DNA Barcodes and Genetic Analyses
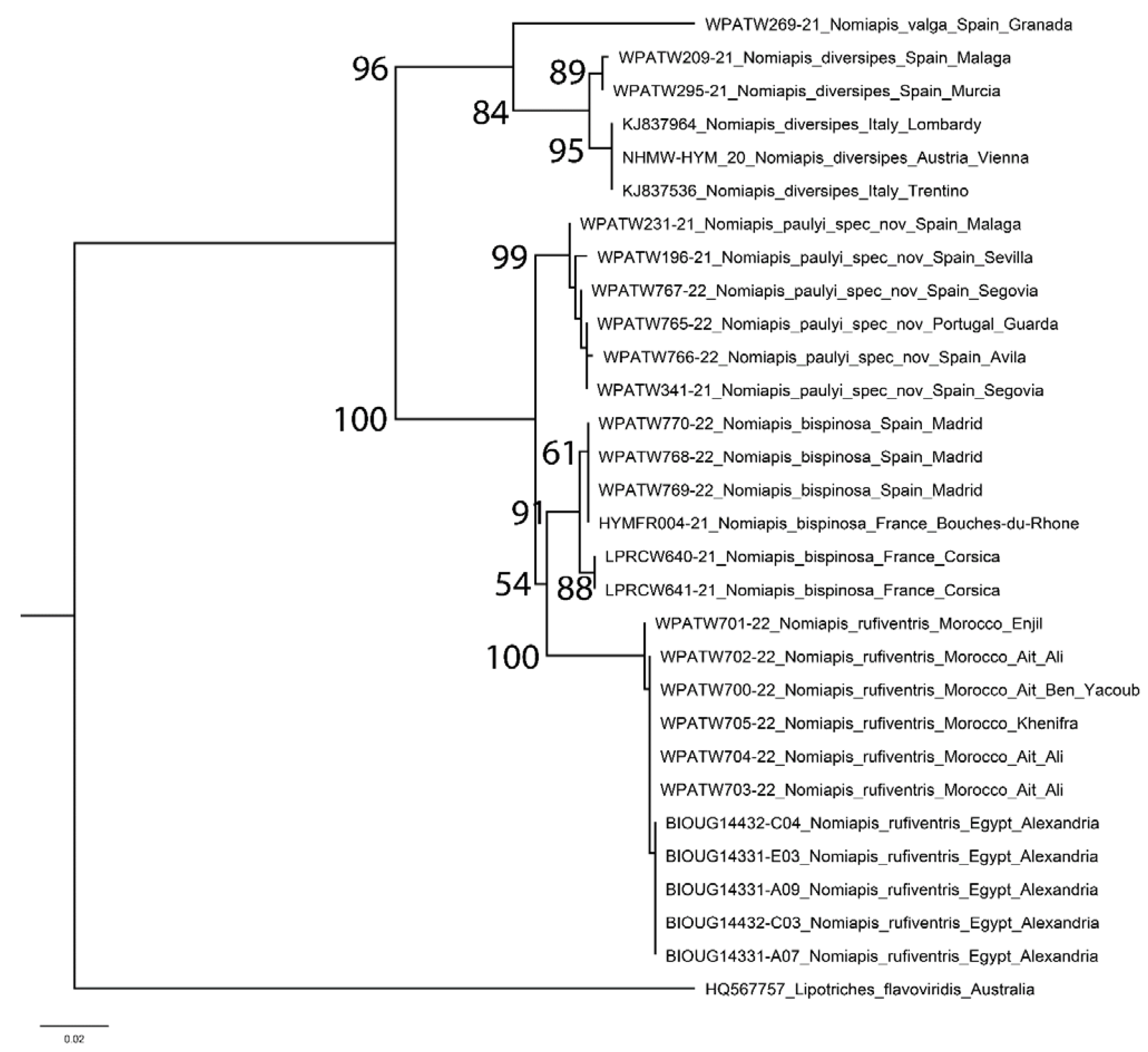
For the genus
Nomiapis, five taxa were identified (
Figure 1).
Nomiapis diversipes (Latreille, 1806) was recovered as a monophyletic clade with bootstrap support of 84. Iberian specimens and Italian/Austrian specimens have identical sequences within each region, but between region distance was an average of 0.96%, leading to high bootstrap support for each clade (89 and 95, respectively) despite low overall differentiation.
Nomiapis bispinosa s.l. contained three distinct lineages. Material from Morocco and Egypt strongly clustered together, showing an average intraspecific distance of 0.14% (range 0.00–0.38%,
Table 2), this clade having bootstrap support of 100. This clade is referable to as
Nomiapis rufiventris (Spinola, 1838)
sp. resurr. (see below). Material from Portugal and Spain also strongly clustered together, showing an average intraspecific distance of 0.34% (range 0.00–0.77%), this clade having bootstrap support of 99. This clade represents a previously undescribed species,
Nomiapis paulyi spec. nov. (see below). This new species is separated from
Nomiapis rufiventris by an average of 3.42% (range 2.87–3.83%).
Nomiapis bispinosa s. str. formed a monophyletic clade with a lower bootstrap support of 91. This is due to the variation of sequences from specimens from Corsica, which were identical to each other, but which varied by 0.77% from the sequences from Spain and mainland France, which were themselves identical to each other. These two clades within
Nomiapis bispinosa s. str. have moderate bootstrap support (88 and 61, respectively), though as overall genetic differentiation is low, it is not considered to correspond to a species-specific difference.
Nomiapis bispinosa s. str. as a whole is separated from
Nomiapis rufiventris by 2.83% (range 2.49–3.07) and from the new
Nomiapis species by 2.14% (range 1.72–2.68%). Across all
Nomiapis species, average interspecific differences were consistently higher (range 3.60–7.27%) than average intraspecific differences (range 0.14–0.57%), and hence a five taxon conclusion was drawn.
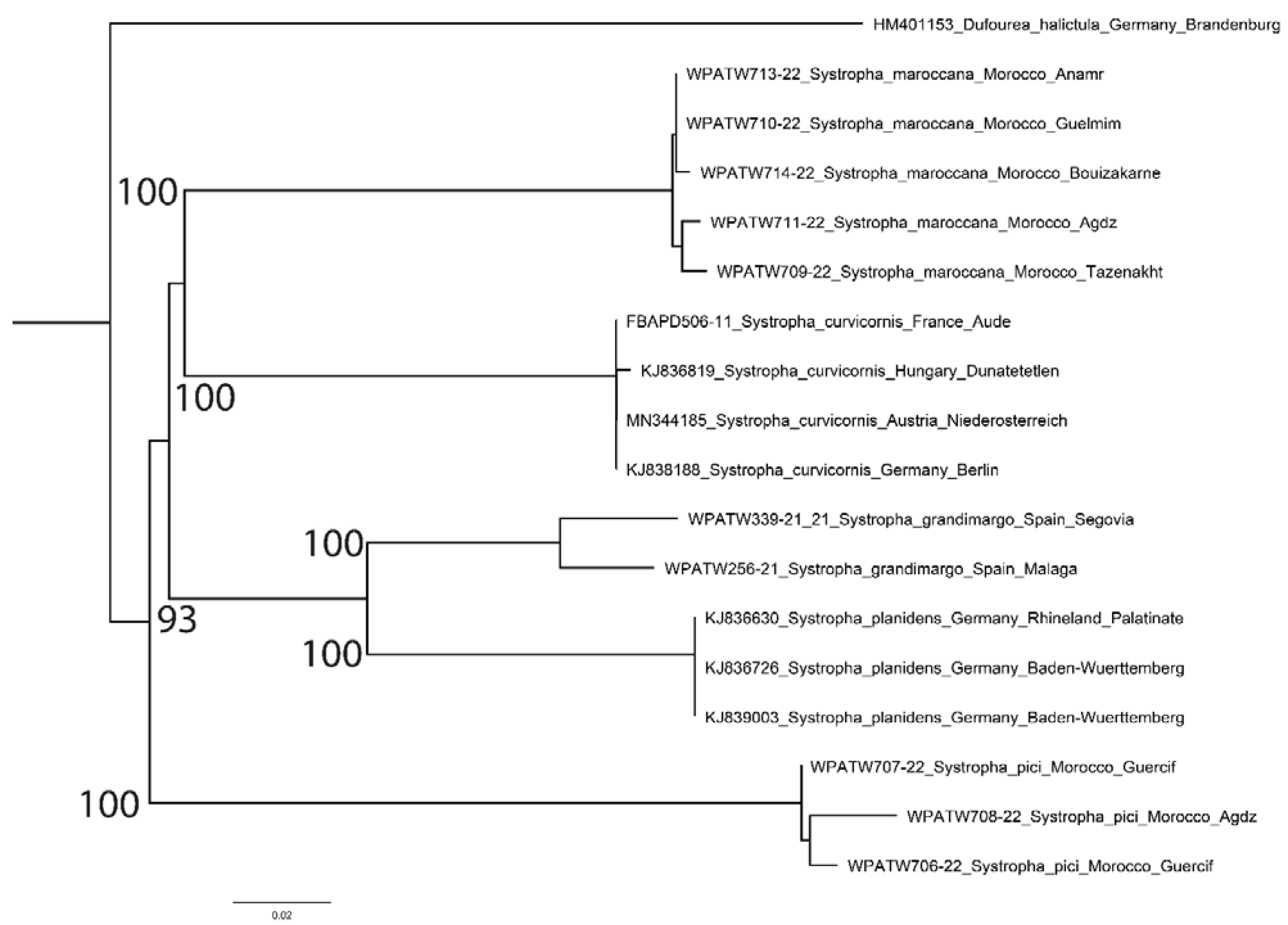
For the genus
Systropha, all species were well-resolved, with bootstrap values of 100 for five taxa, including
S. planidens grandimargo (
Figure 2). Intraspecific distances were low, typically less than 2%, but reaching 4.23% in
S. p. grandimargo (
Table 2). However, average interspecific distances between
Systropha species were very high. The species showing the lowest divergence,
S. p. grandimargo, had average interspecific distances between 10.27–19.34%, whereas the species showing the greatest distances,
S. pici, had average interspecific distance between 18.13–22.96%.
Systropha p. grandimargo was separated from its nearest relative
S. planidens s. str. by an average of 10.73% (range 10.23–11.18%). It is therefore treated here as a distinct species,
S. grandimargo spec. resurr. Across all
Systropha species, average interspecific differences were consistently higher (range 16.06–20.22%) than average intraspecific differences (range 0.00–4.23%), and hence a five-taxon conclusion was drawn.
