
Predicting the Potential Suitable Area of the Invasive Ant Linepithema humile in China under Future Climatic Scenarios Based on Optimized MaxEnt
Abstract
:1. Introduction
2. Materials and Methods
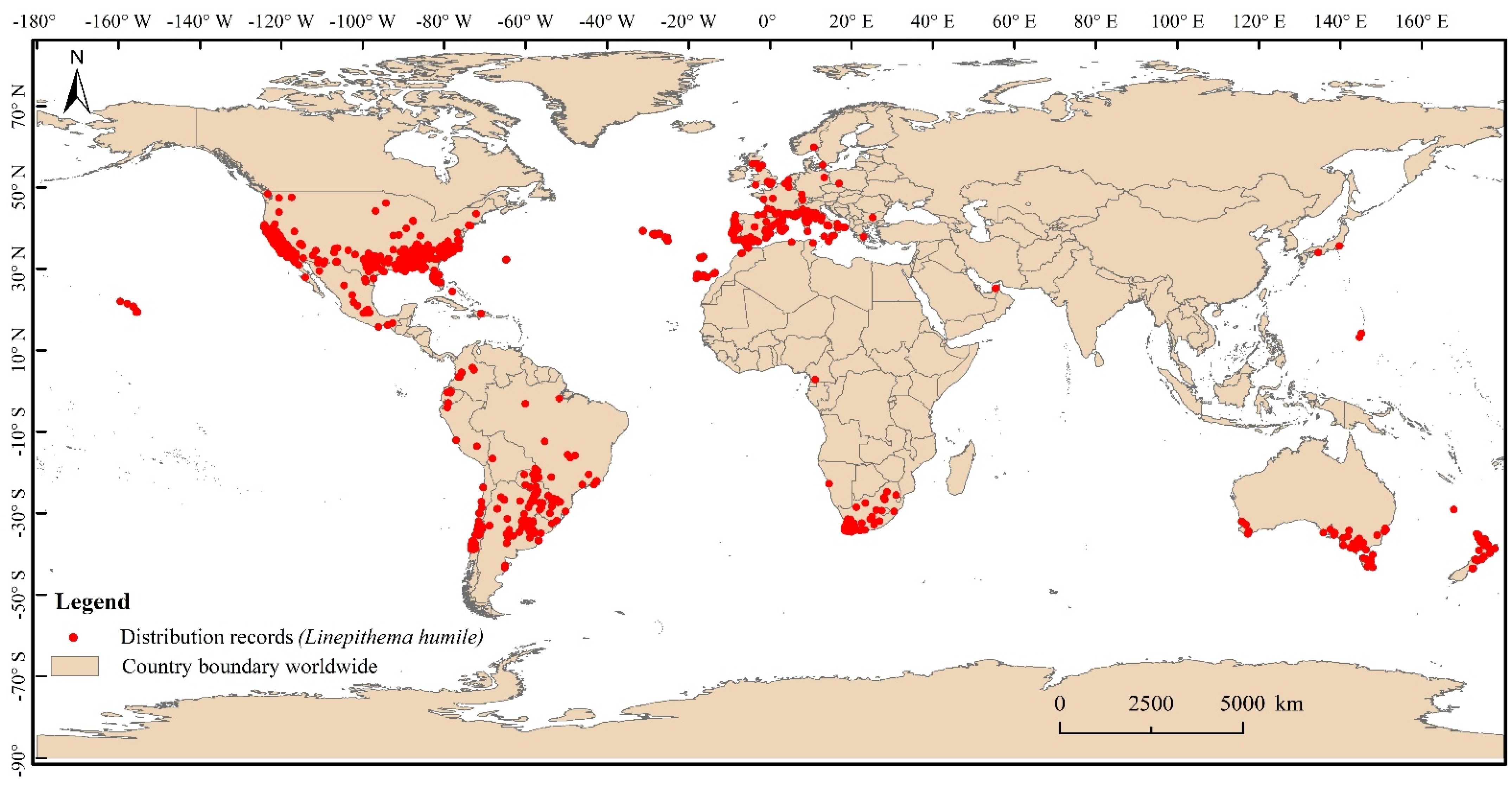
2.1. Global Occurrence Records of Linepithema humile
2.2. Bioclimatic Variables
2.3. Optimized MaxEnt Modeling
2.4. Model Evaluation and Potential Suitable Area Delineation
3. Results
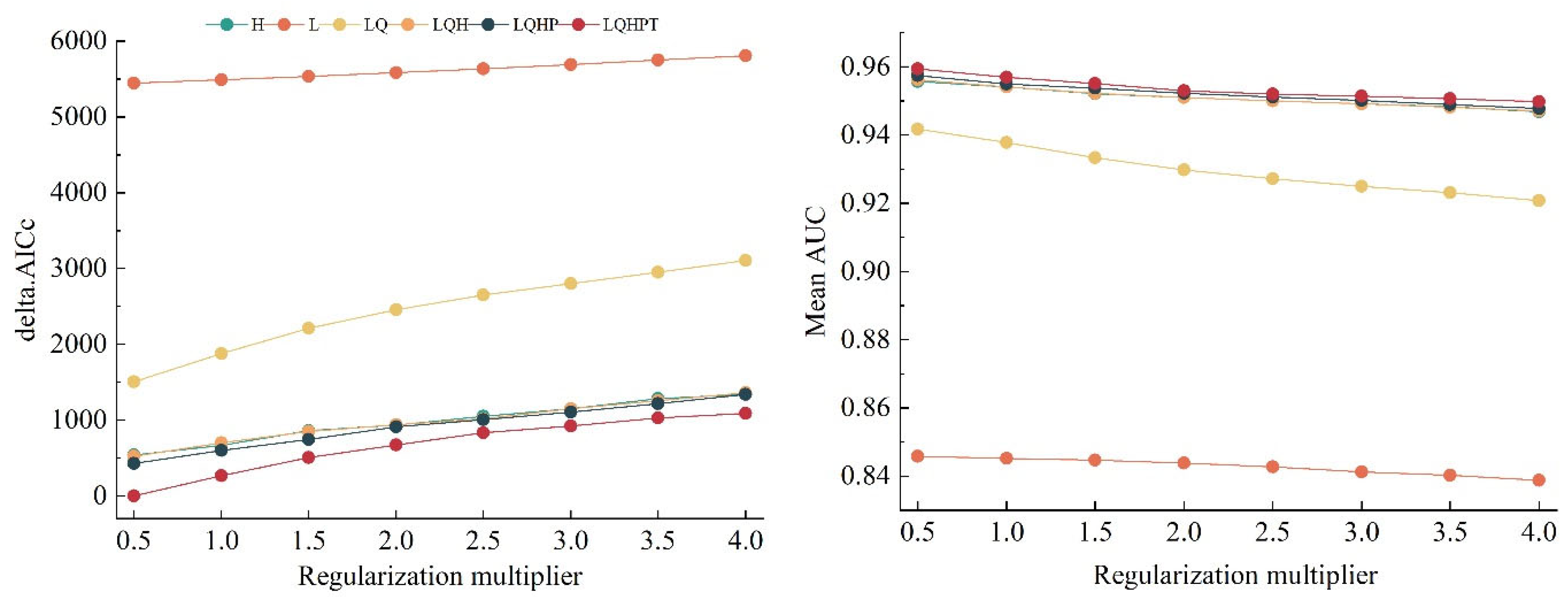
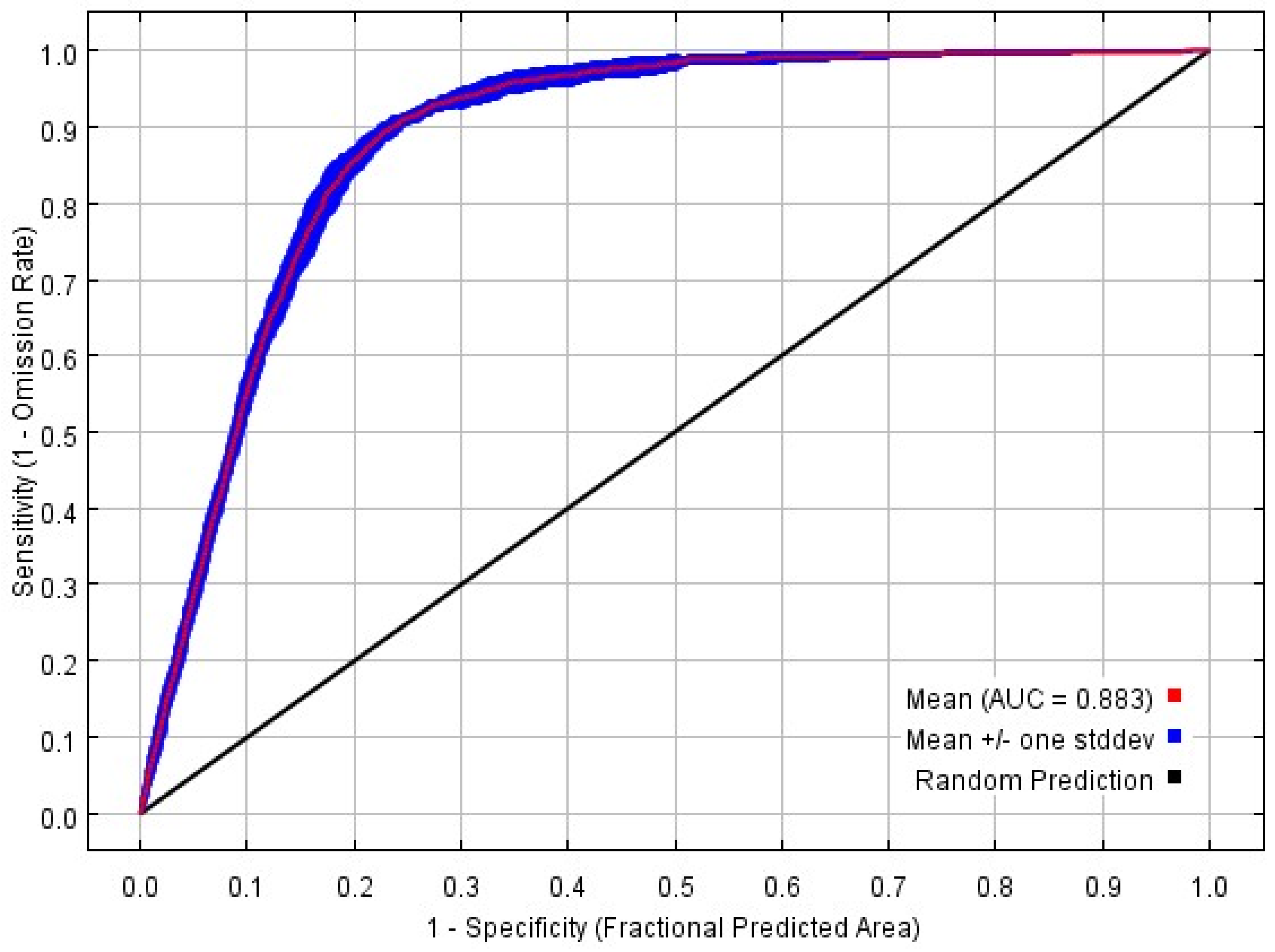
3.1. Model Optimization and Accuracy Evaluation
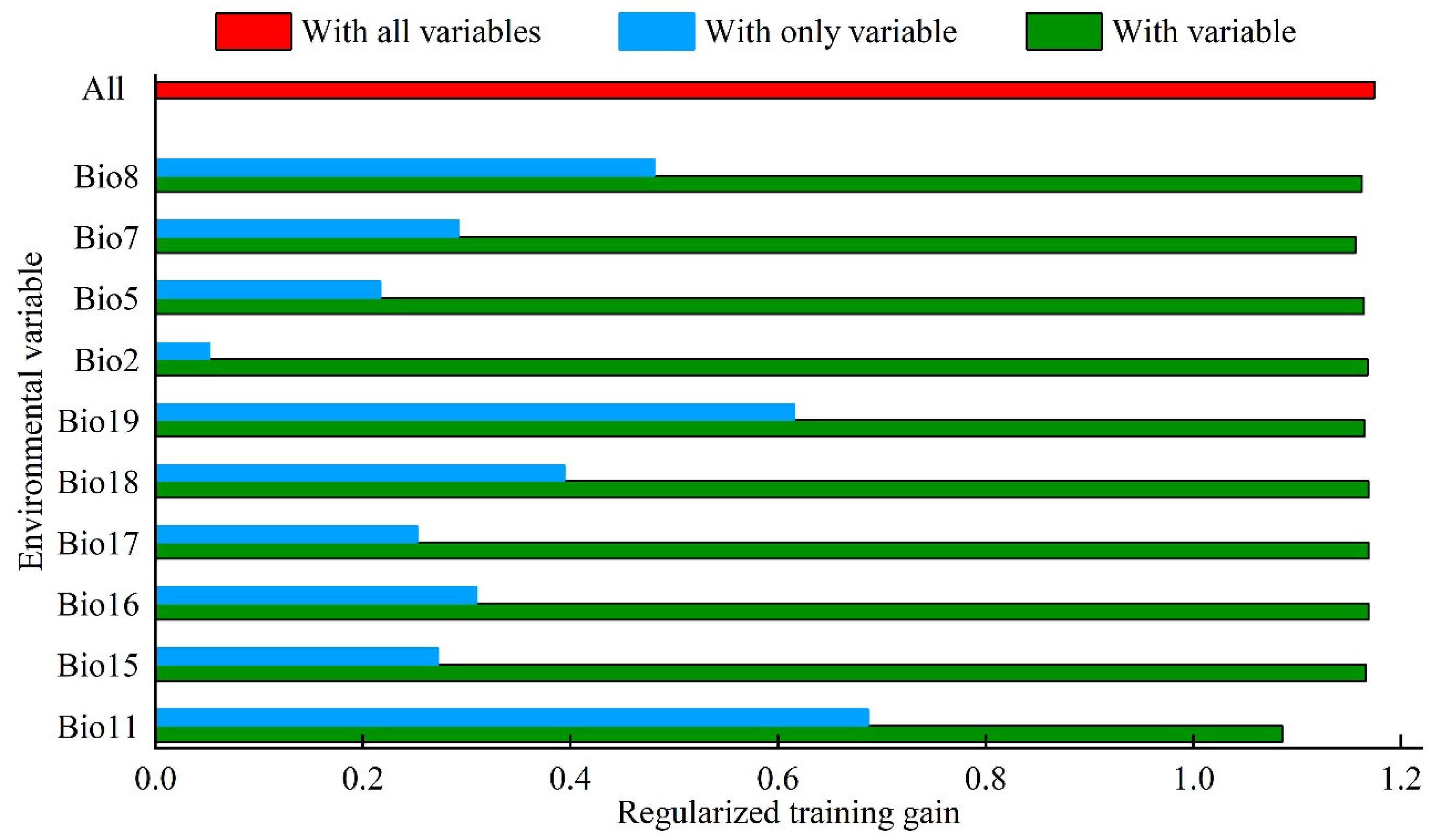
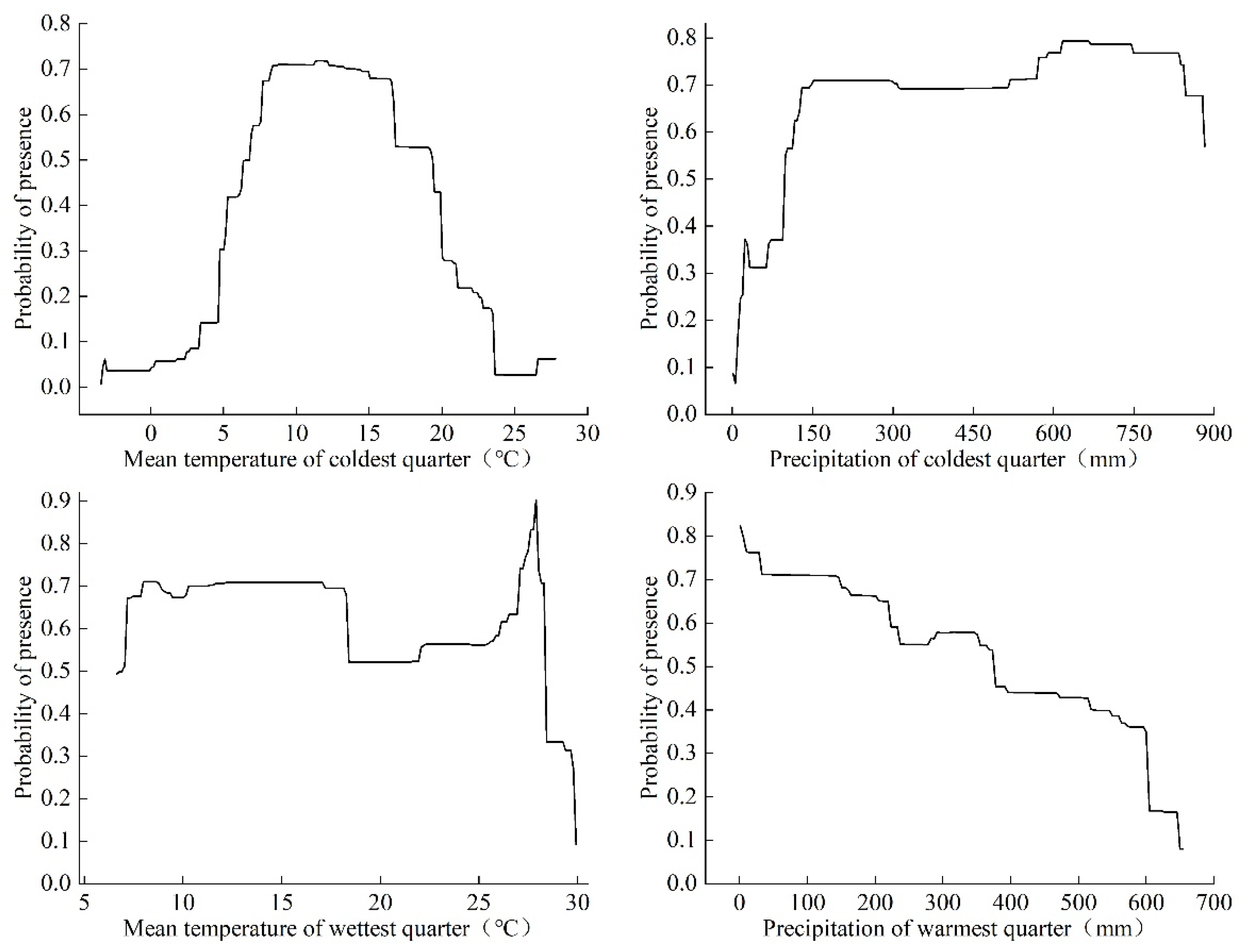
3.2. Bioclimatic Variables Affecting the Potential Suitable Area of Linepithema humile
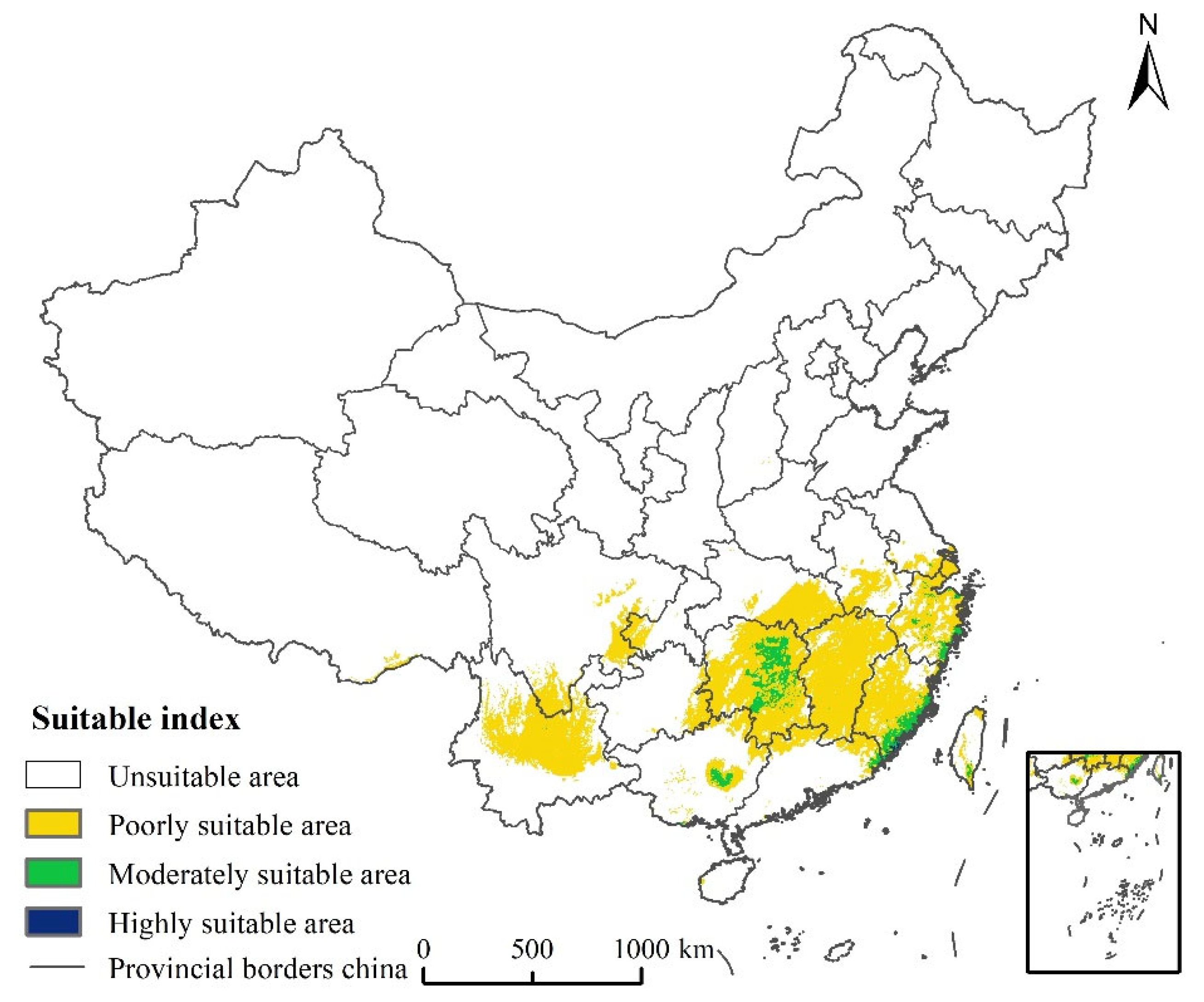
3.3. Current Potential Suitable Areas of Linepithema humile in China
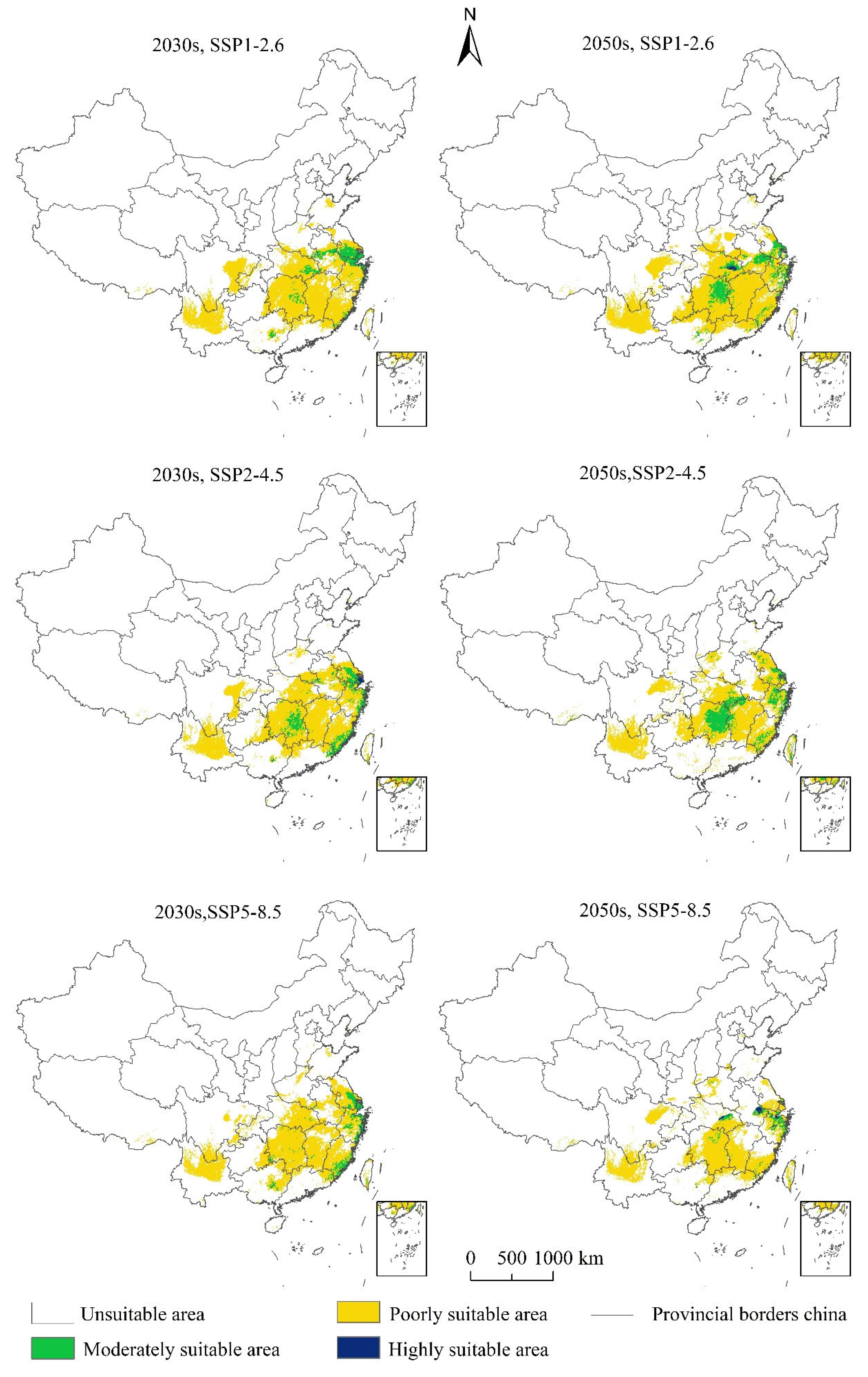
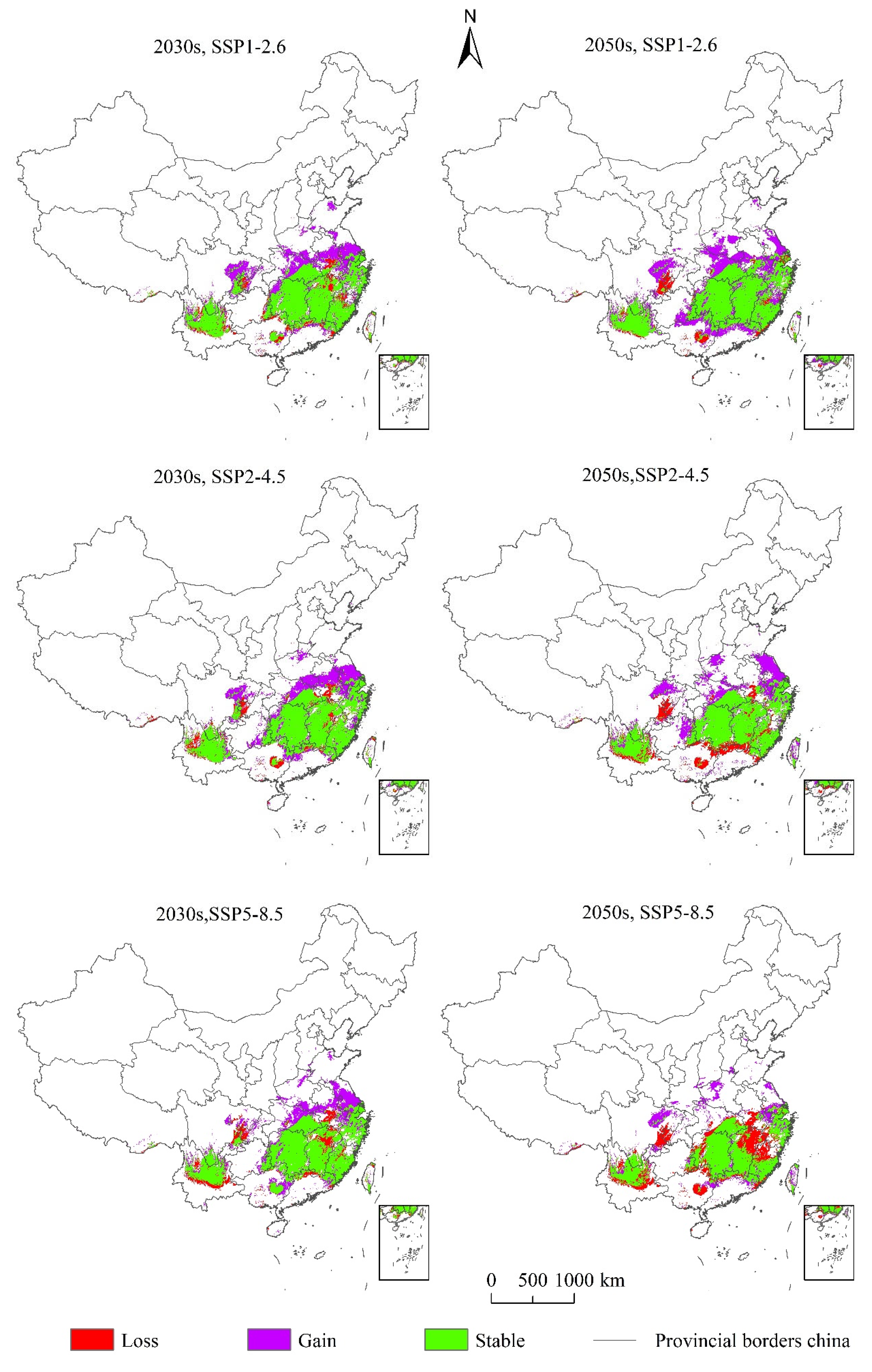
3.4. The Changes Potential Suitable Areas of Linepithema humile under Future Climatic Scenarios
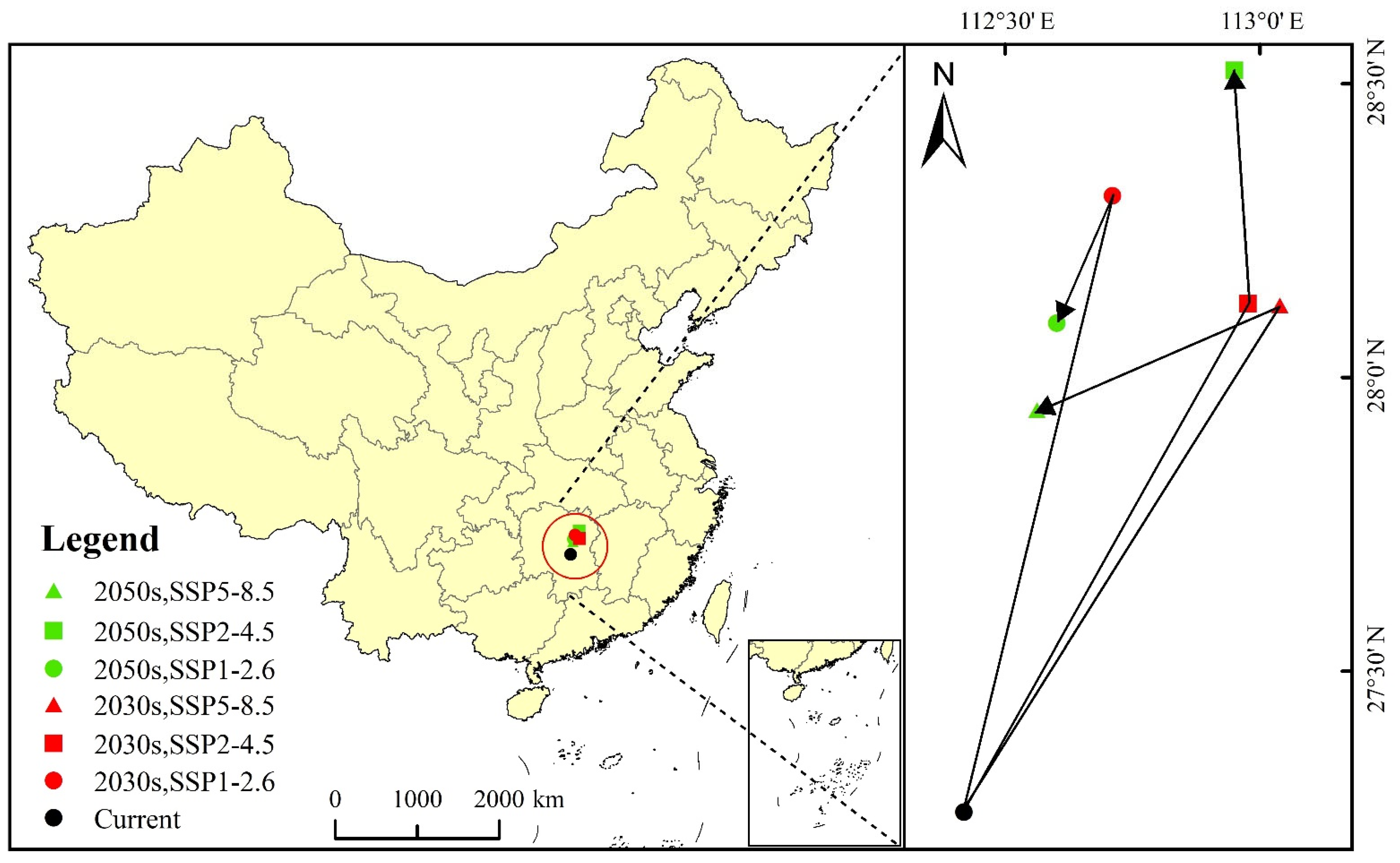
3.5. The Centroid Migration of Potential Suitable Areas for Linepithema humile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, B.X.; Sun, Y.F.; Han, Z.H.; Huang, H.K.; Zhang, H.B.; Li, Y.K.; Zhang, G.L.; Liu, W.X. Challenges in preventing and controlling invasive alien species in China. J. Biosaf. 2020, 29, 157–163. [Google Scholar]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef] [PubMed]
- Seebens, H.; Essl, F.; Blasius, B. The intermediate distance hypothesis of biological invasions. Ecol. Lett. 2017, 20, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Li, H.R.; Yan, J.; Du, C.; Yan, X.L. Current status and suggestions of research on invasive risk assessment of alien plants in China. Acta Ecol. Sin. 2022, 42, 6451–6463. [Google Scholar]
