
Genome-Wide Identification of HSF Gene Family in Kiwifruit and the Function of AeHSFA2b in Salt Tolerance


Abstract
:1. Introduction
2. Results
2.1. Identification of the AeHSF Genes in Kiwifruit
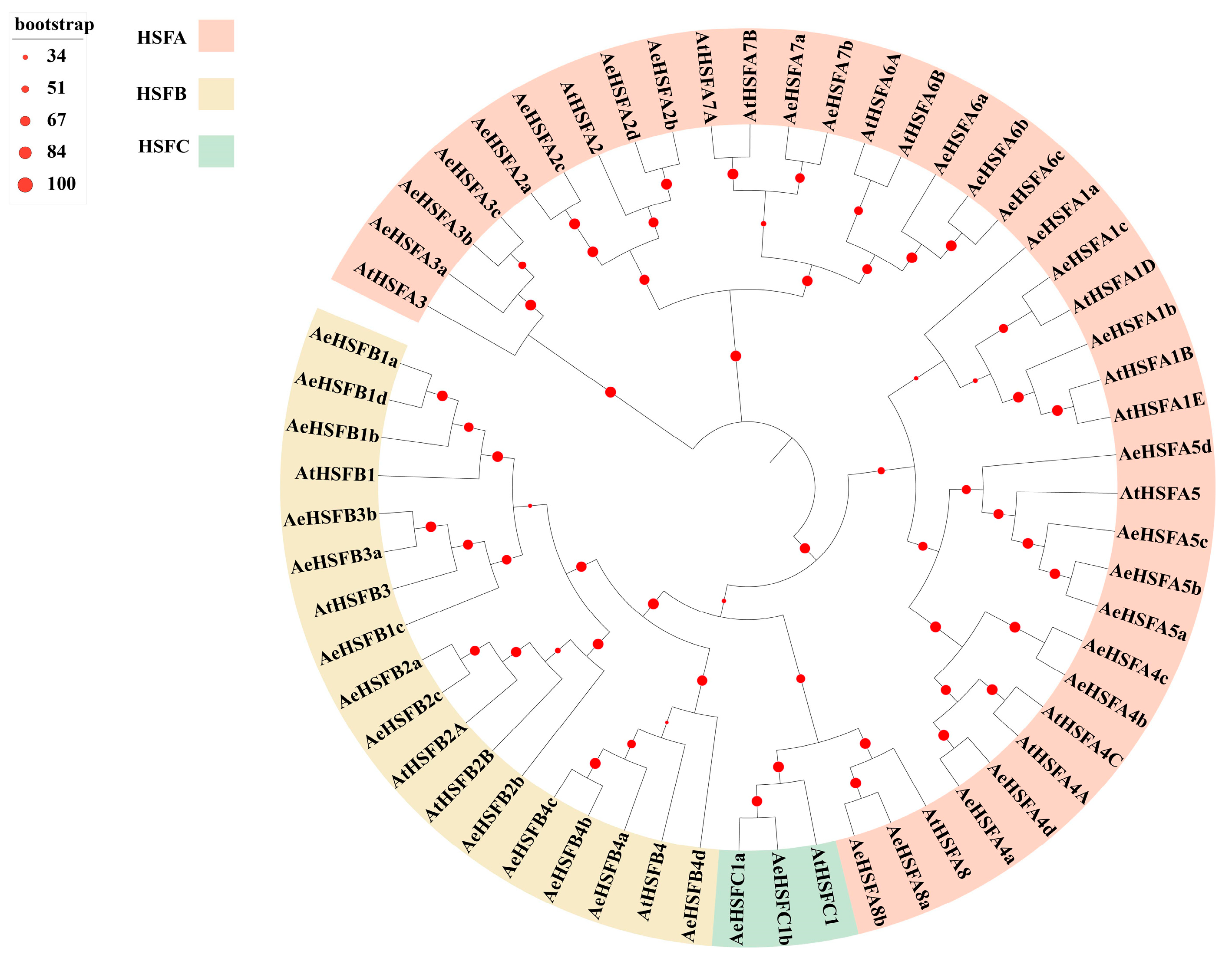
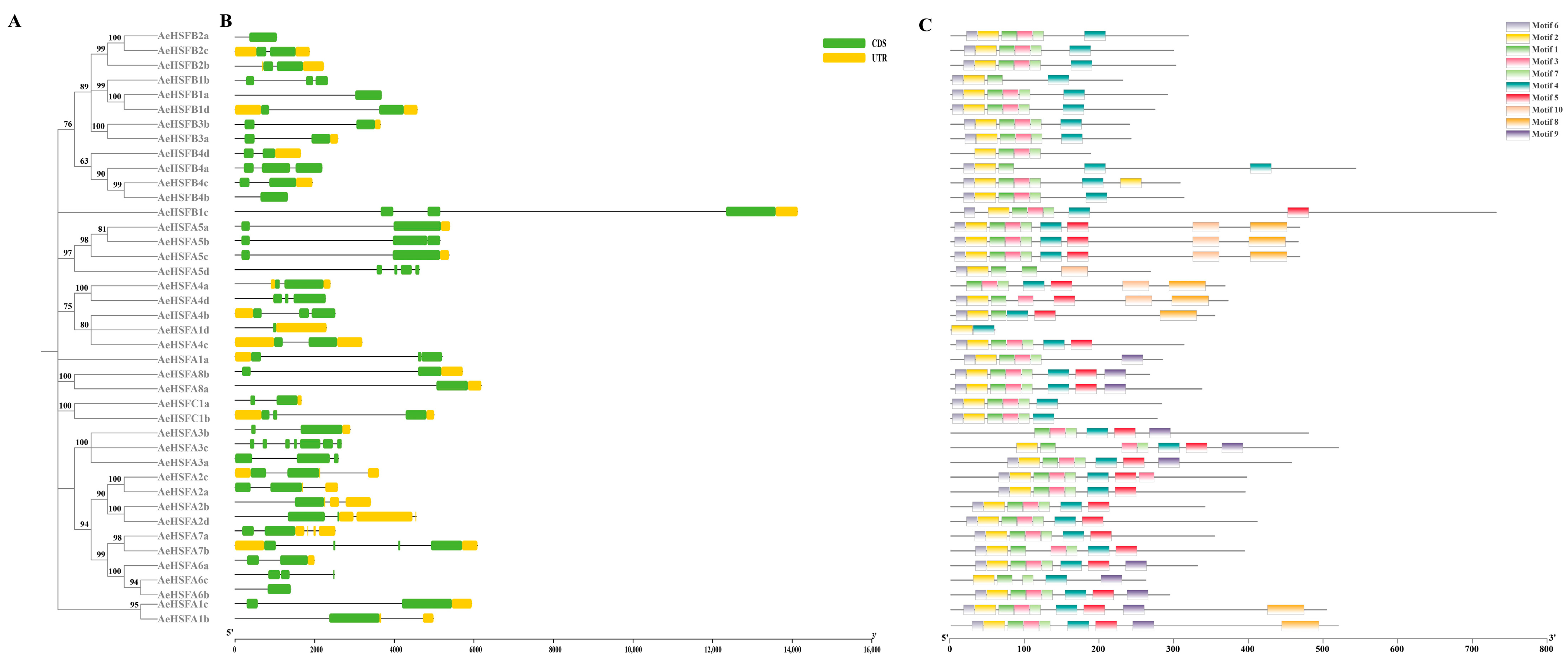
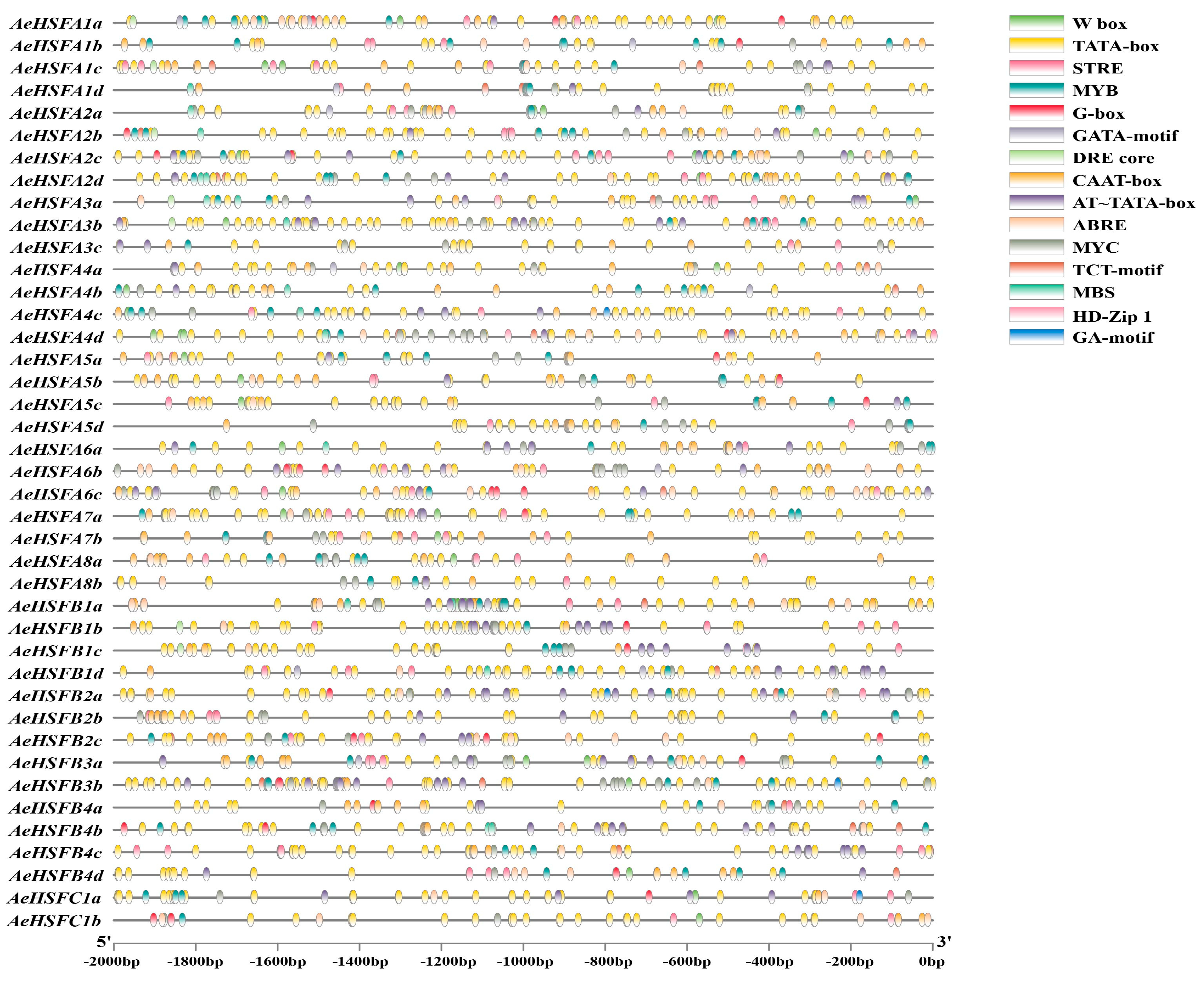
2.2. Phylogenetic Structure and Motif of AeHSF Genes
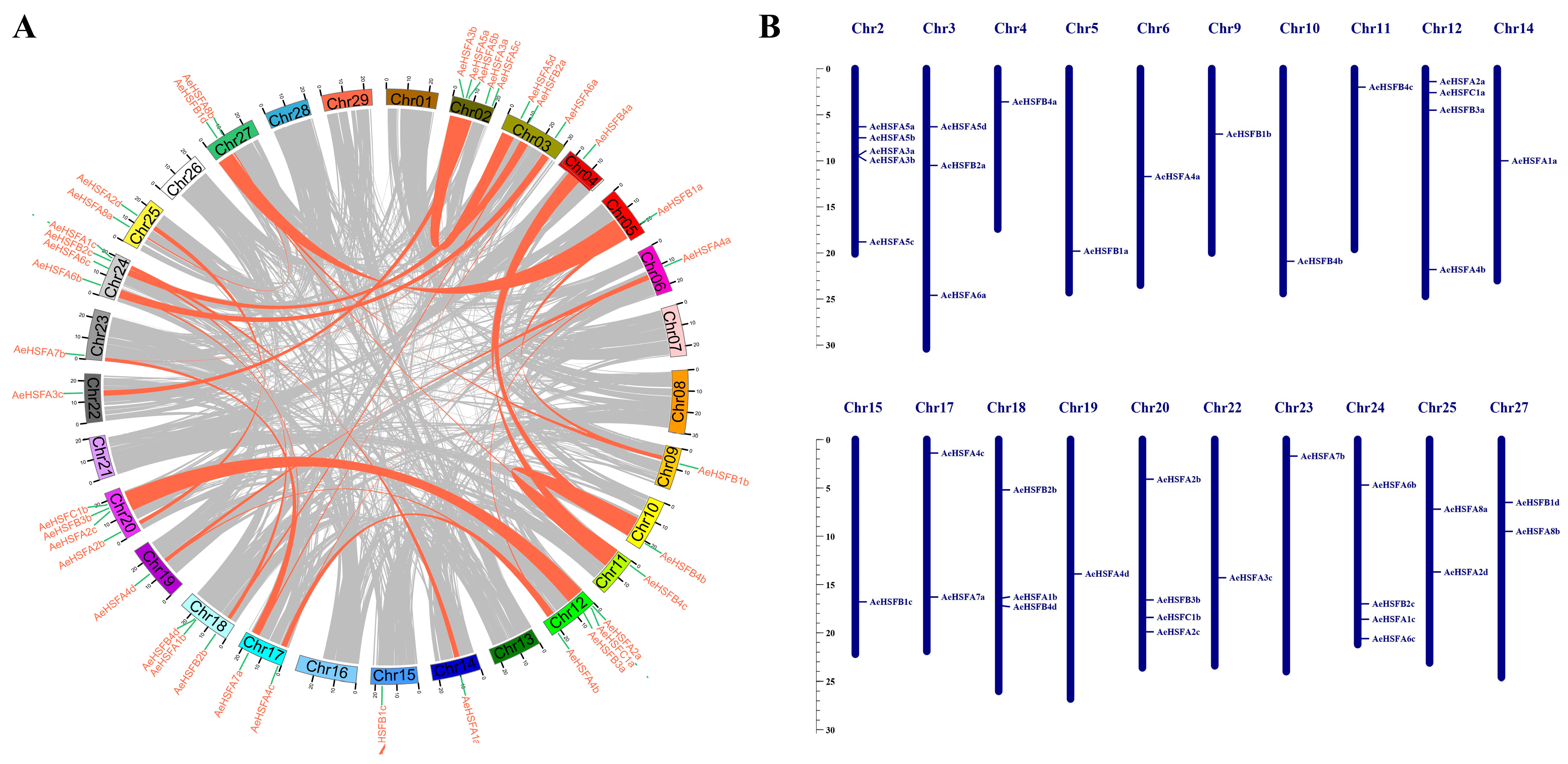
2.3. Duplication and Chromosomal Location of the AeHSF Genes
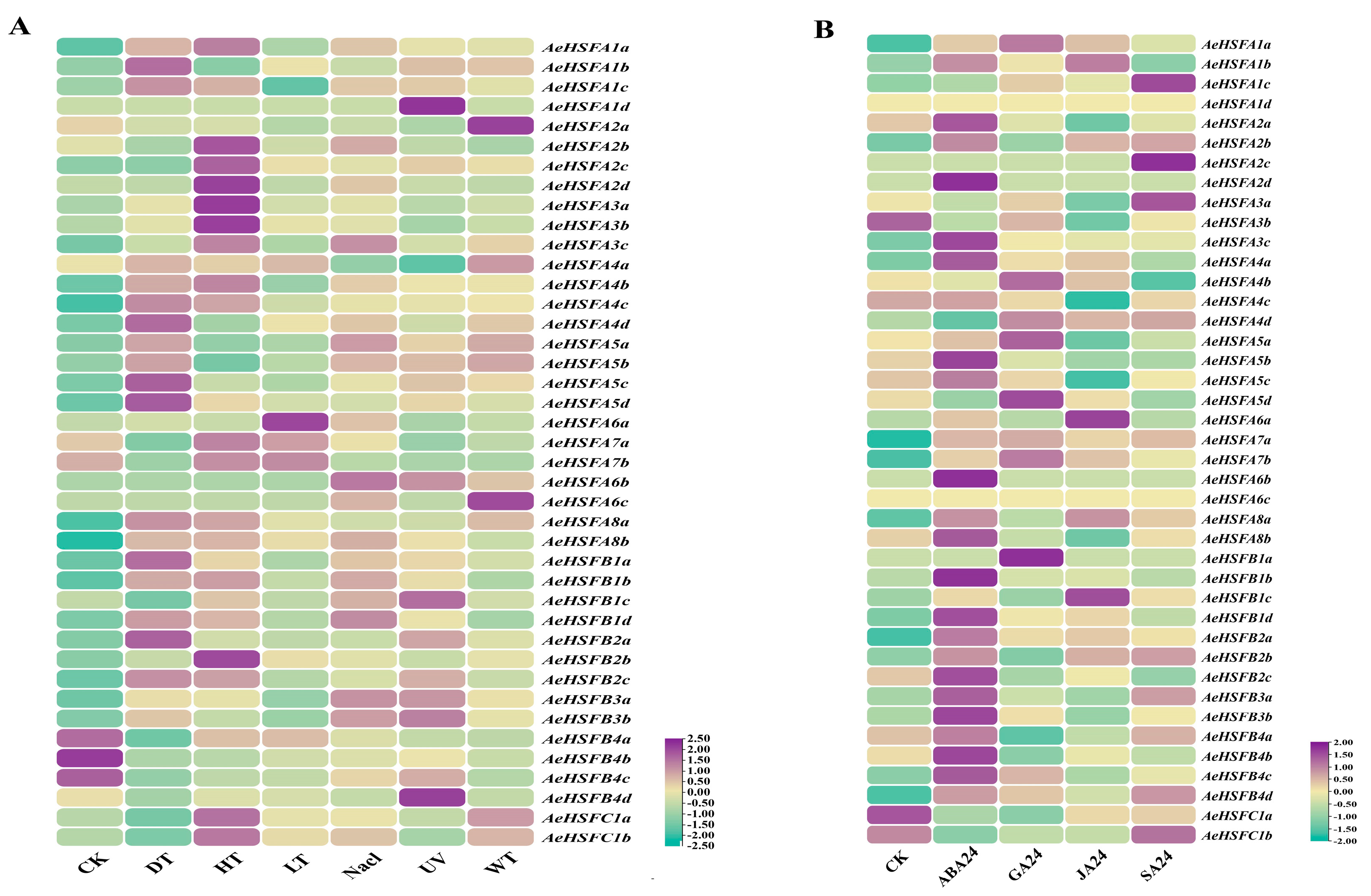
2.4. Expression of AeHSF Genes in Response to Abiotic Stresses and Hormone Treatments
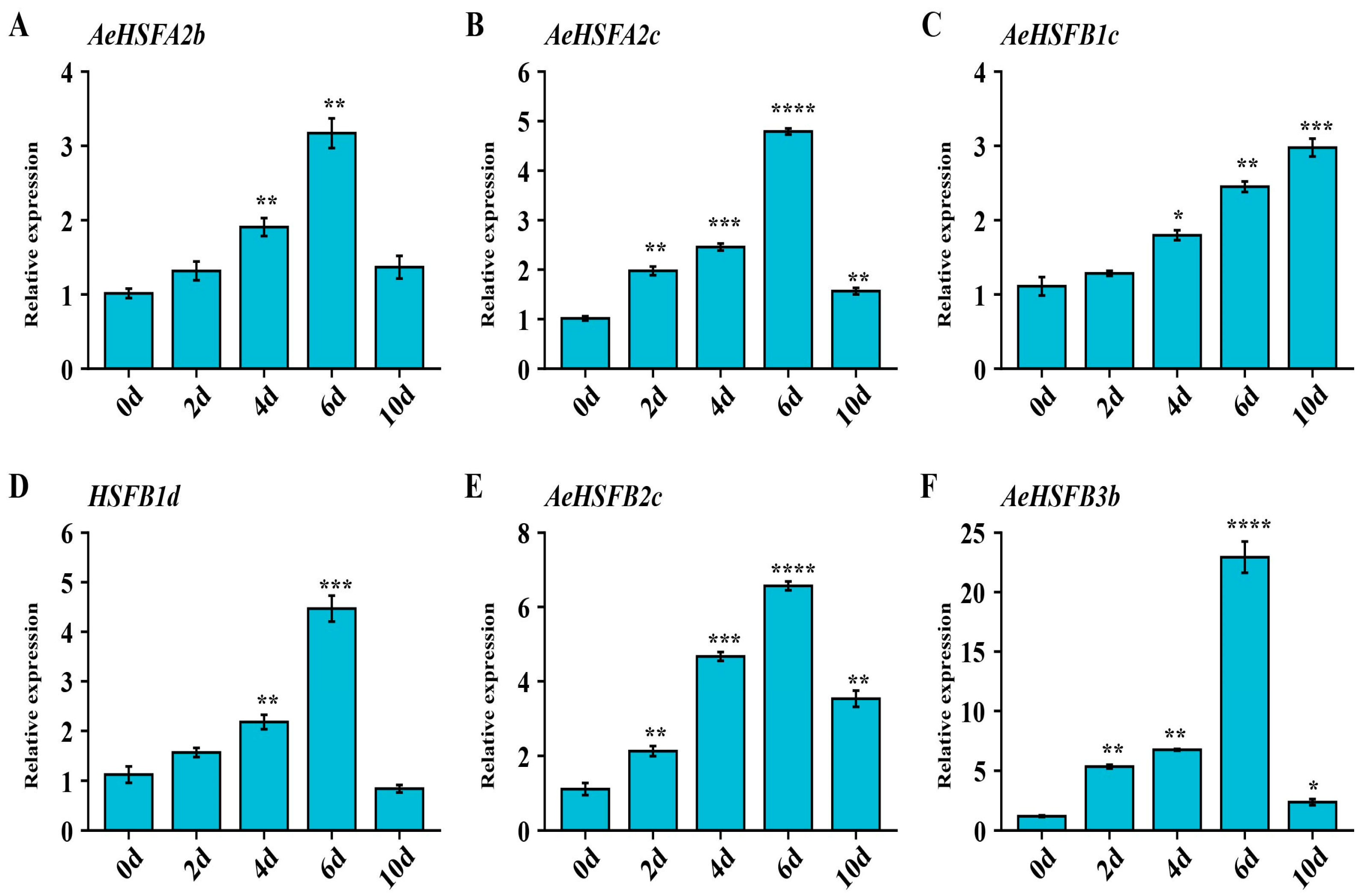
2.5. Verification of Key AeHSF Genes Expression under NaCl Stress by qRT−PCR
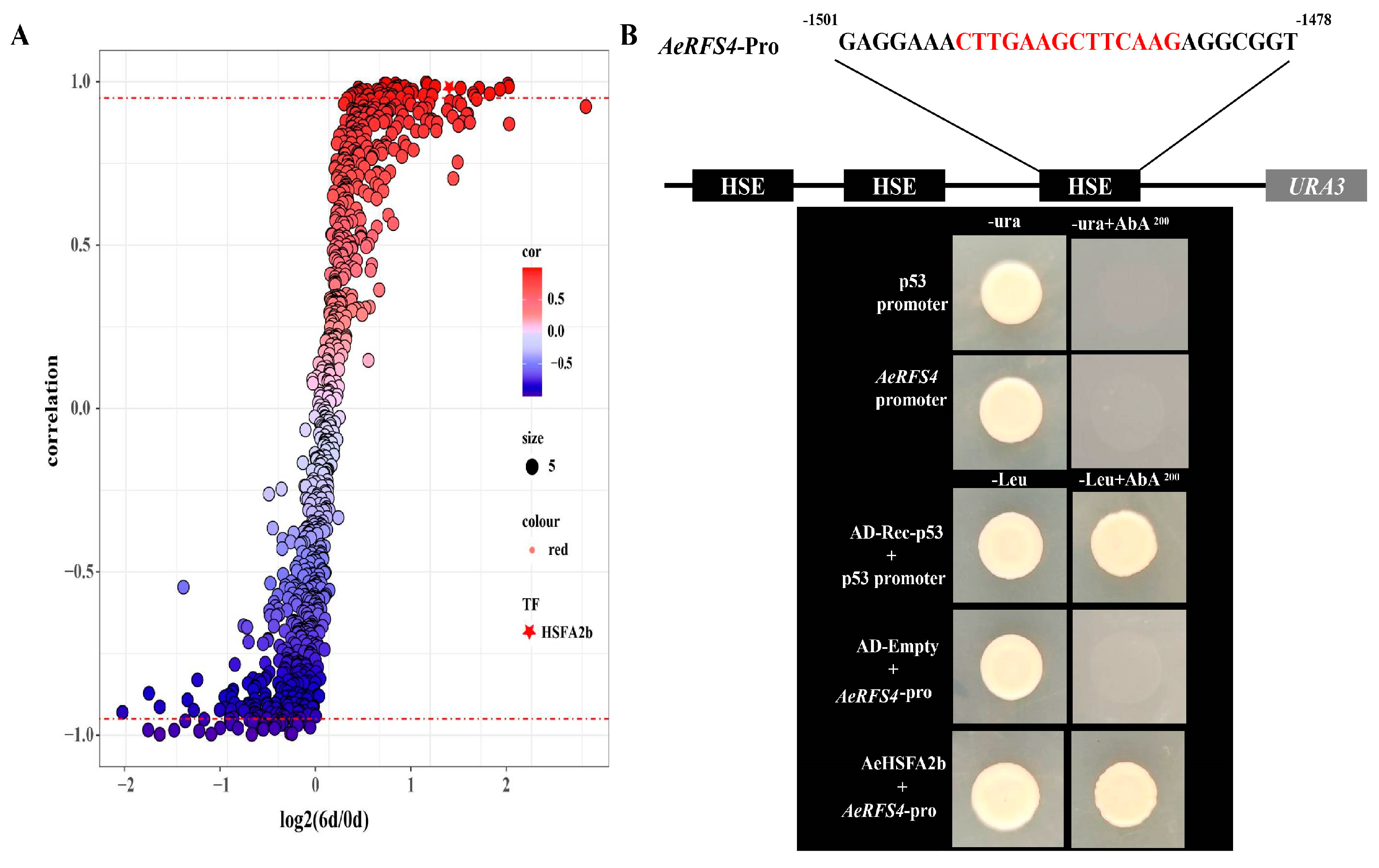
2.6. Correlation of All Transcription Factors with the Expressional Pattern of RFS4 under Salt Treatment and Transcriptional Activation of AeHSFA2b
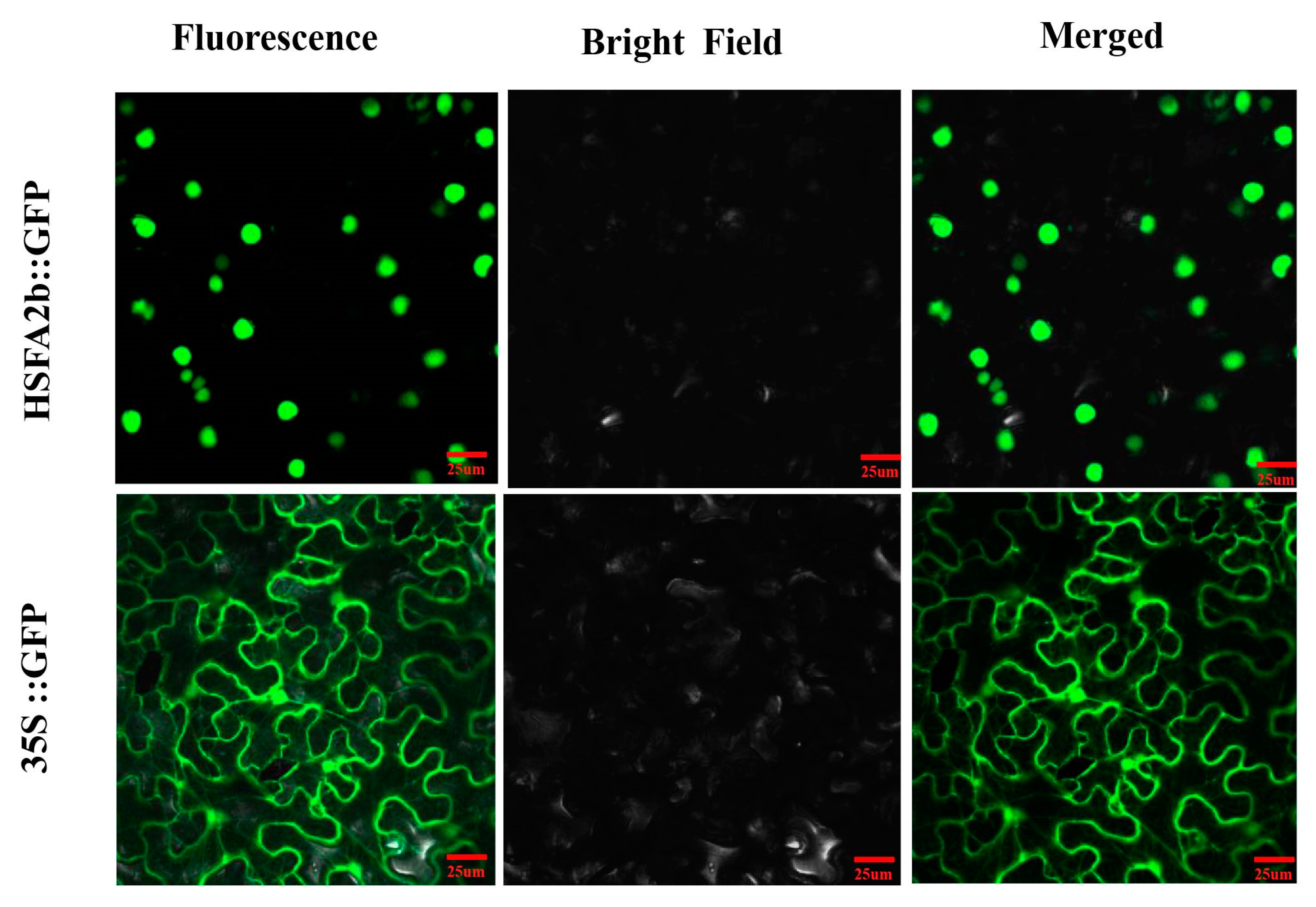
2.7. Subcellular Localization of the AeHSFA2b Protein
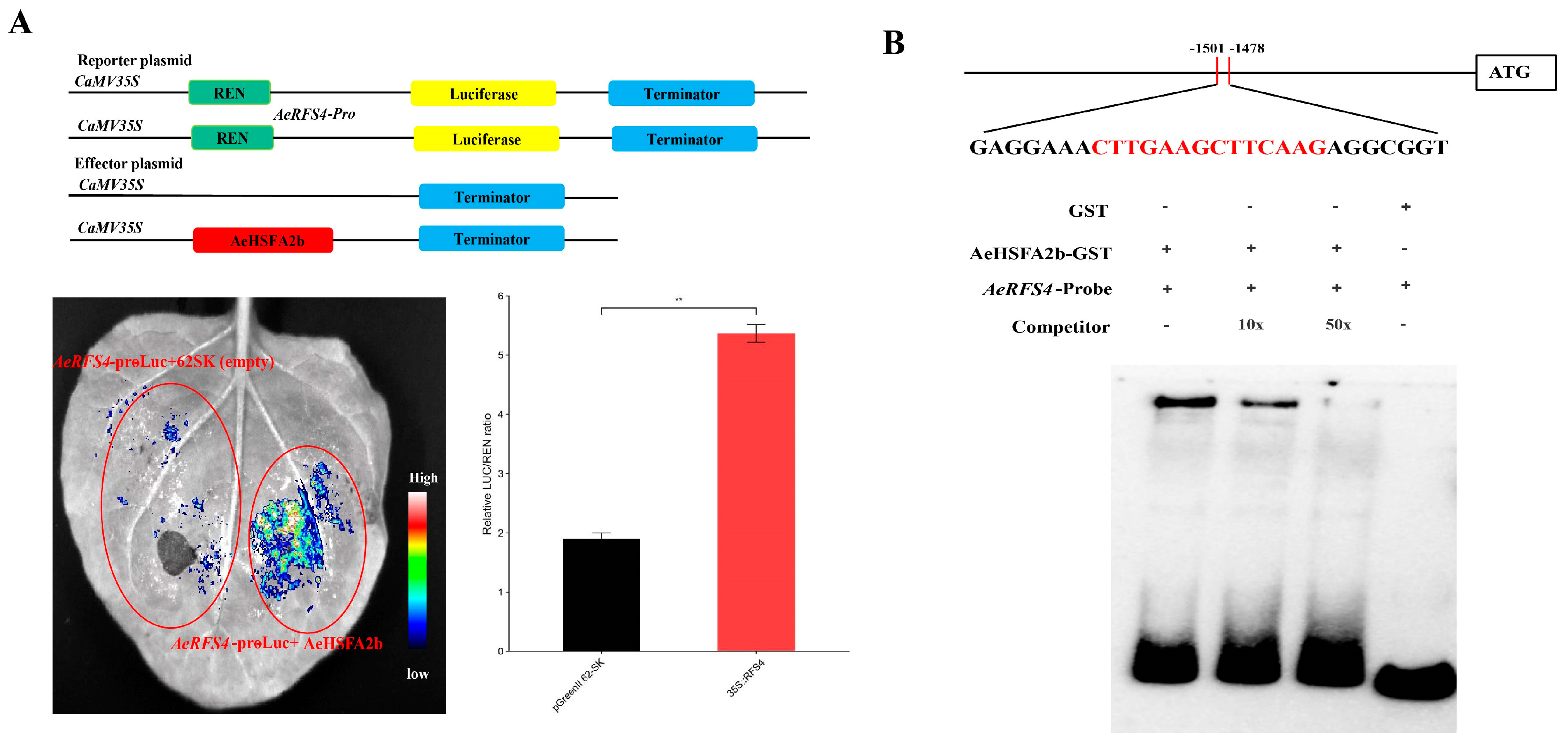
2.8. AeHSFA2b Regulates RFS4 Expression by Directly Activating and Binding to Its Promoter
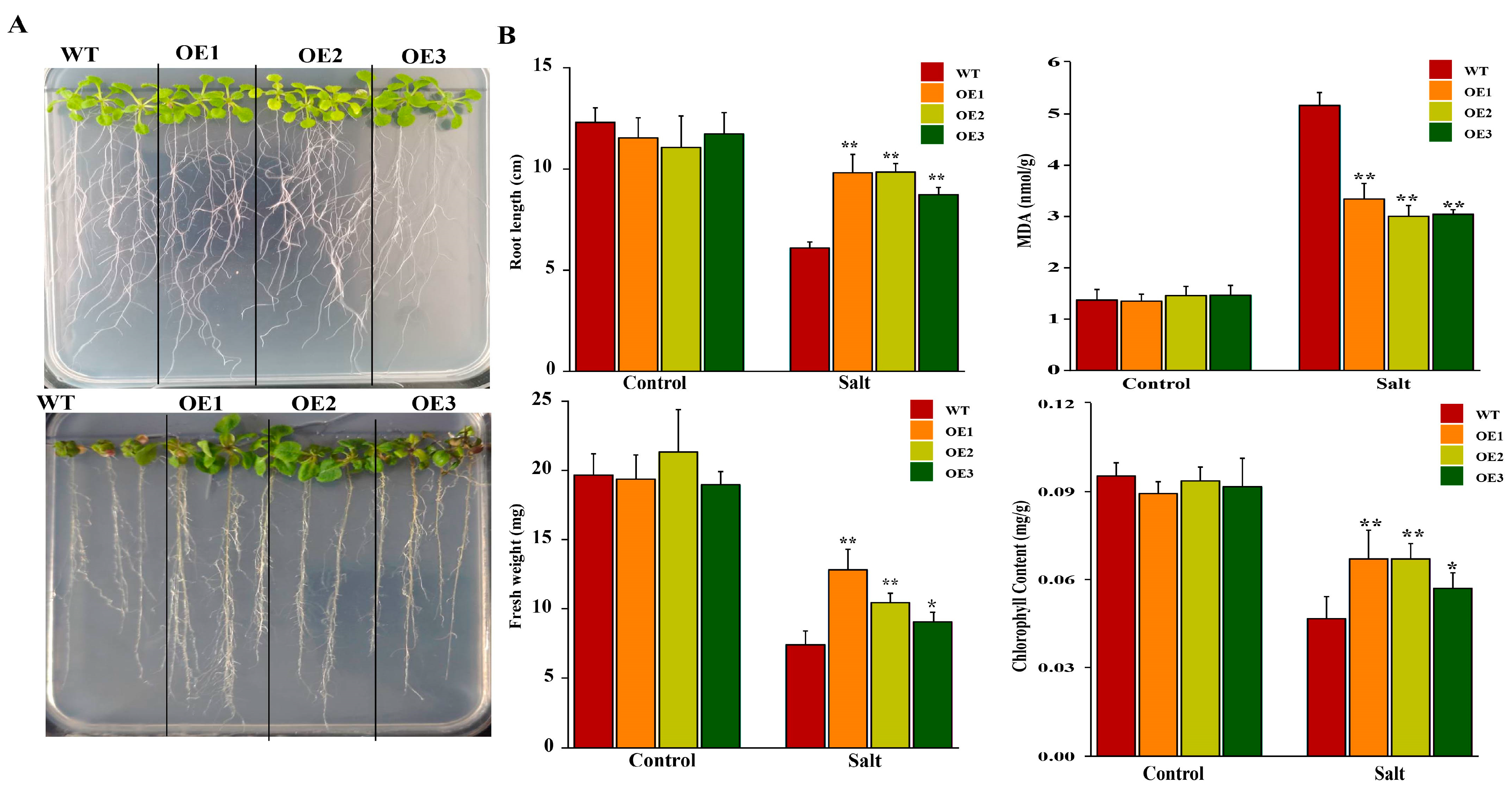
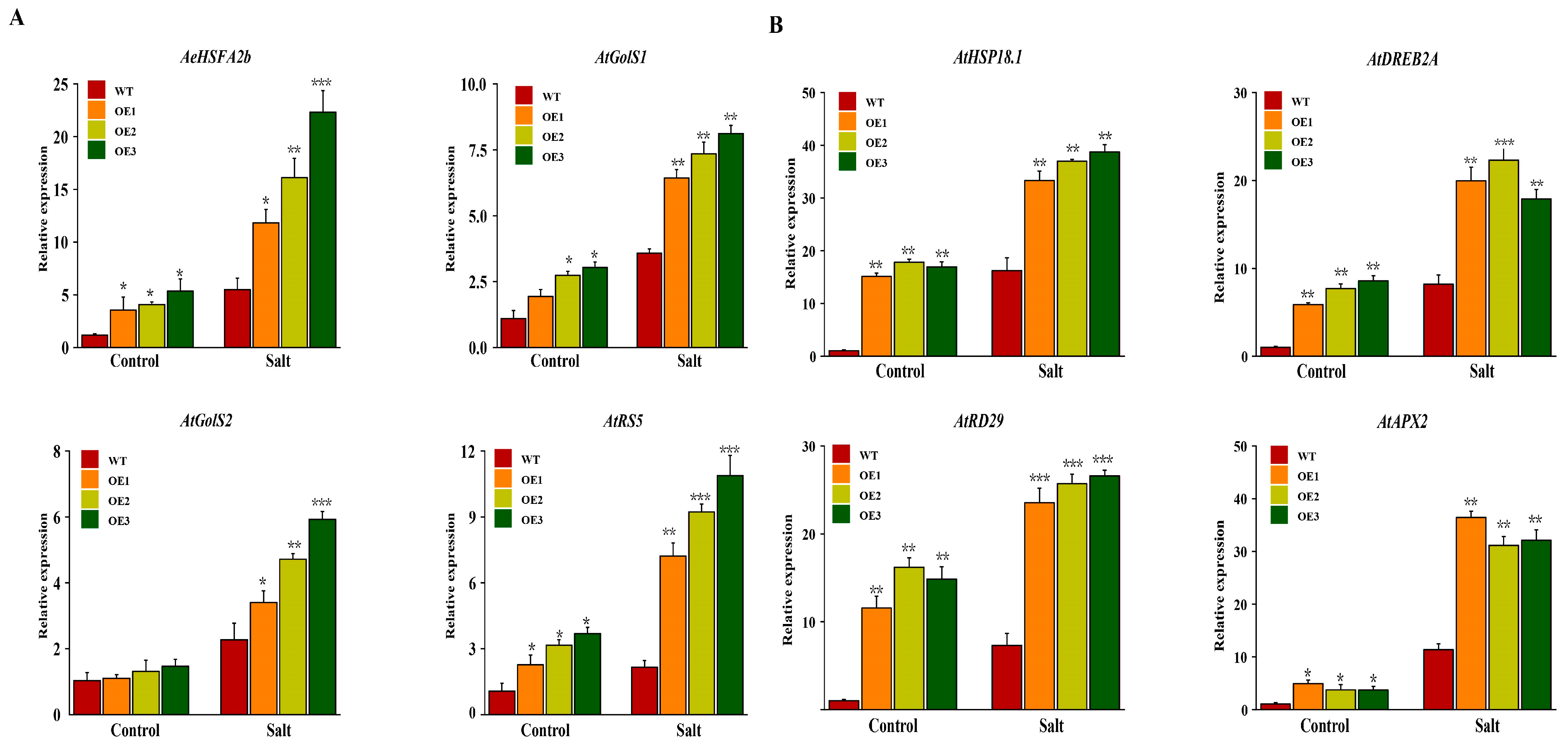
2.9. The Effects of Overexpression of AeHSFA2b in Arabidopsis
3. Discussion
4. Materials and Methods
4.1. Identification and Phylogenetic Relationships Analysis of A. eriantha AeHSF Genes
4.2. Gene Duplication and Chromosomal Locations
4.3. Expression Profiles Analysis of AeHSF Genes in Different Tissue, Hormone Treatments, and Abiotic Stress Treatments
4.4. Correlation Analysis of All Transcription Factors with the Expressional Pattern of AeRFS4 under Salt Treatment and Transcriptional Activation Analysis of AeHSFA2b
4.5. Dual-Luciferase Reporter Assay and AeHSFA2b Protein Subcellular Localization Assay
4.6. EMSA Protein Expression and Electrophoretic Mobility Shift Assay (EMSA)
4.7. AeHSFA2b-Overexpressing Arabidopsis Generation and Salt Stress Tolerance Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.-K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and Salt Tolerance in Plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription Factors Associated with Abiotic and Biotic Stress Tolerance and Their Potential for Crops Improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef] [PubMed]
