
TOE1/TOE2 Interacting with GIS to Control Trichome Development in Arabidopsis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
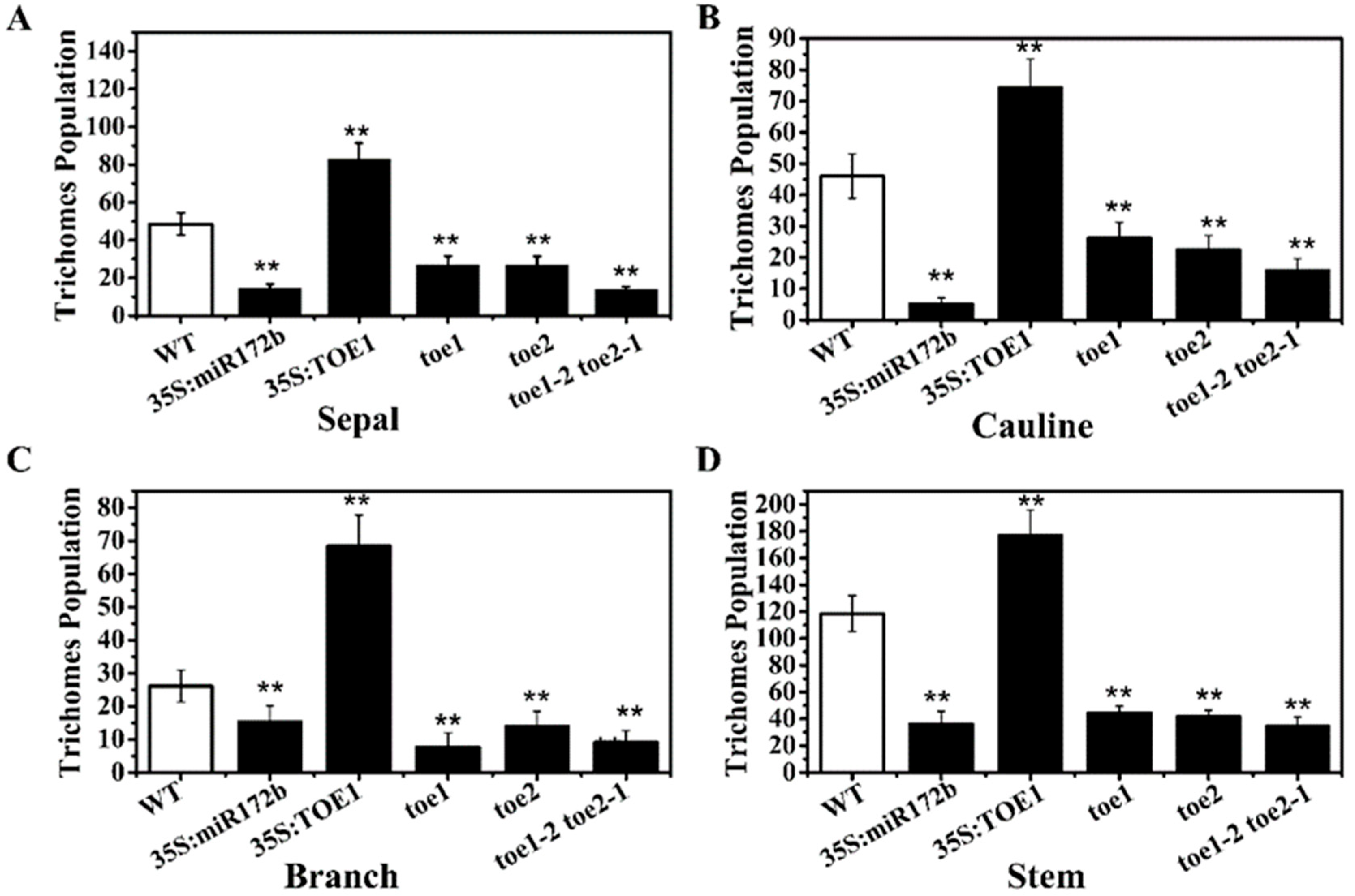
2.1. TOE1/TOE2 Are Involved in Main-Stem Inflorescence Trichome Initiation in Arabidopsis
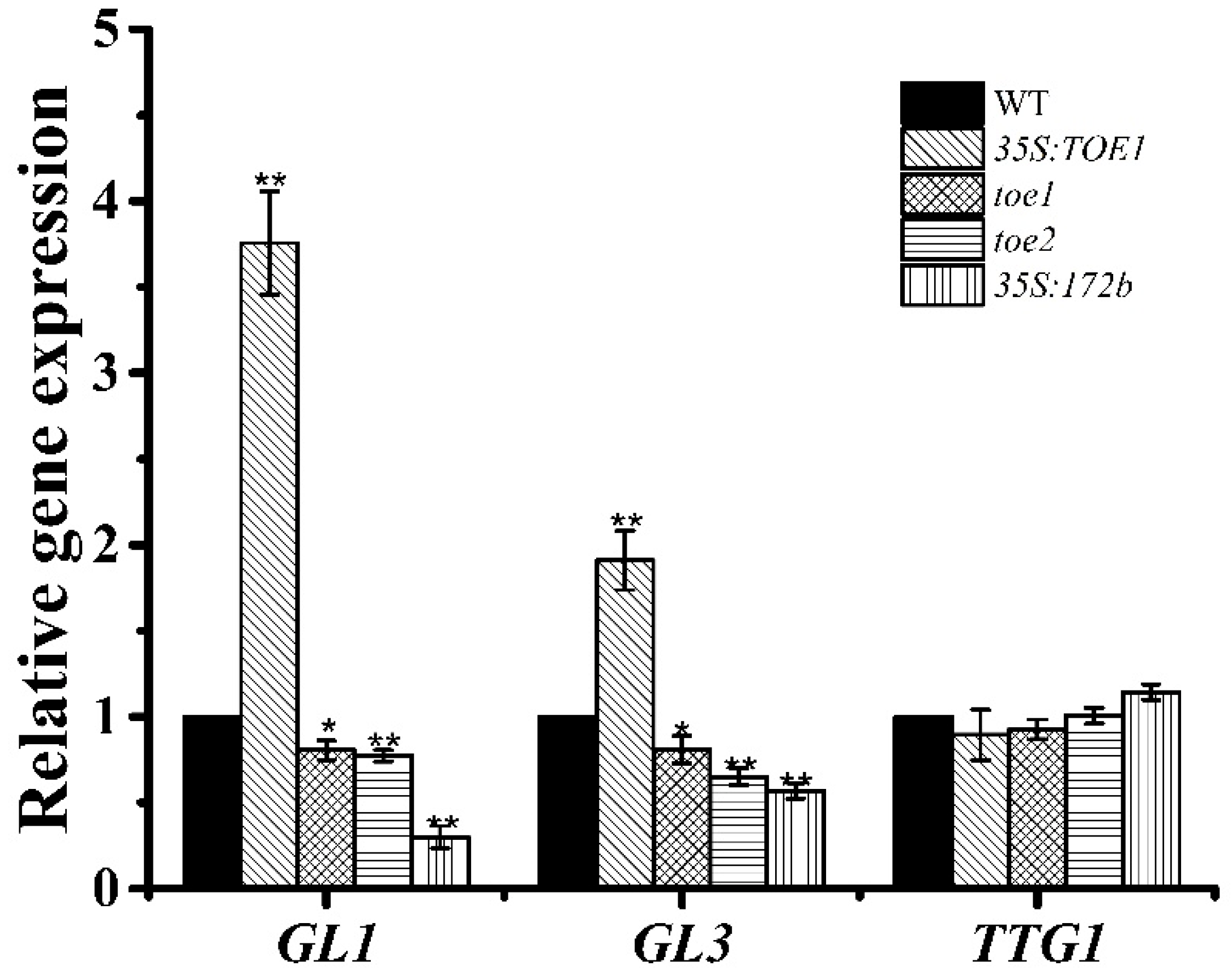
2.2. TOE1/TOE2 Act Upstream of GL1 and GL3
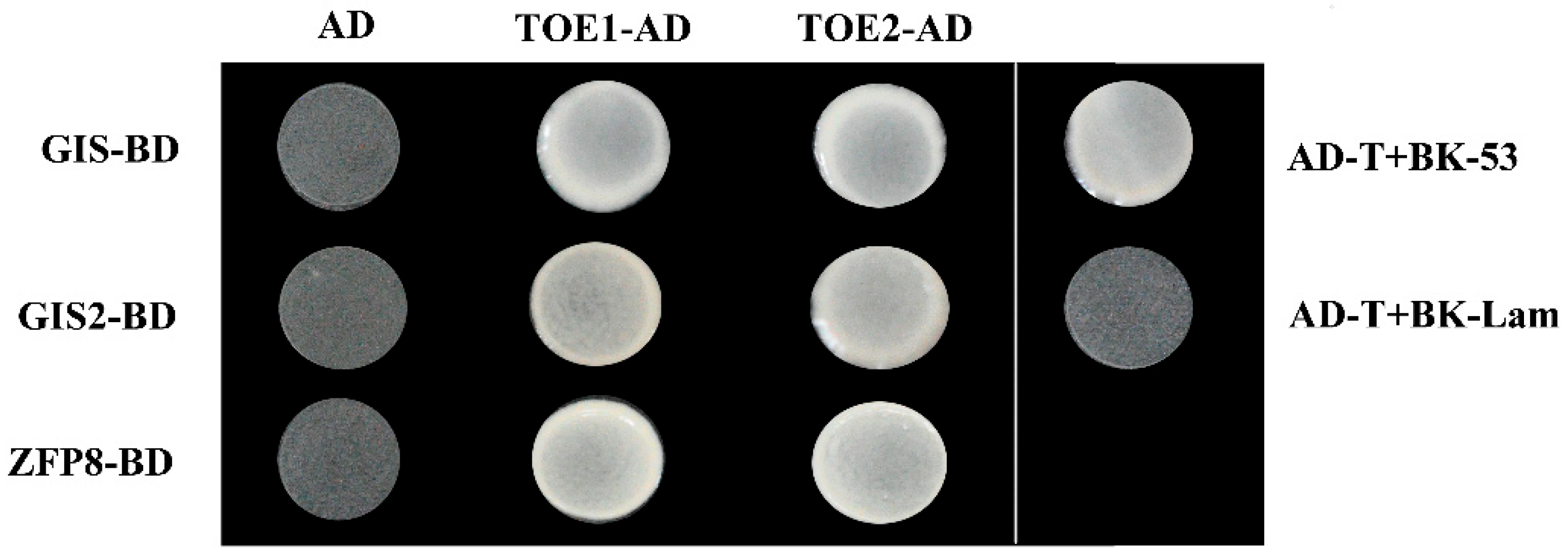
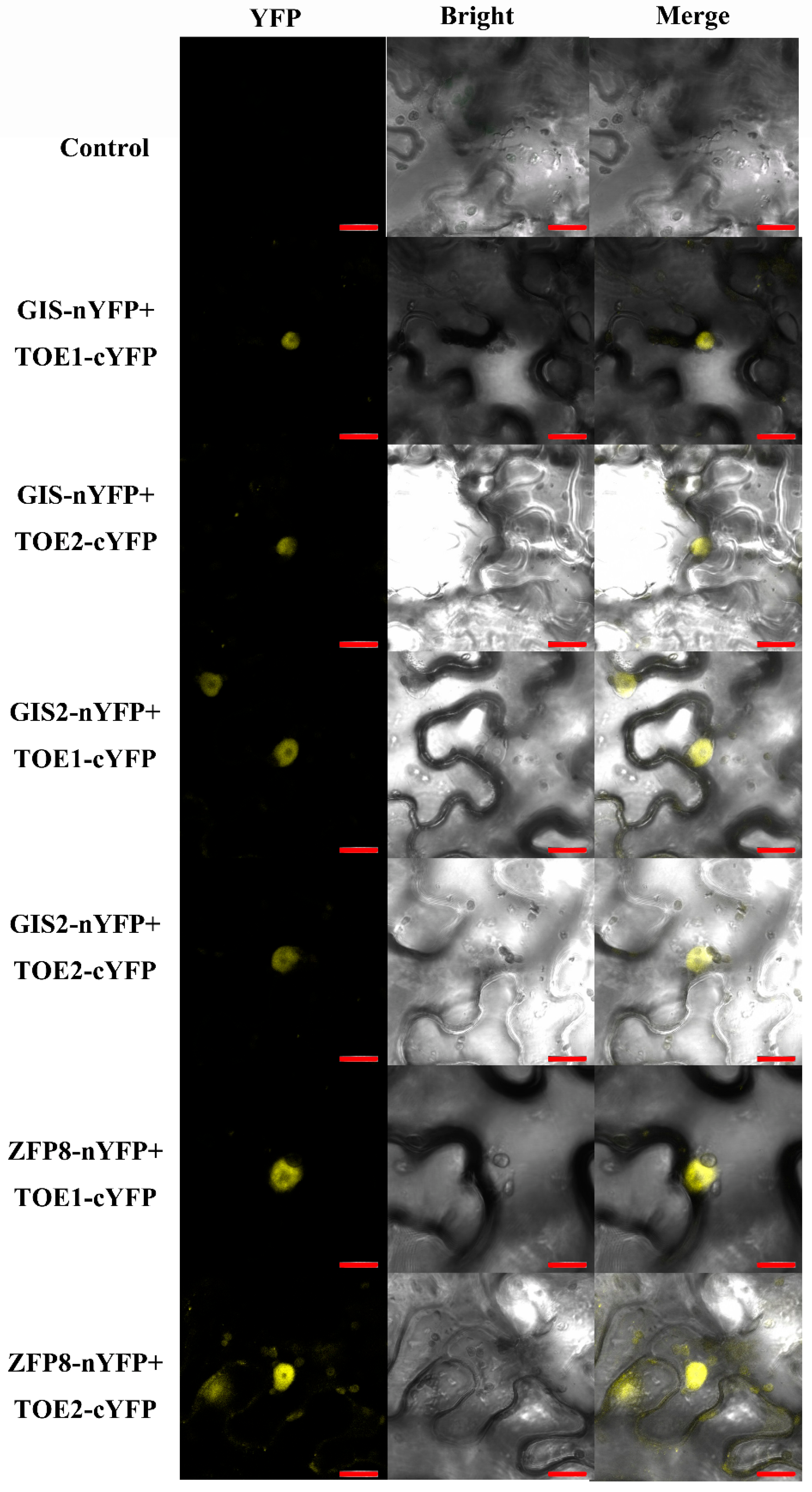
2.3. TOE1/TOE2 Interact with GIS
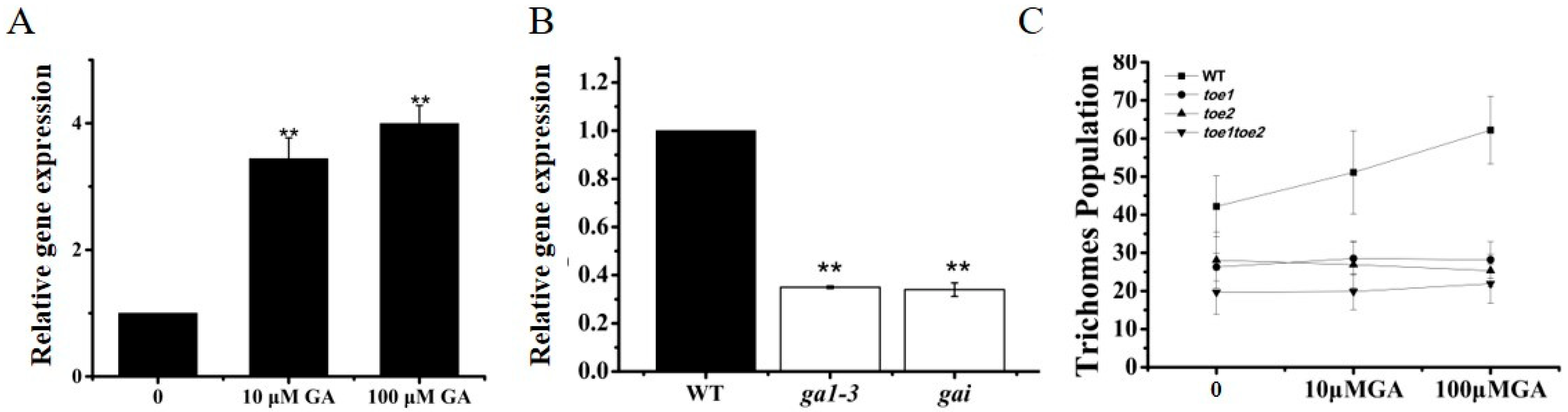
2.4. TOE1 Regulates Trichome Initiation through GA Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. RNA Extraction and Quantitative Real-Time PCR (qPCR)
4.3. Gene Cloning and Vector Construction
4.4. Trichome Production Analysis
4.5. Yeast Two-Hybridization Assay
4.6. Bimolecular Fluorescent Complementation (BiFC) Assay
4.7. GA Treatment
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chalvin, C.; Drevensek, S.; Dron, M.; Bendahmane, A.; Boualem, A. Genetic Control of Glandular Trichome Development. Trends Plant Sci. 2020, 25, 477–487. [Google Scholar] [CrossRef]
- Li, S.; Tosens, T.; Harley, P.C.; Jiang, Y.F.; Kanagendran, A.; Grosberg, M.; Jaamets, K.; Niinemets, U. Glandular trichomes as a barrier against atmospheric oxidative stress: Relationships with ozone uptake, leaf damage, and emission of LOX products across a diverse set of species. Plant Cell Environ. 2018, 41, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Pattanaik, S.; Patra, B.; Singh, S.K.; Yuan, L. An overview of the gene regulatory network controlling trichome development in the model plant, Arabidopsis. Front. Plant Sci. 2014, 5, 259. [Google Scholar] [CrossRef] [Green Version]
- Werker, E. Trichome diversity and development. Adv. Bot. Res. 2000, 31, 1–35. [Google Scholar] [CrossRef]
- Yang, C.; Ye, Z. Trichomes as models for studying plant cell differentiation. Cell. Mol. Life Sci. 2013, 70, 1937–1948. [Google Scholar] [CrossRef] [PubMed]
