
Characterization of the Moroccan Barley Germplasm Preserved in the Polish Genebank as a First Step towards Selecting Forms with Increased Drought Tolerance
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
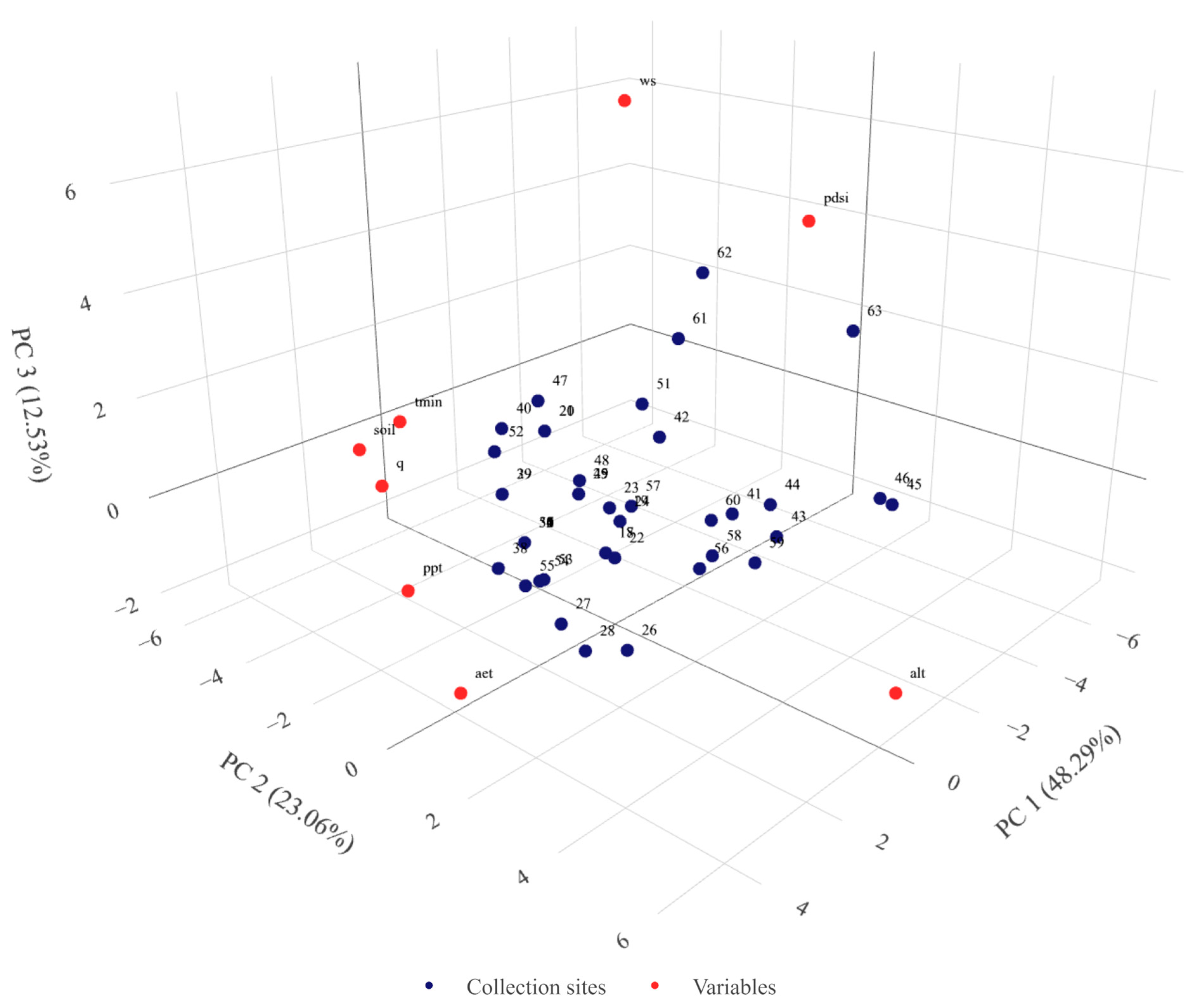
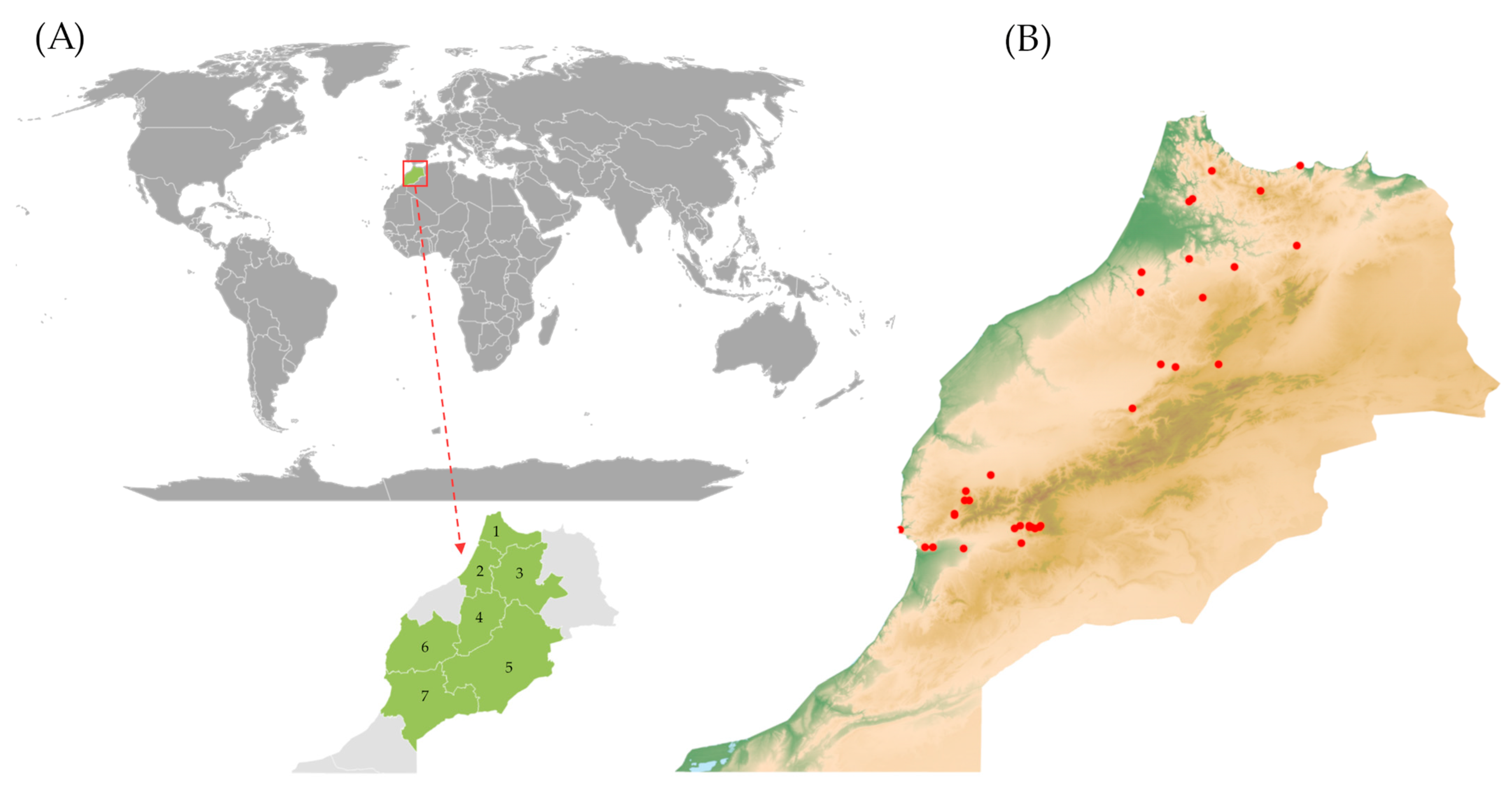
2.1. Eco-Geographical Data Summary
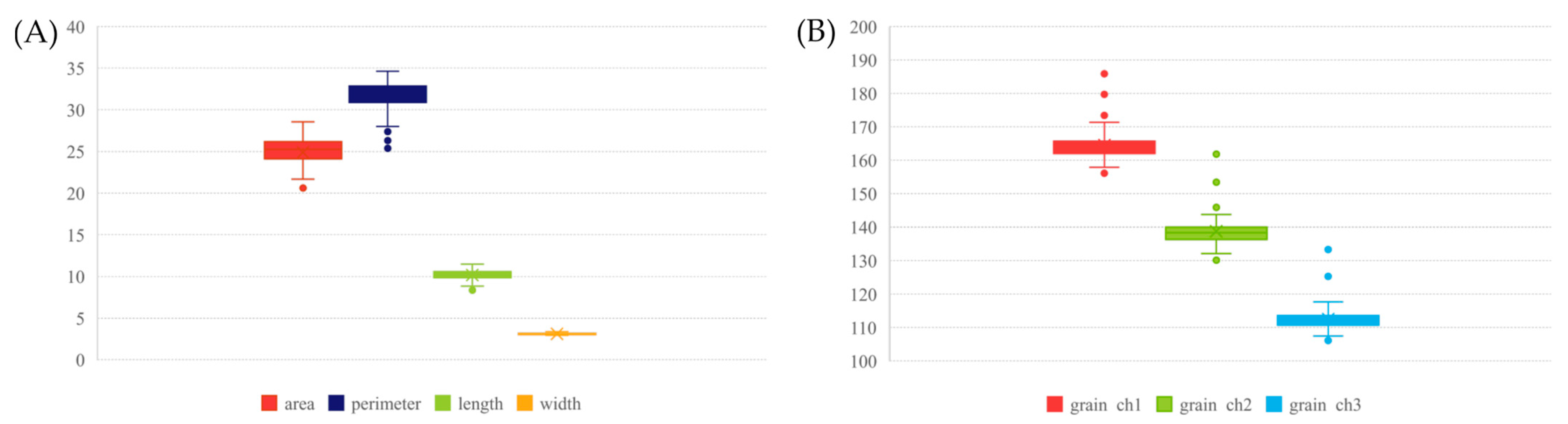
2.2. Grain Morphometry
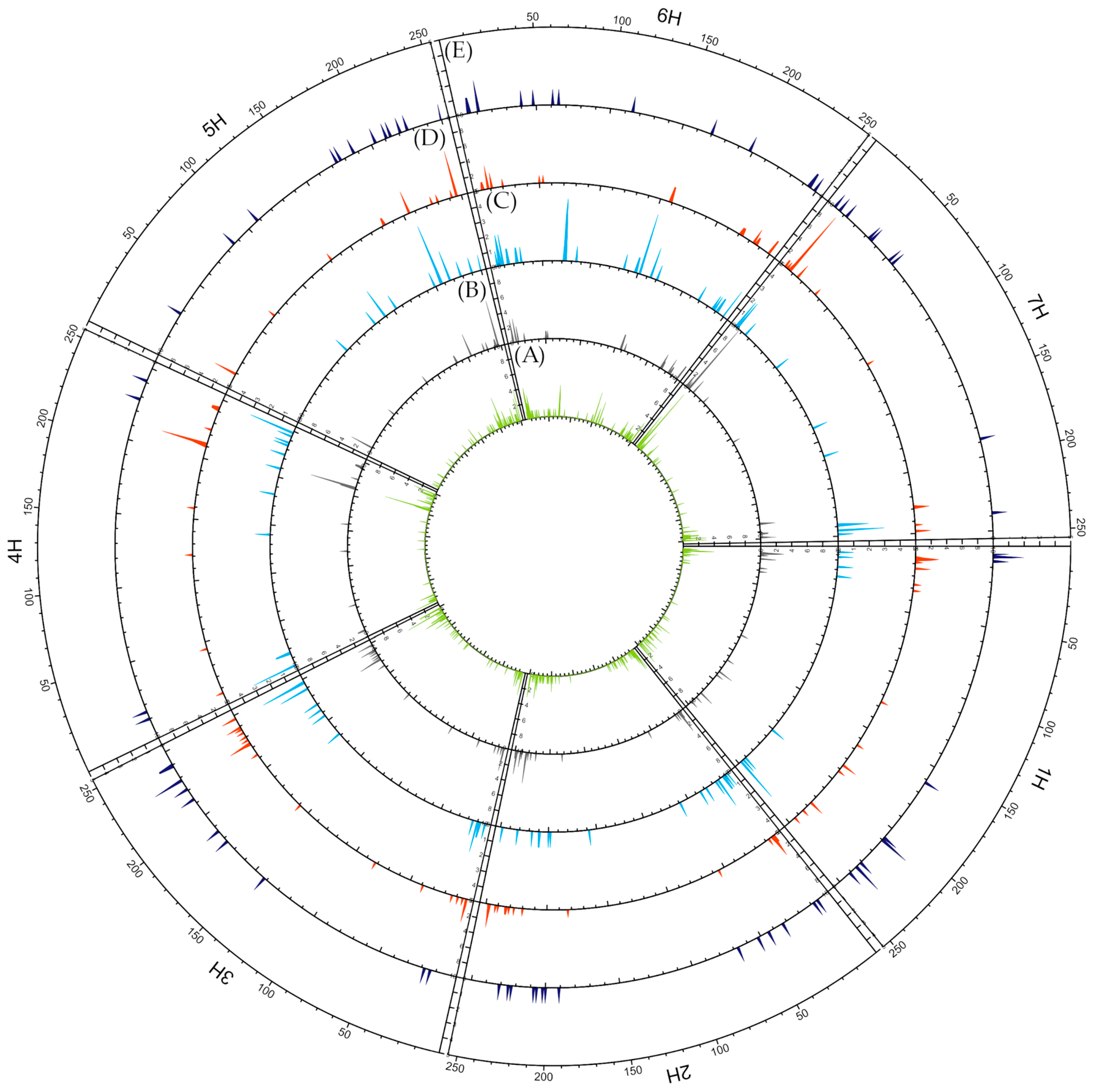
2.3. Genetic Analysis
2.3.1. Data Quality
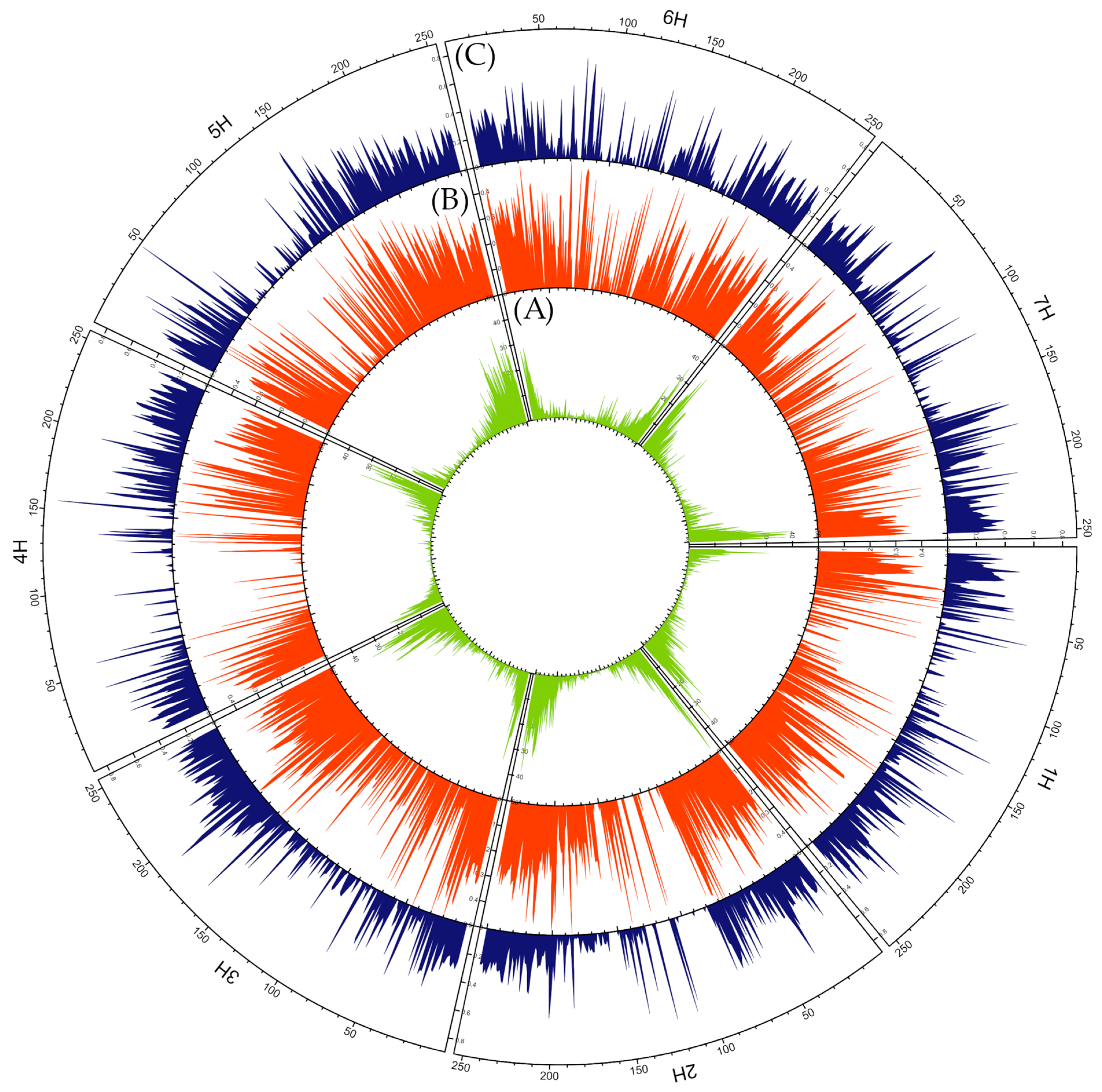
2.3.2. Mutation Types
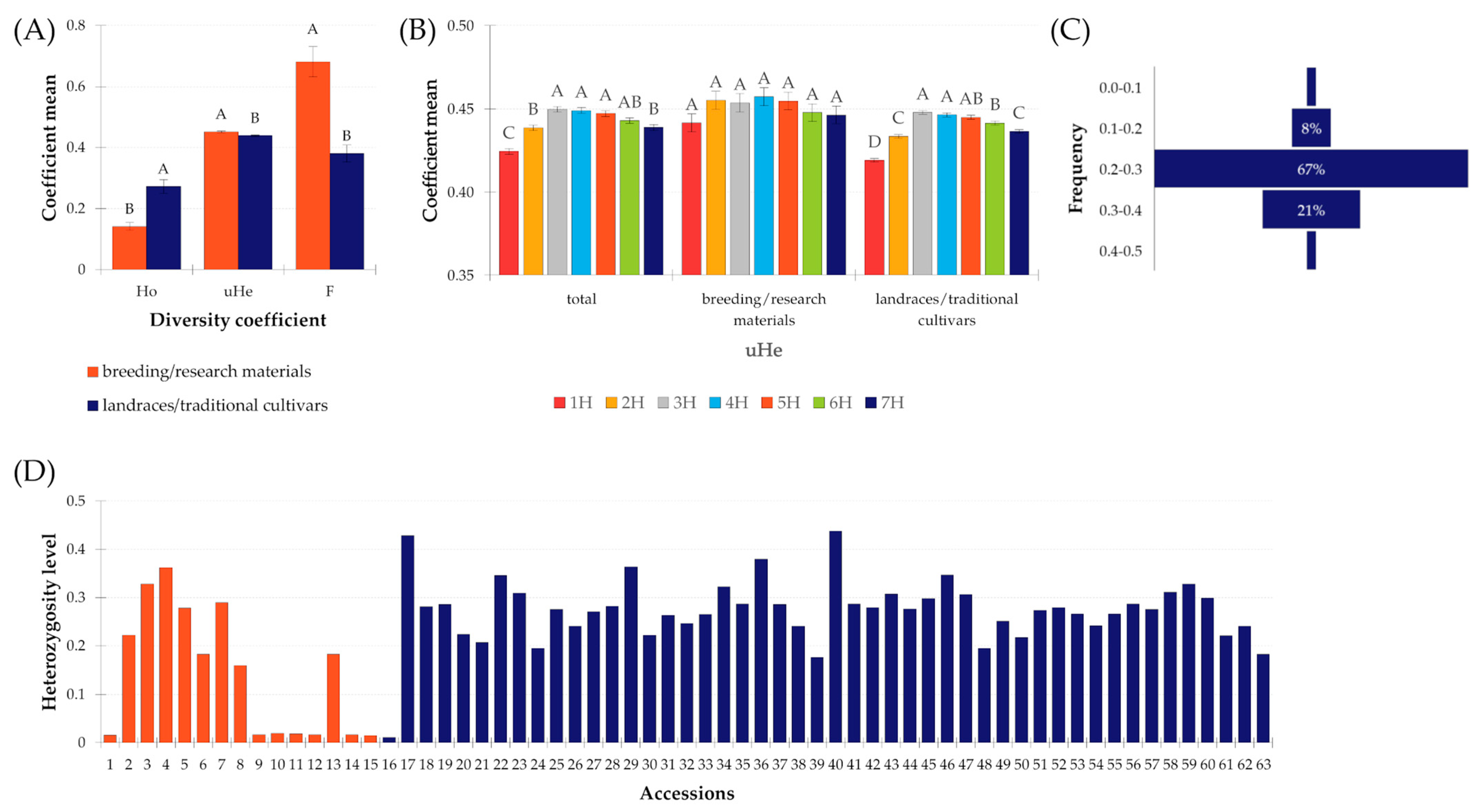
2.3.3. Diversity
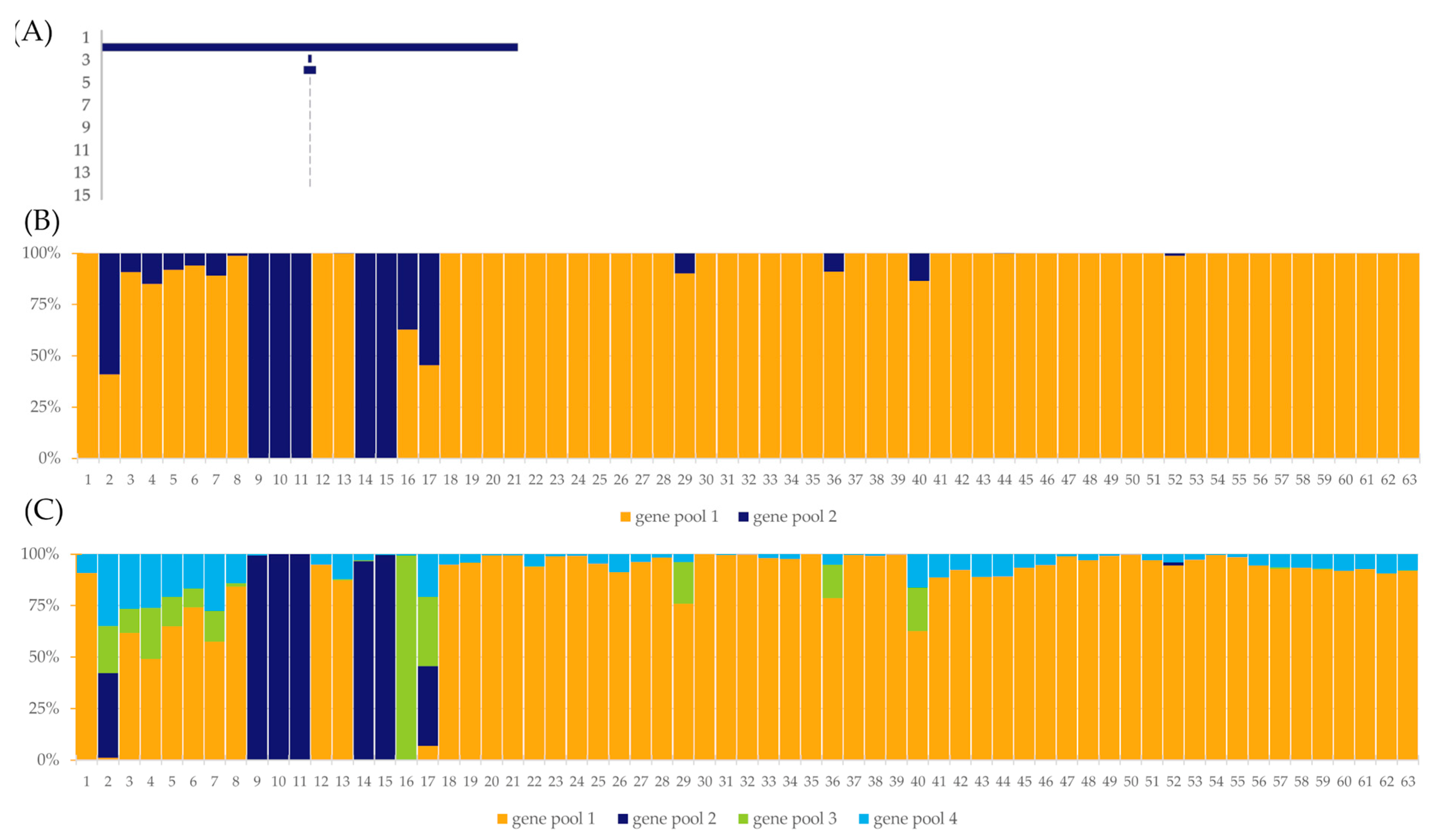
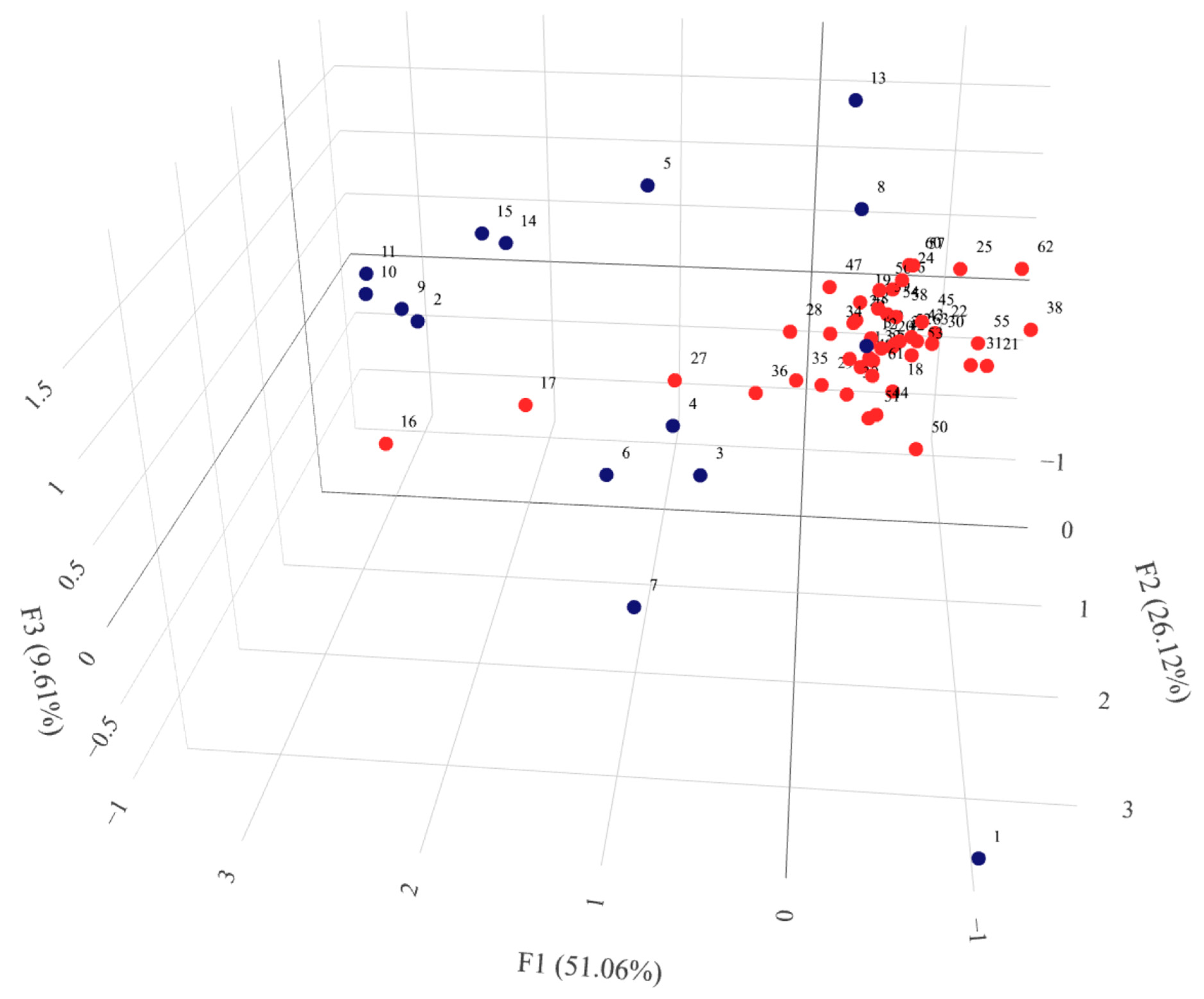
2.3.4. Population Structure
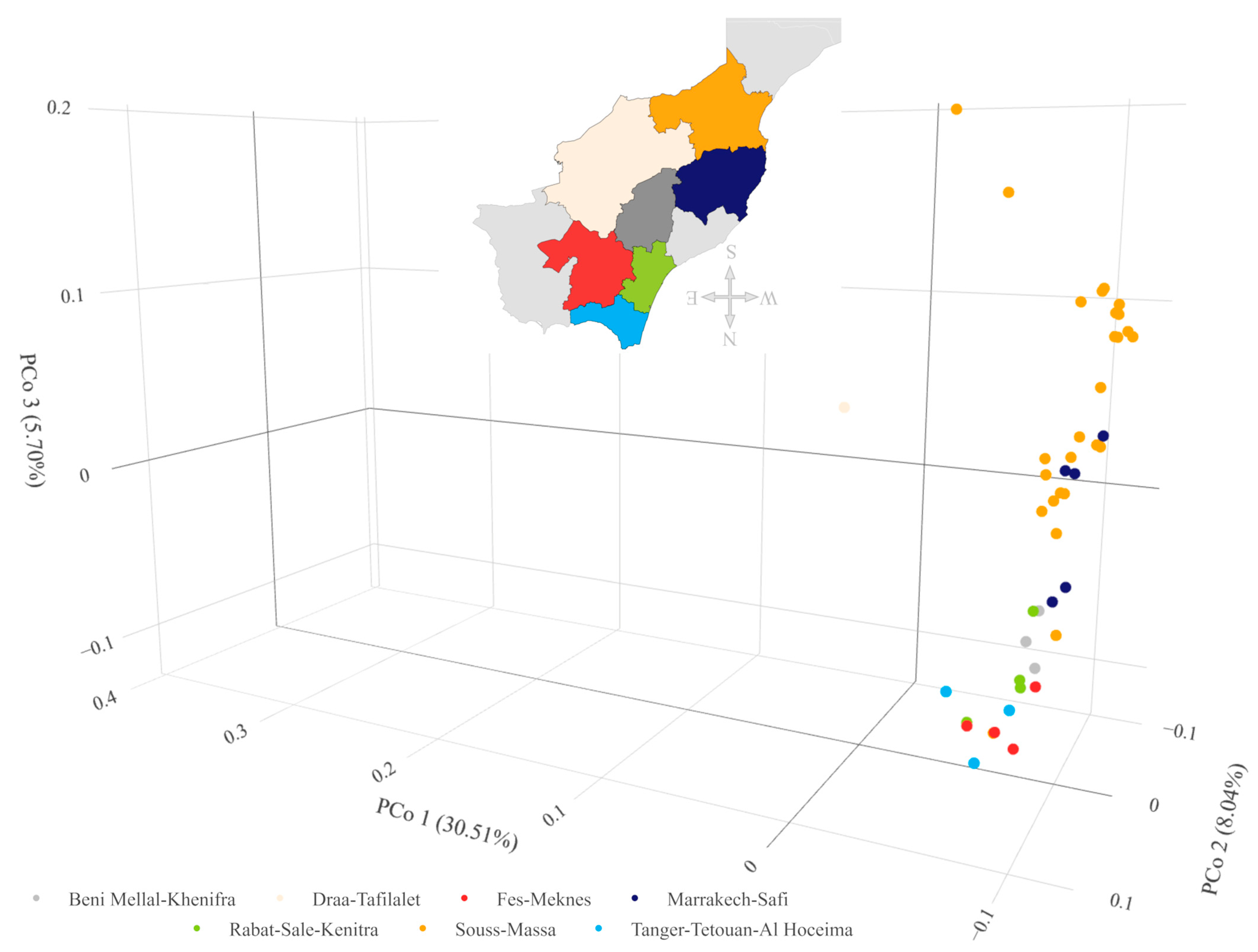
2.4. Association of Variability with Environment
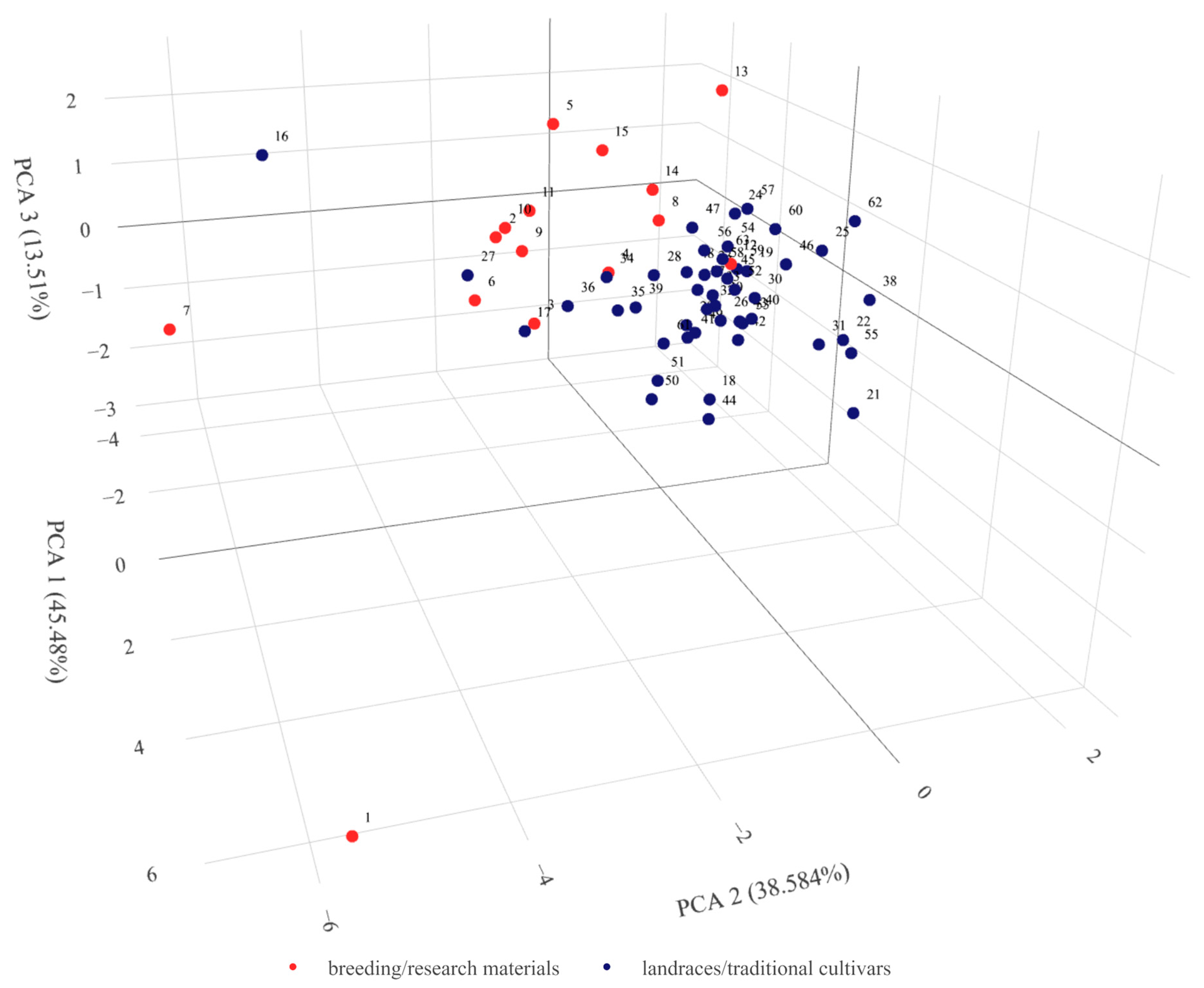
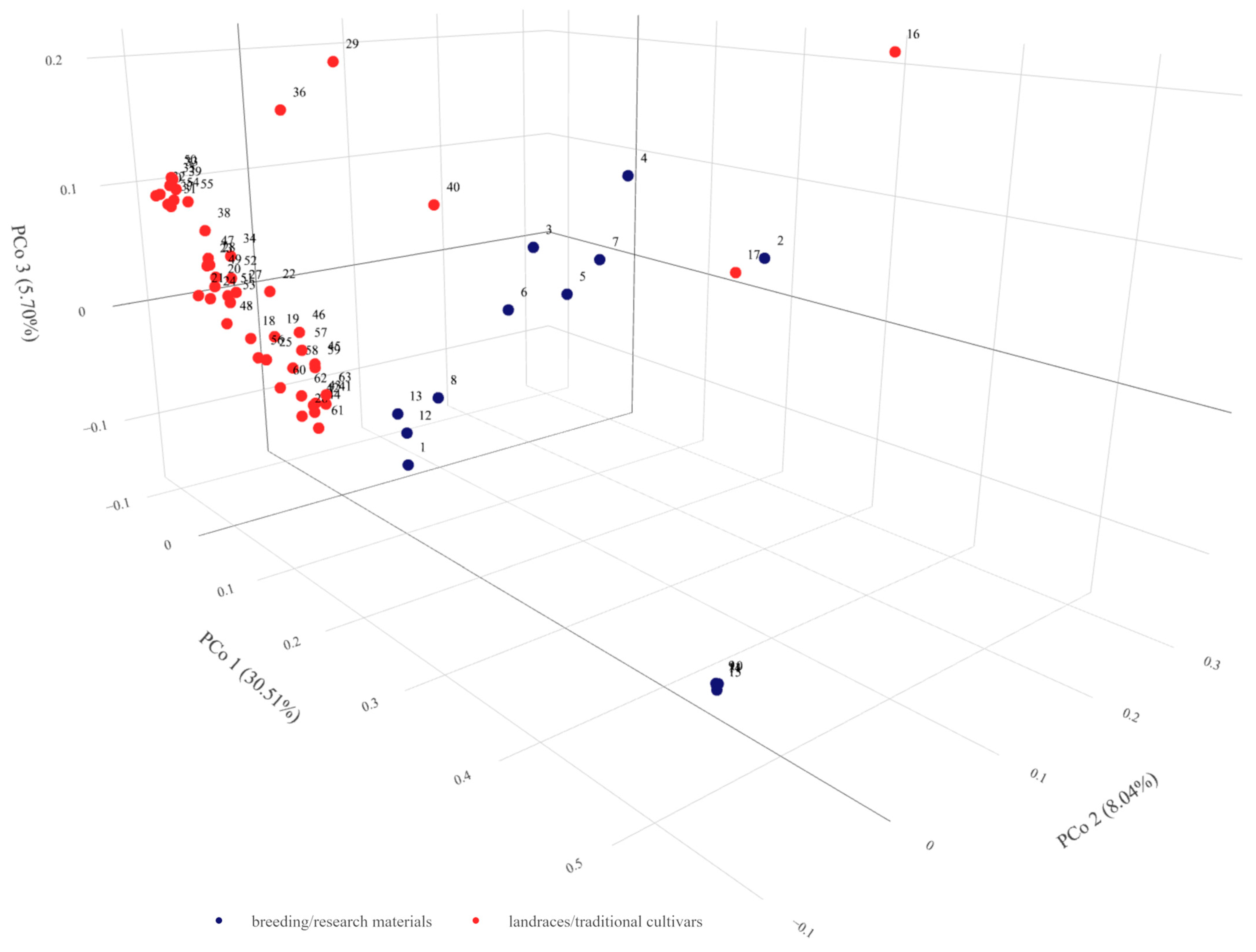
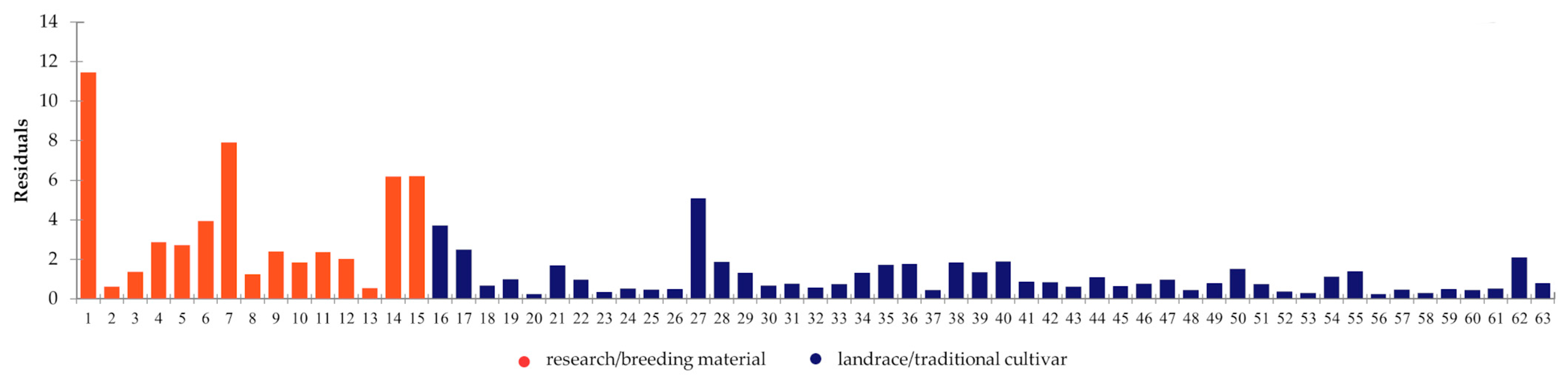
2.5. Combined and Comparative Evaluation
3. Discussion
3.1. Moroccan Accessions Diversity
3.2. Genetic Structure
3.3. Water Deficit Adaptation and Its Potential for Breeding
4. Materials and Methods
4.1. Plant Material
4.2. DNA Isolation
4.3. DArTseq Genotyping
4.4. Grain Morphometry
4.5. Eco-Geographical Data
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Myers, N. Threatened biotas: “Hot spots” in tropical forests. Environmentalist 1988, 8, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Sloan, S.; Jenkins, C.N.; Joppa, L.N.; Gaveau, D.L.; Laurance, W.F. Remaining natural vegetation in the global biodiversity hotspots. Biol. Conserv. 2014, 177, 12–24. [Google Scholar] [CrossRef]