Taxonomic accounts
Genus Nomiapis Cockerell, 1919
Nomiapis bispinosa (Brullé, 1832)
Nomia bispinosa Brullé, 1832: 348; ♂ surroundings of Mistra [Mystras], Greece [MNHN, lectotype by present designation]
Nomia ruficornis Spinola, 1838: 514; ♂ Egypt [MRSN, lectotype by present designation]
Nomia aureocincta Costa, 1861: 8; ♂♀ San Luca, Calabria Ulteriore, Italy [IEAN, types undesignated, not examined]. Synonymy with
Nomia unidentata by Warncke [
8].
Nomia polita Costa, 1861: 11; ♀ Napoli, Italy [exact type location unknown, see Baker [
11]]. Synonymy with
Nomia unidentata by Warncke [
8].
Nomia basalis Smith, 1875: 55; ♀. India [NHMUK, holotype, examined]. Synonymy by Baker [
11].
Nomia aureocincta var.
turcomanica Radoszkowski, 1893: 54; ♂♀ “
Askhabad ou de ses environs”, Turkmenistan, [IZK, ♀ lectotype designated by Astafurova & Pesenko [
19]]. Synonymy with
Nomia rufiventris Spinola, 1838 by Popov (1935), then automatically with
Nomiapis bispinosa by Baker [
11].
Nomia fletcheri Cockerell, 1920: 207; ♀. Peshawar District, Tarnab, India [USNM, holotype, not examined]. Synonymy by Pauly [
12].
Nomia basalicincta Cockerell, 1922: 663; nom. nov. pro Nomia basalis Smith, 1875.
Type material examined.—Lectotype of
Nomia bispinosa Brullé (present designation): 767 [species code referring to the original description]//Brullé Morée//Au Mu-séum//
N. bispinosa Br. 767.//A. PAULY DT 1984
Nomiapis unidentata (Olivier)//Lectotype
Nomia bispinosa Brullé Le Divelec des. 2022//
Nomiapis bispinosa (Brullé, 1832) ♂ Le Divelec det. 2022//MNHN, Paris EY33638 (
Figure 3).
–Lectotype of
Nomia ruficornis Spinola (present designation): ♂//
Nomia ruficornis ♂ Spinola, 1838 LECTOTYPE des. WOOD 2022//
Nomia ruficornis, m. ♂, ann. soc. ent. D. Waltl. Egypte (
Figure 4).
Other material examined (illustrative, see Table S1). AFGHANISTAN: 1♂, Herat province, Koshk district, 24.vi.1997, leg. G.G.M. Schulten, RMNH;
BULGARIA: 1♀, Warna [Varna], 5.viii.1956, leg. W.J. Pulawski, RMNH;
CROATIA: 1♀, Lussingrande [Veli Lošinj], 24.vii.1924, leg. Dr. Fudakowski, RMNH;
CYPRUS: 1♂, Akrotiri, SW of Limassol, 25.vi.1971, leg. M.J. & J.P. Duffels, RMNH;
EGYPT: 1♂, Ghizeh, Reg. du Caire, 15.iv.1958, leg. W.J. Pulawski, RMNH;
FRANCE: 1♂, 2♀, Pyr. Or., la Barcarès (Lido), 8.viii.1950, leg. P.M.F. Verhoeff, RMNH;
GREECE: 17♀, Cyclades, Naxos, dunes S. of Naxos (village), 24–26.vii.1975, leg. Ph. Pronk, RMNH;
HUNGARY: 1♂, Budapest, Mocsáry, RMNH;
IRAN: 1♀, Mollasani, Ahwaz, 28.ii.1943, MNHN;
ISRAEL: 1♂, Negev, Bir Reckene [Bir Rekhme, =Be’er Yeruham], 14.vi.1919, leg. Bytinski-Salz, RMNH;
ITALY: 1♂, Bibione/Adria, 3–15.viii.1969, leg. H. Wolf, RMNH;
KAZAKHSTAN: 1♂, 1♀, Tschelkar, Grosse-Barsuki, Barbier collection, MNHN;
MALTA: 1♀, N. Gozo, NW of Zaghra, nr. Ir. Ramla [=Ramla Bay], 30.ix.1980, leg. Ph. Pronk, RMNH;
MOROCCO: 2♂, Tanger, 1–30.viii.1895, Barbier collection, MNHN;
PORTUGAL: 1♀, Algarve, Mexilhoeira Grande, 19.viii.2015, leg. J. D’Haeseleer, TJWC;
ROMANIA: 1♀, Mamaia, 6.vi.1961, K. Bleyl, RMNH;
SERBIA: 2♀, Deliblato, 23.vii.1886, André collection, MNHN;
SPAIN: 24♂, 2♀, Almería, El Alquián, 20.v.1960, RMNH;
TURKEY: 2♂, Pamukale, 15 km N of Denizli, 13.vii.1980, H. v. Oorschot, RMNH;
TURKMENISTAN: 1♂, Achkhabad, Grosse-Barsuki, 25.v.1916, Barbier collection, MNHN;
UKRAINE: 1♂, Измаилъская oбл [Izmail Oblast] oкр г. Килия [district of Kiliya], 11.vi.1951, leg. А. Осыцнюк [A. Osytshnjuk], RMNH.
Remarks. The type material of
Nomiapis bispinosa was considered to be lost [
8,
11]. Careful examination of the MNHN collection has resulted in the discovery of one male from the Brullé collection collected during the ‘Morée,’ which was the Morea expedition [
Expédition de Morée] by the French army during 1828–1833 to the Peloponnese, which was accompanied by a scientific commission [
Expédition Scientifique de Morée]. The specimen was labelled as
Nomia bispinosa by Brullé (code 767). Vachal [
31] said he examined the type of
Nomia bispinosa and, given that most species described by Brullé are single specimens, it is probably a holotype. It is not possible to conclude this from the original description, so the present specimen is designated as the lectotype.
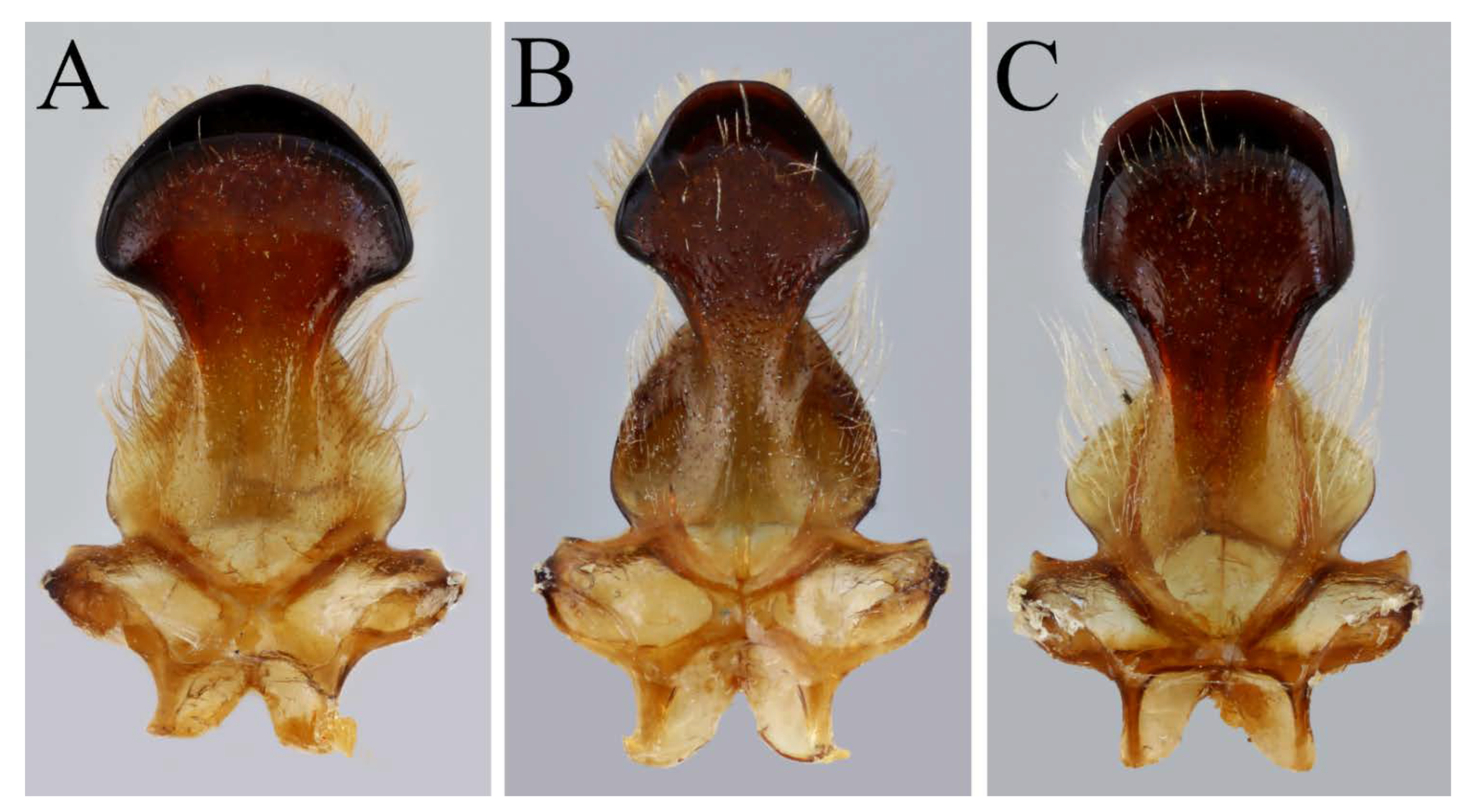
Nomiapis bispinosa can be separated from its two closely related sister species predominantly by the density of the punctures of the scutum in the female sex and the shape of the pair of raised tubercles on S6 in the male sex, see
Table 3.
