
Catalase (CAT) Gene Family in Oil Palm (Elaeis guineensis Jacq.): Genome-Wide Identification, Analysis, and Expression Profile in Response to Abiotic Stress
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of EgCAT Genes
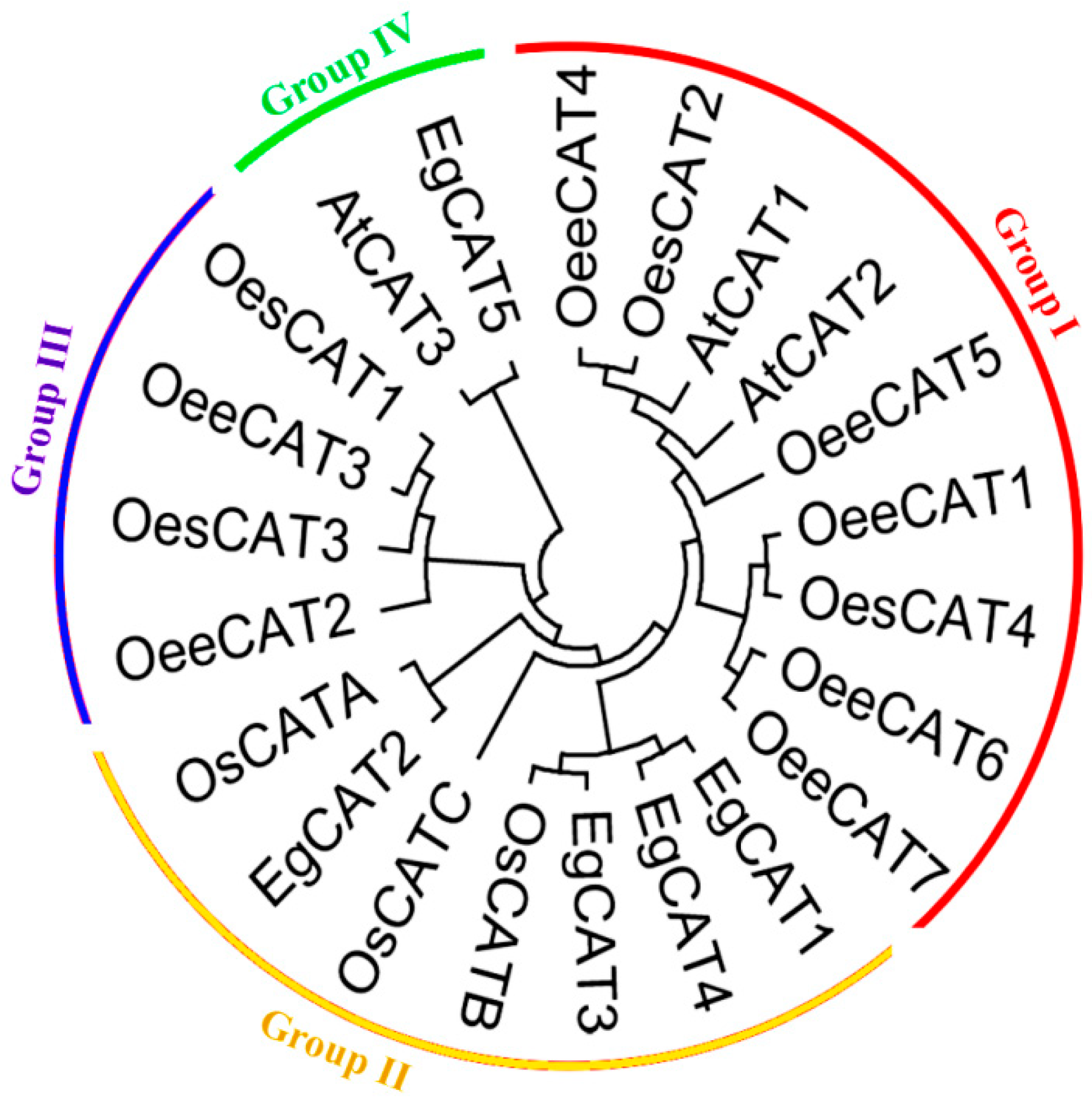
2.2. Phylogenetic Relationship and Structure Analysis of EgCAT Genes
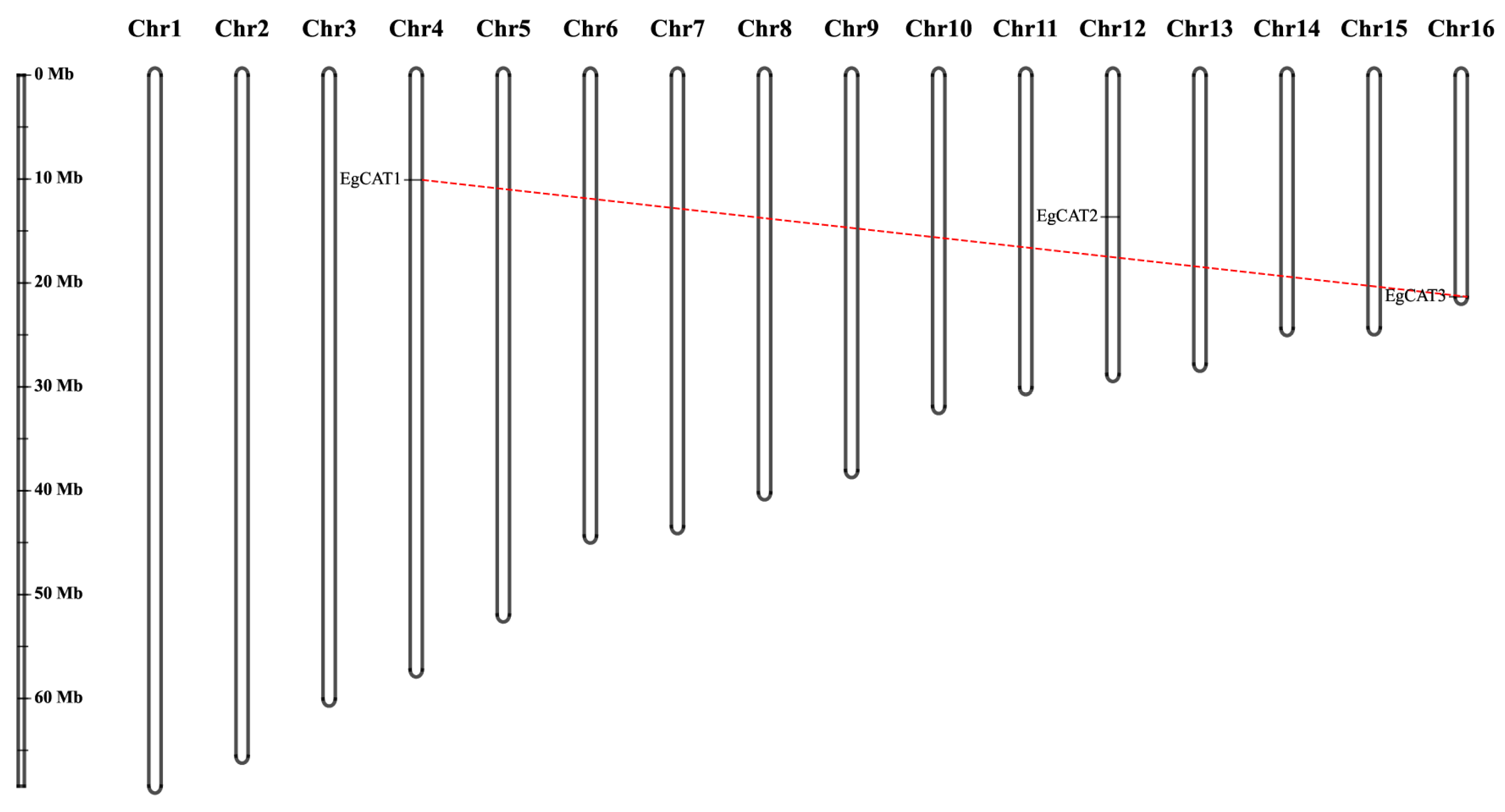
2.3. Chromosomal Distribution of EgCAT Genes
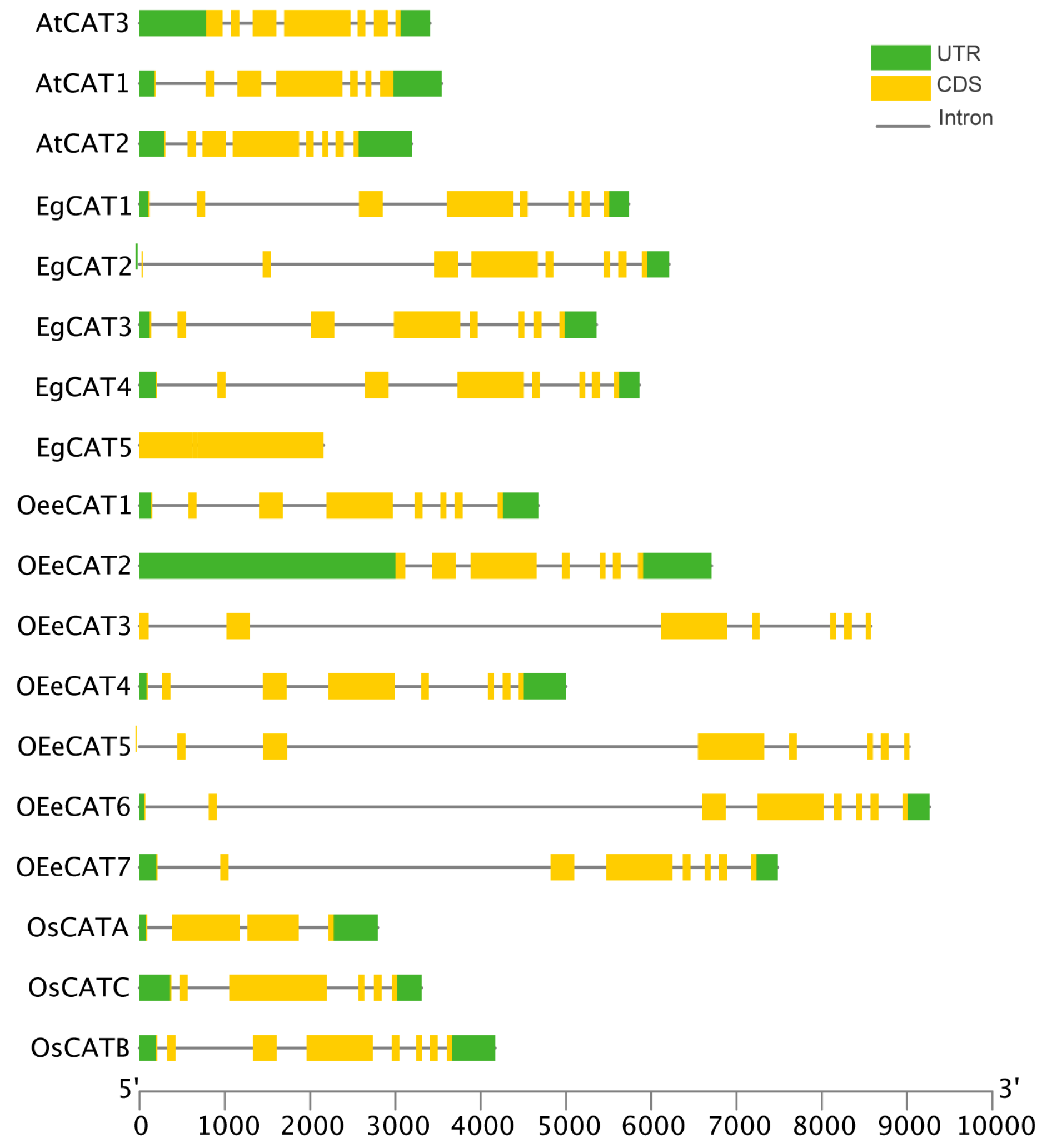
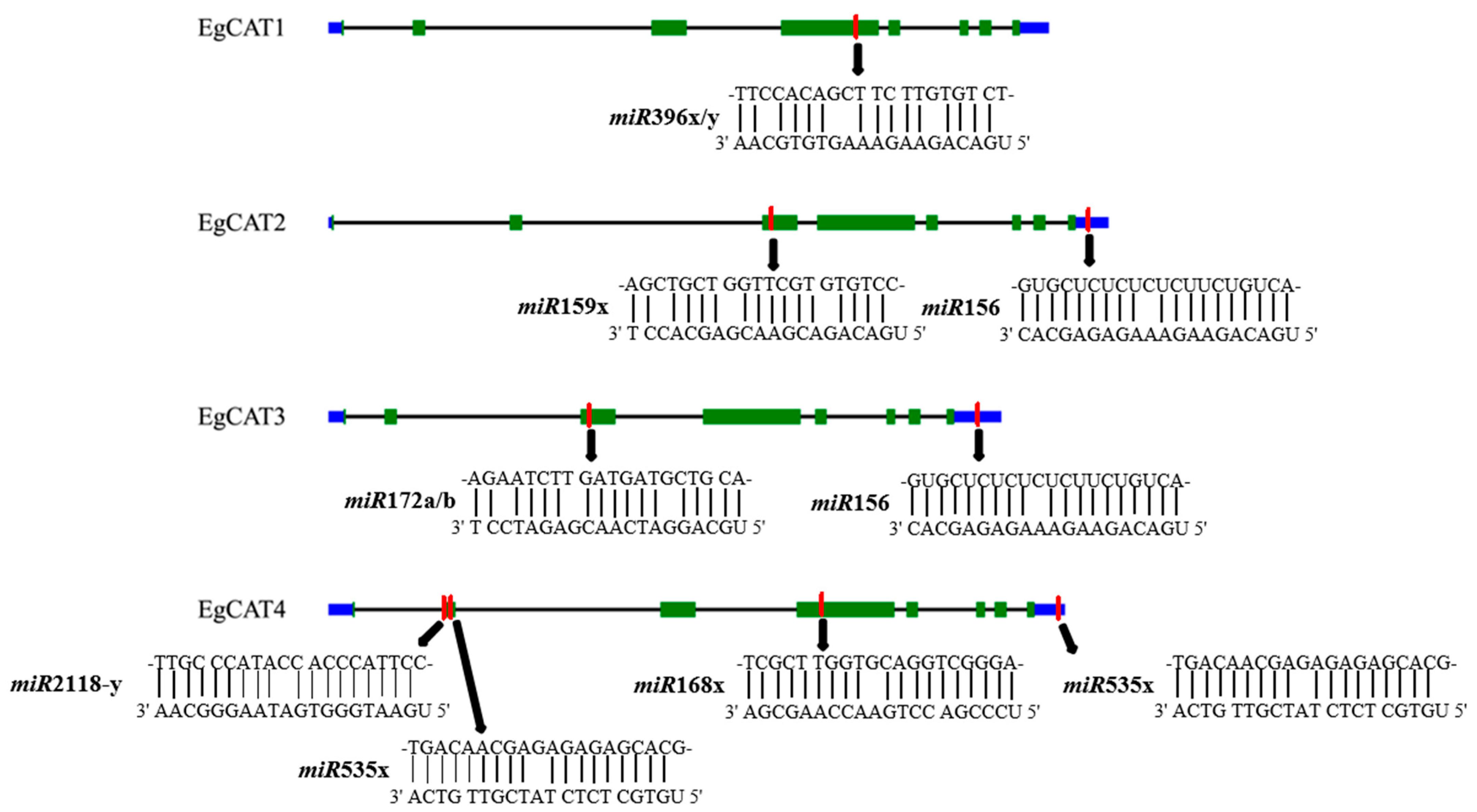
2.4. Gene Structure and Conserved Motif Analysis of EgCAT Genes
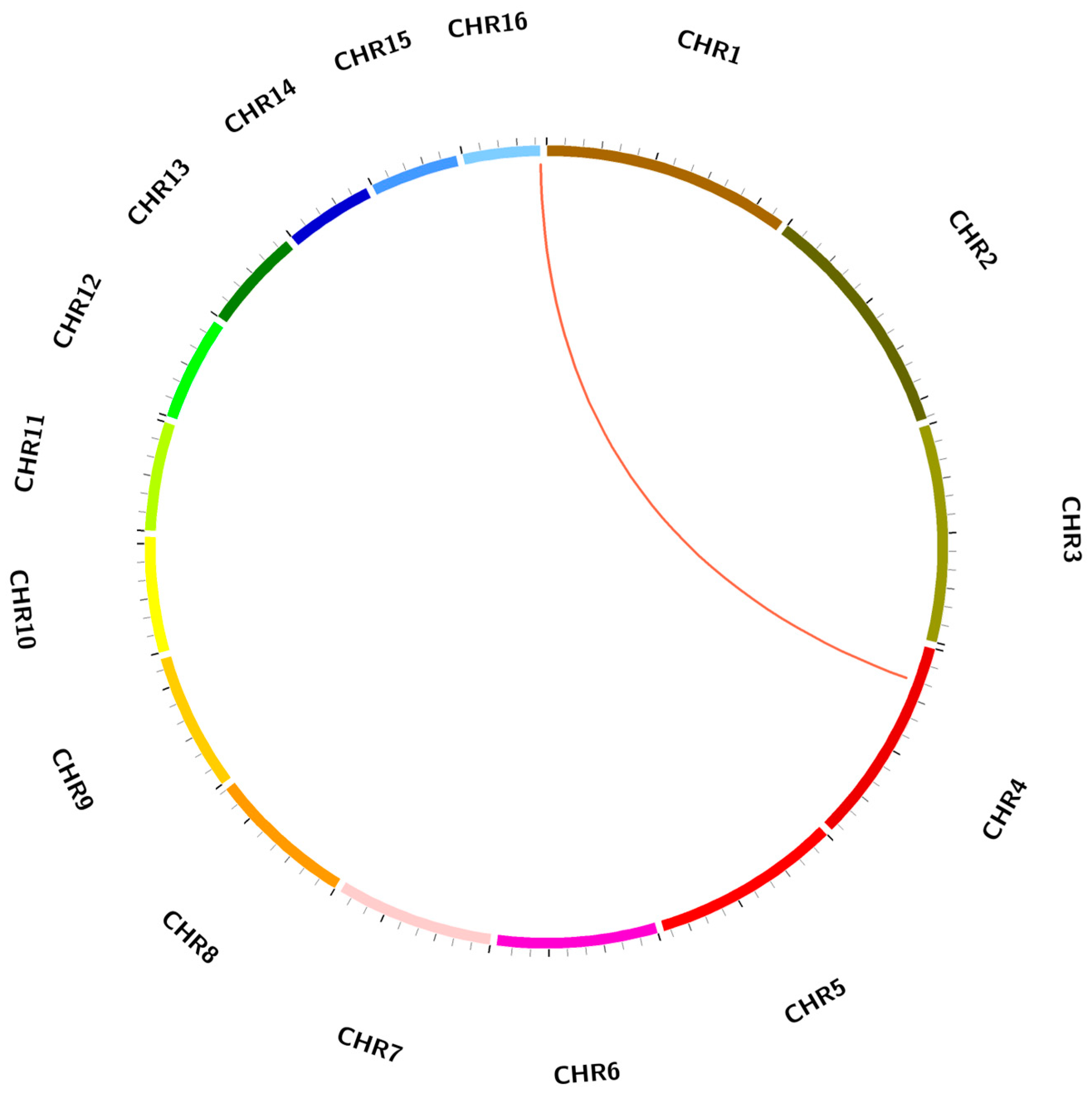
2.5. Duplication Analysis of EgCAT Genes
2.6. Analysis of cis-Acting Elements in EgCAT Gene Promoter Regions
2.7. Prediction of miRNA-Targeting Genes in Oil Palm Genome
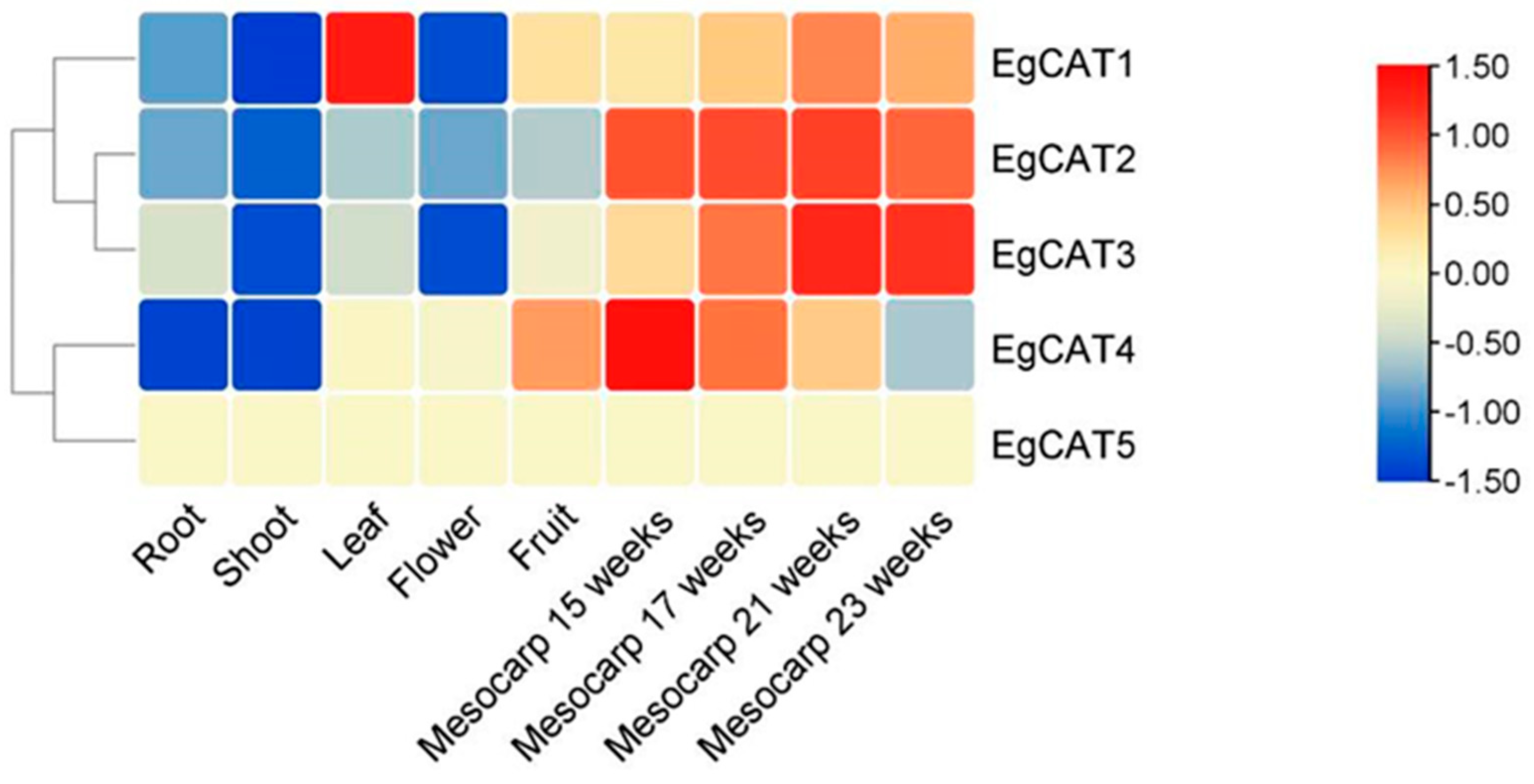
2.8. Expression Profile Analysis of EgCAT Genes
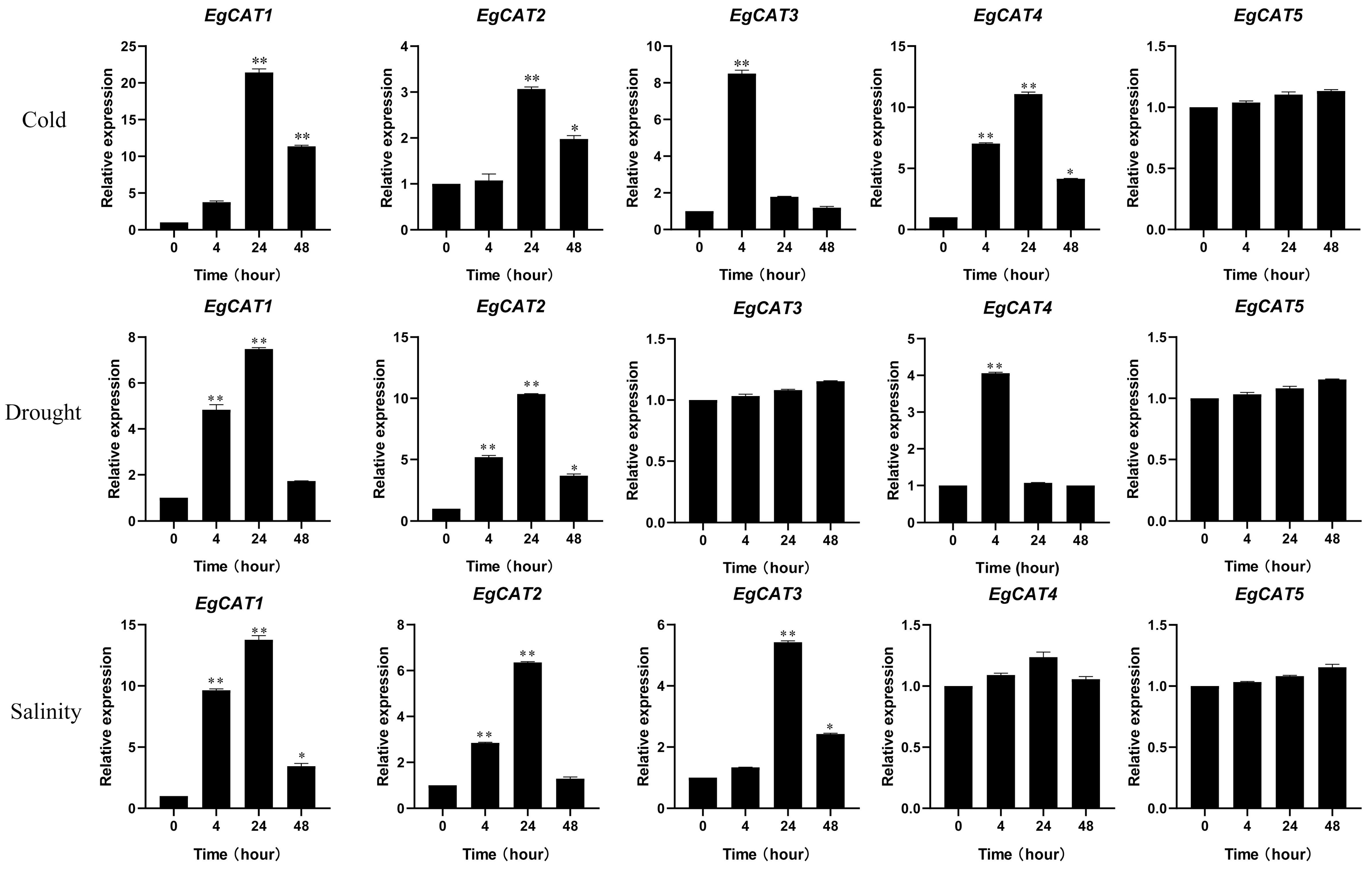
2.9. Expression Analysis of EgCAT Genes of Oil Palm under Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Identification of EgCAT Genes
4.2. EgCAT Gene Structure and Conserved Motif Analysis
4.3. Phylogenetic Relationship, Chromosomal Distribution, and cis-Acting Elements Prediction of EgCAT Genes
4.4. Tissue-Specific Expression of EgCAT Genes Based on Available Transcriptome Datasets
4.5. Plant Material and Abiotic Stress Treatments
4.6. RNA Isolation and Real-Time qPCR Assays
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Soydam-Aydın, S.; Büyük, İ.; Cansaran-Duman, D.; Aras, S. Roles of catalase (CAT) and ascorbate peroxidase (APX) genes in stress response of eggplant (Solanum melongena L.) against Cu(+2) and Zn(+2) heavy metal stresses. Environ. Monit. Assess. 