
Developing Anti-Babesia bovis Blood Stage Vaccines: A New Perspective Regarding Synthetic Vaccines

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
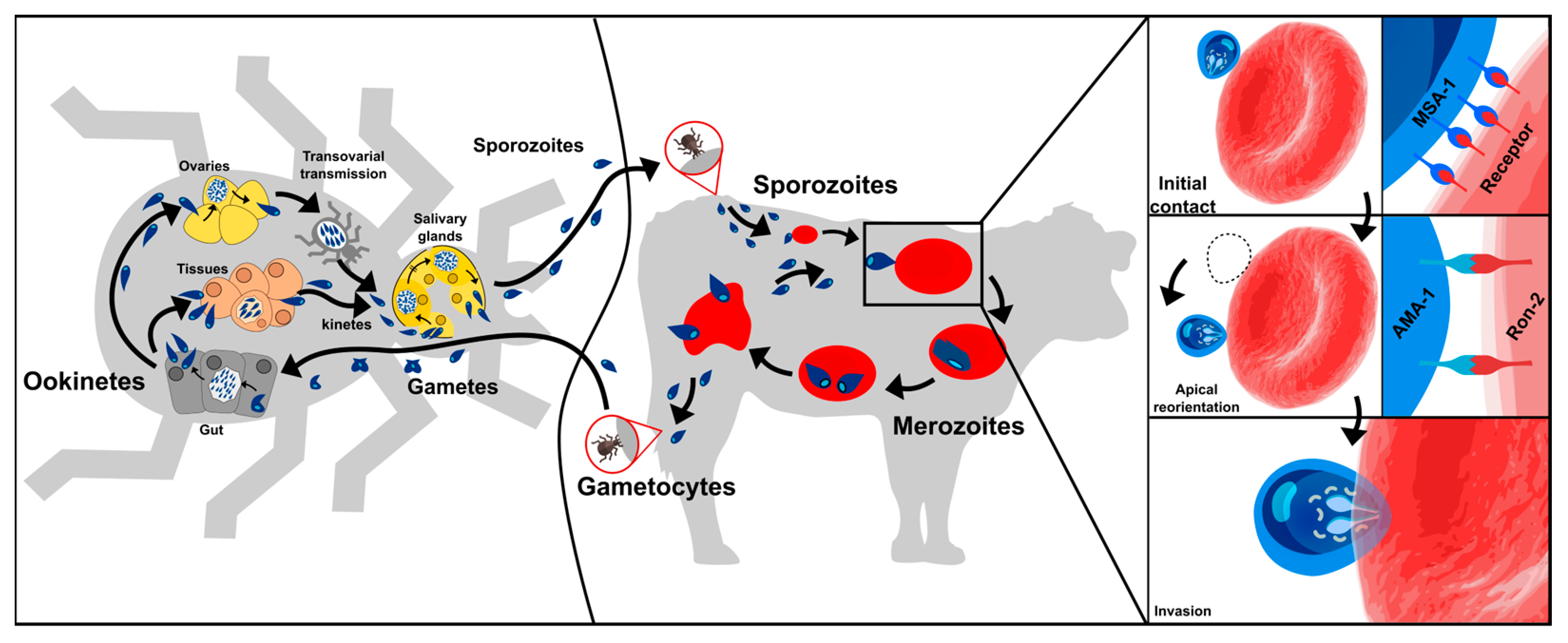
:1. Introduction
2. Anti-Babesia Vaccines
2.1. Live Attenuated Parasite Vaccines
2.2. Protein-Based Vaccines
2.3. Recombinant Viral Vector Vaccines
2.4. Antigenic Peptide-Based Vaccines
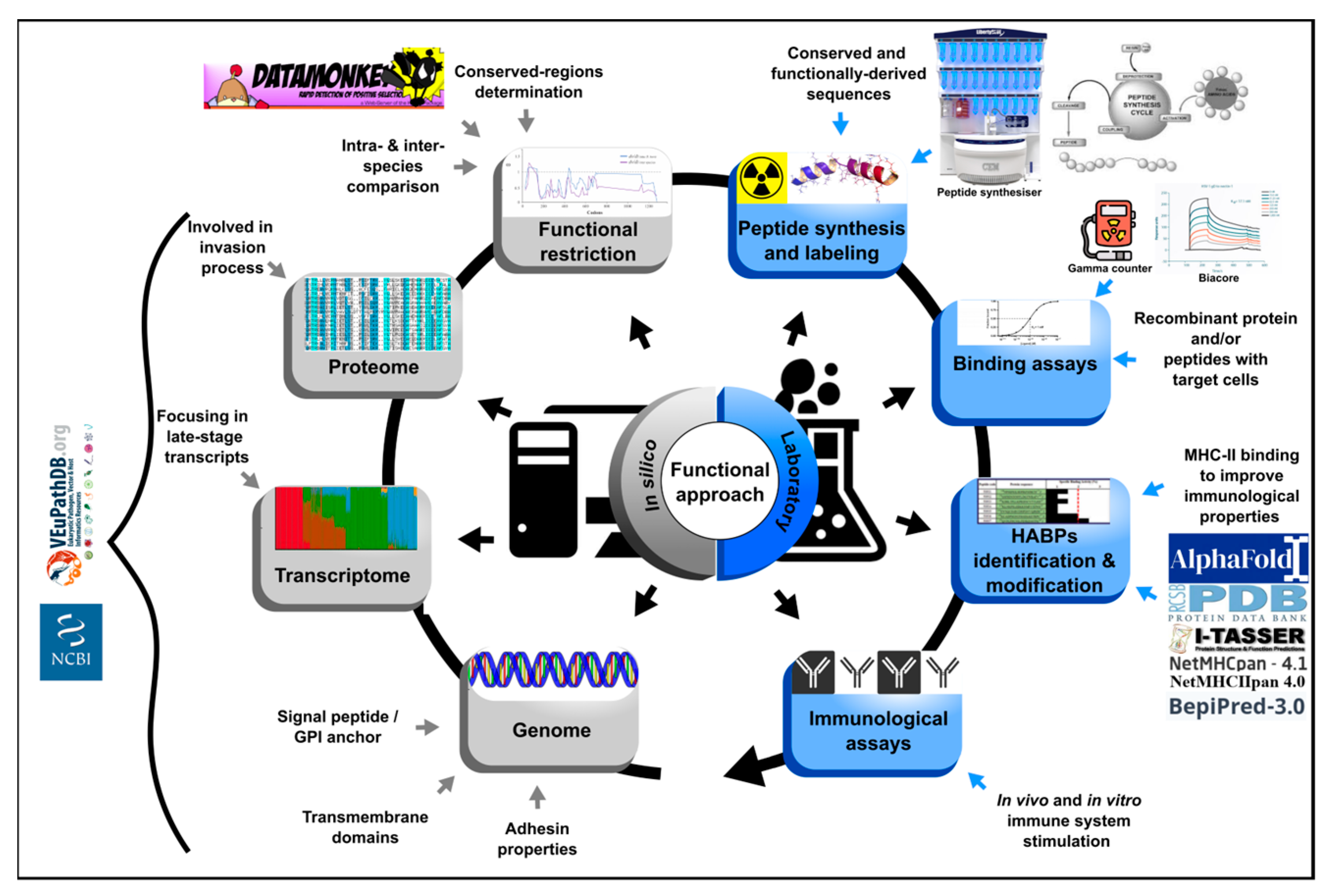
3. Functional Approach
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arisue, N.; Hashimoto, T. Phylogeny and evolution of apicoplasts and apicomplexan parasites. Parasitol. Int. 2015, 64, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Sibley, L.D. The moving junction, a key portal to host cell invasion by apicomplexan parasites. Curr. Opin. Microbiol. 2012, 15, 449–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargieri, D.; Lagal, V.; Andenmatten, N.; Tardieux, I.; Meissner, M.; Menard, R. Host cell invasion by apicomplexan parasites: The junction conundrum. PLOS Pathog. 2014, 10, e1004273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, C.A.; Rodriguez, M.; Cursino-Santos, J.R. Babesia and red cell invasion. Curr. Opin. Hematol. 2012, 19, 170–175. [Google Scholar] [CrossRef]
- Marques, R.; Kruger, R.F.; Peterson, A.T.; de Melo, L.F.; Vicenzi, N.; Jimenez-Garcia, D. Climate change implications for the distribution of the babesiosis and anaplasmosis tick vector, Rhipicephalus (Boophilus) microplus. Vet. Res. 2020, 51, 81. [Google Scholar] [CrossRef]
- Nyangiwe, N.; Horak, I.G.; Van der Mescht, L.; Matthee, S. Range expansion of the economically important Asiatic blue tick, Rhipicephalus microplus, in South Africa. J. South Afr. Vet. Assoc. 2017, 88, e1–e7. [Google Scholar] [CrossRef] [Green Version]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, A.; Moreau, E.; Bonnet, S.; Plantard, O.; Malandrin, L. Babesia and its hosts: Adaptation to long-lasting interactions as a way to achieve efficient transmission. Vet. Res. 2009, 40, 37. [Google Scholar] [CrossRef] [Green Version]
