. For example, a fourhorn sculpin from the Baltic Sea has one pore in the coronal commissure. In other species of the genus
Myoxocephalus (
M. stelleri,
M. brandtii,
M. polyacanthocephalus,
M. jaok,
M. verrucosus,
M. tuberculatus,
M. ochotoensis,
M. scorpioides. M. octodecemspinosus,
M. aeneus, and
M. niger), canaliculus is branched and contains multiple pores [
7]. While the Baltic fourhorn sculpin has only one pore in the coronal commissure,
M. stelleri (type species of the genus
Myoxocephalus) has 14 pores. These differences served as the basis for the transfer of
M. quadricormus to the genus
Triglopsis Girard, 1881 [
7]. In studies of fishes of the Russian Arctic, Anatoliy Andriashev and Natalia Chernova (1994) as well as Igor Chereshnev (1996) assigned the fourhorn sculpins to the genus
Triglopsis [
8,
9]. The level of differences between them was assessed as the rank of subspecies. The White Sea subspecies was named
Triglopsis quadricornis polaris [
10], but this name is unacceptable. Molecular genetic studies of mtDNA genes were conducted for the Baltic form of
T. quadricornis [
11]. The complete mitochondrial genome was sequenced for the White Sea form [
12]. Comparative analysis of the genetic data of these two forms was not carried out. The study is devoted to elucidating the morphological and genetic differences between two forms of the fourhorn sculpin from the Baltic and White Seas in order to establish their taxonomic status.
3. Result
3.1. Morphological Characteristics
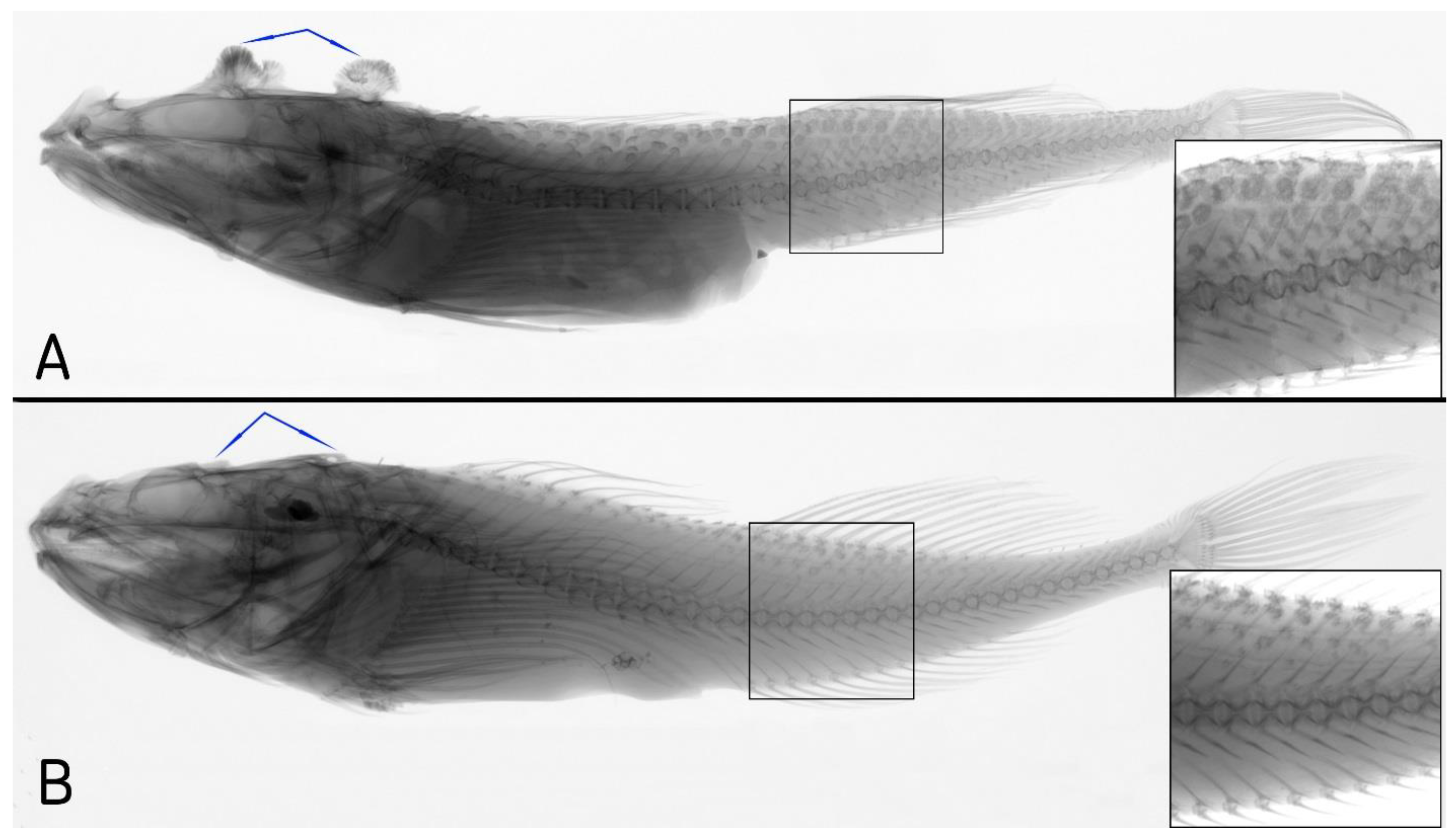
The fourhorn sculpin
T. quadricornis from the Baltic Sea (a type locality) has a characteristic complex of morphological features. There is an abundance on the body of rounded flaps with numerous small prickles at the top. The flaps form four rows and cover a major portion of the body (
Figure 2A and
Figure 3A). The caudal peduncle is long and narrow in its depth, on average 2.5 times less than its length. The head is large, 36%
SL on average, flattened in the dorso-ventral direction. On the top surface of the head, there are four mushroom-shaped bony outgrowths. They have a spongy bone texture with numerous prickles forming into groups. The anterior pair of postorbital outgrowths is smaller than the occipital pair. Their diameters are 6% and 8.3%
SL, respectively (
Figure 2A and
Figure 3A). The eyes are small (5.7%
SL on average); the interorbital space is wide, nearly equal to eye diameter. The mouth is large; the maxillary bone is 52.1% of the head length, almost reaching the vertical of the posterior edge of eye. The armament on the head is well developed; there are four large sharp spines on the preoperculum. There are three front spines of similar length. The first spine is right forward, the other two are directed to the side, perpendicular to the bone. The posterior spine is the longest, averaging 5.6%
SL, which is comparable to the eye diameter.
There is a wide gap between the dorsal fins (from 5.1 to 10.1% SL, average 7.4%). Among the fins, the longest is the anal fin. The base slightly exceeds that of the second dorsal fin. The pelvic fins are short at 18.5% SL (range 16.6–22.5%).
