
Power Lines and Birds: Drivers of Conflict-Prone Use of Pylons by Nesting White Storks (Ciconia ciconia)


Abstract
:1. Introduction
2. Materials and Methods
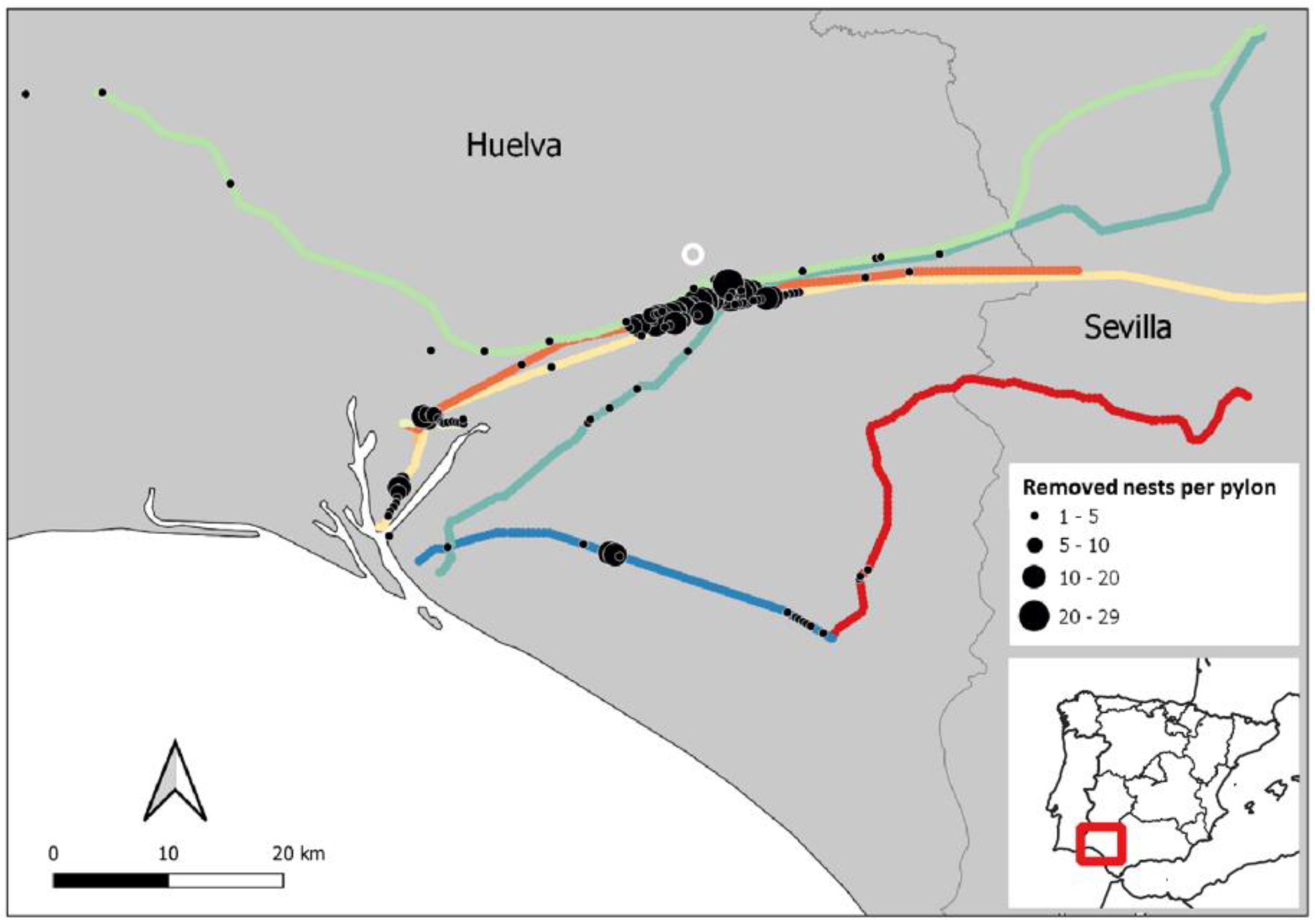
2.1. Study Area and Data Source
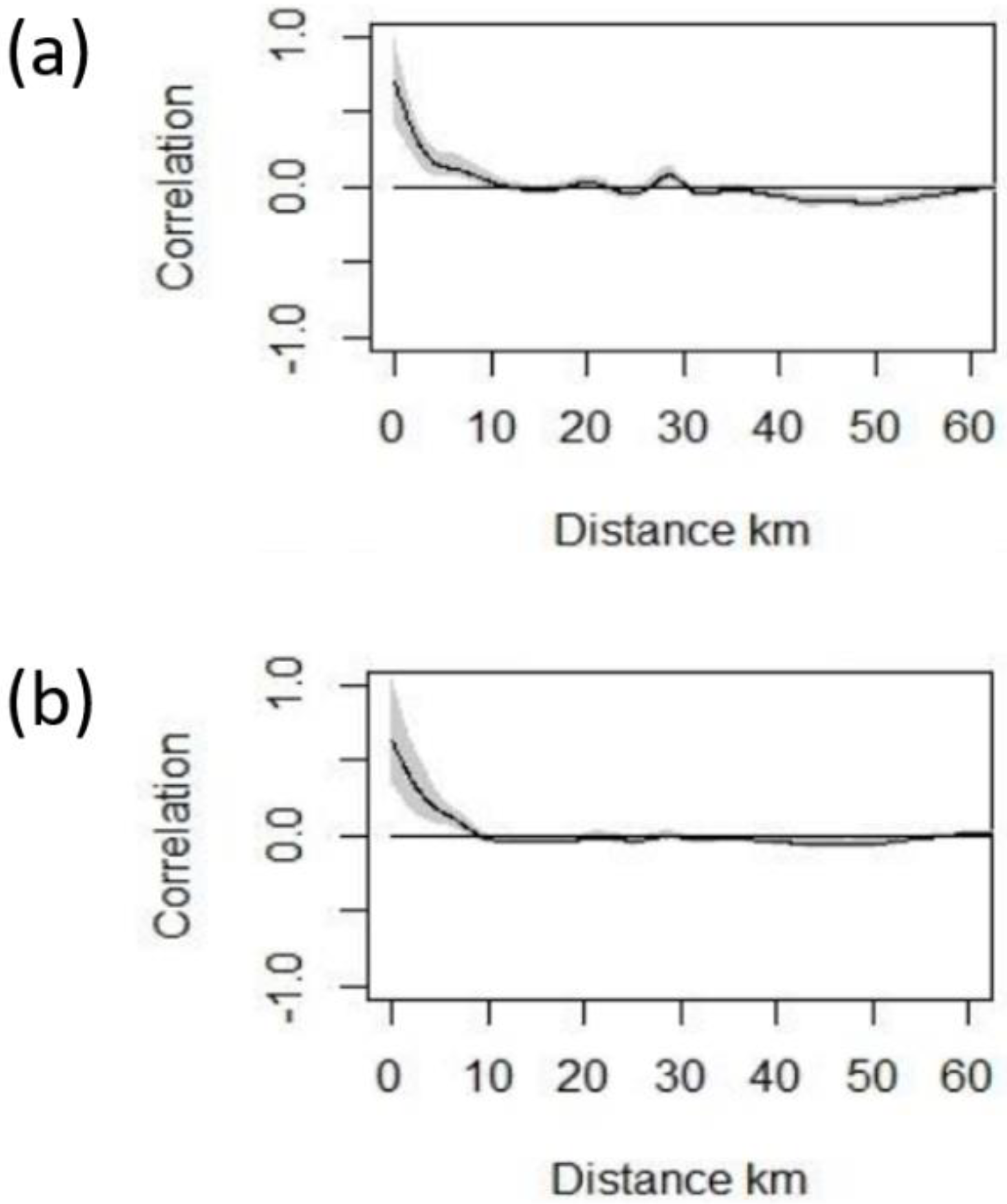
2.2. Data Analysis
2.3. Model Selection
3. Results
3.1. Predictors of Occurrence of Problematic Pylons
3.2. Drivers of Abundance of Problematic Nests
4. Discussion
4.1. Spatial Distribution of Pylon Use
4.2. Environmental/Ecological Drivers of Pylon Nesting
4.3. Potential Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanderson, E.W.; Jaiteh, M.; Levy, M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The Human Footprint and the Last of the Wild. BioScience 2002, 52, 891. [Google Scholar] [CrossRef] [Green Version]
- Butchart, S.H.M. Global Biodiversity: Indicators of recent decline. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Hovick, T.J.; Elmore, R.D.; Dahlgren, D.K.; Fuhlendorf, S.D.; Engle, D.M.; Hill, M. Evidence of negative effects of anthropogenic structures on wildlife: A review of grouse survival and behavior. J. Appl. Ecol. 2014, 51, 1680–1689. [Google Scholar] [CrossRef]
- Ferrer, M.; De Lucas, M.; Hinojosa, E.; Morandini, V. Transporting Biodiversity Using Transmission Power Lines as Stepping-Stones? Diversity 2020, 12, 439. [Google Scholar] [CrossRef]
- De Lucas, M.; Janss, G.; Whitfield, P.; Ferrer, M. Collision fatality of raptors in wind farms does not depend on raptor abundance. J. Appl. Ecol. 2008, 45, 1695–1703. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.C.; Ascensa, F.; Amico, M.D. Bird.On the wire: Landscape planning considering costs and benefits for bird populations coexisting with power lines. Ambio 2018, 47, 650–656. [Google Scholar] [CrossRef]
- Ferrer, M.; Morandini, V.; Baumbush, R.; Muriel, R.; De Lucas, M.; Calabuig, C. Efficacy of different types of “bird flight diverter” in reducing bird mortality due to collision with transmission power lines. Glob. Ecol. Conserv. 2020, 23, e01130. [Google Scholar] [CrossRef]
- Lior, N. Energy resources and use: The present situation and possible paths to the future. Energy 2008, 33, 842–857. [Google Scholar] [CrossRef]
- Kalt, G.; Thunshirn, P.; Haberl, H. A global inventory of electricity infrastructures from 1980 to 2017: Country-level data on power plants, grids and transformers. Data Brief 2021, 38, e107351. [Google Scholar] [CrossRef]
- Jenkins, A.R.; Smallie, J.J.; Diamond, M. Avian collisions with power lines: A global review of causes and mitigation with a South African perspective. Bird Conserv. Int. 2010, 20, 263–278. [Google Scholar] [CrossRef]