- Noss, R.F.; Platt, W.J.; Sorrie, B.A.; Weakley, A.S.; Means, D.B.; Costanza, J.; Peet, R.K. How global biodiversity hotspots may go unrecognized: Lessons from the North American Coastal Plain. Divers. Distrib. 2015, 21, 236–244. [Google Scholar] [CrossRef]
- Habel, J.C.; Rasche, L.; Schneider, U.A.; Engler, J.O.; Schmid, E.; Rödder, D.; Meyer, S.T.; Trapp, N.; Sos del Diego, R.; Eggermont, H. Final countdown for biodiversity hotspots. Conserv. Lett. 2019, 12, e12668. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Rankou, H.; Culham, A.; Sghir Taleb, M.; Ouhammou, A.; Martin, G.; Jury, S.L. Conservation assessments and Red Listing of the endemic Moroccan flora (monocotyledons). Bot. J. Linn. Soc. 2015, 177, 504–575. [Google Scholar] [CrossRef]
- Valdés, B. Catalogue des Plantes Vasculaires du Nord du Maroc, Incluant des clés D’Identification; CSIC: Madrid, Spain, 2002; Volume 1. [Google Scholar]
- Rankou, H.; Culham, A.; Jury, S.L.; Christenhusz, M.J. The endemic flora of Morocco. Phytotaxa 2013, 78, 1–69. [Google Scholar] [CrossRef]
- Di Castri, F.; Mooney, H.A. Mediterranean Type Ecosystems: Origin and Structure; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 7. [Google Scholar]
- Médail, F.; Myers, N. Mediterranean basin. In Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; CEMEX: Mexico City, Mexico, 2004; pp. 144–147. [Google Scholar]
- Valdés, B. Andalucia and the Rif. Floristic links and a common flora. Bot. Chron 1991, 10, 117–124. [Google Scholar]
- Rodríguez-Sánchez, F.; Pérez-Barrales, R.; Ojeda, F.; Vargas, P.; Arroyo, J. The Strait of Gibraltar as a melting pot for plant biodiversity. Quat. Sci. Rev. 2008, 27, 2100–2117. [Google Scholar] [CrossRef]