- Hulskamp, M.; Misra, S.; Jurgens, G. Genetic dissection of trichome cell development in Arabidopsis. Cell 1994, 76, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Luo, Y.; Zhang, W.; Jian, W.; Zhang, L.; Gao, X.; Hu, X.; Yuan, Y.; Wu, M.; Xu, X.; et al. SlMYB75-centred transcriptional cascade regulates trichome formation and sesquiterpene accumulation on tomato. J. Exp. Bot. 2021, 72, 3806–3820. [Google Scholar] [CrossRef]
- Liu, S.; Fan, L.; Liu, Z.; Yang, X.; Zhang, Z.; Duan, Z.; Liang, Q.; Imran, M.; Zhang, M.; Tian, Z. A Pd1-Ps-P1 Feedback Loop Controls Pubescence Density in Soybean. Mol. Plant 2020, 13, 1768–1783. [Google Scholar] [CrossRef]
- Andama, J.B.; Mujiono, K.; Hojo, Y.; Shinya, T.; Galis, I. Nonglandular silicified trichomes are essential for rice defense against chewing herbivores. Plant Cell Environ. 2020, 43, 2019–2032. [Google Scholar] [CrossRef]
- Benatto, A.; Mogor, A.F.; Penteado, S.C.; Pereira, L.S.; Salas, F.J.S.; Zawadneak, M.A.C. Influence of Trichomes in Strawberry Cultivars on the Feeding Behavior of Chaetosiphon fragaefolii (Cockerell) (Hemiptera: Aphididae). Neotrop. Entomol. 2018, 47, 569–576. [Google Scholar] [CrossRef]
- Feng, Z.; Bartholomew, E.S.; Liu, Z.; Cui, Y.; Dong, Y.; Li, S.; Wu, H.; Ren, H.; Liu, X. Glandular trichomes: New focus on horticultural crops. Hortic. Res. 2021, 8, 158. [Google Scholar] [CrossRef]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant Glandular Trichomes: Natural Cell Factories of High Biotechnological Interest[OPEN]. Plant Physiol. 2017, 175, 6–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, N.; Yang, L.; Luo, S.; Wang, X.T.; Wang, W.; Cheng, Y.X.; Tian, H.N.; Zheng, K.J.; Cai, L.; Wang, S.C. Genetic evidence suggests that GIS functions downstream of TCL1 to regulate trichome formation in Arabidopsis. BMC Plant Biology 2018, 18, 63. [Google Scholar] [CrossRef] [PubMed]