- Ferrer, M.; Hiraldo, F. Man-induced sex-biased mortality in the Spanish imperial eagle. Biol. Conserv. 1992, 60, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Ferrer, M.; Janss, G. Birds and Power Lines; Editorial Quercus: Madrid, Spain, 1999. [Google Scholar]
- Ferrer, M. Birds and Power Lines: From Conflict to the Solution; Migres Foundation: Seville, Spain, 2012. [Google Scholar]
- Ferrer, M.; De La Riva, M.; Castroviejo, J. Electrocution of raptors on power lines in Southern Spain. J. Field Ornithol. 1991, 62, 181–190. [Google Scholar]
- Bevanger, K. Biological and conservation aspects of bird mortality caused by electricity power lines: A review. Biol. Conserv. 1989, 86, 67–76. [Google Scholar] [CrossRef]
- Silva, J.P.; Manuel, J.; Alcazar, R.; Correia, R.; Delgado, A.; Moreira, F. A spatially explicit approach to assess the collision risk between birds and overhead power lines: A case study with the little bustard. Biol. Conserv. 2014, 170, 256–263. [Google Scholar] [CrossRef]
- Morelli, F.; Beim, M.; Jerzak, L.; Jones, D.; Tryjanowski, P. Can roads, railways and related structures have positive effects on birds?—A review. Transp. Res. Part D Transp. Environ. 2014, 30, 21–31. [Google Scholar] [CrossRef]
- Moreira, F.; Martins, R.C.; Catry, I.; D’Amico, M. Drivers of power line use by white storks: A case study of birds nesting on anthropogenic structures. J. Appl. Ecol. 2018, 55, 2263–2273. [Google Scholar] [CrossRef]
- Mainwaring, M.C. The use of man-made structures as nesting sites by birds: A review of the costs and benefits. J. Nat. Conserv. 2015, 25, 17–22. [Google Scholar] [CrossRef]
- Janiszewski, T.; Minias, P.; Wojciechowski, Z. Selective forces responsible for transition to nesting on electricity poles in the white stork Ciconia ciconia. Ardea 2015, 103, 39–50. [Google Scholar] [CrossRef]
- Steenhof, K.; Kochert, M.N.; Roppe, J.A. Nesting by raptors and common ravens on electricity transmission line towers. J. Wildl. Manag. 1993, 57, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Jakubiec, Z. Causes of breeding losses and adult mortality in white stork Ciconia ciconia in Poland. Stud. Nat. 1991, 37, 107–124. [Google Scholar]
- Garrido, J.R.; Fernández-Cruz, M. Effects of power lines on a white stork Ciconia ciconia population in Central Spain. Ardeola 2003, 50, 191–200. [Google Scholar]
- Balmori, A. Possible Effects of Electromagnetic Fields from Phone Masts on a Population of White Stork (Ciconia ciconia). Electromagn. Biol. Med. 2005, 24, 109–119. [Google Scholar] [CrossRef]