- Wu, J.Q.; Smith, M.T. Successful Approaches for Battling Invasive Species in Developed Countries. Acta Agric. Univ. Jiangxiensis 2010, 32, 1040–1055. [Google Scholar]
- Angulo, E.; Hoffmann, B.D.; Ballesteros-Mejia, L.; Taheri, A.; Balzani, P.; Bang, A.; Renault, D.; Cordonnier, M.; Bellard, C.; Diagne, C.; et al. Economic costs of invasive alien ants worldwide. Biol. Invasions. 2022, 24, 2041–2060. [Google Scholar] [CrossRef]
- Luque, G.M.; Bellard, C.; Cleo, B.; Bonnaud, E.; Genovesi, P.; Simberloff, D.; Courchamp, F. The 100th of the world’s worst invasive alien species. Biol. Invasions. 2014, 16, 981–985. [Google Scholar] [CrossRef]
- Mothapo, N.P.; Wossler, T.C. Behavioural and chemical evidence for multiple colonisation of the Argentine ant, Linepithema humile, in the Western Cape, South Africa. BMC Ecol. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Ingram, K.K. Plasticity in Queen Number and Social Structure in The Invasive Argentine Ant (Linepithema Humile). Evolution 2002, 56, 2008–2016. [Google Scholar] [CrossRef]
- Kennedy, T.A. Patterns of an invasion by Argentine ants (Linepithema humile) in a riparian corridor and its effects on ant diversity. Am. Midl. Nat. 1998, 140, 343–350. [Google Scholar] [CrossRef]
- Lach, L. A mutualism with a native membracid facilitates pollinator displacement by Argentine ants. Ecology 2007, 88, 1994–2004. [Google Scholar] [CrossRef]
- Lach, L. Argentine ants displace floral arthropods in a biodiversity hotspot. Divers. Distrib. 2008, 14, 281–290. [Google Scholar] [CrossRef]
- Ward, D.F.; Green, C.; Harris, R.J.; Hartley, S.; Lester, P.J.; Stanley, M.C.; Suckling, D.M.; Toft, R.J. Twenty years of Argentine ants in New Zealand: Past research and future priorities for applied management. N. Z. Entomol. 2010, 33, 68–78. [Google Scholar] [CrossRef]