2015, 187, 726. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Sharma, P.; Gill, S.S.; Hasanuzzaman, M.; Khan, E.A.; Kachhap, K.; Mohamed, A.A.; Thangavel, P.; Devi, G.D.; Vasudhevan, P.; et al. Catalase, and ascorbate peroxidase-representative H2O2-detoxifying heme enzymes in plants. Environ. Sci. Pollut. Res. Int. 2016, 23, 19002–19029. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Khan, S.N.; Chatzicharalampous, C.; Bai, D.; Abu-Soud, H.M. Catalase prevents myeloperoxidase self-destruction in response to oxidative stress. J. Inorg. Biochem. 2019, 197, 110706. [Google Scholar] [CrossRef] [PubMed]
- Gondim, F.A.; Gomes-Filho, E.; Costa, J.H.; Mendes Alencar, N.L.; Prisco, J.T. Catalase plays a key role in salt stress acclimation induced by hydrogen peroxide pretreatment in maize. Plant Physiol. Biochem. 2012, 56, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Y.; Jiang, Q.; Zhang, Y.; Ma, L.; Ma, S.; Wang, J.; Ma, Y.; Du, M.; Li, J.; et al. Study on CAT activity of tomato leaf cells under salt stress based on microhyperspectral imaging and transfer learning algorithm. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2023, 302, 123047. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Liu, Y.; Zhang, X.; Liu, X.; Wang, G.; Jing, X.; Wang, X.F.; Zhang, Z.; Hao, G.F.; Zhang, S.; et al. Oxidative post-translational modification of catalase confers salt stress acclimatization by regulating H2O2 homeostasis in Malus hupehensis. J. Plant Physiol. 2023, 287, 154037. [Google Scholar] [CrossRef]