Spinola [
1] published two
Nomia species based on Egyptian material, citing the French expedition to the country in 1798–1801, “
Exp. d’Eg.,
loc. cit.”, referring to Savigny [
32]. Note that Baker [
11] lists the publication of Spinola as 1839, but this is not the position of other authors (e.g., [
33]). We follow their position that the year of publication is 1838.
Nomia rufiventris Spinola, 1838 was described in the female sex and
Nomia ruficornis Spinola, 1838 was described in the male sex. It was not clear that the two were conspecific, as Gerstäcker [
4] used the name
Nomia ruficornis without mentioning
Nomia rufiventris. Friese [
34] considered that
Nomia rufiventris might be a reddish female of
Nomia ruficornis but did not propose a synonymy. Alfken [
6] synonymised the two names under
Nomia rufiventris, but it is unclear if he examined type material.
We examined the Spinola collection in Turin and were able to locate the type material. Standing above the label of
Nomia rufiventris (individual specimens are unlabelled, and one must refer to the labels pinned within the box itself, see Casolari & Casolari 1980) was a male
Anthidium specimen. This is clearly a mistake since both the sex and family are incorrect. Standing to the right of the
Nomia rufiventris label is a label for ‘
Nomia calceata,’ which is listed (see [
35] for an explanation of the numerical numbering system used in the Spinola collection) as a taxon described by Drège, the specimen coming from ‘
Cap de Bonne-Espérance’ [Cape of Good Hope, South Africa], from the collection of Drège, with nominally zero specimens placed here. However, a single female
Nomiapis specimen with red metasomal terga labelled ‘Egitto?’ [Italian for Egypt] was placed here (
Figure 5). The name ‘
Nomia calceata’ is unpublished, and as
Nomiapis species are unknown from sub-Saharan Africa, this specimen has clearly been misplaced and is, in fact, the type of
Nomia rufiventris. It is here designated as the lectotype. It may automatically be the holotype, but as Spinola did not indicate how many specimens he examined, this is not clear.
Examination of the specimen reveals that it is not conspecific with
Nomiapis bispinosa, as thought by Warncke ([
8], as
Nomia unidentata) and Baker [
11]. The scutal punctation is relatively coarse and sparse, suggesting
Nomiapis bispinosa, but Egyptian specimens of the North African
Nomiapis taxon display this character state when compared to material from Morocco and Algeria (see
Table 3). Importantly, the punctation of the marginal area of T2 covers ½ of its area (
Figure 5D), whereas, in
Nomiapis bispinosa, the punctures cover only 1/3rd of the marginal area of T2 (
Figure 6). The disc of T2 also has two lateral impunctate zones mediolaterally (
Figure 5D), which are absent in
Nomiapis bispinosa (
Figure 6).
Nomiapis rufiventris is, therefore, not conspecific with
Nomiapis bispinosa and is the valid name for the North African
Nomiapis taxon (see below). The red terga are not significant, as bee specimens from hot regions often display red colouring that is not expressed in specimens from cooler climes, as can be seen within
Andrena afzeliella (Kirby, 1802) from Egypt (
Andrena afzeliella var.
heliopolis Friese, 1914, [
17]). A final note, though the description of
Nomia rufiventris Spinola [
1] mentions that the taxon is illustrated in Savigny [
32] on “
pl. 5, Figure 17, ♀”, this illustration displays what appears to be a female
Lipotriches parca (Kohl, 1906). We believe that this is a misnumbering or typographical error by Spinola, as the lectotype specimen is clearly a
Nomiapis with large tegulae.
The type of
Nomia ruficornis was located in its expected place (
Figure 4). This specimen belongs to
Nomiapis bispinosa due to its comparatively large body size, depression of T3 sharply depressed and with a sharp carina separating it from the tergal disc, and the marginal area with only a single line of very narrow punctures basally. It is also designated as a lectotype, following the same logic as for
Nomia rufiventris. Therefore, its synonymy with
Nomiapis bispinosa, as listed by Baker [
11], is confirmed to be correct.
One outstanding issue is the identity of
Nomia aureocincta Costa, 1861. The original description does not allow a clear distinction because the males of
Nomiapis bispinosa and
Nomiapis rufiventris are morphologically extremely similar. Both species occur in Sicily and might occur in the neighbouring region of Calabria. Warncke [
8] synonymised
Nomia aureocincta with
Nomia unidentata, but type revision will be needed to see if
Nomia aureocincta belongs to
Nomiapis bispinosa s. str. or to
Nomiapis rufiventris. We also consider
Nomia polita Costa, 1861 to be a probable synonym of
Nomiapis bispinosa, as suggested by Warncke [
8]. Baker [
11] placed it with doubts among the synonyms of
Nomiapis monstrosa (Costa, 1861) without any justification except that of the misunderstanding of Warncke [
8] regarding the application of the name of
Nomia unidentata. Astafurova & Pesenko [
19] list the taxon as a synonym of
Nomiapis bispinosa. The description of
Nomia polita fits
Nomiapis bispinosa better (hair fringes on T2-4; scutal punctuation sparse), and this taxon is much more common in Italy than
Nomiapis monstrosa and more likely to be collected around Naples, though this should be confirmed by type examination.
In Iberia, France, and Corsica,
Nomiapis bispinosa is strongly halophilic, nesting in saline soils, both on the coast and also inland in central Spain on salt steppe ([
36], as
Pseudapis monstrosa (Costa, 1861), misidentification; RLD & TJW
pers. obs.). It is, therefore, less commonly encountered than
Nomiapis paulyi (see below), which can be found on a wide range of soil types, from coastal sands to calcareous steppe soil. The distribution and nesting ecology of
Nomiapis bispinosa s. str. should be carefully studied in light of these taxonomic changes to confirm if this behaviour is consistent across its distribution, though based on material studied here (
Figure 7), it does appear to be strongly associated with coastal environments or salt steppe at inland localities. It is parasitised by
Pasites maculatus Jurine, 1807 (36; RLD & TJW
pers. obs.). Very few pollen loads were available, but
Nomiapis bispinosa was observed to collect pollen from
Limonium (Plumbaginaceae) in Central Spain, reflecting its halophilic distribution, as well as from
Onopordum (Asteraceae,
Table 4). Additional samples will confirm that
Nomiapis bispinosa is polylectic, as is the case in other
Nomiapis species.