- Zhao, H.X.; Xian, X.Q.; Zhao, Z.H.; Zhang, G.F.; Liu, W.X.; Wan, F.H. Climate Change Increases the Expansion Risk of Helicoverpa zea in China According to Potential Geographical Distribution Estimation. Insects 2022, 13, 79. [Google Scholar] [CrossRef]
- Newman, J.A. Climate change and the fate of cereal aphids in Southern Britain. Glob. Chang. Biol. 2005, 11, 940–944. [Google Scholar] [CrossRef]
- Menzel, F.; Feldmeyer, B. How does climate change affect social insects? Curr. Opin. Insect. Sci. 2021, 46, 10–15. [Google Scholar] [CrossRef]
- Zhao, G.H.; Cui, X.Y.; Sun, J.J.; Li, T.T.; Wang, Q.; Ye, X.Z.; Fan, B.G. Analysis of the distribution pattern of Chinese Ziziphus jujuba under climate change based on optimized biomod2 and MaxEnt models. Ecol. Indic. 2021, 132, 108256. [Google Scholar] [CrossRef]
- Wang, H.R.; Zhang, Q.Z.; Liu, R.F.; Sun, Y.; Xiao, J.H.; Gao, L.; Gao, X.; Wang, H.B. Impacts of changing climate on the distribution of Solenopsis invicta Buren in Mainland China: Exposed urban population distribution and suitable habitat change. Ecol. Indic. 2022, 139, 108944. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Ann. Rev. Ecol. Evol. System. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W.; Zimmermann, N.E. Habitat Suitability and Distribution Models: With Applications in R; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- Zhang, X.Q.; Li, G.Q.; Du, S. Simulating the potential distribution of Elaeagnus angustifolia L. based on climatic constraints in China. Ecol. Eng. 2018, 113, 27–34. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Fan, G.; He, Y. Predicting the current and future distribution of three Coptis herbs in China under climate change conditions, using the MaxEnt model and chemical analysis. Sci. Total Environ. 2020, 698, 134141. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.A.; Sui, X.Y.; Lv, Z.; Chen, Y.F. A prediction of the global habitat of two invasive fishes (Pseudorasbora parva and Carassius auratus) from East Asia using MaxEnt. Biodivers. Sci. 2014, 22, 182–188. [Google Scholar]