- Komina, A.V.; Korostileva, K.A.; Gyrylova, S.N.; Belonogov, R.N.; Ruksha, T.G. Interaction between single nucleotide polymorphism in catalase gene and catalase activity under the conditions of oxidative stress. Physiol. Res. 2012, 61, 655–658. [Google Scholar] [CrossRef]
- Okabe, H.; Uji, Y. Catalase (CAT). Nihon Rinsho 2004, 11, 554–556. [Google Scholar]
- Sofo, A.; Scopa, A.; Nuzzaci, M.; Vitti, A. Ascorbate Peroxidase and Catalase Activities and Their Genetic Regulation in Plants Subjected to Drought and Salinity Stresses. Int. J. Mol. Sci. 2015, 16, 13561–13578. [Google Scholar] [CrossRef]
- González, E. The C-terminal domain of plant catalases Implications for a glyoxysomal targeting sequence. Eur. J. Biochem. 1991, 199, 211–215. [Google Scholar] [CrossRef]
- Drory, A.; Woodson, W.R. Molecular cloning and nucleotide sequence of a cDNA encoding catalase from tomato. Plant Physiol. 1992, 100, 1605. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.J.; Wu, S.D.; Huang, G.J.; Shen, C.Y.; Afiyanti, M.; Li, W.J.; Lin, Y.H. Expression of a cloned sweet potato catalase SPCAT1 alleviates ethephon-mediated leaf senescence and H2O2 elevation. J. Plant Physiol. 2012, 169, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Willekens, H.; Villarroel, R.; Van Montagu, M.; Inzé, D.; Van Camp, W. Molecular identification of catalases from Nicotiana plumbaginifolia (L.). FEBS Lett. 1994, 352, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Ye, Q.; Wu, Z.; Zhang, Q.; Wang, L.; Liu, J.; Hu, X.; Guo, D.; Wang, X.; Zhang, Z.; et al. Analysis of CAT Gene Family and Functional Identification of OsCAT3 in Rice. Genes 2023, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yang, Y.; Jiang, L.; Liu, S. The catalase gene family in cucumber: Genome-wide identification and organization. Genet. Mol. Biol. 2016, 39, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.S.; Hussain, M.A.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef] [PubMed]