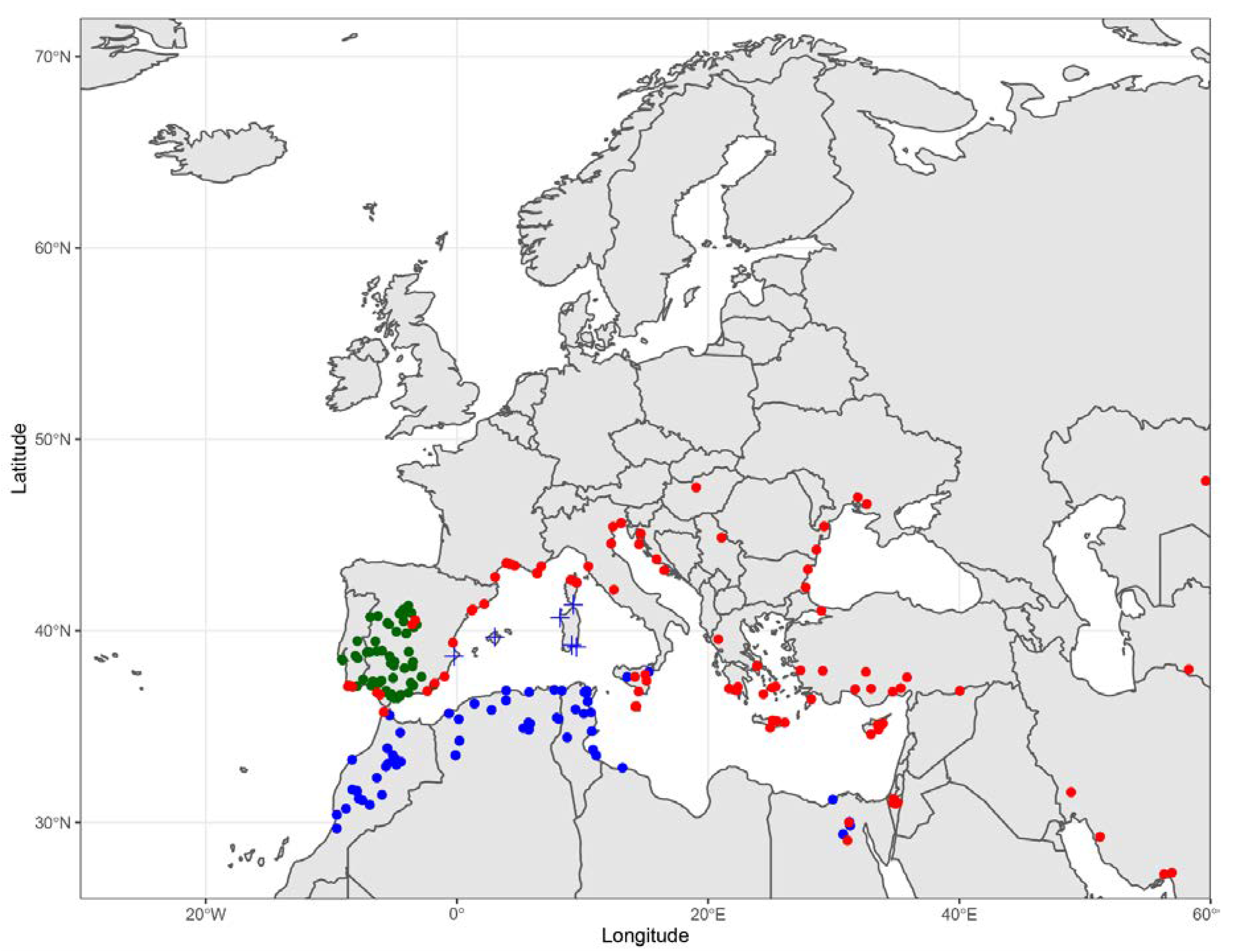
Distribution. In North Africa, we could only confirm its occurrence in Tangier (northern Morocco) and Egypt. The distribution is, therefore, northern Morocco, Iberia, southern France (including Corsica), Italy, southern parts of Central Europe (Hungary, Ukraine), Balkan Peninsula, Turkey, Cyprus, Egypt, Israel (Negev), Iran, the Caucasus to Central Asia, northern India, and north-western China (
Figure 7.
Nomiapis rufiventris (Spinola, 1838) sp. resurr.
Nomia rufiventris Spinola, 1838: 513; ♀ Egypt [MRSN, lectotype by present designation].
Nomia albocincta Lucas, 1849: 187; ♀ La Calle, Lac Tonga, Algeria [MNHN, lectotype by present designation] syn. nov.
Nomia perforata Lucas, 1849: 185; ♂ La Calle, Lac Tonga, Algeria [MNHN] syn. nov.
Type material examined.—Lectotype of
Nomia rufiventris Spinola (present designation): 183//Egitto?//
Nomia rufiventris ♀ Spinola, 1838 LECTOTYPE des. WOOD 2022//
Nomia rufiventris, ♀ m. Ann. Soc. ent. D. Waltl. Egypte (
Figure 5).
–Lectotype of
Nomia albocincta Lucas (present designation): Blue circle [underside with the locality code 1525 referring Lucas’s manuscript)//
Nomia albocincta, Luc. ♀//LECTOTYPE//Lectotype
Nomia albocincta Lucas Le Divelec des. 2022//
Nomiapis albocincta (Lucas, 1849) Le Divelec det. 2022//MNHN, Paris EY33640 (
Figure 8 and
Figure 9).
–Paralectotype of Nomia albocincta Lucas: Blue circle [underside with the locality code 1525 referring Lucas’s manuscript)//Nomia albocincta, Luc.//PARALECTOTYPE//Paralectotype Nomia albocincta Lucas Le Divelec des. 2022//Nomiapis albocincta (Lucas, 1849) Le Divelec det. 2022.
–Holotype of
Nomia perforata Lucas: Blue circle [underside with the locality code 1258 referring Lucas’s manuscript)//
Nomia armata, Luc. ♂//
Nomia albocincta, Lucas est la femelle de la
Nomia armata Lucas ♂//HOLOTYPE//Holotype
Nomia perforata Lucas—Le Divelec 2022//
Nomiapis albocincta (Lucas, 1849) Le Divelec det. 2022//MNHN, Paris EY33639 (
Figure 10).
Other material examined (illustrative, see Table S1). ALGERIA: 1♀, La Calle [El Kala], 26.vii.1910, Ferton collection, MNHN;
EGYPT: 1♀, Cairo, 13.vi.1939, MNHN;
ITALY: 1♂, Sicily, Pérez collection, MNHN;
LIBYA: 1♂, Tripolis, OÖLM;
MOROCCO: 1♂, 4♀, Fès-Meknès, Midelt, R503, W of Aït Ali, Oued Taourda, 1550 m, leg. T.J. Wood, TJWC;
TUNISIA: 1♂, Djerba, 14.iv.1955, leg. P.M.F. Verhoeff, RMNH.
Unclear material (Nomiapis cf. rufiventris). FRANCE: 1♀, Corsica, Bonifacio, Cavallo Island, 24.ix.1898, Ferton collection, MNHN; ITALY: 1♂, Sardinia, Cagliari, 15–20.vii.1959, leg. H. Wolf, RMNH; 1♀, Sardinia, Villasimius, 1–30.vi.1975, leg. Bouček, RMNH; SPAIN: 1♀, Benifato, Alicante, 12.vi.1978, leg. H. Teunissen, RMNH; 2♂, Mallorca, 1–24.vi.1954, leg. Klokke—Moll, RMNH.
Remarks. Genetic results strongly support a North African taxon from Morocco to Egypt, with this taxon extending north into Sicily based on morphology. As discussed above, the oldest name for this taxon is
Nomiapis rufiventris. The next oldest names from North Africa come from Lucas [
2]. Lucas mentioned four specimens for the three species of
Nomia described from Algeria, and these remain in his collection. Two specimens are his females of
Nomia albocincta, one is the holotype of
Nomia flavilabris Lucas, 1849 which is actually a male of
Ancyla oraniensis Lepeletier, 1841 and not a female of
Nomia despite being described as such, and one is a male
Nomiapis labelled as “
armata Lucas”.
Nomia armata Lucas was never described. The collecting station code of this specimen is 1258. It is similar to the code used for
Nomia flavilabris and refers to Lac Tonga in June, according to the unpublished catalogue of Lucas, which is housed at the MNHN. As mentioned by Lucas [
2],
Nomia flavilabris was collected in the same place and at the same time as
Nomia perforata, which was in June 1841, around Lac Tonga. For this reason, we consider the present specimen to be the holotype of
Nomia perforata which was not properly labelled after its description. The holotype matches the original description and drawings. Vachal [
31], who examined the type of
Nomia perforata, considered it to be the same species as
Nomia albocincta. Lucas also added a label to this specimen stating that
Nomia armata Lucas was the male of
Nomia albocincta Lucas. Examination of the type confirms these statements. Both names are synonymised with
Nomiapis rufiventris.
Nomiapis rufiventris can be most easily recognised in the female sex by the punctation of the scutum. The punctation is finer and denser than in
Nomiapis bispinosa but less dense than in
Nomiapis paulyi (see below). It is, therefore, intermediate between the two. Males can be separated from
Nomiapis bispinosa by the smaller tubercles of S6, but separation from males of
Nomiapis paulyi is challenging, and reference is best made to the determination table below (
Table 3). As mentioned, female Egyptian
Nomiapis rufiventris have the punctation of the scutum a little less dense than in specimens from north-western Africa and thus begin to approach the condition in
Nomiapis bispinosa s. str. However, barcoded specimens from Egypt and Morocco showed almost no genetic differentiation (maximum 0.38%) and hence are considered conspecific.
Specimens from the Balearic Islands (Mallorca), Sardinia (reported as
Pseudapis unidentata albocincta by [
37]), a single female specimen from Corsica, and a single female specimen from eastern Spain (Alicante) have an ambiguous morphology that is closest to
Nomiapis rufiventris, but they could potentially belong to
Nomiapis paulyi. In the absence of genetic data from these regions, we refrain from a decisive determination and await further genetic work to decisively settle their placement.