- Galán de Mera, A.; Vicente Orellana, J. Phytogeographical characterization of SW Cádiz (Spain) and its relationship with the Tingitanean Peninsula (Morocco). Lagascalia 1997, 19, 609–616. [Google Scholar]
- Marañón, T.; Ajbilou, R.; Ojeda, F.; Arroyo, J. Biodiversity of woody species in oak woodlands of southern Spain and northern Morocco. For. Ecol. Manag. 1999, 115, 147–156. [Google Scholar] [CrossRef]
- Aoubouazza, M.; Rajel, R.; Essafi, R. Impact of extreme climate events on water resources and agriculture and biodiversity in Morocco. J. Climatol. Weather Forecast. 2013, 1, 2. [Google Scholar] [CrossRef]
- Meliho, M.; Khattabi, A.; Jobbins, G.; Sghir, F. Impact of meteorological drought on agriculture in the Tensift watershed of Morocco. J. Water Clim. Chang. 2020, 11, 1323–1338. [Google Scholar] [CrossRef]
- Ezzine, H.; Bouziane, A.; Ouazar, D. Seasonal comparisons of meteorological and agricultural drought indices in Morocco using open short time-series data. Int. J. Appl. Earth Obs. Geoinf. 2014, 26, 36–48. [Google Scholar] [CrossRef]
- Hmimsa, Y.; Ater, M. Agrodiversity in the traditional agrosystems of the Rif mountains (North of Morocco). Biodiversity 2008, 9, 78–81. [Google Scholar] [CrossRef]
- Agnoletti, M.; Tredici, M.; Santoro, A. Biocultural diversity and landscape patterns in three historical rural areas of Morocco, Cuba and Italy. Biodivers. Conserv. 2015, 24, 3387–3404. [Google Scholar] [CrossRef]
- Savin, R.; Stone, P.J.; Nicolas, M.E. Responses of grain growth and malting quality of barley to short periods of high temperature in field studies using portable chambers. Aust. J. Agric. Res. 1996, 47, 465–477. [Google Scholar] [CrossRef]