- Yong, B.; Wang, X.Y.; Xu, P.; Zheng, H.Y.; Fei, X.T.; Hong, Z.X.; Ma, Q.Q.; Miao, Y.Z.; Yuan, X.H.; Jiang, Y.S.; et al. Isolation and Abiotic Stress Resistance Analyses of a Catalase Gene from Ipomoea batatas (L.) Lam. BioMed Res. Int. 2017, 2017, 6847532. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.C.; Guo, J.L.; Ling, H.; Chen, S.S.; Wang, S.S.; Xu, L.P.; Allan, A.C.; Que, Y.X. Isolation of a novel peroxisomal catalase gene from sugarcane, which is responsive to biotic and abiotic stresses. PLoS ONE 2014, 9, e84426. [Google Scholar] [CrossRef]
- Mhamdi, A.; Queval, G.; Chaouch, S.; Vanderauwera, S.; Van Breusegem, F.; Noctor, G. Catalase function in plants: A focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 2010, 61, 4197–4220. [Google Scholar] [CrossRef]
- Du, Y.Y.; Wang, P.C.; Chen, J.; Song, C.P. Comprehensive functional analysis of the catalase gene family in Arabidopsis thaliana. J. Integr. Plant Biol. 2008, 50, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Contento, A.L.; Bassham, D.C. Increase in catalase-3 activity as a response to use of alternative catabolic substrates during sucrose starvation. Plant Physiol. Biochem. 2010, 48, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Polidoros, A.; Mylona, P.; Scandalios, J. Transgenic tobacco plants expressing the maize Cat2 gene have altered catalase levels that affect plant-pathogen interactions and resistance to oxidative stress. Trans. Res. 2001, 10, 555–569. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zheng, X.; Diao, Y.; Wang, Y.; Zhou, M.; Hu, Z. Molecular Cloning and Expression Analysis of a Catalase Gene (NnCAT) from Nelumbo nucifera. Appl. Biochem. Biotechnol. 2015, 177, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, L.; Yun, L.; Ji, L.; Li, G.; Ji, M.; Shi, Y.; Zheng, X. Catalase (CAT) Gene Family in Wheat (Triticum aestivum L.): Evolution, Expression Pattern and Function Analysis. Int. J. Mol. Sci. 2022, 23, 542. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, Y.D.; Iqbal, A.; Qadri, R.; Shi, P.; Wang, X.; Wu, Y.; Fan, H.K.; Wu, G.J. Correlation analysis of cold-related gene expression with physiological and biochemical indicators under cold stress in oil palm. PLoS ONE 2019, 14, e0225768. [Google Scholar] [CrossRef]
- Ramli, U.S.; Salas, J.J.; Quant, P.A.; Harwood, J.L. Use of metabolic control analysis to give quantitative information on control of lipid biosynthesis in the important oil crop, Elaeis guineensis (oilpalm). New Phytol. 2009, 184, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.A.; Ahmad Farid, M.A.; Shirai, Y.; Ariffin, H.; Othman, M.R.; Samsudin, M.H.; Hasan, M.Y. Oil Palm Biomass Biorefinery for Sustainable Production of Renewable Materials. Biotechnol. J. 2019, 14, e1800394. [Google Scholar] [CrossRef]
- Baker, A.; Lin, C.C.; Lett, C.; Karpinska, B.; Wright, M.H.; Foyer, C.H. Catalase: A critical node in the regulation of cell fate. Free Radic. Biol. Med. 2023, 199, 56–66. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Satheesh, N.; Kherawat, B.S.; Kumar, A.; Kim, H.U.; Chung, S.M.; Kumar, M. Regulation of Reactive Oxygen Species during Salt Stress in Plants and Their Crosstalk with Other Signaling Molecules-Current Perspectives and Future Directions. Plants 2023, 12, 864. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, Y.; Chen, D.; Liu, D.; Hu, M.; Dong, J.; Zhang, X.; Song, L.; Shen, F. The Catalase Gene Family in Cotton: Genome-Wide Characterization and Bioinformatics Analysis. Cells 2019, 8, 86. [Google Scholar] [CrossRef] [PubMed]
- McClung, C. Regulation of Catalases in Arabidopsis. Free Radic. Biol. Med. 1997, 23, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.; Lee, Y.H.; Song, S.I. Rice CatA, CatB, and CatC are involved in environmental stress response, root growth, and photorespiration, respectively. J. Plant Biol. 2014, 57, 375–382. [Google Scholar] [CrossRef]
- Sytykiewicz, H. Transcriptional responses of catalase genes in maize seedlings exposed to cereal aphids’ herbivory. Biochem. Syst. Ecol. 2015, 60, 131–142. [Google Scholar] [CrossRef]
- Kendall, A.C.; Keys, A.J.; Turner, J.C.; Lea, P.J.; Miflin, B.J. The isolation and characterisation of a catalase-deficient mutant of barley (Hordeum vulgare L.). Planta 1983, 159, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Boeva, V. Analysis of genomic sequence motifs for deciphering transcription factor binding and transcriptional regulation in eukaryotic cells. Front. Genet. 2016, 7, 24. [Google Scholar] [CrossRef] [PubMed]
- Faraji, S.; Filiz, E.; Kazemitabar, S.K.; Vannozzi, A.; Palumbo, F.; Barcaccia, G.; Heidari, P. The AP2/ERF Gene Family in Triticum durum: Genome-wide identification and expression analysis under drought and salinity stresses. Genes 2020, 11, 1464. [Google Scholar] [CrossRef]