- Franco-Zorrilla, J.M.; Lopez-Vidriero, I.; Carrasco, J.L.; Godoy, M.; Vera, P.; Solano, R. DNA-binding specificities of plant transcription factors and their potential to define target genes. Proc. Natl. Acad. Sci. USA 2014, 111, 2367–2372. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The Role of Stress-Responsive Transcription Factors in Modulating Abiotic Stress Tolerance in Plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Guo, M.; Liu, J.H.; Ma, X.; Luo, D.X.; Gong, Z.H.; Lu, M.H. The Plant Heat Stress Transcription Factors (HSFs): Structure, Regulation, and Function in Response to Abiotic Stresses. Front. Plant Sci. 2016, 7, 114. [Google Scholar] [CrossRef]
- Shen, Z.; Yao, J.; Sun, J.; Chang, L.; Wang, S.; Ding, M.; Qian, Z.; Zhang, H.; Zhao, N.; Sa, G.; et al. Populus euphratica HSF binds the promoter of WRKY1 to enhance salt tolerance. Plant Sci. 2015, 235, 89–100. [Google Scholar] [CrossRef]
- Scharf, K.D.; Berberich, T.; Ebersberger, I.; Nover, L. The plant heat stress transcription factor (Hsf) family: Structure, function and evolution. Biochim. Biophys. Acta 2012, 1819, 104–119. [Google Scholar] [CrossRef]
- Sorger, P.K.; Pelham, H.R.J.C. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell 1988, 54, 855–864. [Google Scholar] [CrossRef]
- Nover, L.; Bharti, K.; Döring, P.; Mishra, S.K.; Ganguli, A.; Scharf, K.D. Arabidopsis and the heat stress transcription factor world: How many heat stress transcription factors do we need? Cell Stress. Chaperones 2001, 6, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Döring, P.; Treuter, E.; Kistner, C.; Lyck, R.; Chen, A.; Nover, L. The role of AHA motifs in the activator function of tomato heat stress transcription factors HsfA1 and HsfA2. Plant Cell 2000, 12, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Rafudeen, M.; Golldack, D. Physiological aspects of raffinose family oligosaccharides in plants: Protection against abiotic stress. Plant Biol. 2013, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Thole, J.M.; Nielsen, E. Phosphoinositides in plants: Novel functions in membrane trafficking. Curr. Opin. Plant Biol. 2008, 11, 620–631. [Google Scholar] [CrossRef]
- Lahuta, L.B.; Pluskota, W.E.; Stelmaszewska, J.; Szablinska, J. Dehydration induces expression of GALACTINOL SYNTHASE and RAFFINOSE SYNTHASE in seedlings of pea (Pisum sativum L.). J. Plant Physiol. 2014, 171, 1306–1314. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Vieira, L.G.E. Involvement of the galactinol synthase gene in abiotic and biotic stress responses: A review on current knowledge. Plant Gene 2020, 24, 100258. [Google Scholar] [CrossRef]
- Fan, Y.; Yu, M.; Liu, M.; Zhang, R.; Sun, W.; Qian, M.; Duan, H.; Chang, W.; Ma, J.; Qu, C.; et al. Genome-Wide Identification, Evolutionary and Expression Analyses of the GALACTINOL SYNTHASE Gene Family in Rapeseed and Tobacco. Int. J. Mol. Sci. 2017, 18, 2768. [Google Scholar] [CrossRef]