Distribution. Morocco, Algeria, Tunisia, Italy (Sicily and southern Italy), Libya, and Egypt (
Figure 7). Based on re-examined material (RMNH), records of
Pseudapis (
Nomiapis)
unidentata albocincta from Malta [
37] are
Nomiapis bispinosa (see
Table S1). In Europe,
Nomiapis rufiventris occurs in Sicily. This record is based on two males. The identity of Sicilian material, as well as ambiguous material from the Balearics, Corsica, and Sardinia, should be confirmed in the future by genetic barcodes.
Nomiapis paulyi Wood & Le Divelec, spec. nov.
Holotype. SPAIN: 1♀, Segovia, Camino Natural Vía Verde Valle del Eresma, 40.9286° N, −4.1210° W, 18.vii.2019, leg. TJWC, RMNH. Barcoded, reference TJW_028, BOLD WPATW767-22.
Paratypes. PORTUGAL: 1♂, 2♀, Alg. [Algarve] Pr. do Castela [Praia do Castelo], 1–15.vii.1977, leg. P.M.F. Verhoeff, RMNH; 1♂, Alto Alentejo, bij Aquaduct van Elvas, langs N 4, 28.vii.1970, leg. E.N. Kuijper, RMNH; 3♂, 4♀, Alto Alentejo, lands E 4 tuss. Montemor O Novo & Arraiolos, 28.vii.1970, leg. Ph. Pronk, RMNH; 3♂, Evora, 24.vii.1953; leg. P.M.F. Verhoeff, RMNH; 1♂, Gavião, Portall.P., 31.v.1979, leg. H. Teunissen, RMNH; 1♂, Algarve, Santa Bárbara de Nexe, 20.ix.1980, leg. H. Teunissen, RMNH; 2♂, Algarve, Lagos, 3.viii.1968, leg. K. Warncke, OÖLM; 1♂, Algarve, Alcoutim, 24.viii.1968, leg. K. Warncke, OÖLM; SPAIN: 1♀, 10 km SW El Escorial, 1200 m, 23.vii.1972, leg. M.J. & J.P. Duffels, RMNH; 1♂, Alcaracejos, Córdoba, 19.vi.1961, leg. J. v. d. Vecht, RMNH; 1♀, Aznalcázar, S of Pinares de Aznalcázar, 21.v.2021, leg. T.J. Wood, TJWC; 7♂, Badajoz, Mérida, 29–30.vi.1968, leg. P.M.F. Verhoeff, RMNH; 1♂, Benalauría, Puerto de Benalauría, 1.vi.2021, leg. G. Ghisbain, G. Ghisbain collection (Mons, Belgium); 1♂, Málaga, Benaoján, 2.viii.1978, leg. R. Leys, RMNH; 1♂, Segovia, Bernuy de Porreros, Fuente de los Caños, 20.vii.2021, leg. T.J. Wood, TJWC; 10♂, 1♀, Cádiz, Jerez de la Frontera, 10.ix.1961, leg. P.M.F. Verhoeff; RMNH; 1♀, Jerez de la Frontera, 1.viii.1964, leg. P.M.F. Verhoeff; RMNH; 2♂, 1♀, Jerez de la Frontera, 15.ix.1965, leg. P.M.F. Verhoeff; RMNH; 1♂, Segovia, Carbonero el Mayor, 2 km SW, Calle las Tejeras to Río Eresma, 18.vii.2021, leg. T.J. Wood, TJWC; 1♀, Segovia, Carbonero el Mayor, 500 m W, Calle las Tejeras, 18.vii.2021, leg. T.J. Wood, TJWC; 1♂, Écija, 18.ix.1952, leg. Bär, Blöte, de Jong, Osse, RMNH; 1♂, El Bosque, Cádiz, 27.vii.1978, leg. R. Leys, RMNH; 1♂, 1♀, Ávila, El Losar del Barco, 2 km N, 17.vii.2020, leg. T.J. Wood, TJWC; 1♂, 3♀, env. Calahonda, betw. Fuengirola-Marbella; 4–9.ix.1975, leg. C. v. Heijningen, RMNH; 5♀, env. La Carolina (Jaén), bridge Rio Guarrizas, 14.vi.1961, leg. Ph. Pronk, J. v. d. Vecht, C. v. Heijningen, RMNH; 1♂, 1♀, Córdoba, Espiel, 19.vi.1961, leg. J. Wiebes & J. v. d. Vecht, RMNH; 1♂, Extremadura, Badajoz, 25.vii.1953, leg. P.M.F. Verhoeff; RMNH; 1♀, Granada, Baños de Zújar, 19.vi.1970, leg. M.C. & G. Kruseman, RMNH; 1♂, Granada, Canales, 22.vii.1978, leg. R. Leys, RMNH; 2♂, 2♀, Granada, Pantano de Cubillas, 27.v-9.vi.1978, leg. R. Leys, RMNH; 1♂, Helechar [Helechal], Badajoz, 20.vi.1961, leg. J. v. d. Vecht, RMNH; 5♀, Jaén, env. of Sta. Elena, 15.vi.1961, leg. C. v. Heijningen, RMNH; 1♂, Málaga, Júzcar, Júzcar to Sendero de la Eras, 29.v.2021, leg. T.J. Wood, TJWC; 1♀, La Aliseda [La Aliseda de Tormes], Ávila, 23.vi.1961, leg. C. v. Heijningen, RMNH; 2♂, Las Correderas (Jaén), 25.v.1958, RMNH; 1♀, Las Correderas (Jaén), 16.vi.1961, leg. J. v. d. Vecht, RMNH; 1♂, Lozoyuela [Lozoyuela-Navas-Sieteiglesias], 10.ix.1952, leg. Bär, Blöte, de Jong, Osse, RMNH; 2♀, Madrid, Chinchón, 6 km N, M-311, 8.vii.2021, leg. T.J. Wood, TJWC; 1♀, Málaga, Benalmádena, 29.v.1962, leg. Jeekel & Wiering, RMNH; 1♂, Málaga, Nerja, 18.ix.1969, leg. C.A.W. & A.M. Jeekel, RMNH; 3♂, Málaga, San Julián 8 km SW of Málaga, 25.v.1962, leg. Jeekel & Wiering, RMNH; 5♂, Málaga, San Pedro de Alcántara, 12–15.v.1960, RMNH; 1♀, Malpartida de Cáceres, 6.vi.1979, leg. H. Teunissen, RMNH; 3♀, Marmolejo, Jaén, 27.v.1979, leg. H. Teunissen, RMNH; 1♂, Martín de Yeltes, Salamanca, 22.vi.1981, leg. H. Teunissen, RMNH; 1♀, Mazagón, Huelva, 22.iv.1985, leg. H. Teunissen, RMNH; 1♀, Medellín, bed of Río Guadiana (Badajoz), 20.vi.1961, leg. J. v. d. Vecht, RMNH; 1♀, Madrid, Pozuelo del Rey, 2 km NW, 10.vii.2021, leg. T.J. Wood, TJWC; 1♂, 2♀, Segovia, Sanchidrián, Esquistos con metarcosas del Precámbrico, 19.vii.2021, leg. T.J. Wood, TJWC; 2♂, 1♀, Sevilla, 29.vii.1950, leg. P.M.F. Verhoeff, RMNH; 3♂, Sevilla, 19.vi.1963, leg. P.M.F. Verhoeff, RMNH; 1♂, Sevilla, Villamanrique [Villamanrique de la Condesa], 8 km SW Pilas, 13.vii.1969, leg. H. Overbeek, RMNH; 1♂, 1♀, Sierra-Morena, El Soldado [Villanueva del Duque], 1.vii.1927, leg. A. Seyrig, MNHN; 1♂, Toledo, 25.vii.1967, leg. P.M.F. Verhoeff, RMNH; 4♂, 1♀, Toledo, 16–18.viii.1967, leg. P.M.F. Verhoeff, RMNH; 2♀, Toledo, 2–3.vii.1968, leg. P.M.F. Verhoeff, RMNH; 1♀, Toledo, 9–10.vi.1968, leg. P.M.F. Verhoeff, RMNH; 2♂, 1♀, Toledo, Alto de Tiendas, 28.v.1995, leg. H. & J.E. Wiering, RMNH; 1♂, Toledo, Talavera de la Reina, 14.ix.1960, leg. H. Wiering, RMNH; 3♂, Villamartín [Cádiz], 30.vii.1950, leg. P.M.F. Verhoeff, RMNH; 2♀, Segovia, Villar del Olmo, 2 km SE, Arroyo de Val, 11.vii.2021, leg. T.J. Wood, TJWC; 1♂, Segovia, Villaseca, Ermita de San Frutos, fields to south and east, 24.vii.2021, leg. T.J. Wood, TJWC; 2♂, 1♀, Huelva, Niebla, 13.viii.1968, leg. K. Warncke, OÖLM;
Diagnosis.Nomiapis paulyi can be recognised within
Nomiapis as being part of the
bispinosa group in the female sex due to the strongly punctate and dull disc of T1 and the weakly and obscurely punctate disc of T3 that strongly contrasts the discs of T1-2 (
Figure 11). In the male sex, it can be recognised by the shape of the hind leg, with a ‘normal’ hind femur (not grossly enlarged as in other
Nomiapis species) which has only very short squamous hairs ventrally (shorter than the width of a flagellum) and the hind tibia is compact, with the apex only produced into a short rounded extension ventrally, not grossly enlarged or forming a long extension (
Figure 12C).