- Vaitkuvienė, D.; Dagys, M. Possible effects of electromagnetic field on White Storks Ciconia ciconia breeding on low-voltage electricity line poles. Zool. Ecol. 2014, 24, 289–296. [Google Scholar] [CrossRef]
- Maricato, L.; Faria, R.; Madeira, V.; Carreira, P.; De Almeida, A.T. White stork risk mitigation in high voltage electric distribution networks. Ecol. Eng. 2016, 91, 212–220. [Google Scholar] [CrossRef]
- Newman, J.; Newman, C.; Lindsay, J.; Merchant, B.; Avery, M.; Pruett-Jones, S. Monk parakeets: An expanding problem on power lines and other electrical utility structures. In Environment Concerns in Rights-of-Way Management 8th International Symposium; Goodrich-Mahoney, J.W., Abrahamson, L., Ballard, J., Tikalsky, S., Eds.; Elsevier: New York, NY, USA, 2004; pp. 355–364. [Google Scholar]
- McIvor, G.E.; Rowe, C.; Healy, S.D. Deterring hooded crows from re-nesting on power poles. Wildl. Soc. Bull. 2012, 36, 729–734. [Google Scholar] [CrossRef]
- Red Eléctrica de España. Red Eléctrica and Birdlife: 15 Years of Applied Research; REE, S.A.: Madrid, Spain, 2005. [Google Scholar]
- Harness, R.E. Raptor Nest Management on Power Lines. In Proceedings of the Fifth IASTED International Conference on Power and Energy Systems, Benalmádena, Spain, 15–17 June 2005; pp. 534–538. [Google Scholar]
- Polat, Ö. An overview of bird related issues in electrical power systems. IOP Conf. Ser. Mater. Sci. Eng. 2016, 161, 012091. [Google Scholar] [CrossRef] [Green Version]
- Burnham, J.; Carlton, R.; Cherney, E.; Couret, G.; Eldridge, K.; Farzaneh, M.; Frazier, S.; Gorur, R.; Harness, R.; Shaffner, D.; et al. Preventive Measures to Reduce Bird-Related Power Outages—Part I: Electrocution and Collision. IEEE Trans. Power Deliv. 2004, 19, 1843–1847. [Google Scholar] [CrossRef]
- Kaluga, I.; Sparks, T.H.; Tryjanowski, P. Reducing death by electrocution of the white stork Ciconia ciconia. Conserv. Lett. 2011, 4, 483–487. [Google Scholar] [CrossRef]
- Tryjanowski, P.; Kosicki, J.Z.; Kuźniak, S.; Sparks, T.H. Long-term changes and breeding success in relation to nesting structures used by the White Stork, Ciconia ciconia. Ann. Zool. Fenn. 2009, 46, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Muriel, R.; Ferrer, M.; Casado, E.; Calabuig, C. First successful breeding of reintroduced ospreys Pandion haliaetus in mainland Spain. Ardeola 2010, 57, 175–180. [Google Scholar]
- Gyalus, A.; Végvári, Z.; Csörgő, T. Changes in the nest sites of white stork (Ciconia ciconia) in Hungary. Ornis Hung. 2018, 26, 65–88. [Google Scholar] [CrossRef] [Green Version]
- Molina, B.; Del Moral, J.C. La Cigüeña Blanca en España. VI Censo Internacional; SEO/BirdLife: Madrid, Spain, 2005. [Google Scholar]