- Teshome, A.; Brown, A.; Hodgkin, T. Diversity in landraces of cereal and legume crops. Plant Breed. Rev. 2010, 21, 221–261. [Google Scholar]
- Saidi, S.; Amri, A.; Jarvis, D.; Sbeih, D.; Saad, A. On-farm conservation of barley landraces in Morocco. In Conference Proceedings: African Regional Workshop on Sustainable Use (Focus on Agriculture), Nairobi, Kenya; ICARDA: Aleppo, Syria, 2006. [Google Scholar]
- Jarvis, D.I.; Hodgkin, T.; Sthapit, B.R.; Fadda, C.; Lopez-Noriega, I. An heuristic framework for identifying multiple ways of supporting the conservation and use of traditional crop varieties within the agricultural production system. Crit. Rev. Plant Sci. 2011, 30, 125–176. [Google Scholar] [CrossRef]
- Van Mele, P.; Guéi, R.G. African Seed Enterprises: Sowing the Seeds of Food Security; CABI: Wallingford, UK, 2011. [Google Scholar]
- Jensen, H.R.; Belqadi, L.; De Santis, P.; Sadiki, M.; Jarvis, D.I.; Schoen, D.J. A case study of seed exchange networks and gene flow for barley (Hordeum vulgare subsp. vulgare) in Morocco. Genet. Resour. Crop Evol. 2013, 60, 1119–1138. [Google Scholar] [CrossRef]
- Scholten, M.; Green, N.; Campbell, G.; Maxted, N.; Ford-Lloyd, B.; Ambrose, M.; Spoor, B. Landrace inventory of the UK. In European Landraces: On-farm conservation, Management and Use. Bioversity Technical Bulletin 15; 2009; p. 161. Available online: https://research.birmingham.ac.uk/en/publications/landrace-inventory-of-the-uk (accessed on 10 October 2023).