- Matias-Hernandez, L.; Aguilar-Jaramillo, A.E.; Cigliano, R.A.; Sanseverino, W.; Pelaz, S. Flowering and trichome development share hormonal and transcription factor regulation. J. Exp. Bot. 2016, 67, 1209–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jindal, S.; Longchar, B.; Singh, A.; Gupta, V. Promoters of AaGL2 and AaMIXTA-Like1 genes of Artemisia annua direct reporter gene expression in glandular and non-glandular trichomes. Plant Signal. Behav. 2015, 10, e1087629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. TRANSPARENT TESTA GLABRA2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, D.B.; Lloyd, A.M.; Marks, M.D. Progress in the molecular genetic analysis of trichome initiation and morphogenesis in Arabidopsis. Trends Plant Sci. 2000, 5, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Rerie, W.G.; Feldmann, K.A.; Marks, M.D. The GLABRA2 gene encodes a homeo domain protein required for normal trichome development in Arabidopsis. Genes Dev. 1994, 8, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Hyoung, S.; So, W.M.; Shin, J.S. The novel transcription factor TRP interacts with ZFP5, a trichome initiation-related transcription factor, and negatively regulates trichome initiation through gibberellic acid signaling. Plant Mol. Biol. 2018, 96, 315–326. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, D.; Hu, R.; Hua, C.; Ali, I.; Zhang, A.; Liu, B.; Wu, M.; Huang, L.; Gan, Y. AtGIS, a C2H2 zinc-finger transcription factor from Arabidopsis regulates glandular trichome development through GA signaling in tobacco. Biochem. Biophys. Res. Commun. 2017, 483, 209–215. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, A.; Zhou, Z.; Zhao, Y.; Yan, A.; Bao, S.; Yu, H.; Gan, Y. GLABROUS INFLORESCENCE STEMS3 (GIS3) regulates trichome initiation and development in Arabidopsis. New Phytol. 