- Shen, Y.; Jia, B.; Wang, J.; Cai, X.; Hu, B.; Wang, Y.; Chen, Y.; Sun, M.; Sun, X. Functional Analysis of Arabidopsis thaliana Galactinol Synthase AtGolS2 in Response to Abiotic Stress. Mol. Plant Breed. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Liu, L.; Wu, X.; Sun, W.; Yu, X.; Demura, T.; Li, D.; Zhuge, Q. Galactinol synthase confers salt-stress tolerance by regulating the synthesis of galactinol and raffinose family oligosaccharides in poplar. Ind. Crops Prod. 2021, 165, 113432. [Google Scholar] [CrossRef]
- Kito, K.; Yamane, K.; Yamamori, T.; Matsuhira, H.; Tanaka, Y.; Takabe, T. Isolation, functional characterization and stress responses of raffinose synthase genes in sugar beet. J. Plant Biochem. Biotechnol. 2017, 27, 36–45. [Google Scholar] [CrossRef]
- Li, X.; Li, J.; Soejarto, D.D. Advances in the study of the systematics of Actinidia Lindley. Front. Biol. China 2009, 4, 55–61. [Google Scholar] [CrossRef]
- Richardson, D.P.; Ansell, J.; Drummond, L.N. The nutritional and health attributes of kiwifruit: A review. Eur. J. Nutr. 2018, 57, 2659–2676. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Zhang, Y.J.; Li, Z.; Bai, D.F.; Zhong, Y.P.; Fang, J.B. Effect of salt stress on growth, physiological and biochemical characters of four kiwifruit genotypes. Sci. Hortic. 2020, 271, 109473. [Google Scholar] [CrossRef]
- Chen, J.; Fang, J.; Qi, X.; Gu, H.; Lin, M.; Zhang, W.; Wei, C. Research progress on rootstock of kiwifruit. J. Fruit Sci. 2015, 32, 959–968. [Google Scholar]
- Yang, J.; Ling, C.; Liu, Y.; Zhang, H.; Hussain, Q.; Lyu, S.; Wang, S.; Liu, Y. Genome-Wide Expression Profiling Analysis of Kiwifruit GolS and RFS Genes and Identification of AcRFS4 Function in Raffinose Accumulation. Int. J. Mol. Sci. 2022, 23, 8836. [Google Scholar] [CrossRef]
- Scarpeci, T.E.; Zanor, M.I.; Valle, E.M. Investigating the role of plant heat shock proteins during oxidative stress. Plant Signal Behav. 2008, 3, 856–857. [Google Scholar] [CrossRef]
- Arrigo, A.P. Human small heat shock proteins: Protein interactomes of homo- and hetero-oligomeric complexes: An update. FEBS Lett. 2013, 587, 1959–1969. [Google Scholar] [CrossRef]
- Guo, J.; Wu, J.; Ji, Q.; Wang, C.; Luo, L.; Yuan, Y.; Wang, Y.; Wang, J. Genome-wide analysis of heat shock transcription factor families in rice and Arabidopsis. J. Genet. Genom. Yi Chuan Xue Bao 2008, 35, 105–118. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhu, W.; Zhang, H.; Liu, N.; Tian, S. Heat shock factors in tomatoes: Genome-wide identification, phylogenetic analysis and expression profiling under development and heat stress. PeerJ 2016, 4, e1961. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Liu, B.; Zhang, Y.; Li, G.; Guo, X. Genome-wide identification and abiotic stress-responsive pattern of heat shock transcription factor family in Triticum aestivum L. BMC Genom. 2019, 20, 257. [Google Scholar] [CrossRef]