- Rathinasamy, V.; Poole, W.A.; Bastos, R.G.; Suarez, C.E.; Cooke, B.M. Babesiosis Vaccines: Lessons Learned, Challenges Ahead, and Future Glimpses. Trends Parasitol. 2019, 35, 622–635. [Google Scholar] [CrossRef]
- Jalovecka, M.; Sojka, D.; Ascencio, M.; Schnittger, L. Babesia Life Cycle—When Phylogeny Meets Biology. Trends Parasitol. 2019, 35, 356–368. [Google Scholar] [CrossRef]
- Jalovecka, M.; Bonsergent, C.; Hajdusek, O.; Kopacek, P.; Malandrin, L. Stimulation and quantification of Babesia divergens gametocytogenesis. Parasites Vectors 2016, 9, 439. [Google Scholar] [CrossRef] [PubMed]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosqueda, J.; Olvera-Ramirez, A.; Aguilar-Tipacamu, G.; Canto, G.J. Current advances in detection and treatment of babesiosis. Curr. Med. Chem. 2012, 19, 1504–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howell, J.M.; Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Knowles, D.P. Transovarial transmission efficiency of Babesia bovis tick stages acquired by Rhipicephalus (Boophilus) microplus during acute infection. J. Clin. Microbiol. 2007, 45, 426–431. [Google Scholar] [CrossRef] [Green Version]
- Suarez, C.E.; Noh, S. Emerging perspectives in the research of bovine babesiosis and anaplasmosis. Vet. Parasitol. 2011, 180, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Kivaria, F.M. Estimated direct economic costs associated with tick-borne diseases on cattle in Tanzania. Trop. Anim. Health Prod. 2006, 38, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.S.; Estrada-Pena, A.; Zintl, A. Vectors of Babesiosis. Annu. Rev. Entomol. 2019, 64, 149–165. [Google Scholar] [CrossRef]