2015, 206, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Sun, L.; Zhao, Y.; An, L.; Yan, A.; Meng, X.; Gan, Y. Zinc Finger Protein 6 (ZFP6) regulates trichome initiation by integrating gibberellin and cytokinin signaling in Arabidopsis thaliana. New Phytol. 2013, 198, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; An, L.; Sun, L.; Gan, Y. ZFP5 encodes a functionally equivalent GIS protein to control trichome initiation. Plant Signal. Behav. 2012, 7, 28–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Y.; Liu, C.; Yu, H.; Broun, P. Integration of cytokinin and gibberellin signalling by Arabidopsis transcription factors GIS, ZFP8 and GIS2 in the regulation of epidermal cell fate. Development 2007, 134, 2073–2081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, B.; Chang, J.; Xu, Z.; Han, X.; Xu, M.; Yang, M.; Yang, C.; Ye, Z.; Wu, S. HOMEODOMAIN PROTEIN8 mediates jasmonate-triggered trichome elongation in tomato. New Phytol. 2021, 230, 1063–1077. [Google Scholar] [CrossRef]
- Xia, X.C.; Hu, Q.Q.; Li, W.; Chen, Y.; Han, L.H.; Tao, M.; Wu, W.Y.; Li, X.B.; Huang, G.Q. Cotton (Gossypium hirsutum) JAZ3 and SLR1 function in jasmonate and gibberellin mediated epidermal cell differentiation and elongation. Plant Cell Tissue Organ Cult. 2018, 133, 249–262. [Google Scholar] [CrossRef]
- Inthima, P.; Nakano, M.; Otani, M.; Niki, T.; Nishijima, T.; Koshioka, M.; Supaibulwatana, K. Overexpression of the gibberellin 20-oxidase gene from Torenia fournieri resulted in modified trichome formation and terpenoid metabolities of Artemisia annua L. Plant Cell Tissue Organ Cult. 2017, 129, 223–236. [Google Scholar] [CrossRef]
- Qi, T.; Huang, H.; Wu, D.; Yan, J.; Qi, Y.; Song, S.; Xie, D. ArabidopsisDELLA and JAZ Proteins Bind the WD-Repeat/bHLH/MYB Complex to Modulate Gibberellin and Jasmonate Signaling Synergy. Plant Cell 2014, 26, 1118–1133. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Sano, R.; Wada, T.; Takabayashi, J.; Okada, K. Jasmonic acid control of GLABRA3 links inducible defense and trichome patterning in Arabidopsis. Development 2009, 136, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhou, C.M.; Mai, Y.X.; Li, L.Z.; Gao, J.; Shang, G.D.; Lian, H.; Han, L.; Zhang, T.Q.; Tang, H.B.; et al. A spatiotemporally regulated transcriptional complex underlies heteroblastic development of leaf hairs in Arabidopsis thaliana. EMBO J. 2019, 38, e100063. [Google Scholar] [CrossRef]
- Diaz-Manzano, F.E.; Cabrera, J.; Ripoll, J.J.; Del Olmo, I.; Andres, M.F.; Silva, A.C.; Barcala, M.; Sanchez, M.; Ruiz-Ferrer, V.; de Almeida-Engler, J.; et al. A role for the gene regulatory module microRNA172/TARGET of Early Activation Tagged 1/FLOWERING LOCUS T (miRNA172/TOE1/FT) in the feeding sites induced by Meloidogyne javanica in Arabidopsis thaliana. New Phytol. 2018, 217, 813–827. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.L.; Wang, L.; Zeng, L.P.; Zhang, C.; Ma, H. Arabidopsis TOE proteins convey a photoperiodic signal to antagonize CONSTANS and regulate flowering time. Genes Dev. 2015, 29, 975–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, M.; Munne-Bosch, S. Ethylene Response Factors: A Key Regulatory Hub in Hormone and Stress Signaling. Plant Physiol. 2015, 169, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Licausi, F.; Ohme-Takagi, M.; Perata, P. APETALA2/Ethylene Responsive Factor (AP2/ERF) transcription factors: Mediators of stress responses and developmental programs. New Phytol. 2013, 199, 639–649. [Google Scholar] [CrossRef]
- Nakano, T.; Suzuki, K.; Fujimura, T.; Shinshi, H. Genome-wide analysis of the ERF gene family in Arabidopsis and rice. Plant Physiol. 2006, 140, 411–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.H.; Helliwell, C.A. Regulation of flowering time and floral patterning by miR172. J. Exp. Bot. 2011, 62, 487–495. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.; Won, S.Y.; Suh, S.C.; Kim, H.; Wing, R.; Jeong, Y.; Hwang, I.; Kim, M. The barley ERF-type transcription factor HvRAF confers enhanced pathogen resistance and salt tolerance in Arabidopsis. Planta 2007, 225, 575–588. [Google Scholar] [CrossRef]