- Guo, M.; Lu, J.P.; Zhai, Y.F.; Chai, W.G.; Gong, Z.H.; Lu, M.H. Genome-wide analysis, expression profile of heat shock factor gene family (CaHsfs) and characterisation of CaHsfA2 in pepper (Capsicum annuum L.). BMC Plant Biol. 2015, 15, 151. [Google Scholar] [CrossRef]
- Tang, R.; Zhu, W.; Song, X.; Lin, X.; Cai, J.; Wang, M.; Yang, Q. Genome-Wide Identification and Function Analyses of Heat Shock Transcription Factors in Potato. Front. Plant Sci. 2016, 7, 490. [Google Scholar] [CrossRef]
- Lin, Y.X.; Jiang, H.Y.; Chu, Z.X.; Tang, X.L.; Zhu, S.W.; Cheng, B.J. Genome-wide identification, classification and analysis of heat shock transcription factor family in maize. BMC Genom. 2011, 12, 76. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.; Ren, H.; Zhang, C.; Xiao, D.; Li, Y.; Hou, X.; Liu, T. Regulatory interaction of BcWRKY33A and BcHSFA4A promotes salt tolerance in non-heading Chinese cabbage [Brassica campestris (syn. Brassica rapa) ssp. chinensis]. Hortic. Res. 2022, 9, uhac113. [Google Scholar] [CrossRef]
- Dossa, K.; Diouf, D.; Cisse, N. Genome-Wide Investigation of Hsf Genes in Sesame Reveals Their Segmental Duplication Expansion and Their Active Role in Drought Stress Response. Front. Plant Sci. 2016, 7, 1522. [Google Scholar] [CrossRef]
- Wan, X.; Yang, J.; Guo, C.; Bao, M.; Zhang, J. Genome-wide identification and classification of the Hsf and sHsp gene families in Prunus mume, and transcriptional analysis under heat stress. PeerJ 2019, 7, e7312. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Qi, X.; Zhang, Y.; Zhang, R.; Wang, C.; Sun, T.; Zheng, J.; Lu, Y. Genome-wide analysis of the heat shock transcription factor gene family in Sorbus pohuashanensis (Hance) Hedl identifies potential candidates for resistance to abiotic stresses. Plant Physiol. Biochem. 2022, 175, 68–80. [Google Scholar] [CrossRef]
- Liu, G.; Chai, F.; Wang, Y.; Jiang, J.; Duan, W.; Wang, Y.; Wang, F.; Li, S.; Wang, L. Genome-wide Identification and Classification of HSF Family in Grape, and Their Transcriptional Analysis under Heat Acclimation and Heat Stress. Hortic. Plant J. 2018, 4, 133–143. [Google Scholar] [CrossRef]
- Shen, R.-J.; Xing, M.-Y.; Wang, W.-Q.; Su, W.-Y.; Zhu, J.-Z.; Li, K.-F.; Zhang, Y.; Allan, A.C.; Grierson, D.; Yin, X.-R.; et al. Over-expression of heat shock factor AcHsfA2-1 upregulates transcripts of multiple genes and enhances heat tolerance of kiwifruit plants. Environ. Exp. Bot. 2023, 207, 105196. [Google Scholar] [CrossRef]
