
Roles of AGD2a in Plant Development and Microbial Interactions of Lotus japonicus


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
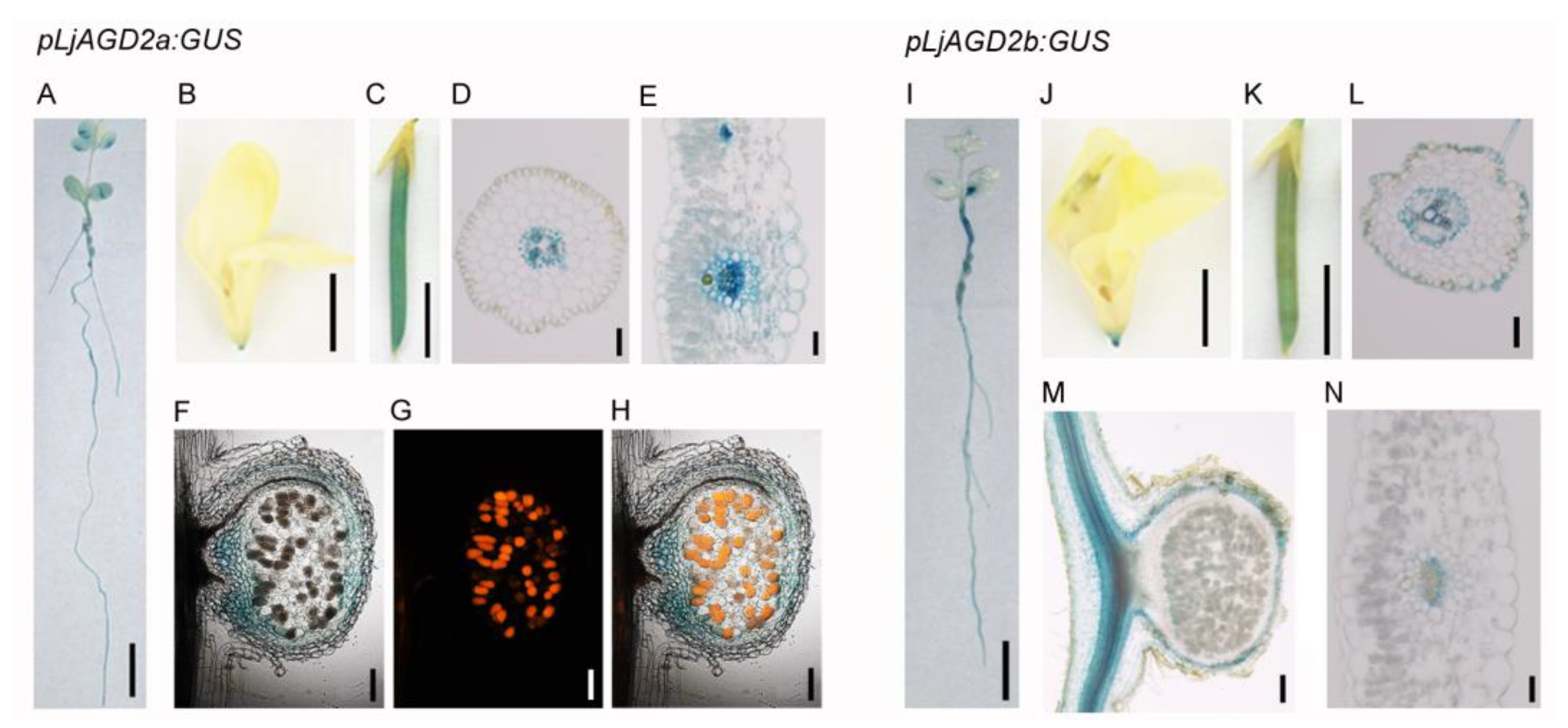
2.1. Expression Pattern of LjAGD2a in L. japonicus
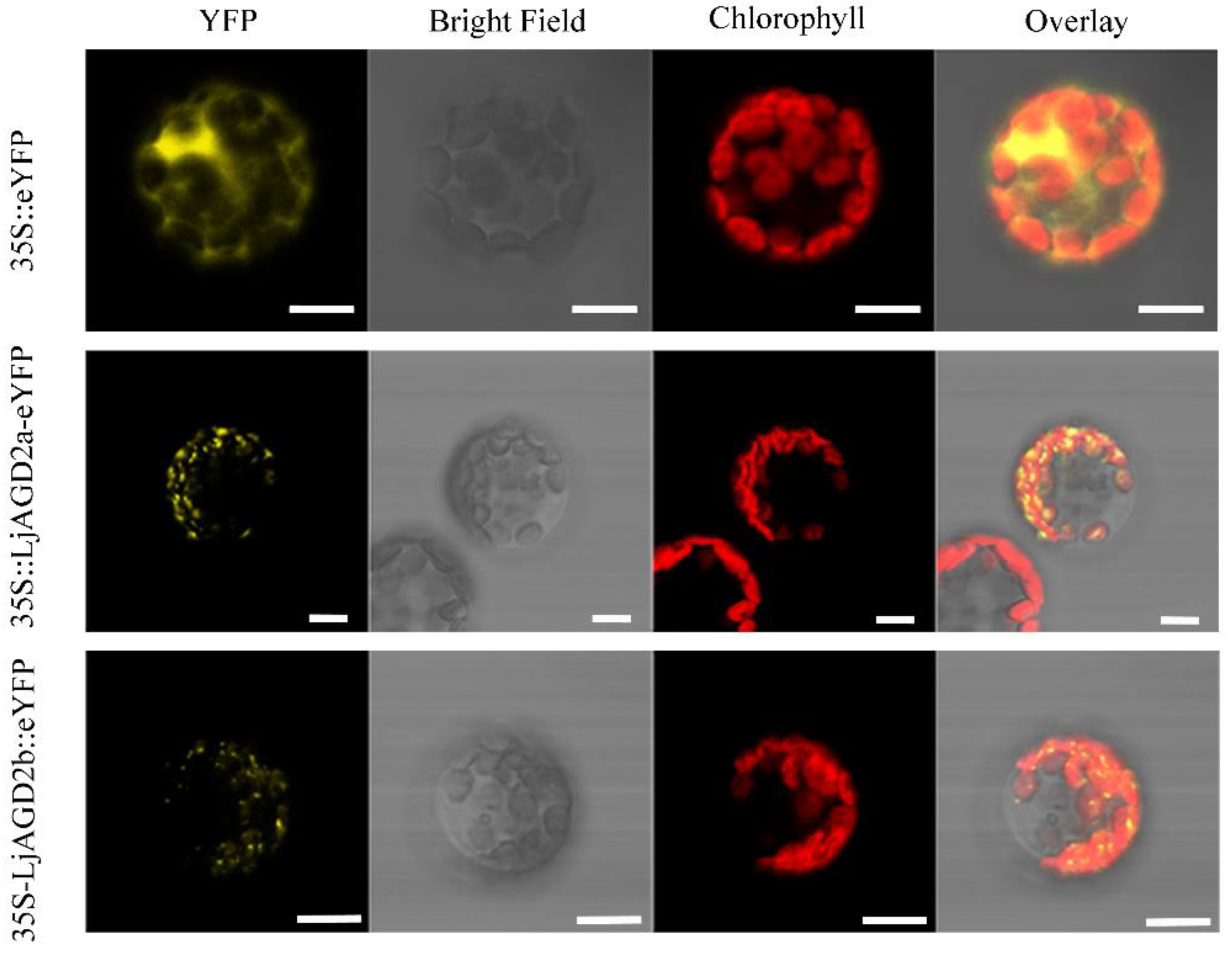
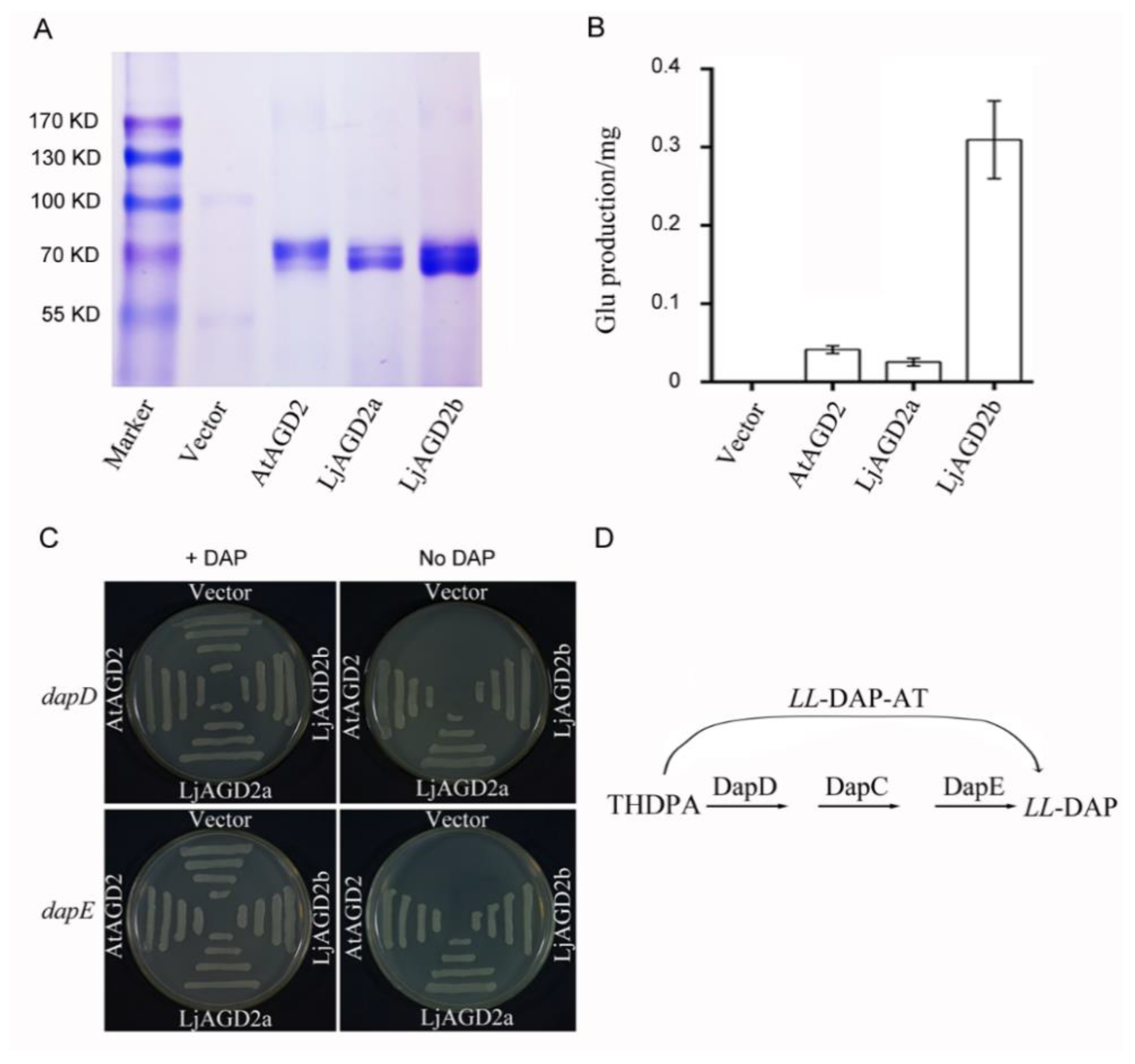
2.2. LjAGD2s Are Localized in Chloroplasts and Function as LL-Diaminopimelate Aminotransferases
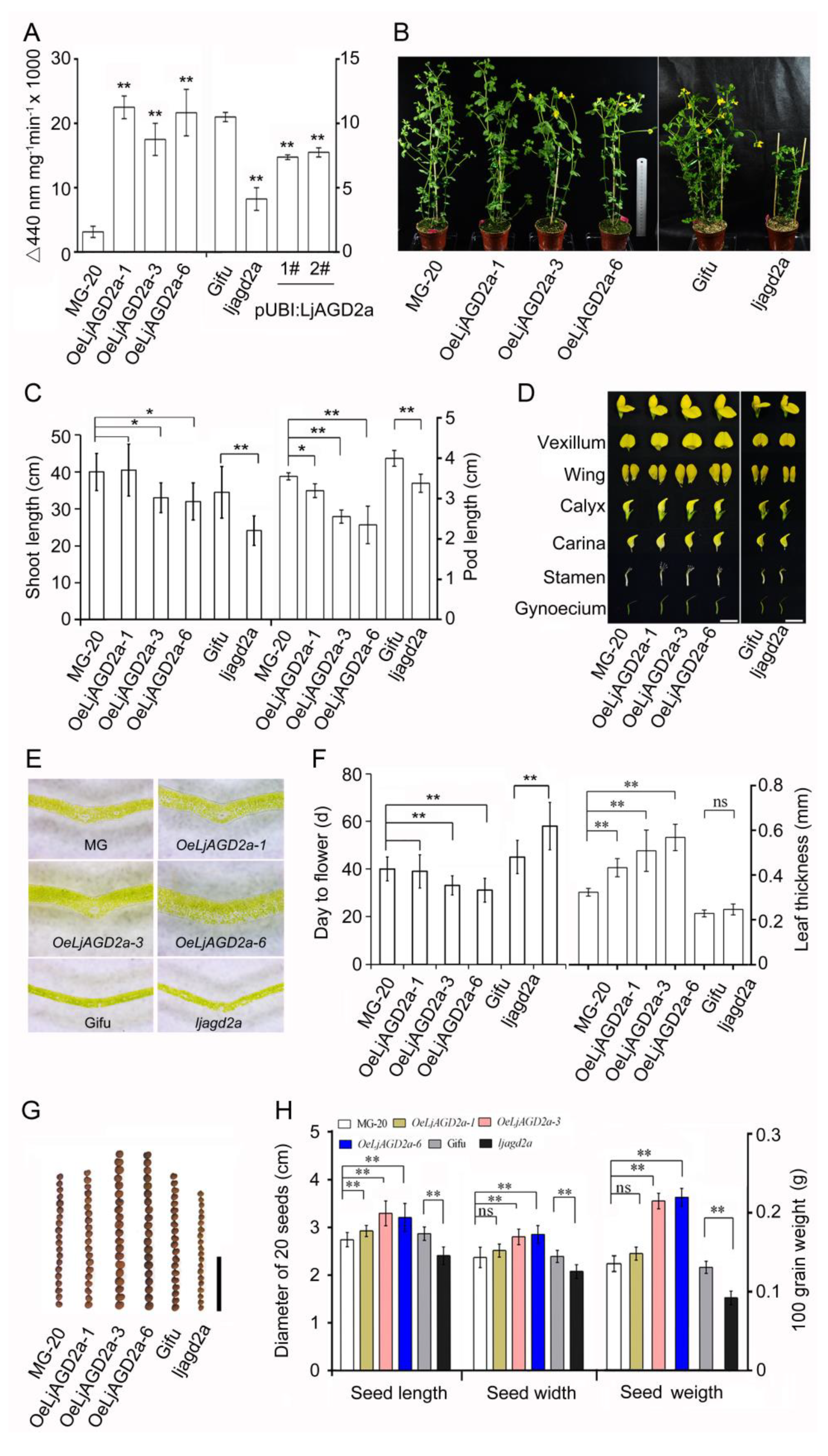
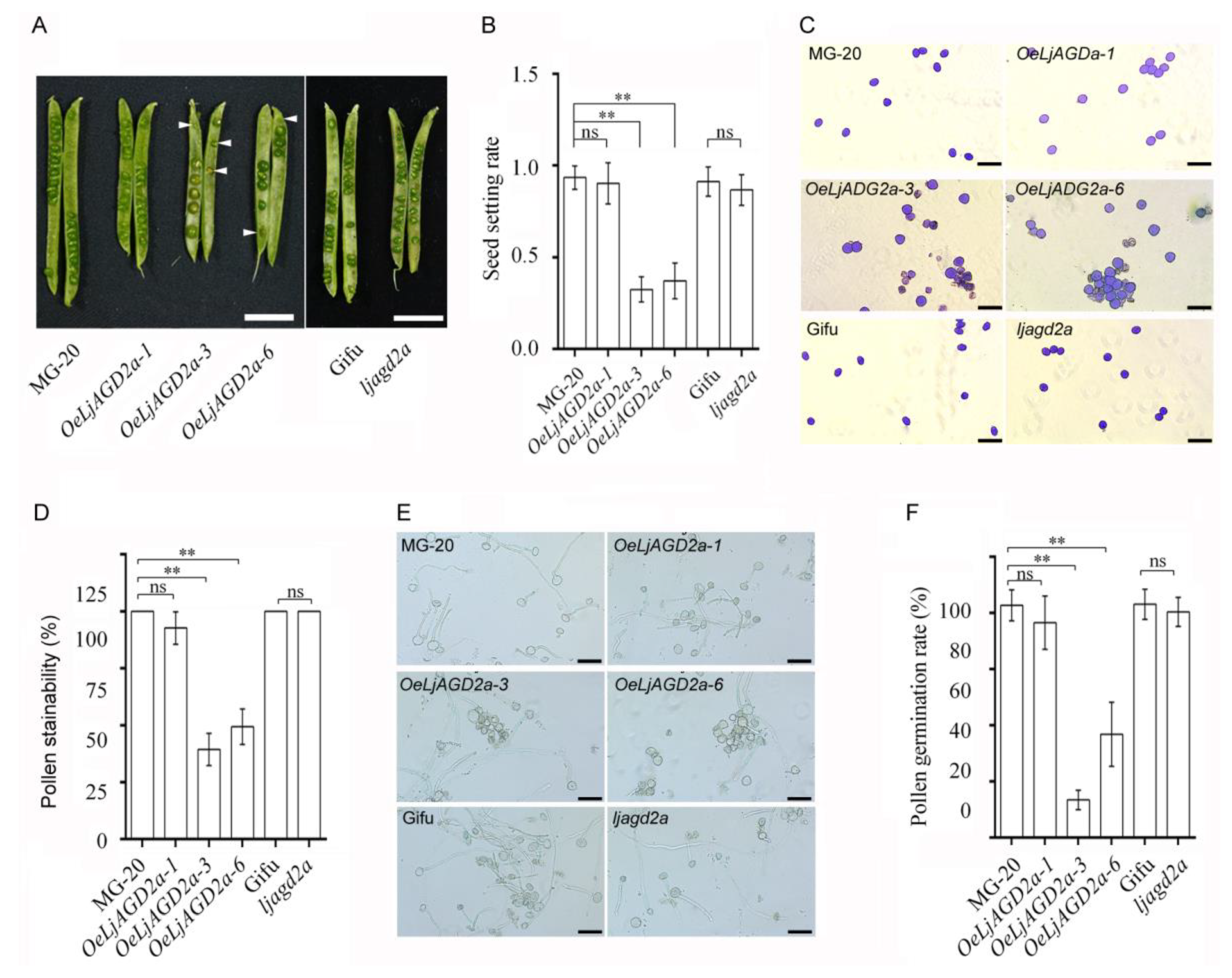
2.3. LjAGD2a Affects Plant Morphology and Development
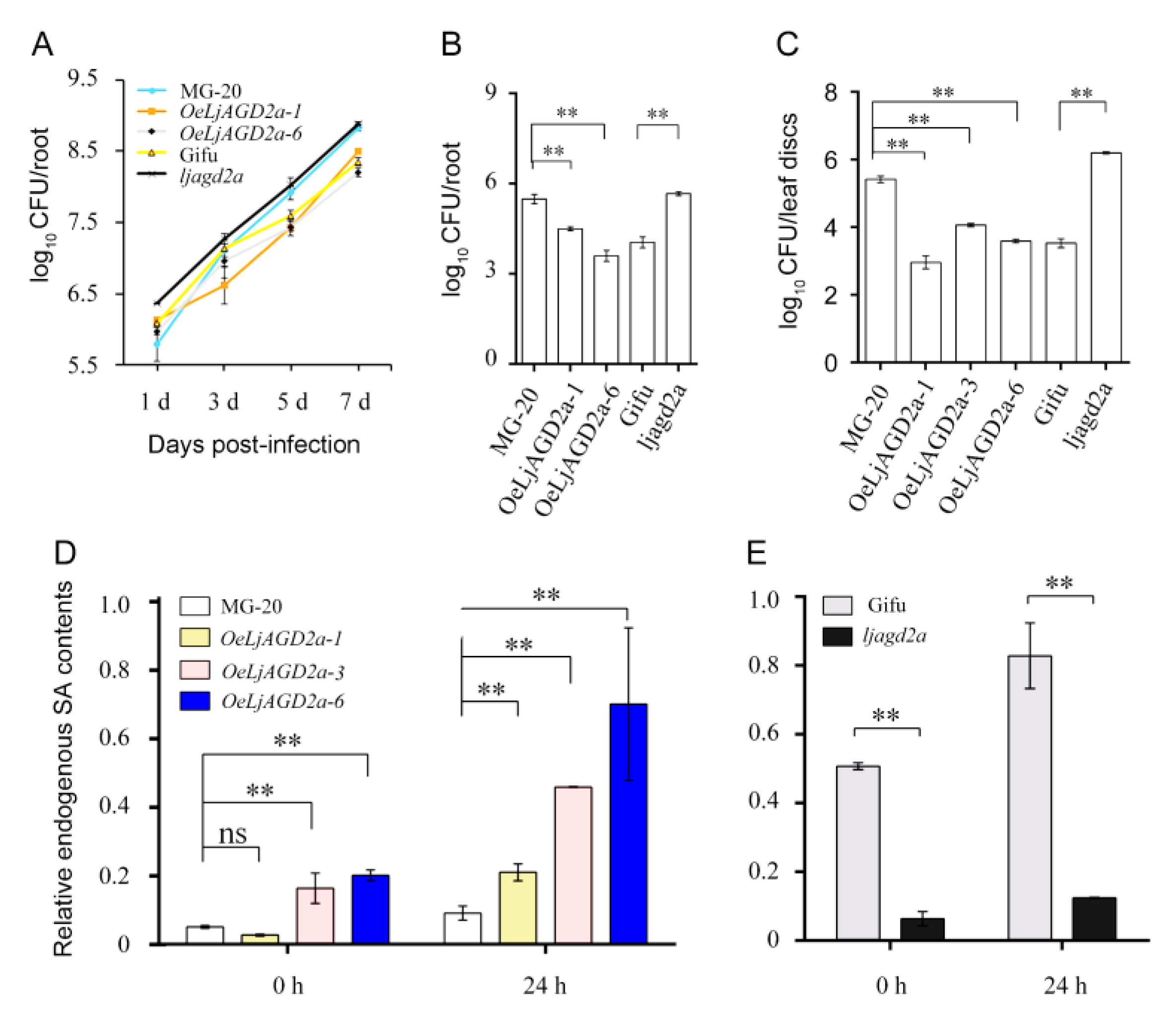
2.4. Alteration of LjAGD2a Expression in L. japonicus Changed Resistance to the Bacterial Pathogen R. Solanacearum
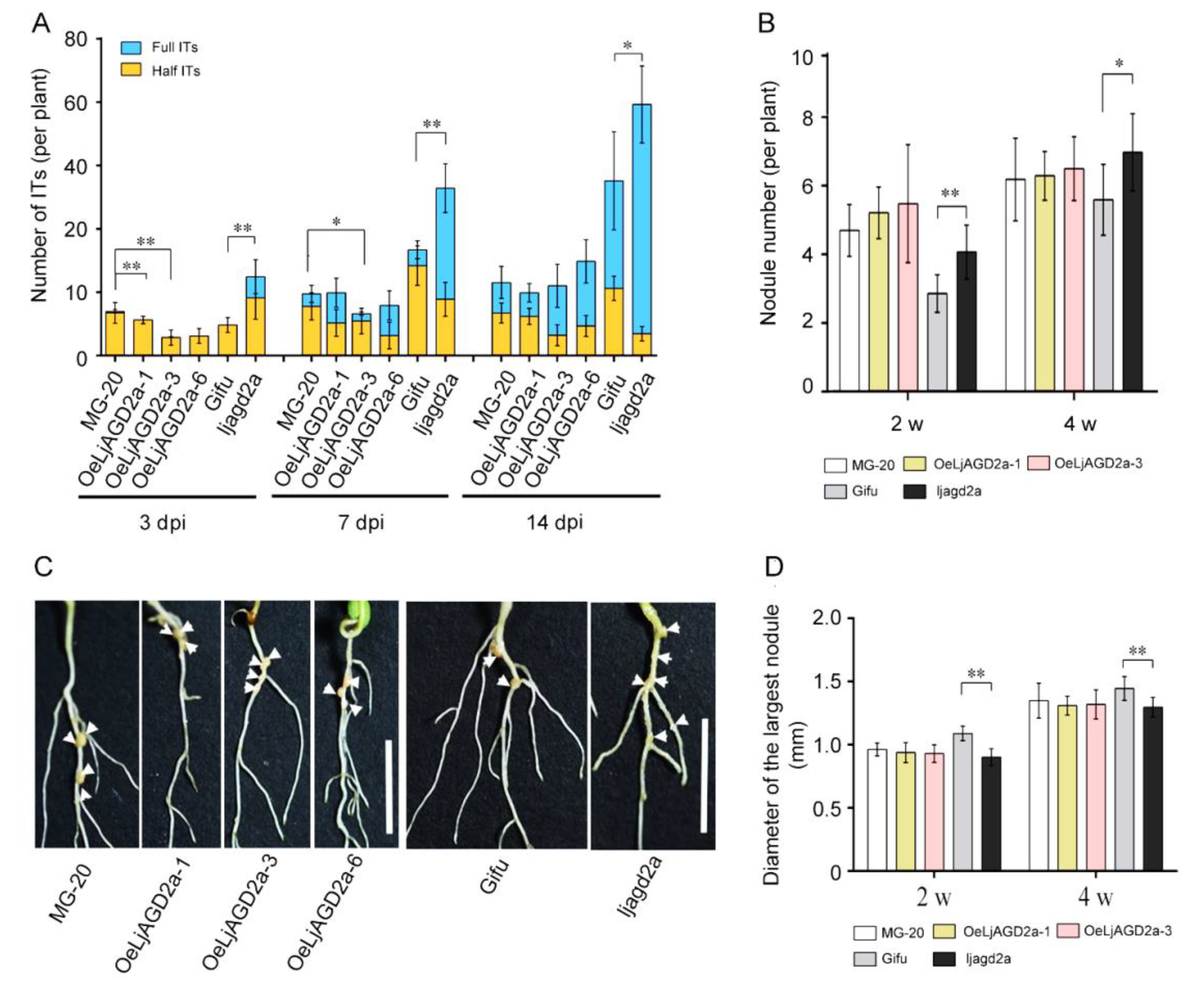
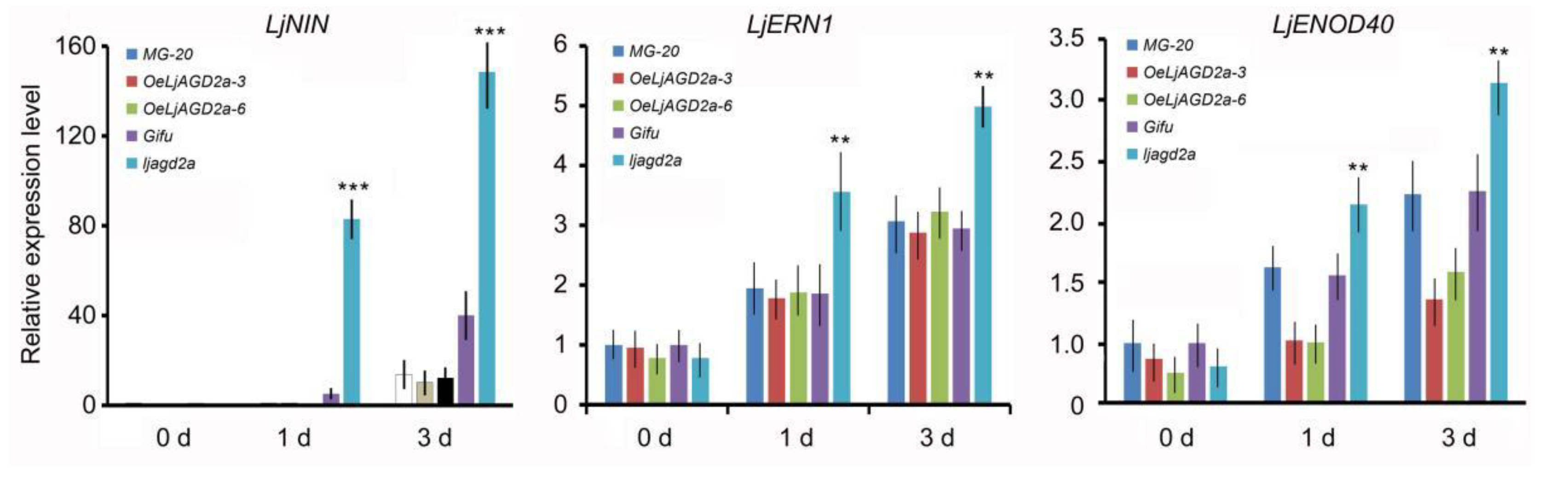
2.5. Knockout Mutation of LjAGD2a Increased Infection Thread and Nodule Numbers in L. japonicus
3. Discussion
3.1. LjAGD2s Function as LL-DAP-Ats
3.2. LjAGD2s May Have Functional Redundancy in Regulation of Plant Development
3.3. AGD2a Affects Disease Resistance and Nodulation via the Lys Biosynthesis Pathway
4. Materials and Methods
4.1. Plant Growth Conditions and Treatments