- Zhang, H.; Song, J.Y.; Zhao, H.X.; Li, M.; Han, W.H. Predicting the distribution of the invasive species Leptocybe invasa: Combining MaxEnt and Geodetector models. Insects 2021, 12, 92. [Google Scholar] [CrossRef]
- Liu, X.Y.; Zhao, C.Y.; Li, F.F.; Zhu, J.F.; Gao, K.X.; Hu, Y.B. Prediction of potential geographical distribution of Solenopsis invicta Buren in China based on MaxEnt. Plant Quar. 2019, 33, 70–76. [Google Scholar]
- Zhang, Y.J.; Ma, F.Z.; Xu, H.G.; Fan, J.Y.; Sun, H.Y.; Ding, H. Prediction of potential geographic distribution of Anoplolepis gracilipes (Homoptera: Formicinae) in China using MaxEnt model. Chin. J. Ecol. 2018, 37, 3364–3370. [Google Scholar]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Wright, A.N.; Seifert, S.N.; Shaffer, H.B. Incorporating model complexity and spatial sampling bias into ecological niche models of climate change risks faced by 90 California vertebrate species of concern. Divers. Distrib. 2014, 20, 334–343. [Google Scholar] [CrossRef]
- Zhu, G.P.; Qiao, H.J. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Bowen, A.K.M.; Stevens, M.H.H. Temperature, topography, soil characteristics, and NDVI drive habitat preferences of a shade-tolerant invasive grass. Ecol. Evol. 2020, 10, 10785–10797. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A., Jr. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Zhu, G.P.; Liu, Q.; Gao, Y.B. Improving ecological niche model transferability to predict the potential distribution of invasive exotic species. Biodivers. Sci. 2014, 22, 223–230. [Google Scholar]