The White Sea sculpin differs from the Baltic Sea sculpin in a set of morphological characteristics. The body has one long and two short rows of large but thin scale-like flaps (
Figure 2B and
Figure 3B). The flaps do not cover the body; they occupy only a small part of the surface. The head is high, not flattened in the dorso-ventral direction, and is well sculpted. The outer edges of the frontal bones are raised and form ridges above the eyes. There are postorbital and occipital bony outgrowths on the head. This fish is fundamentally different from those of the Baltic form (
Figure 2B and
Figure 3B). The outgrowths are small in size and have a triangular form. This type of bony outgrowth was never found in
T. quadricornis from the Baltic Sea. The eye is larger in
Triglopsis sp. from the White Sea (16–20.1% vs. 14.5–17.7% of head length). The interorbital space is concave and narrow, smaller than the eye diameter. There is a narrow gap between the first and second dorsal fins (3.6–6.0%
SL, average 4.9%). It is much less than the gap in the Baltic Sea sculpin (5.1–10.1%
SL, average 7.4%). The length of the base of the second dorsal fin is similar to the base length of the anal fin: 23.3–28.3%
SL (average 25.7%) and 22.7–27.9%
SL (average 25.3%), respectively.
3.2. Morphometric Characteristics
To find morphological fluctuation of body form, the ranges of variation and average values of 25 relative measurements were analyzed (
Table 1). The most noticeable differences were found in five characteristics: length of the caudal peduncle, base length of the second dorsal fin, distance between dorsal fins, head width, and interorbital space. The median values of three features (head width, interorbital space, and distance between dorsal fins) were higher in White Sea individuals than in Baltic ones. The difference of medians for the first two features averaged 9.7%. The great discrepancy in median values (34%) was found for only one characteristic: the gap between the dorsal fins. In the Baltic Sea specimens, dorsal fins are located at noticeably greater distance from each other than that of the White Sea fourhorn sculpins. Differences between medians of two other characters (length of caudal peduncle and base length of the second dorsal fin) in the Baltic and White Sea specimens were 9 and 10%, respectively.
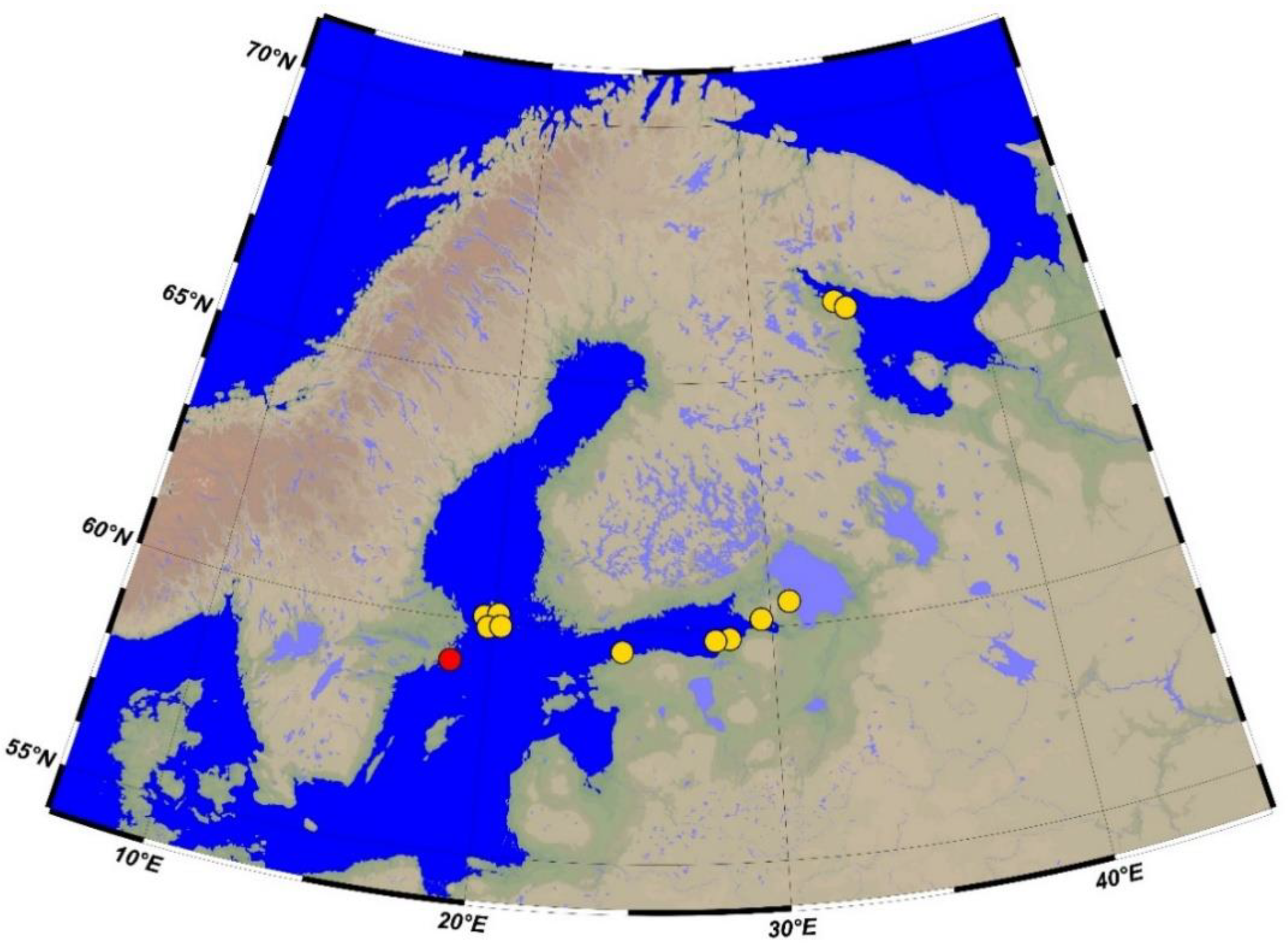
Data on the variation in the number of rays in the dorsal, anal, and pectoral fins in the White and Baltic Sea fourhorn sculpins are presented in
Table 2. A comparison of the obtained data showed that individuals from both localities have a similar level of variation in the number of fin rays. The number of rays in each fin varied by 2 to 4 values. The syntype from the Baltic Sea had nine rays in the first dorsal fin. The number of rays in non-type individuals from five localities in the Baltic Sea (
Figure 1) varied from six to nine (
Table 2). About 43% of specimens had eight rays in the first dorsal fin. The number of rays in the first dorsal fin of the White Sea fourhorn sculpin varied from 7 to 10 with a modal number of 9, which is more than in the Baltic Sea individuals. The number of rays in the second dorsal fin of the White Sea sculpin ranged from 14 to 16 with a modal value of 15. In the Baltic Sea fourhorn sculpin, this character ranged from 13 to 14 with a modal value of 13. Such differences in the modal numbers of rays are the greatest between two studied forms.