- Fragkostefanakis, S.; Mesihovic, A.; Simm, S.; Paupière, M.J.; Hu, Y.; Paul, P.; Mishra, S.K.; Tschiersch, B.; Theres, K.; Bovy, A.; et al. HsfA2 Controls the Activity of Developmentally and Stress-Regulated Heat Stress Protection Mechanisms in Tomato Male Reproductive Tissues. Plant Physiol. 2016, 170, 2461–2477. [Google Scholar] [CrossRef]
- Ogawa, D.; Yamaguchi, K.; Nishiuchi, T. High-level overexpression of the Arabidopsis HsfA2 gene confers not only increased themotolerance but also salt/osmotic stress tolerance and enhanced callus growth. J. Exp. Bot. 2007, 58, 3373–3383. [Google Scholar] [CrossRef]
- Xiang, J.; Ran, J.; Zou, J.; Zhou, X.; Liu, A.; Zhang, X.; Peng, Y.; Tang, N.; Luo, G.; Chen, X. Heat shock factor OsHsfB2b negatively regulates drought and salt tolerance in rice. Plant Cell Rep. 2013, 32, 1795–1806. [Google Scholar] [CrossRef]
- Huang, Y.C.; Niu, C.Y.; Yang, C.R.; Jinn, T.L. The Heat Stress Factor HSFA6b Connects ABA Signaling and ABA-Mediated Heat Responses. Plant Physiol. 2016, 172, 1182–1199. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jia, H.; Li, J.; Li, Y.; Lu, M.; Hu, J. Molecular evolution and expression divergence of the Populus euphratica Hsf genes provide insight into the stress acclimation of desert poplar. Sci. Rep. 2016, 6, 30050. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujita, Y.; Sayama, H.; Kidokoro, S.; Maruyama, K.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 2010, 61, 672–685. [Google Scholar] [CrossRef]
- Qi, C.; Dong, D.; Li, Y.; Wang, X.; Guo, L.; Liu, L.; Dong, X.; Li, X.; Yuan, X.; Ren, S.; et al. Heat shock-induced cold acclimation in cucumber through CsHSFA1d-activated JA biosynthesis and signaling. Plant J. 2022, 111, 85–102. [Google Scholar] [CrossRef]
- Liu, M.; Huang, Q.; Sun, W.; Ma, Z.; Huang, L.; Wu, Q.; Tang, Z.; Bu, T.; Li, C.; Chen, H. Genome-wide investigation of the heat shock transcription factor (Hsf) gene family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 871. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wan, X.L.; Yu, J.Y.; Wang, K.L.; Zhang, J. Genome-Wide Identification, Classification, and Expression Analysis of the Hsf Gene Family in Carnation (Dianthus caryophyllus). Int. J. Mol. Sci. 2019, 20, 5233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Huang, C.; Zhang, L.; Liu, H.; Yu, J.; Hu, Z.; Hua, W. Systematic Analysis of Hsf Family Genes in the Brassica napus Genome Reveals Novel Responses to Heat, Drought and High CO2 Stresses. Front. Plant Sci. 2017, 8, 1174. [Google Scholar] [CrossRef]