- Moreno, R.; Zamora, R.; Molina, J.R.; Vasquez, A.; Herrera, M.Á. Predictive modeling of microhabitats for endemic birds in South Chilean temperate forests using Maximum entropy (Maxent). Ecol. Inform. 2011, 6, 364–370. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.Z.; Ye, X.Z.; Li, J.H.; Liu, Y.P.; Chen, S.P.; Liu, B. Prediction of potential suitable area of Ulmus elongate in China under climate change scenarios. Chin. J. Ecol. 2021, 40, 3822–3835. [Google Scholar]
- Abril, S.; Oliveras, J.; Gómez, C. Effect of temperature on the development and survival of the Argentine ant, Linepithema humile. J. Insect Sci. 2010, 10, 97. [Google Scholar] [CrossRef] [Green Version]
- Brightwell, R.J.; Labadie, P.E.; Silverman, J. Northward expansion of the invasive Linepithema humile (Hymenoptera: Formicidae) in the Eastern United States is constrained by winter soil temperatures. Environ. Entomol. 2010, 39, 1659–1665. [Google Scholar] [CrossRef]
- Abril, S.; Oliveras, J.; Gómez, C. Effect of temperature on the oviposition rate of Argentine ant queens (Linepithema humile Mayr) under monogynous and polygynous experimental conditions. J. Insect Physiol. 2007, 54, 265–272. [Google Scholar] [CrossRef]
- Battisti, A.; Stastny, M.; Netherer, S.; Robinet, C.; Schopf, A.; Roques, A.; Larsson, S. Expansion of geographic range in the pine processionary moth caused by increased winter temperatures. Ecol. Appl. 2005, 15, 2084–2096. [Google Scholar] [CrossRef]
- Zhu, H.; Wang, D.L.; Wang, L.; Fan, J.; Sun, W.; Ren, B.Z. Effects of altered precipitation on insect community composition and structure in a meadow steppe. Ecol. Entomol. 2014, 39, 453–461. [Google Scholar] [CrossRef]
- Chang, X.N.; Gao, X.N.; Cheng, F.J.; Zhai, B.P. Effects of environmental moisture and precipitation on insects: A review. Chin. J. Ecol. 2008, 27, 619–625. [Google Scholar]