- O’Maoileidigh, D.S.; Stewart, D.; Zheng, B.; Coupland, G.; Wellmer, F. Floral homeotic proteins modulate the genetic program for leaf development to suppress trichome formation in flowers. Development 2018, 145, dev157784. [Google Scholar] [CrossRef] [Green Version]
- Daviere, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Yu, H.; Peng, J.; Broun, P. Genetic and molecular regulation by DELLA proteins of trichome development in Arabidopsis. Plant Physiol. 2007, 145, 1031–1042. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.Q.; Lian, H.; Tang, H.; Dolezal, K.; Zhou, C.M.; Yu, S.; Chen, J.H.; Chen, Q.; Liu, H.; Ljung, K.; et al. An intrinsic microRNA timer regulates progressive decline in shoot regenerative capacity in plants. Plant Cell 2015, 27, 349–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, J.C.; Sussex, I.M. Differential regulation of trichome formation on the adaxial and abaxial leaf surfaces by Gibberellins and photoperiod in Arabidopsis thaliana (L) Heynh. Plant Physiol. 1996, 111, 1321–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Y.; Kumimoto, R.; Liu, C.; Ratcliffe, O.; Yu, H.; Broun, P. Glabrous Inflorescence Stems modulates the regulation by gibberellins of epidermal differentiation and shoot maturation in Arabidopsis. Plant Cell 2006, 18, 1383–1395. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.; Cai, W.J.; Wang, S.; Shan, C.M.; Wang, L.J.; Chen, X.Y. Temporal Control of Trichome Distribution by MicroRNA156-Targeted SPL Genes in Arabidopsis thaliana. Plant Cell Online 2010, 22, 2322–2335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; An, L.; Sun, L.; Zhu, S.; Xi, W.; Broun, P.; Yu, H.; Gan, Y. Zinc finger protein5 is required for the control of trichome initiation by acting upstream of zinc finger protein8 in Arabidopsis. Plant Physiol. 2011, 157, 673–682. [Google Scholar] [CrossRef] [Green Version]