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Vallage, V.; Hari, S. Domestication of plants in the old World—The origin and spread of cultivated plants in West Asia, Europe, and the Nile Valley. Genet. Resour. Crop. Evol. 1979, 123, 256–275. [Google Scholar]
- Salamini, F.; Özkan, H.; Brandolini, A.; Schäfer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 2002, 3, 429–441. [Google Scholar] [CrossRef]
- Molina-Cano, J.-L.; Russell, J.; Moralejo, M.; Escacena, J.; Arias, G.; Powell, W. Chloroplast DNA microsatellite analysis supports a polyphyletic origin for barley. Theor. Appl. Genet. 2005, 110, 613–619. [Google Scholar] [CrossRef]
- Molina-Cano, J.L.; Moralejo, M.; Igartua, E.; Romagosa, I. Further evidence supporting Morocco as a centre of origin of barley. Theor. Appl. Genet. 1999, 98, 913–918. [Google Scholar] [CrossRef]
- Tanno, K.; Taketa, S.; Takeda, K.; Komatsuda, T. A DNA marker closely linked to the vrs1 locus (row-type gene) indicates multiple origins of six-rowed cultivated barley (Hordeum vulgare L.). Theor. Appl. Genet. 2002, 104, 54–60. [Google Scholar] [CrossRef]
- Clark, H.H. The origin and early history of the cultivated barleys: A botanical and archaeological synthesis. Agric. Hist. Rev. 1967, 15, 1–18. [Google Scholar]
- Kramer, S.N. History Begins at Sumer: Thirty-Nine Firsts in Recorded History; University of Pennsylvania Press: Philadelphia, PA, USA, 1981. [Google Scholar]
- Newman, R.K.; Newman, C.W. Barley for Food and Health: Science, Technology, and Products; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- FAO. Faostat, Statistical Database. Available online: https://www.fao.org/faostat/en/#home (accessed on 10 October 2023).