- Menke, S.B.; Holway, D.A. Abiotic factors control invasion by Argentine ants at the community scale. J. Animal Ecol. 2006, 75, 368–376. [Google Scholar] [CrossRef] [Green Version]
- Vega, S.J.; Rust, M.K. The Argentine ant–a significant invasive species in agricultural, urban and natural environment. Sociobiology 2001, 37, 3–25. [Google Scholar]
- Wang, F.; Wang, D.; Guo, G.; Zhang, M.X.; Lang, J.Y.; Wei, J.F. Potential distributions of the invasive barnacle scale Ceroplastes cirripediformis (Hemiptera: Coccidae) under climate change and implications for its management. J. Econ. Entomol. 2020, 114, 82–89. [Google Scholar] [CrossRef]
- Fekrat, L.; Farashi, A. Impacts of climatic changes on the worldwide potential geographical dispersal range of the leopard moth, Zeuzera pyrina (L.) (Lepidoptera: Cossidae). Glob. Ecol. Conserv. 2022, 34, e02050. [Google Scholar] [CrossRef]
- Buczkowski, G.; Bertelsmerier, C. Invasive termites in a changing climate: A global perspective. Ecol. Evol. 2017, 7, 974–985. [Google Scholar] [CrossRef]
- Li, X.; Ge, X.Z.; Chen, L.H.; Zhang, L.J.; Wang, T.; Zong, S.X. Climate change impacts on the potential distribution of Eogystia hippophaecolus in China. Pest. Manag. Sci. 2018, 75, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Song, J.Y.; Zhang, H.; Li, M.; Han, W.H.; Yin, Y.X.; Lei, J.P. Prediction of spatiotemporal invasive risk of the red import fire ant, Solenopsis invicta (Hymenoptera: Formicidae), in China. Insects 2021, 12, 874. [Google Scholar] [CrossRef]
- Ji, Y.L.; Su, X.Y.; Yu, Z.J. Potential habitat prediction of Hyphantria cunea based on random forest model in China. J. Nanjing Univ. Nat. Sci. 2019, 43, 121–128. [Google Scholar]
- Zhang, V.M.; Punzalan, D.; Rowe, L. Climate change has different predicted effects on the range shifts of two hybridizing ambush bug (Phymata, Family Reduviidae, Order Hemiptera) species. Ecol. Evol. 2020, 10, 12036–12048. [Google Scholar] [CrossRef]