- Vial, H.J.; Gorenflot, A. Chemotherapy against babesiosis. Vet. Parasitol. 2006, 138, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Nari, A. Strategies for the control of one-host ticks and relationship with tick-borne diseases in South America. Vet. Parasitol. 1995, 57, 153–165. [Google Scholar] [CrossRef]
- Andreotti, R.; Guerrero, F.D.; Soares, M.A.; Barros, J.C.; Miller, R.J.; Leon, A.P. Acaricide resistance of Rhipicephalus (Boophilus) microplus in State of Mato Grosso do Sul, Brazil. Rev. Bras. Parasitol. Vet. 2011, 20, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Higa Lde, O.; Garcia, M.V.; Barros, J.C.; Koller, W.W.; Andreotti, R. Evaluation of Rhipicephalus (Boophilus) microplus (Acari: Ixodidae) resistance to different acaricide formulations using samples from Brazilian properties. Rev. Bras. Parasitol. Vet. 2016, 25, 163–171. [Google Scholar] [CrossRef]
- Patarroyo, M.E.; Bermudez, A.; Patarroyo, M.A. Structural and immunological principles leading to chemically synthesized, multiantigenic, multistage, minimal subunit-based vaccine development. Chem. Rev. 2011, 111, 3459–3507. [Google Scholar] [CrossRef] [PubMed]
- Florin-Christensen, M.; Suarez, C.E.; Rodriguez, A.E.; Flores, D.A.; Schnittger, L. Vaccines against bovine babesiosis: Where we are now and possible roads ahead. Parasitology 2014, 141, 1563–1592. [Google Scholar] [CrossRef]
- Moreno, J.M. Evaluación Clínica e Inmunológica de una Vacuna Anabasan Limor Contra Hemoparásitos Bovinos; Universidad de La Salle: Bogotá, Colombia, 1998. [Google Scholar]
- Vacuna Contra la Anaplasmosis y la Babesiosis Bovina. Available online: https://www.argentina.gob.ar/inta/tecnologias/vacuna-contra-la-anaplasmosis-y-la-babesiosis-bovina-0 (accessed on 24 November 2022).