- Ghorbel, M.; Zribi, I.; Besbes, M.; Bouali, N.; Brini, F. Catalase Gene Family in Durum Wheat: Genome-Wide Analysis and Expression Profiling in Response to Multiple Abiotic Stress Conditions. Plants 2023, 12, 2720. [Google Scholar] [CrossRef]
- Yadav, A.; Sanyal, I.; Rai, S.P.; Lata, C. An overview on miRNA-encoded peptides in plant biology research. Genomics 2021, 113, 2385–2391. [Google Scholar] [CrossRef]
- Ajila, V.; Colley, L.; Ste-Croix, D.T.; Nissan, N.; Golshani, A.; Cober, E.R.; Mimee, B.; Samanfar, B.; Green, J.R. P-TarPmiR accurately predicts plant-specific miRNA targets. Sci. Rep. 2023, 13, 332. [Google Scholar] [CrossRef]
- Sabzehzari, M.; Naghavi, M.R. Phyto-miRNA: A molecule with beneficial abilities for plant biotechnology. Gene 2019, 683, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Jain, D.; Pandey, J.; Yadav, M.; Bansal, K.C.; Singh, I.K. Deciphering the role of miRNA in reprogramming plant responses to drought stress. Crit. Rev. Biotechnol. 2023, 43, 613–627. [Google Scholar] [CrossRef]
- Sarcheshmeh, M.K.; Abedi, A.; Aalami, A. Genome-wide survey of catalase genes in Brassica rapa, Brassica oleracea, and Brassica napus: Identification, characterization, molecular evolution, and expression profiling of BnCATs in response to salt and cadmium stress. Protoplasma 2023, 260, 899–917. [Google Scholar] [CrossRef]
- Verma, D.; Kaushal, N.; Balhara, R.; Singh, K. Genome-wide analysis of Catalase gene family reveal insights into abiotic stress response mechanism in Brassica juncea and B. Rapa. Plant Sci. 2023, 330, 111620. [Google Scholar] [CrossRef]