- Chapman, J.W.; Bell, J.R.; Burgin, L.E.; Reynolds, D.R.; Pettersson, L.B.; Hill, J.K.; Bonsall, M.B.; Thomas, J.A. Seasonal migration to high latitudes results in major reproductive benefits in an insect. Proc. Natl. Acad. Sci. USA 2012, 109, 14924–14929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpintero, S.; Reyes-López, J.; Reyna, L.A.D. Impact of Argentine ants (Linepithema humile) on an arboreal ant community in Doñana National Park, Spain. Biodivers. Conserv. 2005, 14, 151–163. [Google Scholar] [CrossRef]
- Suarez, A.V.; Holway, D.A.; Case, T.J. Patterns of spread in biological invasions dominated by long-distance jump dispersal: Insights from Argentine ants. Proc. Natl. Acad. Sci. USA 2001, 98, 1095–1100. [Google Scholar] [CrossRef] [Green Version]
- CABI. Invasive Species Compendium of the Center for Agriculture and Bioscience International; CAB International: Wallingford, UK, 2022; Available online: https://www.cabi.org/isc (accessed on 28 June 2022).
- Orr, M.R.; Seike, S.H.; Benson, W.W.; Dahlsten, D.L. Host specificity of Pseudacteon (Diptera: Phoridae) parasitoids that attack Linepithema (Hymenoptera: Formicidae) in South America. Environ. Entomol. 2001, 30, 742–747. [Google Scholar] [CrossRef]
- Rust, M.K. Insecticides and their use in urban structural pest control. In Handbook of Pesticide Toxicology, 2nd ed.; Academic Press: San Diego, CA, USA, 2001; pp. 243–250. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description | Unit | Whether to Use for Modeling |
|---|---|---|---|
| Bio1 | Annual mean temperature | °C | No |
| Bio2 | Mean diurnal air temperature area | °C | Yes |
| Bio3 | Isothermality (bio2/bio7) (*100) | — | No |