4.2. Plasmid Constructs and Plant Transformation
4.3. Functional Complementation of E. coli Dap Mutants and Enzyme Assays
4.4. Phylogenetic Tree Construction
4.5. RNA Isolation and Expression Analysis
4.6. Subcellular Localization of LjAGD2s
4.7. Histochemical Staining and Microscopic Analysis
4.8. Observation of Pollen Morphology and Germination
4.9. Cultivation of Bacteria and Plant Inoculation
4.10. Measurement of Salicylic Acid and Free Amino Acid Contents
4.11. Statistical Significance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartmann, M.; Zeier, J. L-lysine metabolism to N-hydroxypipecolic acid: An integral immune-activating pathway in plants. Plant J. 2018, 96, 5–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruda, P.; Kemper, E.L.; Papes, F.; Leite, A. Regulation of lysine catabolism in higher plants. Trends Plant Sci. 2000, 5, 324–330. [Google Scholar] [CrossRef]
- Bunsupa, S.; Yamazaki, M.; Saito, K. Quinolizidine alkaloid biosynthesis: Recent advances and future prospects. Front. Plant Sci. 2012, 3, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arruda, P.; Barreto, P. Lysine catabolism through the saccharopine pathway: Enzymes and intermediates involved in plant responses to abiotic and biotic stress. Front. Plant Sci. 2020, 11, 587. [Google Scholar] [CrossRef]