The unique type specimen from the Baltic Sea had 14 rays in the second dorsal fin. The number of rays in the anal fin varied from 13 to 15. Only one specimen from the White Sea contained 16 rays in the second dorsal fin (
Table 2). The modal number of anal-fin rays in both the White and Baltic Sea fourhorn sculpin (including the syntype) was 14. Such a number of rays in the anal fin was typical for most of the studied fish; 14 rays was found in 64% of the Baltic Sea specimens and in 43% of the White Sea fourhorn sculpins.
In the pectoral fin, 47% of all Baltic and White Sea specimens had 15 rays and 53% had 16 rays. The unique type specimen (Linnaeus, 1758) contained 17 rays in the pectoral fin. Therefore, the number of rays in the anal and pectoral fins of all the studied specimens from the Baltic and White Seas had similar levels of variation and the same modal number of rays. Differences in the modal number of rays were found in two dorsal fins; greater numbers of rays were more common in the White Sea fourhorn sculpin.
3.3. Axial Skeleton
The axial skeleton of
T. quadricornis syntype from the Baltic Sea had 40 vertebrae: 13 abdominal and 27 caudal, while the pleural ribs are absent. In non-type specimens from the Baltic Sea, the total number of vertebrae varied from 39 to 41, with a modal value of 40 (
Table 3). There are 13–14 ventral vertebrae and 27–28 caudal vertebrae. In the Baltic Sea fish, 57% of specimens were observed to have 27 caudal vertebrae.
The White Sea form had the same modal number of vertebrae as those of the Baltic Sea form. However, the variability in the number of vertebrae was completely different. The number of vertebrae in the White Sea specimens showed a downward trend (
Table 3). The greatest difference corresponding to interspecies dissimilarity was found in the number of pleural ribs. In 93% of the Baltic Sea specimens, pleural ribs on the last abdominal vertebrae were absent. In contrast, 81% of the White Sea specimens had one or two pairs of pleural ribs.
3.4. Lateral Line System and Fontanelles in Bones of Skull
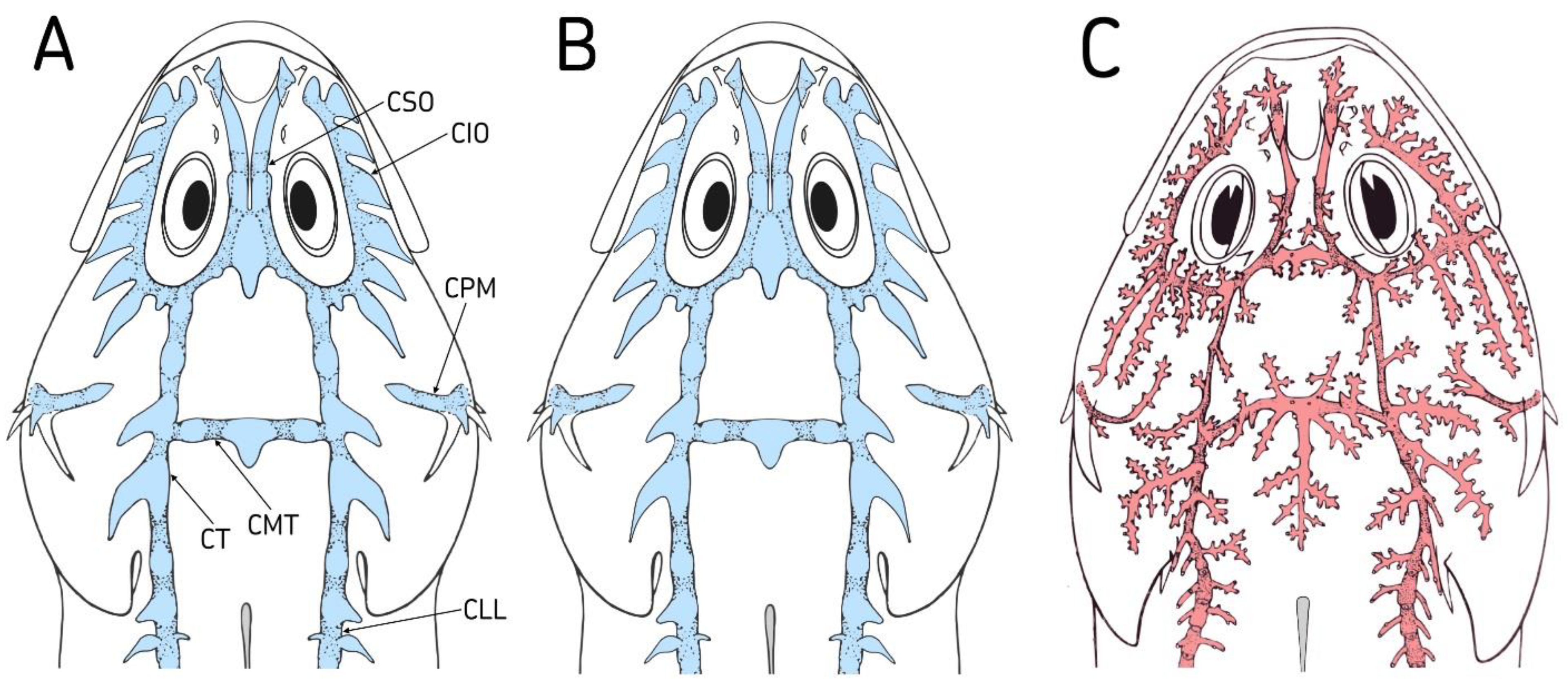
The lateral line system of
T. quadricornis from the Baltic Sea consisted of wide sensory canals, which are interconnected and form a unified system [
7]. The exception is the preopercular-mandibular canal, which is separated from other sensory canals of the head (
Figure 4A). Sensory canals have forms of cavities connected in series with each other and covered with a skin membrane from above. The distal parts of the cavities are elongated and form a skin canaliculus. The end of the canaliculus is usually open. However, sometimes the canaliculus ends blindly, in which case a pore is absent. The pore size is extremely small. The supraorbital canal has two cavities, an anterior one in the interorbital space and a posterior one in the position of coronal commissure connecting the left and right supraorbital canals.