- Shkap, V.; de Vos, A.J.; Zweygarth, E.; Jongejan, F. Attenuated vaccines for tropical theileriosis, babesiosis and heartwater: The continuing necessity. Trends Parasitol. 2007, 23, 420–426. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Rojas, C.; Figueroa, J.V. An Overview of Current Knowledge on in vitro Babesia Cultivation for Production of Live Attenuated Vaccines for Bovine Babesiosis in Mexico. Front. Vet. Sci. 2020, 7, 364. [Google Scholar] [CrossRef]
- Connoway, J.W.; Francines, M. Texas fever: Experiments made by the Texas Experiment Station in immunizing Northern breeding cattle against Texas fever. In Station; Texas Agricultural Experiment Station; J.J. Pastoriza Printing & Litho. Co.: Houston, TX, USA, 1902; pp. 56–106. [Google Scholar]
- Callow, L.L.; Mellors, L.T.; McGregor, W. Reduction in virulence of Babesia bovis due to rapid passage in splenectomized cattle. Int. J. Parasitol. 1979, 9, 333–338. [Google Scholar] [CrossRef]
- Callow, L.A.; Dalgliesh, R. The development of effective, safe vaccination against babesiosis and anaplasmosis in Australia. In Proceedings of the 56th Annual Conference of the Australian Veterinary Association, Townsville, Australia, 14–18 May 1979. [Google Scholar]
- Bock, R.; Jackson, L.; de Vos, A.; Jorgensen, W. Babesiosis of cattle. Parasitology 2004, 129 (Suppl. S1), S247–S269. [Google Scholar] [CrossRef]
- Yunker, C.E.; Kuttler, K.L.; Johnson, L.W. Attenuation of Babesia bovis by in vitro cultivation. Vet. Parasitol. 1987, 24, 7–13. [Google Scholar] [CrossRef]
- Alarcón, G.; Millán, J.; Martínez, J.; Aragón, J.; Vega, C. Capacidad inmunoprotectora de una clona irradiada de Babesia bovis derivada de cultivo in vitro. Tec. Pecu. Mex. 1996, 34, 127. [Google Scholar]
- Mangold, A.J.; Vanzini, V.R.; Echaide, I.E.; de Echaide, S.T.; Volpogni, M.M.; Guglielmone, A.A. Viability after thawing and dilution of simultaneously cryopreserved vaccinal Babesia bovis and Babesia bigemina strains cultured in vitro. Vet. Parasitol. 1996, 61, 345–348. [Google Scholar] [CrossRef]
- Alarcón, J.; Alvaréz, J.; Ramírez, E.; Aragón, J.; Gualito, J. Protection against bovine babesiosis with a mixed vaccine of Babesia bovis and Babesia bigemina derived from in vitro culture under field confrontation. Immun. Dis. Free. Area 2003, 34, 11. [Google Scholar]
- Rojas, C.; Figueroa, J.V.; Alvarado, A.; Mejia, P.; Mosqueda, J.J.; Falcon, A.; Vega, C.A.; Alvarez, A. Bovine babesiosis live vaccine production: Use of gamma irradiation on the substrate. Ann. N. Y. Acad. Sci. 2006, 1081, 405–416. [Google Scholar] [CrossRef]
- de Waal, D.T.; Combrink, M.P. Live vaccines against bovine babesiosis. Vet. Parasitol. 2006, 138, 88–96. [Google Scholar] [CrossRef]
- Fish, L.; Leibovich, B.; Krigel, Y.; McElwain, T.; Shkap, V. Vaccination of cattle against B. bovis infection with live attenuated parasites and non-viable immunogens. Vaccine 2008, 26 (Suppl. S6), G29–G33. [Google Scholar] [CrossRef]