- An, L.; Zhou, Z.; Yan, A.; Gan, Y. Progress on trichome development regulated by phytohormone signaling. Plant Signal. Behav. 2011, 6, 1959–1962. [Google Scholar] [CrossRef] [Green Version]
- Gan, Y.; Filleur, S.; Rahman, A.; Gotensparre, S.; Forde, B.G. Nutritional regulation of ANR1 and other root-expressed MADS-box genes in Arabidopsis thaliana. Planta 2005, 222, 730–742. [Google Scholar] [CrossRef]
- Bao, S.; An, L.; Su, S.; Zhou, Z.; Gan, Y. Expression patterns of nitrate, phosphate, and sulfate transporters in Arabidopsis roots exposed to different nutritional regimes. Botany 2011, 89, 647–653. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, D.; Hua, C.; Yan, A.; Liu, B.; Wu, M.; Liu, Y.; Huang, L.; Ali, I.; Gan, Y. The Arabidopsis Gene zinc finger protein 3(ZFP3) Is Involved in Salt Stress and Osmotic Stress Response. PLoS ONE 2016, 11, e0168367. [Google Scholar] [CrossRef] [Green Version]
- Wakeel, A.; Ali, I.; Upreti, S.; Azizullah, A.; Liu, B.H.; Khan, A.R.; Huang, L.L.; Wu, M.J.; Gan, Y.B. Ethylene mediates dichromate-induced inhibition of primary root growth by altering AUX1 expression and auxin accumulation in Arabidopsis thaliana. Plant Cell Environ. 2018, 41, 1453–1467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huang, L.L.; Yan, A.; Liu, Y.H.; Liu, B.H.; Yu, C.Y.; Zhang, A.D.; Schiefelbein, J.; Gan, Y.B. Multiple phytohormones promote root hair elongation by regulating a similar set of genes in the root epidermis in Arabidopsis. J. Exp. Bot. 2016, 67, 6363–6372. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.Y.; Liu, Y.H.; Zhang, A.D.; Su, S.; Yan, A.; Huang, L.; Ali, I.; Liu, Y.; Forde, B.G.; Gan, Y.B. MADS-box Transcription Factor OsMADS25 Regulates Root Development through Affection of Nitrate Accumulation in Rice. PLoS ONE 2015, 10, e0135196. [Google Scholar] [CrossRef]