| Bio4 | Temperature seasonality (standard deviation *100) | — | No |
| Bio5 | Max temperature of warmest month | °C | Yes |
| Bio6 | Min temperature of coldest month | °C | No |
| Bio7 | Temperature annual area (bio5-bio6) | °C | Yes |
| Bio8 | Mean temperature of wettest quarter | °C | Yes |
| Bio9 | Mean temperature of driest quarter | °C | No |
| Bio10 | Mean temperature of warmest quarter | °C | No |
| Bio11 | Mean temperature of coldest quarter | °C | Yes |
| Bio12 | Annual precipitation | mm | No |
| Bio13 | Precipitation of wettest month | mm | No |
| Bio14 | Precipitation of driest month | mm | No |
| Bio15 | Precipitation seasonality (coefficient of variation) | — | Yes |
| Bio16 | Precipitation of wettest quarter | mm | Yes |
| Bio17 | Precipitation of driest quarter | mm | Yes |
| Bio18 | Precipitation of warmest quarter | mm | Yes |
| Bio19 | Precipitation of coldest quarter | mm | Yes |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Xian, X.; Zhao, H.; Xue, L.; Chen, B.; Huang, H.; Wan, F.; Liu, W. Predicting the Potential Suitable Area of the Invasive Ant Linepithema humile in China under Future Climatic Scenarios Based on Optimized MaxEnt. Diversity 2022, 14, 921. https://doi.org/10.3390/d14110921
Li M, Xian X, Zhao H, Xue L, Chen B, Huang H, Wan F, Liu W. Predicting the Potential Suitable Area of the Invasive Ant Linepithema humile in China under Future Climatic Scenarios Based on Optimized MaxEnt. Diversity. 2022; 14(11):921. https://doi.org/10.3390/d14110921
Chicago/Turabian StyleLi, Ming, Xiaoqing Xian, Haoxiang Zhao, Lin Xue, Baoxiong Chen, Hongkun Huang, Fanghao Wan, and Wanxue Liu. 2022. "Predicting the Potential Suitable Area of the Invasive Ant Linepithema humile in China under Future Climatic Scenarios Based on Optimized MaxEnt" Diversity 14, no. 11: 921. https://doi.org/10.3390/d14110921