- Zhang, H.; Jin, J.; Fan, D.; Zhao, X.; Jia, P.; Hao, Q.; Yang, L.; Niu, J. Identification and expression analysis of HSF transcription factor family in wild jujube under heat stress. ISHS Acta Hortic. 2021, 1350, 205–214. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Z.; Guo, C.; Zhao, X.; Li, Z.; Mou, Y.; Sun, Q.; Wang, J.; Yuan, C.; Li, C.; et al. Hsf transcription factor gene family in peanut (Arachis hypogaea L.): Genome-wide characterization and expression analysis under drought and salt stresses. Front. Plant Sci. 2023, 14, 1214732. [Google Scholar] [CrossRef] [PubMed]
- Peng, S.; Zhu, Z.; Zhao, K.; Shi, J.; Yang, Y.; He, M.; Wang, Y. A novel heat shock transcription factor, VpHsf1, from Chinese wild Vitis pseudoreticulata is involved in biotic and abiotic stresses. Plant Mol. Biol. Report. 2013, 31, 240–247. [Google Scholar] [CrossRef]
- Ma, H.; Wang, C.; Yang, B.; Cheng, H.; Wang, Z.; Mijiti, A.; Ren, C.; Qu, G.; Zhang, H.; Ma, L. CarHSFB2, a class B heat shock transcription factor, is involved in different developmental processes and various stress responses in chickpea (Cicer arietinum L.). Plant Mol. Biol. Report. 2016, 34, 1–14. [Google Scholar] [CrossRef]
- Yun, L.; Zhang, Y.; Li, S.; Yang, J.; Wang, C.; Zheng, L.; Ji, L.; Yang, J.; Song, L.; Shi, Y.; et al. Phylogenetic and expression analyses of HSF gene families in wheat (Triticum aestivum L.) and characterization of TaHSFB4-2B under abiotic stress. Front. Plant Sci. 2022, 13, 1047400. [Google Scholar] [CrossRef] [PubMed]
- Bi, H.; Zhao, Y.; Li, H.; Liu, W. Wheat Heat Shock Factor TaHsfA6f Increases ABA Levels and Enhances Tolerance to Multiple Abiotic Stresses in Transgenic Plants. Int. J. Mol. Sci. 2020, 21, 3121. [Google Scholar] [CrossRef]
- Schramm, F.; Ganguli, A.; Kiehlmann, E.; Englich, G.; Walch, D.; von Koskull-Döring, P. The heat stress transcription factor HsfA2 serves as a regulatory amplifier of a subset of genes in the heat stress response in Arabidopsis. Plant Mol. Biol. 2006, 60, 759–772. [Google Scholar] [CrossRef]
- Gu, L.; Jiang, T.; Zhang, C.; Li, X.; Wang, C.; Zhang, Y.; Li, T.; Dirk, L.M.A.; Downie, A.B.; Zhao, T. Maize HSFA2 and HSBP2 antagonistically modulate raffinose biosynthesis and heat tolerance in Arabidopsis. Plant J. 2019, 100, 128–142. [Google Scholar] [CrossRef]
- Song, C.; Chung, W.S.; Lim, C.O. Overexpression of Heat Shock Factor Gene HsfA3 Increases Galactinol Levels and Oxidative Stress Tolerance in Arabidopsis. Mol. Cells 2016, 39, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Lang, S.; Liu, X.; Xue, H.; Li, X.; Wang, X. Functional characterization of BnHSFA4a as a heat shock transcription factor in controlling the re-establishment of desiccation tolerance in seeds. J. Exp. Bot. 2017, 68, 2361–2375. [Google Scholar] [CrossRef]