- Fardaoussi, M. Grain and Feed Update, Morocco. Available online: https://apps.fas.usda.gov/ (accessed on 10 October 2023).
- Koornneef, M.; Alonso-Blanco, C.; Peeters, A. Genetic approaches in plant physiology. New Phytol. 1997, 137, 1–8. [Google Scholar] [CrossRef]
- Baik, B.-K.; Ullrich, S.E. Barley for food: Characteristics, improvement, and renewed interest. J. Cereal Sci. 2008, 48, 233–242. [Google Scholar] [CrossRef]
- Bothmer, R.v.; Jacobsen, N.; Baden, C.; Jorgensen, R.; Linde-Laursen, I. An Ecogeographical Study of the Genus Hordeum; IPGRI: Rome, Italy, 1995. [Google Scholar]
- Badr, A.; Sch, R.; Rabey, H.E.; Effgen, S.; Ibrahim, H.; Pozzi, C.; Rohde, W.; Salamini, F. On the origin and domestication history of barley (Hordeum vulgare). Mol. Biol. Evol. 2000, 17, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Cai, S.; Chen, M.; Ye, L.; Chen, Z.; Zhang, H.; Dai, F.; Wu, F.; Zhang, G. Tissue metabolic responses to salt stress in wild and cultivated barley. PLoS ONE 2013, 8, e55431. [Google Scholar] [CrossRef] [PubMed]
- Forster, B.; Ellis, R.; Moir, J.; Talame, V.; Sanguineti, M.; Tuberosa, R.; This, D.; Teulat-Merah, B.; Ahmed, I.; Mariy, S. Genotype and phenotype associations with drought tolerance in barley tested in North Africa. Ann. Appl. Biol. 2004, 144, 157–168. [Google Scholar] [CrossRef]
- Madidi, S.E.; Diani, Z.; Aameur, F.B. Variation of agro-morphological characters in Moroccan barley landraces under near optimal and drought conditions. Genet. Resour. Crop Evol. 2005, 52, 831–838. [Google Scholar] [CrossRef]
- Oukarroum, A.; El Madidi, S.; Strasser, R.J. Differential heat sensitivity index in barley cultivars (Hordeum vulgare L.) monitored by chlorophyll a fluorescence OKJIP. Plant Physiol. Biochem. 2016, 105, 102–108. [Google Scholar] [CrossRef]
- Czembor, J.H. Sources of powdery mildew resistance in barley landraces from morocco. Arch. Phytopathol. Plant Prot. 2000, 33, 111–130. [Google Scholar] [CrossRef]
- Piechota, U.; Czembor, P.C.; Słowacki, P.; Czembor, J.H. Identifying a novel powdery mildew resistance gene in a barley landrace from Morocco. J. Appl. Genet. 2019, 60, 243–254. [Google Scholar] [CrossRef] [PubMed]
- NCPGR. EGISET. Available online: https://wyszukiwarka.ihar.edu.pl/pl (accessed on 10 October 2023).
- Mascher, M.; Gundlach, H.; Himmelbach, A.; Beier, S.; Twardziok, S.O.; Wicker, T.; Radchuk, V.; Dockter, C.; Hedley, P.E.; Russell, J. A chromosome conformation capture ordered sequence of the barley genome. Nature 2017, 544, 427–433. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-B.; Dong, Y.-B. Genetic erosion under modern plant breeding: Case studies in Canadian crop gene pools. Genet. Divers. Eros. Plants Indic. Prev. 2015, 7, 89–104. [Google Scholar]
- Dziurdziak, J.; Podyma, W.; Bujak, H.; Boczkowska, M. Tracking changes in the spring barley gene pool in Poland during 120 years of breeding. Int. J. Mol. Sci. 2022, 23, 4553. [Google Scholar] [CrossRef] [PubMed]
- Rufo, R.; Alvaro, F.; Royo, C.; Soriano, J.M. From landraces to improved cultivars: Assessment of genetic diversity and population structure of Mediterranean wheat using SNP markers. PLoS ONE 2019, 14, e0219867. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Schulthess, A.W.; Bassi, F.M.; Badaeva, E.D.; Neumann, K.; Graner, A.; Özkan, H.; Werner, P.; Knüpffer, H.; Kilian, B. Introducing beneficial alleles from plant genetic resources into the wheat germplasm. Biology 2021, 10, 982. [Google Scholar] [CrossRef]
- de Oliveira, L.; Martins, E.R. A quantitative assessment of genetic erosion in ipecac (Psychotria ipecacuanha). Genet. Resour. Crop Evol. 2002, 49, 607–617. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Yeken, M.Z.; Tekin, M.; Mustafa, Z.; Hatipoğlu, R.; Aktaş, H.; Alsaleh, A.; Cabi, E.; Habyarimana, E.; Zencirci, N. Contribution of Landraces in Wheat Breeding. In Wheat Landraces; Springer International Publishing: Cham, Switzerland, 2021; pp. 215–258. [Google Scholar] [CrossRef]
- Wendler, N.; Mascher, M.; Nöh, C.; Himmelbach, A.; Scholz, U.; Ruge-Wehling, B.; Stein, N. Unlocking the secondary gene-pool of barley with next-generation sequencing. Plant Biotechnol. J. 2014, 12, 1122–1131. [Google Scholar] [CrossRef]
- Begna, T.; Begna, T. Role and economic importance of crop genetic diversity in food security. Int. J. Agric. Sci. Food Technol. 2021, 7, 164–169. [Google Scholar]
- Degu, H.D.; Tehelku, T.F.; Kalousova, M.; Sato, K. Genetic diversity and population structure of barley landraces from Southern Ethiopia’s Gumer district: Utilization for breeding and conservation. PLoS ONE 2023, 18, e0279737. [Google Scholar] [CrossRef]
- Gadissa, F.; Abebe, M.; Bekele, T. Agro-morphological traits-based genetic diversity assessment in Ethiopian barley (Hordeum vulgare L.) landrace collections from Bale highlands, Southeast Ethiopia. Agric. Food Secur. 2021, 10, 1–14. [Google Scholar] [CrossRef]
- Dziurdziak, J.; Gryziak, G.; Groszyk, J.; Podyma, W.; Boczkowska, M. DArTseq Genotypic and Phenotypic Diversity of Barley Landraces Originating from Different Countries. Agronomy 2021, 11, 2330. [Google Scholar] [CrossRef]
- Almerekova, S.; Genievskaya, Y.; Abugalieva, S.; Sato, K.; Turuspekov, Y. Population structure and genetic diversity of two-rowed barley accessions from Kazakhstan based on SNP genotyping data. Plants 2021, 10, 2025. [Google Scholar] [CrossRef]
- Jayakodi, M.; Padmarasu, S.; Haberer, G.; Bonthala, V.S.; Gundlach, H.; Monat, C.; Lux, T.; Kamal, N.; Lang, D.; Himmelbach, A. The barley pan-genome reveals the hidden legacy of mutation breeding. Nature 2020, 588, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Milner, S.G.; Jost, M.; Taketa, S.; Mazón, E.R.; Himmelbach, A.; Oppermann, M.; Weise, S.; Knüpffer, H.; Basterrechea, M.; König, P. Genebank genomics highlights the diversity of a global barley collection. Nat. Genet. 2019, 51, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, M.; Wonneberger, R.; Haaning, A.M.; Coulter, M.; Russell, J.; Himmelbach, A.; Fiebig, A.; Muehlbauer, G.J.; Stein, N.; Waugh, R. Genomic resources for a historical collection of cultivated two-row European spring barley genotypes. bioRxiv 2023. [Google Scholar] [CrossRef]
- Wonneberger, R.; Schreiber, M.; Haaning, A.; Muehlbauer, G.J.; Waugh, R.; Stein, N. Major chromosome 5H haplotype switch structures the European two-rowed spring barley germplasm of the past 190 years. Theor. Appl. Genet. 2023, 136, 174. [Google Scholar] [CrossRef]
- Hill, C.B.; Angessa, T.T.; Zhang, X.Q.; Chen, K.; Zhou, G.; Tan, C.; Wang, P.; Westcott, S.; Li, C. A global barley panel revealing genomic signatures of breeding in modern Australian cultivars. Plant J. 2021, 106, 419–434. [Google Scholar] [CrossRef]
- Hassani, S.M.R.; Talebi, R.; Pourdad, S.S.; Naji, A.M.; Fayaz, F. In-depth genome diversity, population structure and linkage disequilibrium analysis of worldwide diverse safflower (Carthamus tinctorius L.) accessions using NGS data generated by DArTseq technology. Mol. Biol. Rep. 2020, 47, 2123–2135. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Guizado, S.J.V.; Shahid, M.Q.; Nawaz, M.A.; Habyarimana, E.; Ercişli, S.; Ali, F.; Karaköy, T.; Aasim, M.; Hatipoğlu, R. In-depth genetic diversity and population structure of endangered Peruvian Amazon Rosewood germplasm using genotyping by sequencing (GBS) Technology. Forests 2021, 12, 197. [Google Scholar] [CrossRef]
- Baloch, F.S.; Alsaleh, A.; Shahid, M.Q.; Çiftçi, V.; de Miera, L.E.S.; Aasim, M.; Nadeem, M.A.; Aktaş, H.; Özkan, H.; Hatipoğlu, R. A whole genome DArTseq and SNP analysis for genetic diversity assessment in durum wheat from central fertile crescent. PLoS ONE 2017, 12, e0167821. [Google Scholar] [CrossRef] [PubMed]
- Egea, L.A.; Mérida-García, R.; Kilian, A.; Hernandez, P.; Dorado, G. Assessment of Genetic Diversity and Structure of Large Garlic (Allium sativum) Germplasm Bank, by Diversity Arrays Technology “Genotyping-by-Sequencing” Platform (DArTseq). Front. Genet. 2017, 8, 98. [Google Scholar] [CrossRef]
- Ketema, S.; Tesfaye, B.; Keneni, G.; Amsalu Fenta, B.; Assefa, E.; Greliche, N.; Machuka, E.; Yao, N. DArTSeq SNP-based markers revealed high genetic diversity and structured population in Ethiopian cowpea [Vigna unguiculata (L.) Walp] germplasms. PLoS ONE 2020, 15, e0239122. [Google Scholar] [CrossRef]
- Lukanda, M.M.; Dramadri, I.O.; Adjei, E.A.; Arusei, P.; Gitonga, H.W.; Wasswa, P.; Edema, R.; Ssemakula, M.O.; Tukamuhabwa, P.; Tusiime, G. Genetic Diversity and Population Structure of Ugandan Soybean (Glycine max L.) Germplasm Based on DArTseq. Plant Mol. Biol. Report. 2023, 41, 417–426. [Google Scholar] [CrossRef]
- Abbasov, M.; Sansaloni, C.P.; Burgueño, J.; Petroli, C.D.; Akparov, Z.; Aminov, N.; Babayeva, S.; Izzatullayeva, V.; Hajiyev, E.; Rustamov, K.; et al. Genetic diversity analysis using DArTseq and SNP markers in populations of Aegilops species from Azerbaijan. Genet. Resour. Crop Evol. 2020, 67, 281–291. [Google Scholar] [CrossRef]
- Boczkowska, M.; Bączek, K.; Kosakowska, O.; Rucińska, A.; Podyma, W.; Węglarz, Z. Genome-Wide Diversity Analysis of Valeriana officinalis L. Using DArT-seq Derived SNP Markers. Agronomy 2020, 10, 1346. [Google Scholar] [CrossRef]
- Dziurdziak, J.; Bolc, P.; Wlodarczyk, S.; Puchta, M.; Gryziak, G.; Podyma, W.; Boczkowska, M. Multifaceted Analysis of Barley Landraces Collected during Gene Bank Expeditions in Poland at the End of the 20th Century. Agronomy 2020, 10, 1958. [Google Scholar] [CrossRef]
- Ruiz Corral, J.A.; Durán Puga, N.; Sanchez Gonzalez, J.d.J.; Ron Parra, J.; Gonzalez Eguiarte, D.R.; Holland, J.; Medina Garcia, G. Climatic adaptation and ecological descriptors of 42 Mexican maize races. Crop Sci. 2008, 48, 1502–1512. [Google Scholar] [CrossRef]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef]
- Tokatlidis, I.; Vlachostergios, D. Sustainable stewardship of the landrace diversity. Diversity 2016, 8, 29. [Google Scholar] [CrossRef]