The female can be separated from
Nomiapis bispinosa (alternative character state in parentheses) by the densely punctate scutum with medial punctures separated by 0.5–2 puncture diameters but generally by 1 puncture diameter (scutum irregularly punctate with many large interspaces that are usually greater than 2 puncture diameters medially), and the depression of T2 is punctate basally for more than ½ of its length (depression of T2 punctate basally for at most 1/3rd of its length). Males can be separated from
Nomiapis bispinosa by the smaller body size of 7–9 mm (10–12 mm), the small and weakly produced pair of tubercles on S6 (
Figure 13A; S6 with a pair of strongly produced tubercles,
Figure 13B), and the depression of T3 which is weakly differentiated from the disc, with the depression basally possessing around four distinct rows of punctures that extend into the depression itself (depression of T3 strongly differentiated from the disc, the edge of the disc sharp, almost carina-like, the depression of T3 with one to two rows of punctures at most, these almost indistinct, not extending far onto the depression).
The smaller body size places
Nomiapis paulyi closer to
Nomiapis rufiventris that, to our knowledge, cannot be found in sympatry. Females can be separated by comparison of the tergal punctures.
Nomiapis paulyi has T1 without two clear elongate impunctate areas lateromedially on the disc (T1 with two clear elongate impunctate areas lateromedially on the disc), T2 is more densely punctate, without clear elongate impunctate areas lateromedially on the disc (T2 less densely punctate, with two clear elongate impunctate areas lateromedially on the disc), and the medial scutal punctures are denser, typically separated by 1 puncture diameter (medial scutal punctures sparser, typically separated by more than 1 puncture diameter). At the moment, there is no clear character that allows the separation of males of
Nomiapis paulyi and
Nomiapis rufiventris. There may be small differences in the structure of the hind tibia, with this being apically broader in
Nomiapis paulyi and narrower in
Nomiapis rufiventris. For now, males should be separated on the basis of their non-overlapping range (
Figure 7).
Table 3 provides full comparative details to allow the separation of members of the
bispinosa group.
Description. Female. Body length. 8–9 mm (
Figure 11A).
Head. 1.2 times broader than long (
Figure 11B). Clypeus dark, medially with a weak longitudinal impression, densely punctate, punctures separated by 0.5–1 puncture diameter, interspaces shiny, slightly elevated, often joining together to form interconnected longitudinal wrinkles. The paraocular areas and frons have dense white plumose hairs, obscuring the underlying surface. The gena, vertex, and clypeus have longer, sparse, white to brownish hairs. Antennae dark, A3-12 lightened dark brown below, A3 = A4. The vertex is broad, and the ocelloccipital distance equals 2.5 diameters of the lateral ocellus.
Mesosoma. The scutum is shiny and polished, densely punctate anteriorly and laterally with parapsidal lines and punctures that are almost confluent (
Figure 11C). Slightly less densely punctate medially, with punctures separated by 0.5–2 puncture diameters but typically by 1 puncture diameter. The scutellum is densely punctate, with punctures separated by <0.5 puncture diameters, with interspaces weakly shining. The mesepisternum has a strong network of honeycomb-like raised rugosity, underlying surface weakly shining. Honeycomb-like rugosity weakly extending onto dorsolateral parts of propodeum, lateral and posterior faces of propodeum with dense but shallow punctation, puncture separated by 0.5–1 puncture diameters. The scutum anteriorly and posteriorly has a weak hair fringe of dense whitish-yellow hairs, with the metapostnotum entirely covered in dense whitish-yellow hairs; mesepisternum and lateral faces of propodeum with sparser whitish hairs. The tegulae are large, more or less rectangular, equalling the length of the scutum, apically dark brown, laterally and posteriorly yellowish hyaline. The tegulae are impunctate medially at the base of the wings, densely punctate elsewhere, and the punctures are coarsely medially, becoming finer laterally and posteriorly, with punctures confluent. Legs dark, apical tarsal segments lightened dark reddish-brown; general pubescence whitish. The femoral and tibial scopae are composed of uniformly white hairs. The wings are hyaline, with venation and dark orange stigma, and the nervulus is interstitial.
Metasoma. The T1 with disc densely punctate, with punctures almost confluent medially, becoming slightly sparser laterally, separated by up to 1 puncture diameter. The marginal area of T1 is separated by a narrow, almost impunctate area of 1–2 puncture diameters; the marginal area is densely punctate with 5–6 rows of almost confluent punctures in basal 2/3rds, apical 1/3rd impunctate. The T2 with disc is irregularly punctate with punctures of different sizes, with punctures becoming stronger and clearer laterally (
Figure 11D). The marginal area of T2 is densely punctate over the basal two-thirds with 7–8 rows of small punctures, punctures separated by 0–1 puncture diameters, apical 1/3
rd impunctate, hyaline and semi-translucent. The T3 with disc is weakly and obscurely punctate, punctures separated by 1–3 puncture diameters. Marginal area of T3 densely punctate over basal 1/2 with 7–8 rows of small punctures, punctures separated by 0–1 puncture diameters, apical 1/2 impunctate, hyaline and semi-translucent. The lateral corners of T1 and base of T2 with dense, white, adpressed semi-squamous hairs, and these hairs are also present at the base of T3–4, where they are covered by semi-translucent apical margins of T2–3. Marginal areas of T2–4 with very sparse hairs emerging from the junction of marginal area and disc, forming a very weak fringe. The apical fringe of T5 and hairs flanking the pygidial plate are whitish to dark brown. The pygidial plate apically is broadly rounded.
Male. Body length. 7–9 mm. Head. 1.2 times broader than long. Clypeus dark, medially densely punctate with punctures confluent, punctures becoming sparser apically, here separated by 1 puncture diameter. The frons, paraocular area, supraclypeal area and clypeus are covered with dense whitish-yellowish plumose hairs. Gena and vertex with sparser light brownish hairs. Scape and pedicel dark, flagellar segments bright orange ventrally, dark red dorsally, A3 = 0.8 × A4. The vertex is broad, and the ocelloccipital distance equals 2.5 diameters of the lateral ocellus. Mesosoma. The scutum and scutellum are uniformly densely punctate, with punctures confluent, and the interspaces are dull, lateral corners of the scutellum, each with a small pointed spine. The mesepisternum and propodeum are structurally as in the female. The scutum apically and posteriorly and the metapostnotum has light brown hairs forming a weak fringe, not as strongly produced as in the female. The tegulae are large, more or less rectangular, equalling the length of the scutum, are apically dark brown and laterally and posteriorly yellowish hyaline; entirely densely punctate, with punctures becoming slightly finer laterally and posteriorly. The legs with femorae are dark over the majority of their area and lightened orange apically; the tibiae are predominantly orange with a dark mark medially. The basitarsi are orange with ivory white marks on the outer face, and the remaining tarsal segments are orange, becoming darker apically. The hind femorae are not grossly expanded, apico-ventrally produced into small pointed teeth, ventral face densely covered by white adpressed squamous-flattened hairs. Hind tibiae compact, broad, apico-ventrally produced into a short rounded projection, this projection ivory white. The wings are hyaline, the venation and stigma are dark orange, and the nervulus is interstitial. Metasoma. Tergal discs strongly and densely punctate, punctures separated by 0.5–1 puncture diameters. T1 with marginal area densely punctate over basal ½, with 5 rows of confluent punctures. Marginal areas of T2–4 densely punctate over basal 1/3rd with 3–4 rows of confluent punctures, apical 2/3rds lightened hyaline translucent. T1 on the basal part of the disc with long whitish hairs, T2 basally with dense, white, adpressed semi-squamous hairs forming a complete hair band, these hairs present at the base of T3–5 but covered by semi-translucent marginal areas of T2–4.
Etymology. The name is to commend the Belgian entomologist Alain Pauly for his outstanding, detailed, and far-reaching work on bees of the family Halictidae.
Remarks. This is the taxon reported from Portugal and Spain as
Pseudapis unidentata albocincta by van der Zanden [
37] and
Pseudapis bispinosa albocincta by Wood et al. [
21], but which is not conspecific with North African material due to the average 3.42% genetic distance, as well as the morphological differences described above.
Nomiapis paulyi is clearly polylectic, as expected for this genus (
Table 4). Nearly half of the pollen was collected from Asteraceae, with almost none collected from Fabaceae. This contrasts with
Nomiapis diversipes which is also polylectic but which collected some 35% of its pollen from Fabaceae. More study is required to establish if this is a genuine pattern, particularly at sites where the two taxa occur in sympatry.
Nomiapis paulyi is typically recorded between mid-May and mid-September. Since males can be found in both May and September, the species is clearly at least facultatively bivoltine.
Distribution. Portugal and Spain, though seemingly absent from northern and eastern Iberia (
Figure 7).
Genus Systropha Illiger, 1806
Systropha planidens Giraud, 1861
Systropha planidens Giraud, 1861: 451; ♂♀ Austria [MNHN, lectotype by present designation]
Type material examined.—Lectotype of
S. planidens Giraud (present designation): Autriche//LECTOTYPE//LECTOTYPE—
Systropha planidens Giraud—Le Divelec des. 2022//MNHN, Paris—Ex-coll. Giraud//
Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022//MNHN, Paris—EY33648 (
Figure 14).