- Brown, W.C.; Palmer, G.H. Designing blood-stage vaccines against Babesia bovis and B. bigemina. Parasitol. Today 1999, 15, 275–281. [Google Scholar] [CrossRef]
- Brown, W.C.; Norimine, J.; Goff, W.L.; Suarez, C.E.; McElwain, T.F. Prospects for recombinant vaccines against Babesia bovis and related parasites. Parasite Immunol. 2006, 28, 315–327. [Google Scholar] [CrossRef]
- Nascimento, I.P.; Leite, L.C. Recombinant vaccines and the development of new vaccine strategies. Braz. J. Med. Biol. Res. 2012, 45, 1102–1111. [Google Scholar] [CrossRef] [Green Version]
- Wright, I.G.; Casu, R.; Commins, M.A.; Dalrymple, B.P.; Gale, K.R.; Goodger, B.V.; Riddles, P.W.; Waltisbuhl, D.J.; Abetz, I.; Berrie, D.A.; et al. The development of a recombinant Babesia vaccine. Vet. Parasitol. 1992, 44, 3–13. [Google Scholar] [CrossRef]
- Hope, M.; Riding, G.; Menzies, M.; Colditz, I.; Reverter, A.; Willadsen, P. Potential for recombinant Babesia bovis antigens to protect against a highly virulent isolate. Parasite Immunol. 2005, 27, 439–445. [Google Scholar] [CrossRef]
- Brown, W.C.; McElwain, T.F.; Ruef, B.J.; Suarez, C.E.; Shkap, V.; Chitko-McKown, C.G.; Tuo, W.; Rice-Ficht, A.C.; Palmer, G.H. Babesia bovis rhoptry-associated protein 1 is immunodominant for T helper cells of immune cattle and contains T-cell epitopes conserved among geographically distant B. bovis strains. Infect. Immun. 1996, 64, 3341–3350. [Google Scholar] [CrossRef] [Green Version]
- Norimine, J.; Suarez, C.E.; McElwain, T.F.; Florin-Christensen, M.; Brown, W.C. Immunodominant epitopes in Babesia bovis rhoptry-associated protein 1 that elicit memory CD4(+)-T-lymphocyte responses in B. bovis-immune individuals are located in the amino-terminal domain. Infect. Immun. 2002, 70, 2039–2048. [Google Scholar] [CrossRef] [Green Version]
- Brown, W.C.; McElwain, T.F.; Palmer, G.H.; Chantler, S.E.; Estes, D.M. Bovine CD4(+) T-lymphocyte clones specific for rhoptry-associated protein 1 of Babesia bigemina stimulate enhanced immunoglobulin G1 (IgG1) and IgG2 synthesis. Infect. Immun. 1999, 67, 155–164. [Google Scholar] [CrossRef] [Green Version]
- Norimine, J.; Mosqueda, J.; Suarez, C.; Palmer, G.H.; McElwain, T.F.; Mbassa, G.; Brown, W.C. Stimulation of T-helper cell gamma interferon and immunoglobulin G responses specific for Babesia bovis rhoptry-associated protein 1 (RAP-1) or a RAP-1 protein lacking the carboxy-terminal repeat region is insufficient to provide protective immunity against virulent B. bovis challenge. Infect. Immun. 2003, 71, 5021–5032. [Google Scholar]
- Hines, S.A.; Palmer, G.H.; Jasmer, D.P.; McGuire, T.C.; McElwain, T.F. Neutralization-sensitive merozoite surface antigens of Babesia bovis encoded by members of a polymorphic gene family. Mol. Biochem. Parasitol. 1992, 55, 85–94. [Google Scholar] [CrossRef]
- Hines, S.A.; Palmer, G.H.; Jasmer, D.P.; Goff, W.L.; McElwain, T.F. Immunization of cattle with recombinant Babesia bovis merozoite surface antigen-1. Infect. Immun. 1995, 63, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Saul, A. Kinetic constraints on the development of a malaria vaccine. Parasite Immunol. 1987, 9, 1–9. [Google Scholar] [CrossRef]