- Gilbert, N.I.; Correia, R.A.; Silva, J.P.; Pacheco, C.; Catry, I.; Atkinson, P.W.; Gill, J.A.; Franco, A.M.A. Are white storks addicted to junk food? Impacts of landfill use on the movement and behaviour of resident white storks (Ciconia ciconia) from a partially migratory population. Mov. Ecol. 2015, 4, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zbyryt, A.; Dylewski, L.; Neubauer, G. Mass of white stork nests predicted from their size: Online calculator and implications for conservation. J. Nat. Conserv. 2021, 60, 125967. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS 3.14.0 Geographic Information System; Open Source Geospatial Foundation: Beaverton, OR, USA, 2020. [Google Scholar]
- Cramp, S.; Simmons, K.E. Birds of the Western Palearctic; Oxford University Press: Oxford, UK, 1977; Volume I. [Google Scholar]
- Alonso, J.C.; Alonso, J.A.; Carrascal, L.M. Habitat selection by foraging white storks, Ciconia ciconia, during the breeding season. Can. J. Zool. 1991, 69, 1957–1962. [Google Scholar] [CrossRef]
- Johst, K.; Brandl, R.; Pfeifer, R. Foraging in a patchy and dynamic landscape: Human land use and the White Stork. Ecol. Appl. 2001, 11, 60–69. [Google Scholar] [CrossRef]
- Lázaro, E.; Chozas, P.; Fernández-Cruz, M. Demografia de la Cigüeña Blanca (Ciconia ciconia) en España. Censo Nacional de 1984. Ardeola 1986, 33, 131–169. [Google Scholar]
- Carrascal, L.M.; Bautista, L.M.; Lázaro, E. Geographical variation in the density of the white stork Ciconia ciconia in Spain: Influence of habitat structure and climate. Biol. Conserv. 1993, 65, 83–87. [Google Scholar] [CrossRef]
- Bruederle, A.; Hodler, R. Nighttime lights as a proxy for human development at the local level. PLoS ONE 2018, 13, e0202231. [Google Scholar] [CrossRef] [Green Version]
- Shi, K.; Huang, C.; Chen, Y.; Li, L. Remotely sensed nighttime lights reveal increasing human activities in protected areas of China mainland. Remote Sens. Lett. 2018, 9, 467–476. [Google Scholar] [CrossRef]
- National Centers for Environmental Information Version 1 VIIRS Day/Night Band Nighttime Lights. Available online: https://ngdc.noaa.gov/eog/viirs/download_dnb_composites.html (accessed on 22 April 2020).
- Bivand, R.S.; Wong, D. Comparing implementations of global and local indicators of spatial association. TEST 2018, 27, 716–748. [Google Scholar] [CrossRef]
- Crase, B.; Liedloff, A.C.; Wintle, B.A. A new method for dealing with residual spatial autocorrelation in species distribution models. Ecography 2012, 35, 879–888. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Real, R.; Muñoz, A.; Brown, J. New measures for assessing model equilibrium and prediction mismatch in species distribution models. Divers. Distrib. 2013, 19, 1333–1338. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Infante, O.; Peris, S. Bird nesting on electric power supports in northwestern Spain. Ecol. Eng. 2003, 20, 321–326. [Google Scholar] [CrossRef]