- Gupta, S.; Dong, Y.; Dijkwel, P.P.; Mueller-Roeber, B.; Gechev, T.S. Genome-Wide Analysis of ROS Antioxidant Genes in Resurrection Species Suggest an Involvement of Distinct ROS Detoxification Systems during Desiccation. Int. J. Mol. Sci. 2019, 20, 3101. [Google Scholar] [CrossRef]
- Wang, L.; Hou, Y.; Wang, Y.; Hu, S.; Zheng, Y.; Jin, P. Genome-wide identification of heat shock transcription factors and potential role in regulation of antioxidant response under hot water and glycine betaine treatments in cold-stored peaches. J. Sci. Food Agric. 2022, 102, 628–643. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; John Martin, J.J.; Li, R.; Zeng, X.; Wu, Q.; Li, Q.; Fu, D.; Li, X.; Liu, X.; Ye, J.; et al. Catalase (CAT) Gene Family in Oil Palm (Elaeis guineensis Jacq.): Genome-Wide Identification, Analysis, and Expression Profile in Response to Abiotic Stress. Int. J. Mol. Sci. 2024, 25, 1480. https://doi.org/10.3390/ijms25031480
Zhou L, John Martin JJ, Li R, Zeng X, Wu Q, Li Q, Fu D, Li X, Liu X, Ye J, et al. Catalase (CAT) Gene Family in Oil Palm (Elaeis guineensis Jacq.): Genome-Wide Identification, Analysis, and Expression Profile in Response to Abiotic Stress. International Journal of Molecular Sciences. 2024; 25(3):1480. https://doi.org/10.3390/ijms25031480
Chicago/Turabian StyleZhou, Lixia, Jerome Jeyakumar John Martin, Rui Li, Xianhai Zeng, Qiufei Wu, Qihong Li, Dengqiang Fu, Xinyu Li, Xiaoyu Liu, Jianqiu Ye, and et al. 2024. "Catalase (CAT) Gene Family in Oil Palm (Elaeis guineensis Jacq.): Genome-Wide Identification, Analysis, and Expression Profile in Response to Abiotic Stress" International Journal of Molecular Sciences 25, no. 3: 1480. https://doi.org/10.3390/ijms25031480