- Yue, J.; Liu, J.; Tang, W.; Wu, Y.Q.; Tang, X.; Li, W.; Yang, Y.; Wang, L.; Huang, S.; Fang, C.; et al. Kiwifruit Genome Database (KGD): A comprehensive resource for kiwifruit genomics. Hortic. Res. 2020, 7, 117. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J.; Niu, X.-Q.; Zheng, X.-L.; Chen, X.; Zheng, G.-H.; Wu, J.-C. Comparative transcriptome analysis reveals key genes potentially related to organic acid and sugar accumulation in loquat. PLoS ONE 2021, 16, e0238873. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Genome ID | Chr ID | Start (bp) | End (bp) | Protein Length (aa) | Molecular Weight (kDa) | PI |
|---|---|---|---|---|---|---|---|
| DTZ79_14g07720 | AeHSFA1a | Chr14 | 10,001,679 | 10,006,882 | 283 | 31,275.38 | 5.47 |
| DTZ79_18g08160 | AeHSFA1b | Chr18 | 16,404,715 | 16,409,700 | 503 | 55,972.83 | 6.19 |
| DTZ79_24g12140 | AeHSFA1c | Chr24 | 18,591,496 | 18,597,442 | 512 | 56,190.77 | 7.76 |
| DTZ79_12g13240 | AeHSFA1d | Chr12 | 23,908,734 | 23,911,031 | 59 | 6772.63 | 7.83 |
| DTZ79_12g01030 | AeHSFA2a | Chr12 | 1,389,789 | 1,392,367 | 394 | 45,138.73 | 4.53 |
| DTZ79_20g03210 | AeHSFA2b | Chr20 | 4,073,776 | 4,077,184 | 340 | 39,266.25 | 6.43 |
| DTZ79_20g13080 | AeHSFA2c | Chr20 | 19,899,248 | 19,902,860 | 396 | 45,173.66 | 10.25 |
| DTZ79_25g04820 | AeHSFA2d | Chr25 | 13,660,194 | 13,664,745 | 410 | 47,170.3 | 6.05 |
| DTZ79_02g08920 | AeHSFA3a | Chr02 | 9,444,887 | 9,447,485 | 456 | 51,324.32 | 5.1 |
| DTZ79_02g08960 | AeHSFA3b | Chr02 | 9,502,675 | 9,505,569 | 479 | 53,705.08 | 8.16 |
| DTZ79_22g06060 | AeHSFA3c | Chr22 | 14,257,374 | 14,260,052 | 519 | 58,377.59 | 6.79 |
| DTZ79_06g06820 | AeHSFA4a | Chr06 | 11,684,714 | 11,687,108 | 367 | 41,842.63 | 7.87 |
| DTZ79_12g12240 | AeHSFA4b | Chr12 | 21,843,255 | 21,845,772 | 353 | 39,637.21 | 5.12 |
| DTZ79_17g00990 | AeHSFA4c | Chr17 | 1,375,483 | 1,378,676 | 312 | 36,435.81 | 5.95 |
| DTZ79_19g05240 | AeHSFA4d | Chr19 | 13,922,263 | 13,924,540 | 371 | 42,169.33 | 8.3 |
| DTZ79_02g06560 | AeHSFA5a | Chr02 | 6,267,378 | 6,272,776 | 467 | 52,808.94 | 5.6 |
| DTZ79_02g07500 | AeHSFA5b | Chr02 | 7,451,281 | 7,456,434 | 465 | 52,607.75 | 9.15 |
| DTZ79_02g14180 | AeHSFA5c | Chr02 | 18,802,710 | 18,808,087 | 467 | 52,763.84 | 8.93 |
| DTZ79_03g06230 | AeHSFA5d | Chr03 | 6,337,642 | 6,342,273 | 267 | 29,710.02 | 4.62 |
| DTZ79_03g18750 | AeHSFA6a | Chr03 | 24,588,106 | 24,590,103 | 330 | 38,174.39 | 4.91 |
| DTZ79_24g03470 | AeHSFA6b | Chr24 | 4,741,552 | 4,742,952 | 293 | 34,385.11 | 6.37 |