–Paralectotypes of S. planidens Giraud: Autriche//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Autriche//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Autriche//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Autriche//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Autriche//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Blue square label//Bergen-staum.//Autriche—Bergenstaum//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Blue square label//Bergen-staum.//Autriche—Bergenstaum//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha planidens Giraud, 1861 ♂—Le Divelec det. 2022.—Illegible square label//green rectangular label//hisp. [Hispania]//Espagne [Hisp.]—Dufour L. leg.//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—Espagne [Hisp.]—Dufour L. leg.//PARALECTOTYPE//PARALECTOTYPE—Systropha planidens Giraud//MNHN, Paris—Ex-coll. Giraud//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.
Other material examined (illustrative, see Table S1). ARMENIA: 1♀, Arzhakan, 24.vii.1963, leg. G. Soika, OÖLM;
BULGARIA: 1♂, 30 km SW Svilengrad, 550 m, 21.vi.2008, leg. M. & Z. Halada, OÖLM;
CZECH REPUBLIC: 2♂, Písek, Pérez Colln., MNHN;
FRANCE: 2♂, Vogezen, Westhalfen [Westhalten], Zinnköpfe, 1.vii.1951, leg. J. v. Assem, RMNH;
GEORGIA: 1♀, E of Tianeti, W Akhmeta, 27.vi.2015, leg. M. Snižek, OÖLM;
GERMANY: 1♂, Würzburg, 1.vii.1876, leg. H. de Vries, RMNH;
GREECE: 6♂, 1♀, Portaria, 3–4.vi.1971, leg. J. v. d. Vecht & P.M.F. Verhoeff, RMNH;
HUNGARY: 1♂, 1♀, Budapest, leg. Mocsáry, RMNH;
IRAN: 1♂, Elburz, Ov Zanak-Ski, 11 km N Ab Ali, 2300 m, 11.vii.1965, leg. G. Soika-Mavromoustakis [G. Soika & G.A. Mavromoustakis], OÖLM;
NORTH MACEDONIA: 1♂, 2♀, Ohrid, 29.v.1972, leg. H. Teunissen, RMNH;
ROMANIA: 1♂, Bucharest, Mihai Bravu, 23.vii.1967, leg. G.P. Kruseman, RMNH;
RUSSIA: 1♀, Sarepta [Volgograd], Vachal Colln., MNHN;
SLOVAKIA: 1♂, Malý Kamenec, 31.vii.1987, leg. Z. Pádr, OÖLM;
TURKEY: 2♂, 1♀, Karakurt/Arastal, 12.vi.1977, leg. K. Warncke, OÖLM;
UKRAINE: 3♂, 2♀, Кирoвoградская oбл. [Kirovogradskaïa], Александрийский р-н. [Oleksandriis’kyi district], Бандурoвка [Bandurivka], 12.vii.1953, leg. А. Осычнык [A. Osytshnjuk], RMNH.
Remarks. Giraud [
3] described
S. planidens from multiple countries; “
Je possède des mâles provenant de l’Espagne … du midi de la France … et quelques uns que j’ai capturés en Autriche. J’en ai deux … dans le Musée de Vienne … d’Amasie [Amasya], dans l’Asie-Mineure”. Baker [
10] concluded that the type series was lost, other than the two specimens from Turkey, which are nominally held at the Vienna Museum. He concluded that there “can be no doubt as to the identity of Giraud’s Austrian syntypes”. Warncke [
9] gave the type locality as ‘Austria’ without locating the type material or designating a lectotype. Given the morphological and genetic evidence supporting the species-specific status of material from Spain and the morphological evidence supporting the specific-specific status of material from Turkey (see below), it is highly likely that Giraud’s original type series was polytypic.
The unpublished catalogue of Giraud (see
https://science.mnhn.fr/catalogue/ey-bib-giraud2/page/224, accessed 12 October 2022) indicates that his collection contains 10 specimens from Austria and Spain. We could indeed find 10 specimens in his boxes, most of them unlabelled, one with the label “Hisp.” for Hispania and two labelled “Bergenstaum,” most probably the name of an Austrian entomologist rather than a locality. As expected, Spanish specimens belong to
S. grandimargo, and Austrian specimens were typical Central European
S. planidens as diagnosed by Warncke. Syntypes from Turkey housed in Vienna were not examined but could belong to either
S. planidens or
S. anatolica (see below for justification of species status). In order to fix the name
S. planidens definitively on the Central European population, an Austrian specimen is designated as a lectotype (
Figure 14), thus preventing the application of this name to Spanish or Turkish syntypic material.
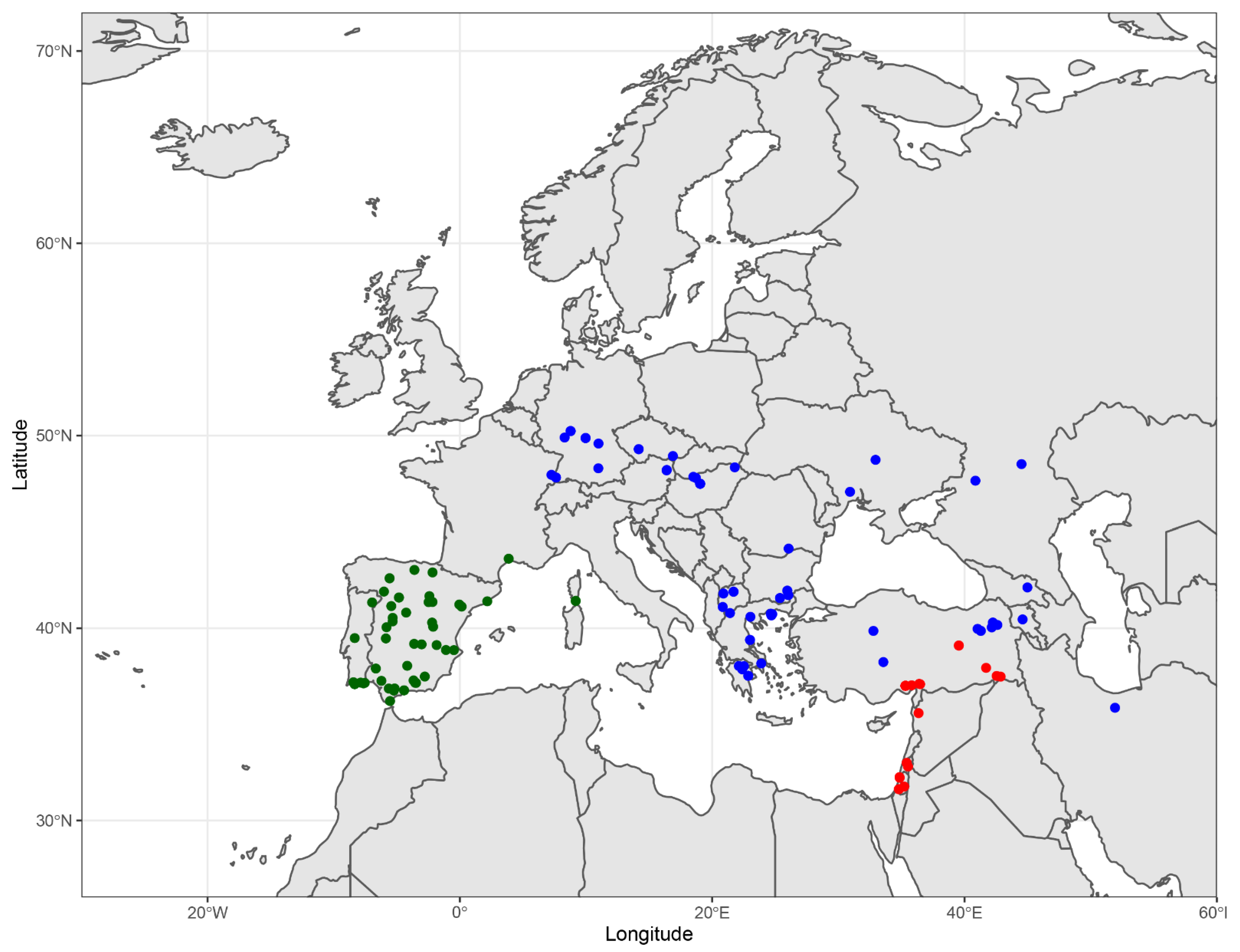
Distribution. France, Germany, Austria, Switzerland, Czech Republic, Poland, Slovakia, Hungary, Romania, Bulgaria, North Macedonia, Greece, Ukraine, Russia, Turkey, Armenia, Georgia, and Iran (see
Table S1,
Figure 15, [
38,
39]). The western limit appears to be the Upper Rhine valley. The eastern limit appears to be the Alborz mountains in northern Iran.
Systropha grandimargo Pérez, 1905 spec. resurr.
Systropha grandimargo Pérez, 1905: 84; ♂♀ Barcelona, Spain [MNHN].
Systropha chrysura Pérez, 1905: 85; ♀ Tarragona, Spain [MNHN] syn. nov.
Type material examined.—Lectotype of
S. grandimargo Pérez: green circle [June]//Barcelone//
grandimargo//MUSEUM PARIS—COLL J.PÉREZ 1915//LECTOTYPE//LECTOTYPE—
Systropha grandimargo J.P.—Warncke [
9] des.//
Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022//MNHN, Paris—EY33641 (
Figure 16).