- FAO. Practical Guide for the Application of the Genebank Standards for Plant Genetic Resources for Food and Agriculture: Conservation of Orthodox Seeds in Seed Genebanks; FAO: Rome, Italy, 2022. [Google Scholar]
- FAO. Genebank Standards for Plant Genetic Resources for Food and Agriculture; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Esquinas-Alcazar, J. Plant genetic resources. In Plant Breeding: Principles and Prospects; Springer: Berlin/Heidelberg, Germany, 1993; pp. 33–51. [Google Scholar]
- Maloy, S.; Hughes, K. Brenner’s Encyclopedia of Genetics; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Kalbarczyk, R.; Kalbarczyk, E. Research into Meteorological Drought in Poland during the Growing Season from 1951 to 2020 Using the Standardized Precipitation Index. Agronomy 2022, 12, 2035. [Google Scholar] [CrossRef]
- Kaczorowska, Z. Opady w Polsce w przekroju wieloletnim (Wyciąg z pracy). Prace i Studia Instytutu Geograficznego Uniwersytetu Warszawskiego. Katedra Klimatologii 1964, 1, 123–127. [Google Scholar]
- Pińskwar, I.; Choryński, A.; Kundzewicz, Z.W. Severe Drought in the Spring of 2020 in Poland—More of the Same? Agronomy 2020, 10, 1646. [Google Scholar] [CrossRef]
- Meresa, H.K.; Osuch, M.; Romanowicz, R. Hydro-meteorological drought projections into the 21-st century for selected Polish catchments. Water 2016, 8, 206. [Google Scholar] [CrossRef]
- Boczkowska, M.; Onyśk, A. Unused genetic resources: A case study of Polish common oat germplasm. Ann. Appl. Biol. 2016, 169, 155–165. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, R.P.S.; Singh, A.; Sharma, H.K.; Devi, G. Barley landraces: Ecological heritage for edaphic stress adaptations and sustainable production. Environ. Sustain. Indic. 2020, 6, 100035. [Google Scholar] [CrossRef]
- Ceccarelli, S. Positive interpretation of genotype by environment interactions in relation to sustainability and biodiversity. In Plant Adaptation and Crop Improvement; CABI: Wallingford, UK, 1996; pp. 467–486. [Google Scholar]
- Aboughadareh, A.P.; NaghaviI, M.R.; Khalili, M. Water deficit stress tolerance in some of barley genotypes and landraces under field conditions. Not. Sci. Biol. 2013, 5, 249–255. [Google Scholar] [CrossRef]
- Denčić, S.; Kastori, R.; Kobiljski, B.; Duggan, B. Evaluation of grain yield and its components in wheat cultivars and landraces under near optimal and drought conditions. Euphytica 2000, 113, 43–52. [Google Scholar] [CrossRef]
- Abtew, W.G.; Lakew, B.; Haussmann, B.I.; Schmid, K.J. Ethiopian barley landraces show higher yield stability and comparable yield to improved varieties in multi-environment field trials. J. Plant Breed. Crop Sci. 2015, 7, 275–291. [Google Scholar]
- Van Oosterom, E.; Ceccarelli, S.; Peacock, J. Yield response of barley to rainfall and temperature in Mediterranean environments. J. Agric. Sci. 1993, 121, 307–313. [Google Scholar] [CrossRef]
- Ceccarelli, S.; Grando, S. Barley Landraces from the Fertile Crescent; Lewis Publisher: Boca Raton, FL, USA, 2000; Volume 1. [Google Scholar]
- Tardy, F.; Créach, A.; Havaux, M. Photosynthetic pigment concentration, organization and interconversions in a pale green Syrian landrace of barley (Hordeum vulgare L., Tadmor) adapted to harsh climatic conditions. Plant Cell Environ. 1998, 21, 479–489. [Google Scholar] [CrossRef]
- Fan, Y.; Shabala, S.; Ma, Y.; Xu, R.; Zhou, M. Using QTL mapping to investigate the relationships between abiotic stress tolerance (drought and salinity) and agronomic and physiological traits. BMC Genom. 2015, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Reinert, S.; Kortz, A.; Léon, J.; Naz, A.A. Genome-wide association mapping in the global diversity set reveals new QTL controlling root system and related shoot variation in barley. Front. Plant Sci. 2016, 7, 1061. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a beneficial macronutrient in higher plants: New roles and regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhou, G.; Shimizu, H. Plant responses to drought and rewatering. Plant Signal. Behav. 2010, 5, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.-W.; Ma, Y.; Liu, W.; Zhang, S.; Wang, Y.; Cai, S.; Zhang, G.; Chater, C.C.; Chen, Z.-H.; Wu, F. Genome resequencing and transcriptome profiling reveal molecular evidence of tolerance to water deficit in barley. J. Adv. Res. 2023, 49, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Singh, V.P. A review of drought concepts. J. Hydrol. 2010, 391, 202–216. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Wells, N.; Goddard, S.; Hayes, M.J. A self-calibrating Palmer drought severity index. J. Clim. 2004, 17, 2335–2351. [Google Scholar] [CrossRef]
- Kabir, N.; Wang, X.; Lu, L.; Qanmber, G.; Liu, L.; Si, A.; Zhang, L.; Cao, W.; Yang, Z.; Yu, Y. Functional characterization of TBL genes revealed the role of GhTBL7 and GhTBL58 in cotton fiber elongation. Int. J. Biol. Macromol. 2023, 241, 124571. [Google Scholar] [CrossRef]
- Bischoff, V.; Nita, S.; Neumetzler, L.; Schindelasch, D.; Urbain, A.; Eshed, R.; Persson, S.; Delmer, D.; Scheible, W.-R. TRICHOME BIREFRINGENCE and its homolog AT5G01360 encode plant-specific DUF231 proteins required for cellulose biosynthesis in Arabidopsis. Plant Physiol. 2010, 153, 590–602. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.; Baenziger, P.S.; Börner, A. Drought stress tolerance in wheat and barley: Advances in physiology, breeding and genetics research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef]
- Kosová, K.; Vítámvás, P.; Urban, M.O.; Kholová, J.; Prášil, I.T. Breeding for enhanced drought resistance in barley and wheat-drought-associated traits, genetic resources and their potential utilization in breeding programmes. Czech J. Genet. Plant Breed. 2014, 50, 247–261. [Google Scholar] [CrossRef]
- Lippold, F.; vom Dorp, K.; Abraham, M.; Hölzl, G.; Wewer, V.; Yilmaz, J.L.; Lager, I.; Montandon, C.; Besagni, C.; Kessler, F. Fatty acid phytyl ester synthesis in chloroplasts of Arabidopsis. Plant Cell 2012, 24, 2001–2014. [Google Scholar] [CrossRef]
- Sharma, R.; Vishal, P.; Kaul, S.; Dhar, M.K. Epiallelic changes in known stress-responsive genes under extreme drought conditions in Brassica juncea (L.) Czern. Plant Cell Rep. 2017, 36, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Graether, S.P. The disordered dehydrin and its role in plant protection: A biochemical perspective. Biomolecules 2022, 12, 294. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-W.; Close, T. A newly identified barley gene, Dhn12, encoding a YSK2 DHN, is located on chromosome 6H and has embryo-specific expression. Theor. Appl. Genet. 2000, 100, 1274–1278. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Bae, J.M.; Huh, G.-H. Transcriptional regulation of the cinnamyl alcohol dehydrogenase gene from sweetpotato in response to plant developmental stage and environmental stress. Plant Cell Rep. 2010, 29, 779–791. [Google Scholar] [CrossRef] [PubMed]
- Preisner, M.; Wojtasik, W.; Kostyn, K.; Boba, A.; Czuj, T.; Szopa, J.; Kulma, A. The cinnamyl alcohol dehydrogenase family in flax: Differentiation during plant growth and under stress conditions. J. Plant Physiol. 2018, 221, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jin, Y.; Li, M.; Dong, L.; Guo, D.; Lu, C.; Qi, H. Analysis of CmCADs and three lignifying enzymes in oriental melon (‘CaiHong7′) seedlings in response to three abiotic stresses. Sci. Hortic. 2018, 237, 257–268. [Google Scholar] [CrossRef]
- Bhasin, M.; Reinherz, E.L.; Reche, P.A. Recognition and classification of histones using support vector machine. J. Comput. Biol. 2006, 13, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ren, Y.; Li, J.; Wang, Z.; Xin, Z.; Lin, T. Knock-down the expression of TaH2B-7D using virus-induced gene silencing reduces wheat drought tolerance. Biol. Res. 2019, 52, 1–9. [Google Scholar] [CrossRef]
- Al Slamh, M. Changes in the Barley Stem Proteome in Response to Drought during Grain Filling. Master’s Thesis, University of Northern Iowa, Cedar Falls, IA, USA, 2022. [Google Scholar]
- Rocha, J.; Sarkis, J.; Thomas, A.; Pitou, L.; Radzimanowski, J.; Audry, M.; Chazalet, V.; de Sanctis, D.; Palcic, M.M.; Block, M.A. Structural insights and membrane binding properties of MGD 1, the major galactolipid synthase in plants. Plant J. 2016, 85, 622–633. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kondo, M.; Fukuda, H.; Nishimura, M.; Ohta, H. Galactolipid synthesis in chloroplast inner envelope is essential for proper thylakoid biogenesis, photosynthesis, and embryogenesis. Proc. Natl. Acad. Sci. USA 2007, 104, 17216–17221. [Google Scholar] [CrossRef]
- Qi, Y.; Yamauchi, Y.; Ling, J.; Kawano, N.; Li, D.; Tanaka, K. Cloning of a putative monogalactosyldiacylglycerol synthase gene from rice (Oryza sativa L.) plants and its expression in response to submergence and other stresses. Planta 2004, 219, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, S.; Qi, L.; Yin, L.; Deng, X. Galactolipid remodeling is involved in drought-induced leaf senescence in maize. Environ. Exp. Bot. 2018, 150, 57–68. [Google Scholar] [CrossRef]
- Paczos-Grzęda, E.; Boczkowska, M.; Sowa, S.; Koroluk, A.; Toporowska, J. Hidden Diversity of Crown Rust Resistance within Genebank Resources of Avena sterilis L. Agronomy 2021, 11, 315. [Google Scholar] [CrossRef]
- Doyle, J. DNA protocols for plants. In Molecular Techniques in Taxonomy; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Whan, A.P.; Smith, A.B.; Cavanagh, C.R.; Ral, J.-P.F.; Shaw, L.M.; Howitt, C.A.; Bischof, L. GrainScan: A low cost, fast method for grain size and colour measurements. Plant Methods 2014, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958–2015. Sci. Data 2018, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ersts, P. Geographic Distance Matrix Generator (v. 1.2. 3); American Museum of Natural History, Center for Biodiversity and Conservation; Available online: http://biodiversityinformatics.amnh.org/open_source/gdmg (accessed on 10 October 2023).