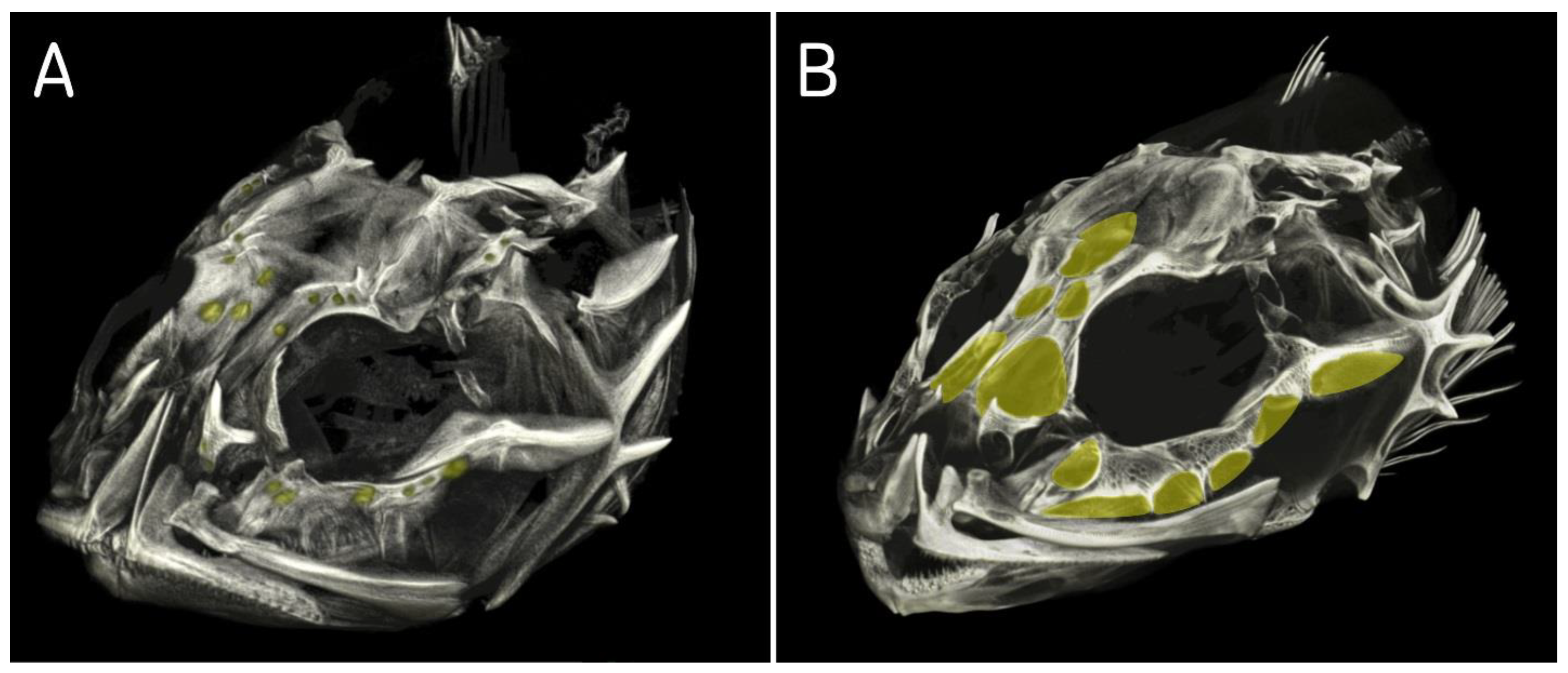
In the tomogram of
T. quadricornis skull, the frontal bones, through which the supraorbital canal passes, show large fontanelles (
Figure 5A). In the supraorbital canal, only one canaliculus opens with a pore. All other canaliculi end blindly and possess no pores. The infraorbital canal consists of eight interconnected extensive cavities. In infraorbital bones, they correspond to fontanelle, which are separated from each other in the upper part by very narrow bony septa (
Figure 5A). In the lower part, the fontanelles are interconnected. All cavities are elongated in the distal part and form skin canaliculi of different lengths. Only the first anterior canaliculus terminates blindly. All other seven canaliculi have pores at the ends.
The temporal canal contains four cavities. The anterior two do not have canaliculi, and therefore, they do not have pores. Wide skin canaliculi depart from the two posterior cavities. One canaliculus terminates blindly, and only the last one has a pore at the end (
Figure 4A). In the occipital commissure which connects temporal canals, there are three cavities. Of these, only the middle cavity has a skin canaliculus, but it does not have a pore. There are also cavities in the preopercular-mandibular canal. In the preoperculum, they are located between spines. The pores in this canal are situated directly on the skin at the roof of the cavity. The mandibular canals on the chin are interconnected by a triangle-shaped commissure. In the middle of the commissure, the canaliculus extends backwards [
7]. Only the lateral canaliculi are developed in the trunk canal. The distal part of each canaliculus in the anterior part of the trunk canal protrudes above the skin surface of the body. Each canaliculus has a pore at the end that sticks out above the surface of the body.
The pattern of lateral line system is similar between fourhorn sculpins from the White and Baltic Seas. The fundamental similarity lies in the presence of extensive cavities in all sensory canals, a small number of pores, and large frontanelles into the bones of the cranium (
Figure 4B and
Figure 5B). For comparison, the lateral line system of
M. stelleri as a type species of genus
Myoxocephalus was studied. The lateral line system of
M. stelleri has a different pattern compared to
Triglopsis from the Baltic and White Seas (
Figure 4C and
Figure 5C). The lateral line system consists of narrow sensory canals without cavities. All sensory canals have narrow openings in the cranium and small fontanelles (
Figure 5C). The skin canaliculi extending from the canals have a fundamentally different pattern. One canaliculus branches and has many additional canaliculi. Each additional canaliculus opens at the end. The species
T. quadricornis has 1 pore in the supraorbital canal, while
M. stelleri has 23 pores. In coronal commissure, these species have 0 and 15 pores, respectively. Seen from above, the scalp of
M. stellerii is dotted with a huge number of holes. The infraorbital canal has the largest number of pores. In this sensory canal, skin canaliculi extend from its outer and inner sides. In addition, the skin canaliculi are unusually long. Only one long canaliculus has 25 additional canaliculi and the same number of pores. The temporal canal and occipital commissure have a composition similar to the supra- and infraorbital canals. The unique composition is characteristic of the preopercular-mandibular canal. The preopercular part of this canal has branched canaliculi. The mandibular part of the canal, located on the lower surface of the head, has a simple canaliculus. One canaliculus corresponds to one pore.