- Sun, W.; Gao, D.; Xiong, Y.; Tang, X.; Xiao, X.; Wang, C.; Yu, S. Hairy Leaf 6, an AP2/ERF Transcription Factor, Interacts with OsWOX3B and Regulates Trichome Formation in Rice. Mol. Plant 2017, 10, 1417–1433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, T.X.; Chen, M.H.; Shen, Q.; Li, L.; Fu, X.Q.; Pan, Q.F.; Tang, Y.L.; Shi, P.; Lv, Z.Y.; Jiang, W.M.; et al. HOMEODOMAIN PROTEIN 1 is required for jasmonate-mediated glandular trichome initiation in Artemisia annua. New Phytol. 2017, 213, 1145–1155. [Google Scholar] [CrossRef]
- Patra, B.; Pattanaik, S.; Yuan, L. Ubiquitin protein ligase 3 mediates the proteasomal degradation of GLABROUS 3 and ENHANCER OF GLABROUS 3, regulators of trichome development and flavonoid biosynthesis in Arabidopsis. Plant J. 2013, 74, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-Domain Proteins Interact with the WD-Repeat/bHLH/MYB Complexes to Regulate Jasmonate-Mediated Anthocyanin Accumulation and Trichome Initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Liu, D.D.; Khan, A.R.; Liu, B.H.; Wu, M.J.; Huang, L.L.; Wu, J.Y.; Song, G.; Ni, H.W.; Ying, H.M.; et al. NbGIS regulates glandular trichome initiation through GA signaling in tobacco. Plant Mol. Biol. 2018, 98, 153–167. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Yang, S.; Khan, A.R.; Gan, Y. TOE1/TOE2 Interacting with GIS to Control Trichome Development in Arabidopsis. Int. J. Mol. Sci. 2023, 24, 6698. https://doi.org/10.3390/ijms24076698
Liu Y, Yang S, Khan AR, Gan Y. TOE1/TOE2 Interacting with GIS to Control Trichome Development in Arabidopsis. International Journal of Molecular Sciences. 2023; 24(7):6698. https://doi.org/10.3390/ijms24076698
Chicago/Turabian StyleLiu, Yihua, Shuaiqi Yang, Ali Raza Khan, and Yinbo Gan. 2023. "TOE1/TOE2 Interacting with GIS to Control Trichome Development in Arabidopsis" International Journal of Molecular Sciences 24, no. 7: 6698. https://doi.org/10.3390/ijms24076698