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Gower, J.C. Generalized procrustes analysis. Psychometrika 1975, 40, 33–51. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annual | Growing Season | |||||||
|---|---|---|---|---|---|---|---|---|
| Min. | Accession | Max. | Accession | Min. | Accession | Max. | Accession | |
| tmax * | 18.9 | PL 42772 | 26.7 | PL 42748 | 16 | PL 42772 | 24.6 | PL 42745 |
| tmin * | 4.7 | PL 43341 | 14.4 | PL 42745 | 1.7 | PL 43341 | 12.1 | PL 43340 |
| ppt ** | 213.7 | PL 42741 | 960.7 | PL 43352 | 197.2 | PL 42741 | 880.9 | PL 43352 |
| def ** | 759.1 | PL 43352 | 1461.7 | PL 42748 | 223.5 | PL 43352 | 756.2 | PL 42745 |
| PDSI * | −0.89 | PL 42766 | −0.54 | PL 43341 | −0.84 | PL 43348 | −0.51 | PL 43351 |
| srad ** | 2450 | PL 42766 | 2688.9 | PL 42759 | 1464.5 | PL 42766 | 1697.5 | PL 42759 |
| pet ** | 1285 | PL 43350 | 1777.4 | PL 42759 | 686.3 | PL 43351 | 1002.1 | PL 42759 |
| q ** | 15.3 | PL 42761 | 379.9 | PL 43352 | 11.9 | PL 42761 | 375.9 | PL 43352 |
| soil ** | 24.5 | PL 42741 | 1017 | PL 43352 | 20.2 | PL 42741 | 883.8 | PL 43352 |
| ws * | 2.8 | PL 43349 | 4.6 | PL 43340 | 2.7 | PL 43348 | 4.5 | PL 43340 |
| aet ** | 191.7 | PL 42741 | 586.9 | PL 42766 | 175.7 | PL 42741 | 495.97 | PL 42766 |
| Parameter | Unit | Mean | Min. | Max. | Total var. | Breeding Material | Landrace |
|---|---|---|---|---|---|---|---|
| Surface area | mm2 | 24.90 | 20.62 (PL 42380) | 28.54 (PL 42759) | 8% | 9% | 6% |
| Perimeter | mm | 31.62 | 25.38 (PL 42735) | 34.63 (PL 42740) | 6% | 6% | 5% |
| Length | mm | 10.15 | 8.33 (PL 42735) | 11.46 (PL 42740) | 7% | 6% | 5% |
| Width | mm | 3.10 | 2.92 (PL 42765) | 3.36 (PL 42735) | 3% | 4% | 3% |
| Color (Red) | 8-bit/channel | 164.54 | 156.11 (PL 42380) | 185.83 (PL 40414) | 3% | 5% | 2% |
| Color (Green) | 8-bit/channel | 138.80 | 130.09 (PL 42380) | 161.83 (PL 40414) | 3% | 6% | 2% |
| Color (Blue) | 8-bit/channel | 112.52 | 106.12 (PL 42380) | 133.29 (PL 40414) | 3% | 6% | 2% |
| Abundance on Chromosomes | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1H | 2H | 3H | 4H | 5H | 6H | 7H | Total | |||
| Transitions | Purines | A > G | 192 | 259 | 232 | 178 | 275 | 189 | 233 | 1558 |
| G > A | 195 | 248 | 220 | 147 | 235 | 180 | 219 | 1444 | ||
| Pyrimidines | C > T | 181 | 233 | 249 | 161 | 252 | 175 | 237 | 1488 | |
| T > C | 140 | 230 | 238 | 145 | 224 | 135 | 213 | 1325 | ||
| Transversion | Purines > Pyrimidines | A > C | 35 | 68 | 65 | 48 | 70 | 48 | 74 | 408 |
| A > T | 32 | 46 | 34 | 34 | 44 | 34 | 34 | 258 | ||
| G > C | 103 | 122 | 122 | 86 | 122 | 89 | 110 | 754 | ||
| G > T | 57 | 94 | 85 | 46 | 87 | 55 | 68 | 492 | ||
| Pyrimidines > Purines | C > A | 49 | 86 | 76 | 47 | 61 | 52 | 77 | 448 | |
| C > G | 89 | 163 | 149 | 82 | 121 | 107 | 147 | 858 | ||
| T > A | 25 | 42 | 42 | 25 | 49 | 27 | 38 | 248 | ||
| T > G | 55 | 95 | 73 | 45 | 64 | 63 | 61 | 456 | ||
| % Ts | 61.41% | 57.53% | 59.24% | 60.44% | 61.47% | 58.84% | 59.70% | 59.72% | ||
| % Tv | 38.59% | 42.47% | 40.76% | 39.56% | 38.53% | 41.16% | 40.30% | 40.28% | ||
| Ts/Tv ratio | 1.59 | 1.35 | 1.45 | 1.53 | 1.60 | 1.43 | 1.48 | 1.48 | ||
| chr | Region | Gene ID | uniProt | Protein |
|---|---|---|---|---|
| 5H | 370151444–371703455 | HORVU5Hr1G047630 | A0A287R3K4 | Trichome birefringence-like N-terminal domain-containing protein |
| 552908671–552945241 | HORVU5Hr1G077120 | A8R7E1 | Putative esterase/lipase/thioesterase family protein | |
| HORVU5Hr1G077150 | M0Z0R4 | Atos-like conserved domain-containing protein | ||
| 6H | 19490420–19611998 | HORVU6Hr1G011030 | Q9XGV5 | Dehydrin 12 |
| 170362846–172463480 | HORVU6Hr1G035420 | O23981 | Cinnamyl alcohol dehydrogenase | |
| 341714489–352940716 | HORVU6Hr1G054420 | Q9FEX6 | coproporphyrinogen oxidase | |
| 561295698–564061223 | HORVU6Hr1G087200 | J7QBS2 | Putative proton-dependent oligopeptide/low-affinity nitrate transporter | |
| HORVU6Hr1G086940 | A0A287V1V4 | BFN domain-containing protein | ||
| HORVU6Hr1G087190 | F2E328 | Histone H2B | ||
| HORVU6Hr1G087090 | F2E586 | monogalactosyldiacylglycerol synthase | ||
| HORVU6Hr1G086910 | M0Y667 | Serine-rich protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boczkowska, M.; Puchta-Jasińska, M.; Bolc, P.; Moskal, K.; Puła, S.; Motor, A.; Bączek, K.; Groszyk, J.; Podyma, W. Characterization of the Moroccan Barley Germplasm Preserved in the Polish Genebank as a First Step towards Selecting Forms with Increased Drought Tolerance. Int. J. Mol. Sci. 2023, 24, 16350. https://doi.org/10.3390/ijms242216350
Boczkowska M, Puchta-Jasińska M, Bolc P, Moskal K, Puła S, Motor A, Bączek K, Groszyk J, Podyma W. Characterization of the Moroccan Barley Germplasm Preserved in the Polish Genebank as a First Step towards Selecting Forms with Increased Drought Tolerance. International Journal of Molecular Sciences. 2023; 24(22):16350. https://doi.org/10.3390/ijms242216350
Chicago/Turabian StyleBoczkowska, Maja, Marta Puchta-Jasińska, Paulina Bolc, Kinga Moskal, Szymon Puła, Adrian Motor, Katarzyna Bączek, Jolanta Groszyk, and Wiesław Podyma. 2023. "Characterization of the Moroccan Barley Germplasm Preserved in the Polish Genebank as a First Step towards Selecting Forms with Increased Drought Tolerance" International Journal of Molecular Sciences 24, no. 22: 16350. https://doi.org/10.3390/ijms242216350