3.5. DNA Analysis
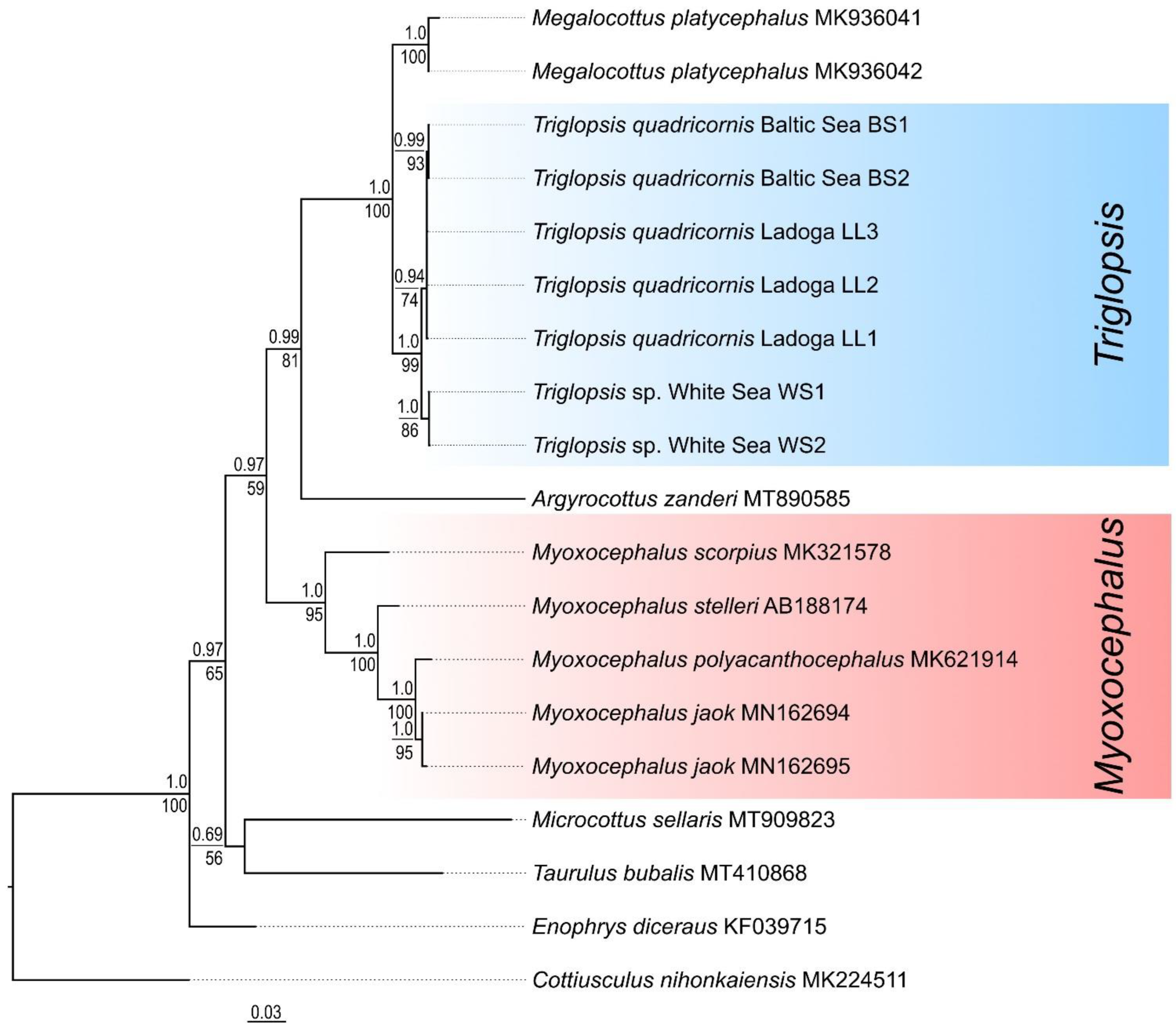
For the genetic characterization of two species of fourhorn sculpins from the Baltic and White Seas, the nucleotide sequences of the non-coding mtDNA CR (846–849 bp), as well as fragments of COI (620 bp) and RAG1 (1276 bp) genes were studied. The Baltic Sea T. quadricornis had one CR haplotype (BS1). A deletion (position 527) was found in one of the three CR sequences. Seven specimens of T. quadricornis from Lake Ladoga had three CR haplotypes (LL1, LL2, LL3) that differed from each other by one nucleotide substitution. The average p-distance between the Baltic Sea and Ladoga individuals was 0.2 ± 0.1%; no common haplotypes were found. This indicates that these are different populations of the same species.
In five specimens of Triglopsis sp. from the White Sea, two haplotypes were identified; one polymorphic site was found. The species T. quadricornis (including a form from Lake Ladoga) differed from the White Sea Triglopsis sp. in five nucleotide substitutions and two deletions in the mtDNA CR. The average p-distance between these species was 0.7 ± 0.3%, indicating their recent divergence. This level of genetic differentiation corresponds to differences between species of the same genus. The average p-distance (CR) between the genera Triglopsis and Myoxocephalus was 11.0 ± 0.8%. In contrast to the genus Triglopsis, genetic distances between marine species of the genus Myoxocephalus ranged from 1.8% to 8.9%.
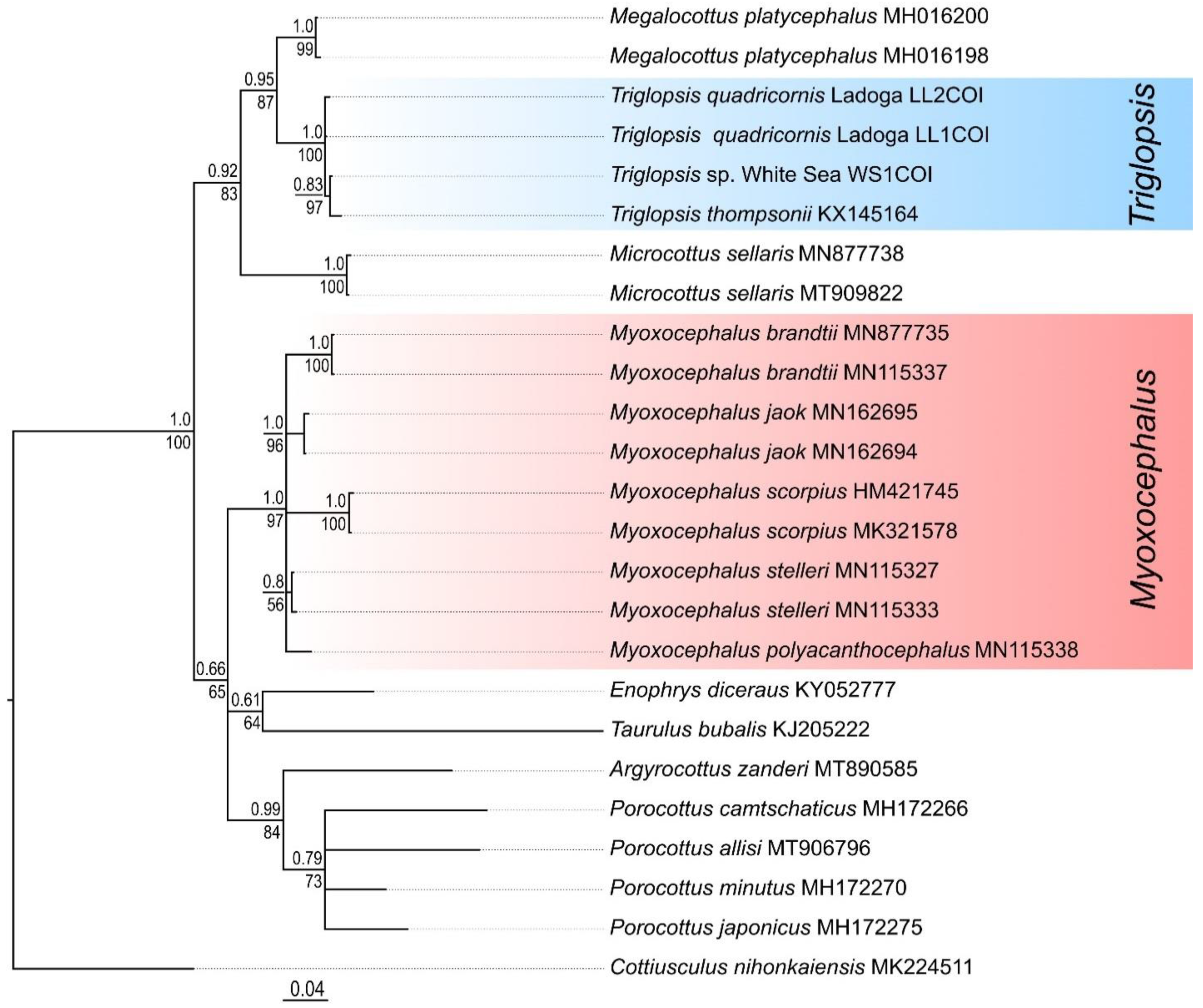
Analysis of COI sequences of the Baltic Sea and Ladoga forms of T. quadricornis revealed the presence of only two haplotypes (LL1COI and LL2COI). They were both found in specimens from the two localities studied. The Baltic Sea fourhorn sculpin differed from Triglopsis sp. from the White Sea by one nucleotide substitution in the fragment of