| DTZ79_24g13410 | AeHSFA6c | Chr24 | 20,568,827 | 20,571,324 | 261 | 30,050.22 | 7.53 |
| DTZ79_17g08930 | AeHSFA7a | Chr17 | 16,311,605 | 16,314,121 | 353 | 40,323.58 | 6.76 |
| DTZ79_23g01840 | AeHSFA7b | Chr23 | 1,719,685 | 1,725,774 | 393 | 45,040.76 | 5.97 |
| DTZ79_25g01580 | AeHSFA8a | Chr25 | 7,169,500 | 7,175,687 | 336 | 38,474.7 | 5.11 |
| DTZ79_27g07960 | AeHSFA8b | Chr27 | 9,529,778 | 9,535,497 | 266 | 30,879.35 | 5.970 |
| DTZ79_05g10500 | AeHSFB1a | Chr05 | 19,792,047 | 19,795,733 | 290 | 32,502.43 | 5.84 |
| DTZ79_09g05760 | AeHSFB1b | Chr09 | 7,085,426 | 7,087,756 | 230 | 25,915.03 | 6.72 |
| DTZ79_15g12130 | AeHSFB1c | Chr15 | 16,819,805 | 16,833,941 | 730 | 83,846.59 | 8.83 |
| DTZ79_27g06080 | AeHSFB1d | Chr27 | 6,538,958 | 6,543,540 | 273 | 30,221.65 | 5.16 |
| DTZ79_03g10230 | AeHSFB2a | Chr03 | 10,487,781 | 10,488,831 | 318 | 35,100.92 | 4.54 |
| DTZ79_18g03510 | AeHSFB2b | Chr18 | 5,178,304 | 5,180,532 | 301 | 33,379.45 | 6.3 |
| DTZ79_24g11010 | AeHSFB2c | Chr24 | 17,046,072 | 17,047,945 | 298 | 33,423.12 | 8.23 |
| DTZ79_12g03600 | AeHSFB3a | Chr12 | 4,511,163 | 4,513,746 | 241 | 27,528.89 | 7.55 |
| DTZ79_20g10710 | AeHSFB3b | Chr20 | 16,559,515 | 16,563,171 | 239 | 27,835.34 | 6.02 |
| DTZ79_04g03520 | AeHSFB4a | Chr04 | 3,612,012 | 3,614,202 | 542 | 60,737.75 | 5.93 |
| DTZ79_10g10590 | AeHSFB4b | Chr10 | 20,942,779 | 20,944,104 | 312 | 35,352.01 | 5.65 |
| DTZ79_11g01990 | AeHSFB4c | Chr11 | 2,045,916 | 2,047,860 | 307 | 34,740.27 | 4.99 |
| DTZ79_18g08770 | AeHSFB4d | Chr18 | 17,220,840 | 17,222,490 | 187 | 21,116.75 | 6.72 |
| DTZ79_12g02240 | AeHSFC1a | Chr12 | 2,633,942 | 2,635,609 | 282 | 32,163.46 | 7.69 |
| DTZ79_20g11770 | AeHSFC1b | Chr20 | 18,439,575 | 18,444,574 | 276 | 31,061.17 | 5.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, C.; Liu, Y.; Yang, Z.; Xu, J.; Ouyang, Z.; Yang, J.; Wang, S. Genome-Wide Identification of HSF Gene Family in Kiwifruit and the Function of AeHSFA2b in Salt Tolerance. Int. J. Mol. Sci. 2023, 24, 15638. https://doi.org/10.3390/ijms242115638
Ling C, Liu Y, Yang Z, Xu J, Ouyang Z, Yang J, Wang S. Genome-Wide Identification of HSF Gene Family in Kiwifruit and the Function of AeHSFA2b in Salt Tolerance. International Journal of Molecular Sciences. 2023; 24(21):15638. https://doi.org/10.3390/ijms242115638
Chicago/Turabian StyleLing, Chengcheng, Yunyan Liu, Zuchi Yang, Jiale Xu, Zhiyin Ouyang, Jun Yang, and Songhu Wang. 2023. "Genome-Wide Identification of HSF Gene Family in Kiwifruit and the Function of AeHSFA2b in Salt Tolerance" International Journal of Molecular Sciences 24, no. 21: 15638. https://doi.org/10.3390/ijms242115638