- Lazo, A.; Castelló, A.; Braña, S.; Conde, J.L. Análisis de los Resultados del Seguimiento de la Nidificación de Cigüeñas Blancas y Eficacia de los Dispositivos Disuasores Instalados en Torres de Transporte de Electricidad de Red Eléctrica de España Entre 1999 y 2014; Red Eléctrica de España: Madrid, Spain, 2016. [Google Scholar]
- Gadenne, H.; Cornulier, T.; Eraud, C.; Barbraud, J.C.; Barbraud, C. Evidence for density-dependent habitat occupancy at varying scales in an expanding bird population. Popul. Ecol. 2014, 56, 493–506. [Google Scholar] [CrossRef]
- Bachir, A.S.; Chenchouni, H.; Djeddou, N.; Barbraud, C.; Céréghino, R.; Santoul, F. Using self-organizing maps to investigate environmental factors regulating colony size and breeding success of the White Stork (Ciconia ciconia). J. Ornithol. 2013, 154, 481–489. [Google Scholar] [CrossRef] [Green Version]
- Vergara, P.; Aguirre, J.I. Age and breeding success related to nest position in a White stork Ciconia ciconia colony. Acta Oecol. 2006, 30, 414–418. [Google Scholar] [CrossRef]
- Djerdali, S.; Guerrero-Casado, J.; Tortosa, F.S. The effects of colony size interacting with extra food supply on the breeding success of the White Stork (Ciconia ciconia). J. Ornithol. 2016, 157, 941–947. [Google Scholar] [CrossRef]
- Zbyryt, A.; Sparks, T.H.; Tryjanowski, P. Whitewashing improves relocated nest occupancy in the white stork: An experimental test of public information. J. Nat. Conserv. 2021, 59, 125929. [Google Scholar] [CrossRef]
- Cuadrado, M.; Sánchez, I.; Barcell, M.; Armario, M. Reproductive data and analysis of recoveries in a population of white stork Ciconia ciconia in southern Spain: A 24-year study. Anim. Biodivers. Conserv. 2016, 39, 37–44. [Google Scholar] [CrossRef]
- Bialas, J.T.; Dylewski, Ł.; Tobolka, M. Determination of nest occupation and breeding effect of the white stork by human-mediated landscape in Western Poland. Environ. Sci. Pollut. Res. 2020, 27, 4148–4158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialas, J.T.; Dylewski, Ł.; Dylik, A.; Janiszewski, T.; Kaługa, I.; Królak, T.; Kruszyk, R.; Pawlukojć, K.; Pestka, Z.; Polakowski, M.; et al. Impact of land cover and landfills on the breeding effect and nest occupancy of the white stork in Poland. Sci. Rep. 2021, 11, 7279. [Google Scholar] [CrossRef] [PubMed]