- Hartmann, M.; Zeier, T.; Bernsdorff, F.; Reichel-Deland, V.; Kim, D.; Hohmann, M.; Scholten, N.; Schuck, S.; Bräutigam, A.; Hölze, T.; et al. Flavin monooxygenase-generated N-hydroxypipecolic acid is a critical element of plant systemic immunity. Cell 2018, 173, 456.e16–469.e16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.C.; Holmes, E.C.; Rajniak, J.; Kim, J.G.; Tang, S.; Fischer, C.R.; Mudgett, M.B.; Sattely, E.S. N-hydroxy-pipecolic acid is a mobile metabolite that induces systemic disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4920–E4929. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Song, Z.Q.; Liu, T.; Guo, L.L.; Li, X.F. De novo assembly and comparative transcriptome analysis provide insight into lysine biosynthesis in Toona sinensis Roem. Int. J. Genom. 2016, 2016, 6735209. [Google Scholar] [CrossRef] [Green Version]
- Hudson, A.O.; Singh, B.K.; Leustek, T.; Gilvarg, C. An LL-diaminopimelate aminotransferase defines a novel variant of the lysine biosynthesis pathway in plants. Plant Physiol. 2006, 140, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Ding, P.; Rekhter, D.; Ding, Y.; Feussner, K.; Busta, L.; Haroth, S.; Xu, S.; Li, X.; Jetter, R.; Feussner, I.; et al. Characterization of a pipecolic acid biosynthesis pathway required for systemic acquired resistance. Plant Cell 2016, 28, 2603–2615. [Google Scholar] [CrossRef] [Green Version]
- Rate, D.N.; Greenberg, J.T. The Arabidopsis aberrant growth and death2 mutant shows resistance to Pseudomonas syringae and reveals a role for NPR1 in suppressing hypersensitive cell death. Plant J. 2001, 27, 203–211. [Google Scholar] [CrossRef]
- Hudson, A.O.; Bless, C.; Macedo, P.; Chatterjee, S.P.; Singh, B.K.; Gilvarg, C.; Leustek, T. Biosynthesis of lysine in plants: Evidence for a variant of the known bacterial pathways. BBA 2005, 1721, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, J.H.F.; Kirma, M.; Barros, J.A.S.; Quinhones, C.G.S.; Pereira-Lima, I.A.; Obata, T.; Nunes-Nesi, A.; Galili, G.; Fernie, A.R.; Avin-Wittenberg, T.; et al. An L,L-diaminopimelate aminotransferase mutation leads to metabolic shifts and growth inhibition in Arabidopsis. J. Exp. Bot. 2018, 69, 5489–5506. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, X.; Tian, L.; Wu, P.; Li, M.; Jiang, H.; Chen, Y.; Wu, G. Knockdown of LjALD1, AGD2-like defense response protein 1, influences plant growth and nodulation in Lotus japonicus. J. Integr. Plant Biol. 2014, 56, 1034–1041. [Google Scholar] [CrossRef] [PubMed]
- Song, J.T.; Lu, H.; Greenberg, J.T. Divergent roles in Arabidopsis thaliana development and defense of two homologous genes, aberrant growth and death2 and AGD2-LIKE DEFENSE RESPONSE PROTEIN1, encoding novel aminotransferases. Plant Cell 2004, 16, 353–366. [Google Scholar] [CrossRef] [Green Version]
- Cecchini, N.M.; Jung, H.W.; Engle, N.L.; Tschaplinski, T.J.; Greenberg, J.T. ALD1 regulates basal immune components and early inducible defense responses in Arabidopsis. Mol. Plant Microbe Interact. 2015, 28, 455–466. [Google Scholar] [CrossRef] [Green Version]
- Bryan, J.K. 5—Advances in the biochemistry of amino acid biosynthesis. In Intermediary Nitrogen Metabolism; Miflin, B.J., Lea, P.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 16, pp. 161–195. [Google Scholar]