COI gene (average p-distance 0.24 ± 0.18%). The low level of differences is due to the high conservatism of this marker in cottids. To establish the genetic similarity of the Baltic Sea T. quadricornis and the White Sea Triglopsis sp. with the type species of the genus Triglopsis (T. thompsonii), the COI gene sequence obtained from Genbank NCBI (Accession number HQ557195) was utilized. The average p-distances between T. thompsonii and the studied species of fourhorn sculpins were 0.5–0.6%. These results testify to the genetic similarity of the species of the genus Triglopsis. The genetic distances (COI) between the type species of the genus Myoxocephalus (M. stelleri, Accession number JF278626) and the fourhorn sculpins from the Baltic and White Seas were 7.6–7.8%. This high level of genetic divergence (12 times the interspecific distances within Triglopsis) is consistent with differences between independent genera. The average p-distance (COI) between the genera Triglopsis and Myoxocephalus was 8.5 ± 1.0%.
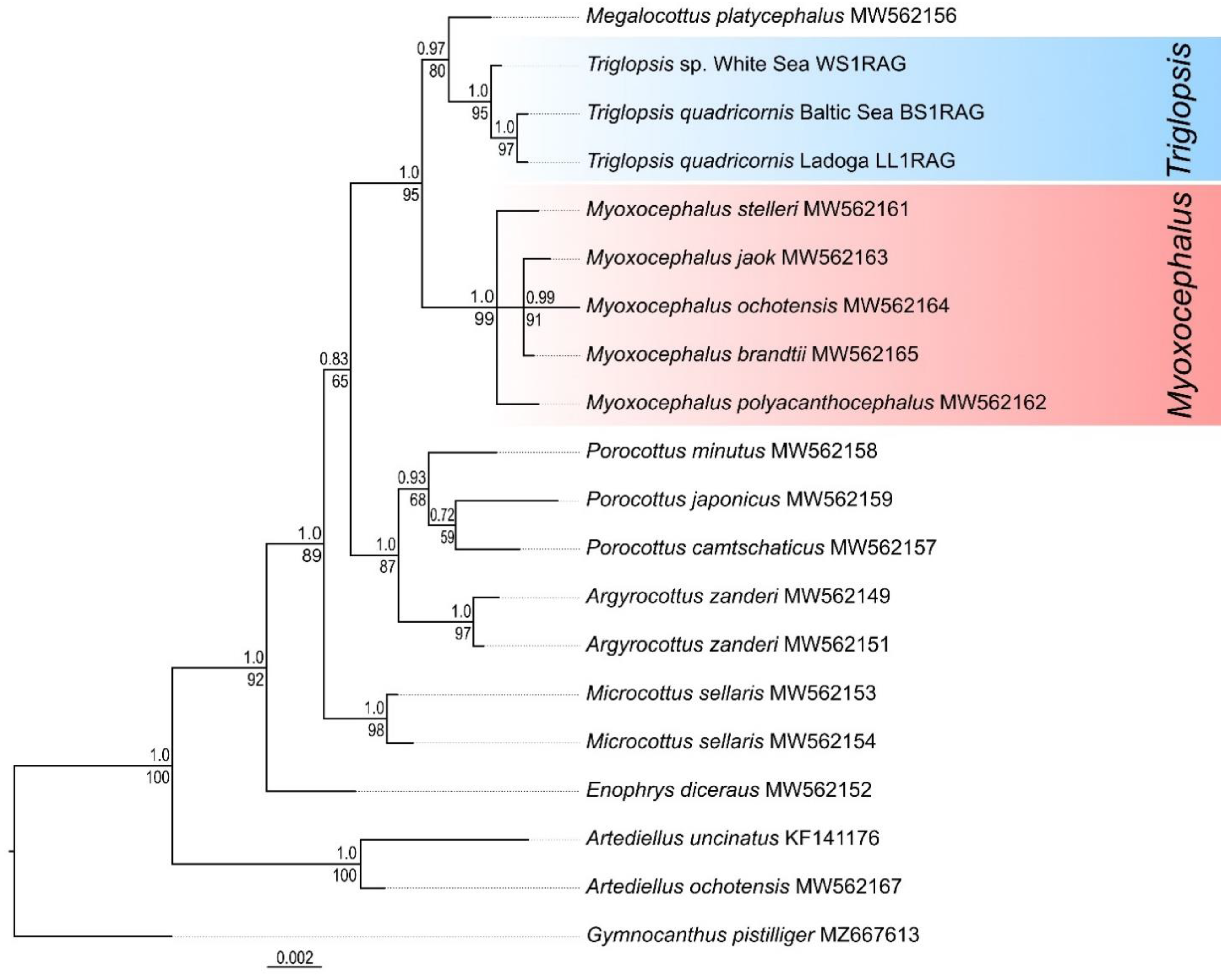
The validity of the independence of Triglopsis and Myoxocephalus was confirmed by nuclear DNA data (RAG1). The average p-distance between these genera was estimated as 0.75 ± 0.23%, which was significantly higher than the interspecies genetic differences. According to the sequences of the nuclear gene RAG1, the distances between the species of the genus Myoxocephalus were 0.08–0.47%. The RAG1 sequences of the Baltic four-horned sculpin and Triglopsis sp. from the White Sea differed by one nucleotide substitution (position 138); thus, the genetic distance between them was 0.08%. This level of interspecies divergence is typical for an ultraconservative gene. A similar distance (0.08%) was found between some marine species of the genus Myoxocephalus (M. brandtii and M. stelleri, M. brandtii and M. jaok).