- Benharzallah, N.; Bachir, A.S.; Barbraud, C. Nest characteristics and food supply affect reproductive output of white storks Ciconia ciconia in semi-arid areas. Biologia 2022, 77, 997–1006. [Google Scholar] [CrossRef]
- Tsachalidis, E.P.; Goutner, V. Diet of the White Stork in Greece in Relation to Habitat. Waterbirds 2002, 25, 417–423. [Google Scholar] [CrossRef]
- Latus, C.; Kujawa, K.; Glemnitz, M. The influence of landscape structure on White Stork’s Ciconia ciconia nest distribution. Acta Ornithol. 2000, 35, 97–102. [Google Scholar]
- Ferreira, E.; Grilo, F.; Mendes, R.; Lourenço, R.; Santos, S.; Petrucci-Fonseca, F. Diet of the White Stork (Ciconia ciconia) in a heterogeneous Mediterranean landscape: The importance of the invasive Red Swamp Crayfish (Procambarus clarkii). Airo 2019, 26, 33–47. [Google Scholar]
- Moritzi, M.; Maumary, L.; Schmid, D.; Steiner, I.; Vallotton, L.; Spaar, R.; Biber, O. Time budget, habitat use and breeding success of White Storks Ciconia ciconia under variable foraging conditions during the breeding season in Switzerland. Ardea 2001, 89, 457–470. [Google Scholar]
- Tryjanowski, P.; Kuzniak, S. Population size and productivity of the White Stork Ciconia ciconia in relation to common vole Microtus arvalis density. Ardea 2002, 90, 213–217. [Google Scholar]
- Kamiński, P.; Jerzak, L.; Kasprzak, M.; Kartanas, E.; Bocheński, M.; Hromada, M.; Baszyński, J.; Kozera, W.; Woźniak, A.; Ulrich, W. Do agricultural environments increase the reproductive success of White Stork Ciconia ciconia populations in South-Western Poland? Sci. Total Environ. 2020, 702, 134503. [Google Scholar] [CrossRef]
- Radović, A.; Tepić, N. Using Corine Land Cover habitat database for the analysis of breeding bird habitat: Case study of white storks (Ciconia ciconia) from northern Croatia. Biologia 2009, 64, 1212–1218. [Google Scholar] [CrossRef]
- Podlaszczuk, M.; Wojciechowski, Z.; Podlaszczuk, P.; Minias, P.; Janiszewski, T.; Wojciechowska, A. Shortening day length as a previously unrecognized selective pressure for early breeding in a bird with long parental care. J. Ornithol. 2014, 156, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Jerzak, L.; Bocheński, M.; Czechowski, P. Unusual feeding behaviour of the White Stork Ciconia ciconia in the Kłopot colony (W Poland). In The White Stork in Poland: Studies in Biology, Ecology and Conservation; Tryjanowski, P., Sparks, T.H., Jerzak, L., Eds.; Bogucki Wydawnictwo Naukowe: Poznań, Poland, 2006; pp. 203–207. [Google Scholar]
- Zbyryt, A.; Sparks, T.H.; Tryjanowski, P. Foraging efficiency of white stork Ciconia ciconia significantly increases in pastures containing cows. Acta Oecol. 2020, 104, 103544. [Google Scholar] [CrossRef]