- Galili, G. New insights into the regulation and functional significance of lysine metabolism in plants. Annu. Rev. Plant Biol. 2002, 53, 27–43. [Google Scholar] [CrossRef]
- Yang, Q.; Zhao, D.; Liu, Q. Connections between amino acid metabolisms in plants: Lysine as an example. Front. Plant Sci. 2020, 11, 928. [Google Scholar] [CrossRef]
- Stacey, G.; McAlvin, C.B.; Kim, S.Y.; Olivares, J.; Soto, M.J. Effects of endogenous salicylic acid on nodulation in the model legumes Lotus japonicus and Medicago truncatula. Plant Physiol. 2006, 141, 1473–1481. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.H.; Galili, G. Lysine metabolism is concurrently regulated by synthesis and catabolism in both reproductive and vegetative tissues. Plant Physiol. 2004, 135, 129–136. [Google Scholar] [CrossRef] [Green Version]
- Malolepszy, A.; Mun, T.; Sandal, N.; Gupta, V.; Dubin, M.; Urbanski, D.; Shah, N.; Bachmann, A.; Fukai, E.; Hirakawa, H.; et al. The LORE1 insertion mutant resource. Plant J. 2016, 88, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Mun, T.; Bachmann, A.; Gupta, V.; Stougaard, J.; Andersen, S.U. Lotus Base: An integrated information portal for the model legume Lotus japonicus. Sci. Rep. 2016, 6, 39447. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Fu, X.; Mei, X.; Zhou, Y.; Cheng, S.; Zeng, L.; Dong, F.; Yang, Z. Proteolysis of chloroplast proteins is responsible for accumulation of free amino acids in dark-treated tea (Camellia sinensis) leaves. J. Proteom. 2017, 157, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, L.; Liu, L.; Yin, Y.; Huang, M.; Chen, Y.; Xu, X.; Wu, P.; Li, M.; Wu, G.; Jiang, H.; et al. Heterogeneity in the expression and subcellular localization of POLYOL/MONOSACCHARIDE TRANSPORTER genes in Lotus japonicus. PLoS ONE 2017, 12, e0185269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Y.; Tian, L.; Li, X.; Huang, M.; Liu, L.; Wu, P.; Li, M.; Jiang, H.; Wu, G.; Chen, Y. The role of endogenous thiamine produced via THIC in root nodule symbiosis in Lotus japonicus. Plant Sci. 2019, 283, 311–320. [Google Scholar] [CrossRef]
- Qin, S.; Tang, Y.; Chen, Y.; Wu, P.; Li, M.; Wu, G.; Jiang, H. Overexpression of the starch phosphorylase-like gene (PHO3) in Lotus japonicus has a profound effect on the growth of plants and reduction of transitory starch accumulation. Front. Plant Sci. 2016, 7, 1315. [Google Scholar] [CrossRef]