- Terkawi, M.A.; Ratthanophart, J.; Salama, A.; AbouLaila, M.; Asada, M.; Ueno, A.; Alhasan, H.; Guswanto, A.; Masatani, T.; Yokoyama, N.; et al. Molecular characterization of a new Babesia bovis thrombospondin-related anonymous protein (BbTRAP2). PLoS ONE 2013, 8, e83305. [Google Scholar] [CrossRef] [Green Version]
- Rittipornlertrak, A.; Nambooppha, B.; Muenthaisong, A.; Apinda, N.; Koonyosying, P.; Srisawat, W.; Chomjit, P.; Sangkakam, K.; Punyapornwithaya, V.; Tiwananthagorn, S.; et al. Immunization of Cattle With Recombinant Structural Ectodomains I and II of Babesia bovis Apical Membrane Antigen 1 [BbAMA-1(I/II)] Induces Strong Th1 Immune Response. Front. Vet. Sci. 2022, 9, 917389. [Google Scholar] [CrossRef]
- Antonio Alvarez, J.; Lopez, U.; Rojas, C.; Borgonio, V.M.; Sanchez, V.; Castaneda, R.; Vargas, P.; Figueroa, J.V. Immunization of Bos taurus steers with Babesia bovis recombinant antigens MSA-1, MSA-2c and 12D3. Transbound. Emerg. Dis. 2010, 57, 87–90. [Google Scholar] [CrossRef]
- Brown, W.C.; McGuire, T.C.; Zhu, D.; Lewin, H.A.; Sosnow, J.; Palmer, G.H. Highly conserved regions of the immunodominant major surface protein 2 of the genogroup II ehrlichial pathogen Anaplasma marginale are rich in naturally derived CD4+ T lymphocyte epitopes that elicit strong recall responses. J. Immunol. 2001, 166, 1114–1124. [Google Scholar] [CrossRef] [Green Version]
- Brown, W.C.; Norimine, J.; Knowles, D.P.; Goff, W.L. Immune control of Babesia bovis infection. Vet. Parasitol. 2006, 138, 75–87. [Google Scholar] [CrossRef]
- Torina, A.; Blanda, V.; Villari, S.; Piazza, A.; La Russa, F.; Grippi, F.; La Manna, M.P.; Di Liberto, D.; de la Fuente, J.; Sireci, G. Immune Response to Tick-Borne Hemoparasites: Host Adaptive Immune Response Mechanisms as Potential Targets for Therapies and Vaccines. Int. J. Mol. Sci. 2020, 21, 8813. [Google Scholar] [CrossRef]
- Clark, T.G.; Cassidy-Hanley, D. Recombinant subunit vaccines: Potentials and constraints. Dev. Biol. 2005, 121, 153–163. [Google Scholar]
- Jaramillo Ortiz, J.M.; Del Medico Zajac, M.P.; Zanetti, F.A.; Molinari, M.P.; Gravisaco, M.J.; Calamante, G.; Wilkowsky, S.E. Vaccine strategies against Babesia bovis based on prime-boost immunizations in mice with modified vaccinia Ankara vector and recombinant proteins. Vaccine 2014, 32, 4625–4632. [Google Scholar] [CrossRef]
- Gomez, C.E.; Najera, J.L.; Krupa, M.; Esteban, M. The poxvirus vectors MVA and NYVAC as gene delivery systems for vaccination against infectious diseases and cancer. Curr. Gene Ther. 2008, 8, 97–120. [Google Scholar] [CrossRef]
- Moss, B. Genetically engineered poxviruses for recombinant gene expression, vaccination, and safety. Proc. Natl. Acad. Sci. USA 1996, 93, 11341–11348. [Google Scholar] [CrossRef] [Green Version]
- Jaramillo Ortiz, J.M.; Molinari, M.P.; Gravisaco, M.J.; Paoletta, M.S.; Montenegro, V.N.; Wilkowsky, S.E. Evaluation of different heterologous prime-boost immunization strategies against Babesia bovis using viral vectored and protein-adjuvant vaccines based on a chimeric multi-antigen. Vaccine 2016, 34, 3913–3919. [Google Scholar] [CrossRef]