3.6. Phylogenetic Analysis
An analysis of phylogenetic relationships of some representatives of Myoxocephalinae was performed based on the
CR,
COI, and
RAG1 sequences using Bayesian and ML methods. For each marker, both approaches for phylogenetic inference generated trees of similar topology. Bayesian trees reconstructed for each of the studied DNA fragments are shown in
Figure 6,
Figure 7 and
Figure 8.
The most basal position within the ingroup was occupied by
Enophrys diceraus, which lives in the northern part of the Pacific Ocean and the adjacent Arctic. On the
CR tree (
Figure 6), the clades corresponding to the two genera (
Triglopsis and
Myoxocephalus) were well isolated from each other and had very high supports. Clusters of
T. quadricornis (Baltic and Ladoga haplotypes) and White Sea
Triglopsis sp. were well supported on the
CR tree.
The clades corresponding to the two genera were also well isolated from each other and had very high supports on the
COI tree (
Figure 7). The subclades corresponding to
T. quadricornis and the White Sea
Triglopsis sp. were not separated within the “
Triglopsis” clade. The topology of the
COI tree differs from that of the
CR. The differences are related to the mutual arrangement of various genera, as well as the presence of many nodes with polytomy. This marker accumulates insufficient substitutions to study recent divergences.
On the phylogenetic tree based on the
RAG1 (ultraconservative genetic marker) (
Figure 8), the clades corresponding to
Triglopsis and
Myoxocephalus were well differentiated from each other and had very high support values. Despite the overall low variability of the
RAG1 gene, closely related
T. quadricornis from the Baltic Sea and
Triglopsis sp. from the White Sea differentiated on a phylogenetic tree with good support values. The
RAG1 tree was close in its topology to the tree reconstructed using the mtDNA
CR sequences.
4. Discussion
According to modern concepts, the fourhorn sculpin has a marine circumpolar distribution with freshwater forms in large lakes of glacial origin. The fourhorn sculpin belongs to a single species with the name
Myoxocephalus quadricornis [
22]. The species with the name
Cottus quadricornis existed from 1758 to 1907. In 1907, this species was transferred to the genus
Myoxocephalus with the type species
M. stelleri [
2]. The description of this species included the following features: smooth skin on the body, tuberous and porous skin on the head, abundance of spines on the top of head, and elongated upper jaw [
23]. The Baltic Sea
C. quadricornis was described with completely different morphological features than the type species
M. stelleri [
2,
23]. However, Gratzianow (1907) indicated a large size of gills fold on the isthmus as the main morphological feature of the genus
Myoxocephalus [
2]. Based upon this character Gratzianow transferred the Baltic Sea
C. quadricornis to the genus
Myoxocephalus. The following characters were specified as diagnostic: opening behind fourth gill arch and four bony outgrowths on the top of head [
2]. The study of the new morphological features of the lateral line system revealed large differences between
M. quadricornis and all other species of the genus
Myoxocephalus. Sensory canals in
M. quadricornis had large cavities that are absent in other species of the genus. Therefore, the species
M. quadricornis was moved to the genus
Triglopsis [
7].
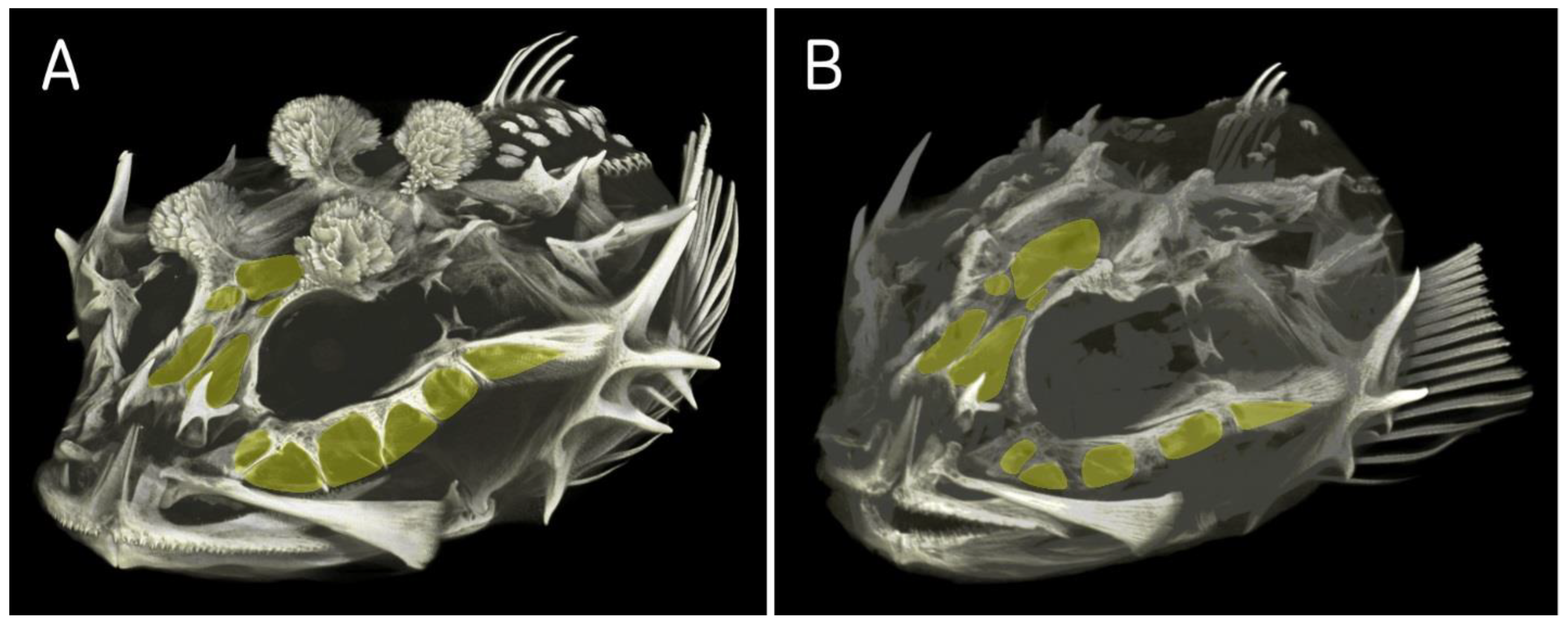
The study of the pattern of the cranium bones using computed tomography confirmed the classification of the Baltic Sea fourhorn sculpin within the genus
Triglopsis (
Figure 5 and
Figure 9). The type species of the genus (
T. thompsonii) has similar extensive cavities in the cranium bones as
T. quadricornis, whereas
M. stelleri lacks cavities and has only small fontanelles (
Figure 9A).
Other morphological features that confirm the differentiation of the Baltic Sea T. quadricornis include the following: the presence of large mushroom-shaped bony outgrowths on the cranium; abundance of dense bone plates on the body; a small number of pores on the head. Morphological differentiation is supported by genetic dissimilarity as well. This was revealed when studying the nucleotide sequences of the mtDNA CR. The genetic distances between T. quadricornis from the Baltic Sea and M. stelleri were 13.4%, while genetic differences between the species within the genus Myoxocephalus varied from 1.8 to 8.8%.