- Tan, X.D.; Dai, X.Q.; Chen, T.; Wu, Y.S.; Yang, D.; Zheng, Y.X.; Chen, H.L.; Wan, X.R.; Yang, Y. Complete genome sequence analysis of Ralstonia solanacearum strain PeaFJ1 provides insights into its strong virulence in peanut plants. Front Microbiol. 2022, 13, 830900. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, M.; Yuan, M.; Sun, C.; Li, M.; Wu, P.; Jiang, H.; Wu, G.; Chen, Y. Roles of AGD2a in Plant Development and Microbial Interactions of Lotus japonicus. Int. J. Mol. Sci. 2022, 23, 6863. https://doi.org/10.3390/ijms23126863
Huang M, Yuan M, Sun C, Li M, Wu P, Jiang H, Wu G, Chen Y. Roles of AGD2a in Plant Development and Microbial Interactions of Lotus japonicus. International Journal of Molecular Sciences. 2022; 23(12):6863. https://doi.org/10.3390/ijms23126863
Chicago/Turabian StyleHuang, Mingchao, Mengru Yuan, Chunyu Sun, Meiru Li, Pingzhi Wu, Huawu Jiang, Guojiang Wu, and Yaping Chen. 2022. "Roles of AGD2a in Plant Development and Microbial Interactions of Lotus japonicus" International Journal of Molecular Sciences 23, no. 12: 6863. https://doi.org/10.3390/ijms23126863