–Lectotype of
S. chrysura Pérez: 147//
chrysura JP//MUSEUM PARIS—COLL J.PÉREZ 1915//LECTOTYPE//LECTOTYPE—
Systropha chrysura J.P.—Warncke [
9] des.//
Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022//MNHN, Paris—EY33642 (
Figure 17).
–Paralectotypes of S. grandimargo Pérez: Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022.—blue circle [May]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022. green circle [June]//Barcelne [Barcelone]//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022.—green circle [June]//Barcelne [Barcelone]//MUSEUM PARIS—COLL J.PÉREZ 1915//Barcelone—Espagne—R. Desmier de Chenon//Systropha planidens Gir. ♀—Warncke 74 [identification in 1974]—R. Desmier de Chenon//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♀—Le Divelec det. 2022.—green circle [June]//Barcelne [Barcelone]//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//grandimargo//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//La Garriga//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//La Garriga//MUSEUM PARIS—COLL J.PÉREZ 1915//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022.—green circle [June]//Barcelone//MUSEUM PARIS—COLL J.PÉREZ 1915//Barcelone—Espagne—R. Desmier de Chenon planidens//Systropha planidens Gir. ♂—Warncke 74 [identification in 1974]—R. Desmier de Chenon//PARALECTOTYPE//Systropha grandimargo Pérez, 1905 ♂—Le Divelec det. 2022 [S7–8 and genitalia glued on a separate label].
Other material examined (illustrative, see Table S1). FRANCE: Montpellier, 1♂, Pérez collection, MNHN; Corsica, Bonifacio, 1♂, 1♀, Pérez collection, MNHN;
PORTUGAL: Algarve, Cacela Velha, 27.iv.2016, 1♂, leg. Wood, det. Wood, TJWC;
SPAIN: Málaga, Benaoján, Cueva del Hundidero, 3.vi.2021, 1♀, leg. Wood, det. Wood, TJWC.
Remarks. Pérez [
5] described
S. grandimargo from “
divers points de la Catalogne” without specifying an exact
locus typicus. Warncke ([
9]:96) designated a lectotype from a male specimen from Barcelona (
Figure 16) and treated
S. grandimargo as a subspecies of
S. planidens. However, Baker [
10] rejected this lectotype designation arguing that Warncke ‘gave no information identifying any particular specimen’. We disagree with this rejection. The specimen chosen by Warncke is from the Pérez collection, is labelled as being from ‘Barcelone’ with Pérez’s distinctive handwriting and small-style label, and is therefore clearly part of the syntypic series. Thus, the information given by Warncke was sufficient to identify the lectotype in combination with the collection of Pérez, and we accept it as valid.
Pérez [
5] described
S. chrysura from Tarragona from the female sex only. Warncke ([
9]:96) designated a female lectotype (
Figure 17) and synonymised the taxon with
S. planidens. It is, however, conspecific with
S. grandimargo, both morphologically and given that it also comes from the same region of Spain (Catalonia).
In addition to the clear genetic differentiation between Iberian specimens of
S. grandimargo and
S. planidens from Central Europe, the two species can be separated morphologically, mostly clearly by the form of the S8 in the male sex. In
S. grandimargo, S8 is more strongly narrowed medially, the outer margins of the sternal disc are less strongly rounded, and the overall shape is narrower and more rhomboidal (
Figure 18B). In
S. planidens s. str., S8 is broader, the disc resembles a semi-circle, and is not rhombiform (
Figure 18A). The full details are given in the identification key below.
Though Baker [
10] did not follow Warncke [
9] in treating
S. grandimargo as a synonym of
S. planidens (in contrast to
S. chrysura), subsequent authors have followed Warncke’s position (e.g., [
22]). Therefore, we formally treat
S. grandimargo spec. resurr. as a distinct species.
Distribution. Portugal, Spain, France (including Corsica) (
Figure 15, [
36,
38]).
Systropha anatolica Warncke, 1977 stat. nov.
Systropha planidens anatolica Warncke, 1977: 96; ♂♀ Tunceli, Turkey [OÖLM].
Type material examined.—Holotype of S. p. anatolica Warncke: Türkei-Tunceli—13-VI-1973—leg. Kl. Warncke//♂//Systropha planidens ssp. anatolica War—det. Kl. Warncke//Holo-typus//coll. K. Warncke—O. O. Landesmuseum—Linz/Austria-cgg. 93.
–Paratype of S. p. anatolica Warncke: Türkei-Tunceli—13-VI-1973—leg. Kl. Warncke//♀//Systropha planidens ssp. anatolica War—det. Kl. Warncke//Para-type//coll. K. Warncke—O. O. Landesmuseum—Linz/Austria-cgg. 93.
Other material examined (illustrative, see Table S1). ISRAEL: 1♂, 1♀, Tiberias, 18–27.v.1967, leg. C.A.W. Jeekel, RMNH;
SYRIA: 1♂, El Rhab, 52 km NW Hama (Orontes) [Al-Ghab Plain], Syrien, 26.v.1952, leg. E. Schmidt, OÖLM;
TURKEY: 1♂, East Turkey, Fevzipaşa, 25 km W, 24.v.1959, RMNH.
Remarks. Warncke [
9] described
S. anatolica as a subspecies of
S. planidens. Though some authors have followed this viewpoint [
22], Baker [
10] argued that Warncke’s description was inadequate to allow separation from the nominal taxon and that males from Bitlis in Turkey were indistinguishable from males from Greece and Iran. Males of
S. anatolica are readily separable from
S. planidens s. str. through a dissection of the hidden sterna (see
Figure 18, identification key below). In eastern Turkey, both
S. anatolica and
S. planidens can be found, and hence we believe that Baker examined only material of
S. planidens s. str. Though we do not have any genetic data for
S. anatolica, given that the differences in the structure of S8 between
S. anatolica and
S. planidens s. str. are at least as significant as the differences between
S. grandimargo and
S. planidens s. str., we treat it as a valid species.
Identification key for Systropha taxa around S. planidens
1-♀♂: The second submarginal cell is elongated and longer than it is wide (the third abscissa of the basal vein is slightly shorter than the second submarginal crossvein,
Figure 19A). ♀: Terga with double punctation (macro and micro), of which the micropunctation is coarse, dense and unevenly distributed but often aggregating in clumps (particularly visible on T1–2,
Figure 19C); this punctation is remarkably dense toward the disc sides where the integument usually is matte over a large area, almost entirely punctate between the macropunctation. Pubescence and therefore appearance darker, hairs of terminal fringe pure black. ♂: Viewed dorsally, the setae on the dorsal surface of S8 are strongly plumose and dense, almost obscuring the underlying integument (
Figure 16D). Viewed ventrally, the process of S8 (apical part) is broadly rhomboidal, the apico-lateral margins slightly sinuate and conspicuously converging toward the apex; the basal part of the process of S8 (the “neck”) basally constricted, with the lateral margins strongly converging; surface convex, densely and coarsely punctate (
Figure 18B) …
grandimargo Pérez.-♀♂: The second submarginal cell sub-square is wider than it is long (the third abscissa of the basal vein is longer than the second submarginal crossvein,
Figure 19B). ♀: The terga have double punctation, compared to which the micropunctation is shallower, finer, and sparser (particularly visible on T1–2,
Figure 19D); this punctation is slightly denser on a small lateral part of the discs where the tegument remains shiny and predominantly impunctate between macropunctation. The pubescence and appearance are lighter, with hairs of the terminal fringe coloured dark to light brown, not pure black. ♂: Viewed dorsally, the setae on the dorsal surface of S8 are not strongly plumose, comparatively sparse, and do not obscure the underlying integument. Viewed ventrally, the process of S8 is broadly semi-circular, the apico-lateral margins more or less evenly rounded; the basal part of the process of S8 is not strongly constricted, with the lateral margins only weakly converging; the surface is flat, finely and sparsely punctate (
Figure 18A,C) … 2.
2-♀: The pilosity of metasoma is usually darker with brown setae on T4–6. The punctation of the frons is comparatively denser and coarser, with many punctures spaced by less than one puncture diameter medially (
Figure 20A). ♂: The pilosity of metasomal terga is brown, most clearly on T4–6. Viewed ventrally, the process of S8 is hemispherical, with lateral margins that are clearly evenly rounded. S8 medially (from base to neck) has semi-translucent convex lateral margins, forming a pear shape. Laterally projecting basal part of S8 without a projecting tooth (
Figure 18A) …
planidens Giraud.-♀: Pilosity of metasoma light fawn, without dark hairs. Punctation of frons sparser, remarkably sparse dorsomedially where punctures separated by less than one puncture diameter are rare (
Figure 20B). ♂: The pilosity of metasomal terga is whitish, at most slightly yellowish at the base of setae. Viewed ventrally, the process of S8 sub-square, lateral margins long, clearly angulate and contrasting truncate apical margin. S8 medially with semi-translucent convex lateral margins of S8, these forming a strong diamond shape. Laterally projecting basal part of S8 with small protruding tooth (
Figure 18C) …
anatolica Warncke.























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}