A comparative morphological analysis of the Baltic Sea and White Sea forms of fourhorn sculpin confirmed the differences between them. These differences were indicated earlier, and the rank of forms at the subspecies level was assessed [
4,
8,
9]. Morphological features characteristic of the White Sea form testify that it belongs to the genus
Triglopsis. These features include a pattern of lateral line systems that consists of sensory canals. The sensory canals are represented by interconnected cavities (
Figure 4B). Each sensory canal of the head has a small number of pores. The lateral line system is nearly analogous to that of
T. quadricornis. The differences of the White Sea fourhorn sculpin are as follows: the reduced number of cavities from eight to seven in the infraorbital canal, the presence of pore in the coronal commissure and first canaliculus of the infraorbital canal. The White Sea form also has extensive cavities in the frontal and infraorbital bones of the cranium (
Figure 5B). Unlike the Baltic Sea form, they are smaller and have wider bony septa. The main morphological features that distinguish this species from others are four triangular bony outgrowths on the head and a small number of bony plates on the body. Morphological features and patterns of the lateral line system reliably show the differences between the genera
Myoxocephalus and
Triglopsis. Classification of the Baltic and White Sea sculpins to the genus
Triglopsis, as well as the morphological isolation of each species
T. qiadricornis and
Triglopsis sp., were established.
The Baltic and White Sea species of the genus
Triglopsis are reproductively isolated. These two seas are not connected by any river systems, and the distance between them is more than 1300 km. So, the species live under different environmental conditions including the water salinity. The Baltic species inhabits desalinated areas of the northern and eastern parts of the sea, where the water salinity is between 2 and 6 ‰ [
24]. The White Sea species is distributed throughout the area with average water salinity 24–26 ‰ [
25].
The compared species differ in maximum age, growth rate, dominant feeding objects, and fecundity. The maximum age of the Baltic species was 11+ years, while for the White Sea species it was 8+ years [
25,
26]. Two species also differ in growth rates. The White Sea fourhorn sculpin grows faster than the Baltic one. Achieving maturity at 4+ years, the White Sea species had an average length of 184.6 mm compared to the 171 mm of the Baltic Sea species. At the maximum age, the length of the White Sea
Triglopsis sp. was 229.8 mm. The Baltic species maximum age length reached 275 mm [
25,
26]. Differences in growth rate are most likely determined by differences in food composition. The diet of White Sea species was dominated by polychaetes (more than 88% in the winter, and 77% in the summer) [
25]. The diet of
T. quadricornis from the Baltic Sea was dominated by isopod
Saduria entomon (86%). The role of fish in the diet of the Baltic fourhorn sculpin was five times less than that of the White Sea species [
27,
28].
The most significant differences between the two species of
Triglopsis were found in the fecundity values. So, at the age of 5+ to 6+ years spawning females of the White Sea fourhorn sculpin had an average number of 6100 eggs. The Baltic Sea sculpin at this age contained 9200 eggs. Upon reaching the maximum age of 11+ years, fecundity increased to 26,100 eggs [
24]. The revealed differences in distribution, habitat selection, food source, as well as fertility, indicate a different adaptive strategy of two species. This level of adaptation to the conditions of a particular waterbody is an additional argument in favor of separating the Baltic and White Sea forms and giving them the status of independent species.
The morphological differentiation of species and their assignment to the genus
Triglopsis was supported by genetic isolation. A comparative analysis of phylogenetic relationships within the genus
Myoxocephalus was performed in an article by Balakirev et al. [
12]. The main result of the study was the confirmation of the groundlessness of assigning the species
M. quadricornis to the genus
Myoxocephalus. The White Sea fourhorn sculpin was included in a cluster separate from all other studied species of
Myoxocephalus. This conclusion was based on analyzes of the ribosomal DNA fragment and the complete mitochondrial genome.
The genetic differences of the Baltic and White Sea species were identified based on our data on the sequences of the mtDNA
CR. The choice of
CR as a marker was due to the fact that the non-coding fragment accumulates a sufficient number of nucleotide substitutions to study recent divergences. This marker was successfully utilized for phylogenetic inferences of various groups of freshwater Cottoidei: Baikal sculpins, among which interspecific genetic distances can be equal to 0.2–0.3% [
11]; species group “
Cottus poecilopus” [
29]; Russian Far East and Japanese sculpins [
30,
31,
32]; European cottoid fishes [
33]. The mtDNA
CR was also used to determine the phylogenetic position of the new species
Cottus cyclophtalmus [
34].
The CR data for the Baltic fourhorn sculpin was expanded with sequences for specimens from Lake Ladoga. This lake is connected with the Gulf of Finland of the Baltic Sea by the Neva River. Specimens from these waterbodies are not geographically or genetically isolated. Therefore, the fourhorn sculpin from Lake Ladoga belongs to the same species T. quadricornis. The average genetic p-distance between individuals from Lake Ladoga and the Baltic Sea was 0.2 ± 0.1%. This corresponds to the level of interpopulation differences. The sequences of T. quadricornis and the White Sea Triglopsis sp. together formed a well-supported clade on the phylogenetic tree. The average genetic distance between T. quadricornis (haplotypes from the Baltic Sea and Lake Ladoga) and the White Sea form was 0.7 ± 0.3%. The level of differences is three times higher than between two Baltic populations and corresponds to interspecies divergence. Genetic difference between the genera Triglopsis and Myoxocephalus turned out to be quite high since the average p-distance was 11.0 ± 0.8%.
To improve result reliability, the analysis was extended by the study of the sequences of two conservative markers: mitochondrial COI and nuclear RAG1. The topology of the tree obtained using the COI marker had some differences from that of the CR. Discrepancies concern the relative position of some genera. The clades corresponding to the genera Triglopsis and Myoxocephalus were still well separated and had high support. The average p-distance (COI) between these genera was 8.5 ± 1.0%. The average genetic distance between the lineages of closely related species T. quadricornis and Triglopsis sp. was 0.24 ± 0.18%. This value is consistent with their relatively recent evolutionary divergence.
The results obtained using the ultraconservative RAG1 gene confirmed the differences between genera and species. According to this marker, the average genetic distance between Triglopsis and Myoxocephalus was 0.75 ± 0.23%. The p-distance between the Baltic Sea and White Sea species of the genus Triglopsis was only 0.08%. This interspecific distance was very small but confirmed the differences between two species. The distances between the species of the genus Myoxocephalus had close values (0.08–0.47%). It is clear that the RAG1 is not very suitable for the genetic identification of species, but it is good at revealing differences between genera. Thus, morphological and ecological differentiation was reliably supported by genetic isolation revealed during the study of the nucleotide sequences of two mtDNA and one nuclear DNA markers.













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}