- Janss, G.F.E.; Ferrer, M. Avian electrocution mortality in relation to pole design and adjacent habitat in Spain. Bird Conserv. Int. 2001, 11, 3–12. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable Name | Description | Mean (Range) |
|---|---|---|
| Dist_to_landfill_km | Distance to the nearest landfill site (km) | 22 (3.5–54) |
| Dist_wetland_km | Distance to nearest freshwater marsh (km) | 13 (0.2–51) |
| P_agroforestry | Proportion of agroforestry land within buffer | 0.06 (0–1) |
| P_other_heterogeneous_agr | Proportion of other heterogenous agricultural systems not agroforestry (i.e., parcels of land which combine permanent and annual crops, agriculture, and natural vegetation) within buffer | 0.01 (0–0.2) |
| P_non_irrigated_arable | Proportion of non-irrigated arable crops within buffer | 0.4 (0–1) |
| P_perm_irrigated_crops | Proportion of permanently irrigated agricultural land within buffer. | 0.1 (0–1) |
| P_grassland | Proportion of natural grassland and agricultural pasture within buffer | 0.03 (0–0.8) |
| Pres_water | A binary variable which denotes the presence of inland freshwater bodies and rivers within buffer | 0.087 (0–1) |
| Nighttime_light_intensity | The mean value of light intensity of pixels within buffer | 1.8 (0–54) |
| Predictor Variable | Problematic Pylons | Abundance of Problematic Nests | |
|---|---|---|---|
| β ± SE | OR | β ± SE | |
| Intercept | −1.702 ± 0.309 | - | 1.49 ± 0.363 *** |
| Dist_landfill | −0.065 ± 0.015 *** | 0.94 | −0.104 ± 0.016 *** |
| Nigttime_light_intensity | 0.039 ± 0.02 * | 1.04 | 0.082 ± 0.026 *** |
| P_grassland | 2.761 ± 0.64 *** | 15.8 | 1.807 ± 0.836 * |
| P_agroforestry | −2.189 ± 1.286 | 0.11 | −4.467 ± 1.205 *** |
| Pres_water | 0.98 ± 0.345 ** | 2.68 | - |
| P_perm_irrigated_crops | - | - | −1.809 ± 0.674 ** |
| D2 (adjusted) | 0.43 | 0.54 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burdett, E.M.; Muriel, R.; Morandini, V.; Kolnegari, M.; Ferrer, M. Power Lines and Birds: Drivers of Conflict-Prone Use of Pylons by Nesting White Storks (Ciconia ciconia). Diversity 2022, 14, 984. https://doi.org/10.3390/d14110984
Burdett EM, Muriel R, Morandini V, Kolnegari M, Ferrer M. Power Lines and Birds: Drivers of Conflict-Prone Use of Pylons by Nesting White Storks (Ciconia ciconia). Diversity. 2022; 14(11):984. https://doi.org/10.3390/d14110984
Chicago/Turabian StyleBurdett, Evan M., Roberto Muriel, Virginia Morandini, Mahmood Kolnegari, and Miguel Ferrer. 2022. "Power Lines and Birds: Drivers of Conflict-Prone Use of Pylons by Nesting White Storks (Ciconia ciconia)" Diversity 14, no. 11: 984. https://doi.org/10.3390/d14110984