- Jaramillo Ortiz, J.M.; Paoletta, M.S.; Gravisaco, M.J.; Lopez Arias, L.S.; Montenegro, V.N.; de la Fourniere, S.A.M.; Valenzano, M.N.; Guillemi, E.C.; Valentini, B.; Echaide, I.; et al. Immunisation of cattle against Babesia bovis combining a multi-epitope modified vaccinia Ankara virus and a recombinant protein induce strong Th1 cell responses but fails to trigger neutralising antibodies required for protection. Ticks Tick Borne Dis. 2019, 10, 101270. [Google Scholar] [CrossRef]
- Francis, M.J. Recent Advances in Vaccine Technologies. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 231–241. [Google Scholar] [CrossRef]
- Hidalgo-Ruiz, M.; Mejia-Lopez, S.; Perez-Serrano, R.M.; Zaldivar-Lelo de Larrea, G.; Ganzinelli, S.; Florin-Christensen, M.; Suarez, C.E.; Hernandez-Ortiz, R.; Mercado-Uriostegui, M.A.; Rodriguez-Torres, A.; et al. Babesia bovis AMA-1, MSA-2c and RAP-1 contain conserved B and T-cell epitopes, which generate neutralizing antibodies and a long-lasting Th1 immune response in vaccinated cattle. Vaccine 2022, 40, 1108–1115. [Google Scholar] [CrossRef]
- Rittipornlertrak, A.; Nambooppha, B.; Muenthaisong, A.; Punyapornwithaya, V.; Tiwananthagorn, S.; Chung, Y.T.; Tuvshintulga, B.; Sivakumar, T.; Yokoyama, N.; Sthitmatee, N. Structural and immunological characterization of an epitope within the PAN motif of ectodomain I in Babesia bovis apical membrane antigen 1 for vaccine development. PeerJ 2021, 9, e11765. [Google Scholar] [CrossRef]
- Hidalgo-Ruiz, M.; Suarez, C.E.; Mercado-Uriostegui, M.A.; Hernandez-Ortiz, R.; Ramos, J.A.; Galindo-Velasco, E.; Leon-Avila, G.; Hernandez, J.M.; Mosqueda, J. Babesia bovis RON2 contains conserved B-cell epitopes that induce an invasion-blocking humoral immune response in immunized cattle. Parasit Vectors 2018, 11, 575. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [Green Version]
- Moyle, P.M.; Toth, I. Modern subunit vaccines: Development, components, and research opportunities. ChemMedChem 2013, 8, 360–376. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Dayananda, K.K.; Achur, R.N.; Gowda, D.C. Epidemiology, drug resistance, and pathophysiology of Plasmodium vivax malaria. J. Vector Borne Dis. 2018, 55, 1–8. [Google Scholar]
- Patarroyo, M.E.; Arevalo-Pinzon, G.; Reyes, C.; Moreno-Vranich, A.; Patarroyo, M.A. Malaria Parasite Survival Depends on Conserved Binding Peptides’ Critical Biological Functions. Curr. Issues Mol. Biol. 2016, 18, 57–78. [Google Scholar]
- Patarroyo, M.A.; Arevalo-Pinzon, G.; Moreno-Perez, D.A. From a basic to a functional approach for developing a blood stage vaccine against Plasmodium vivax. Expert Rev. Vaccines 2020, 19, 195–207. [Google Scholar] [CrossRef]
- Garzon-Ospina, D.; Forero-Rodriguez, J.; Patarroyo, M.A. Inferring natural selection signals in Plasmodium vivax-encoded proteins having a potential role in merozoite invasion. Infect. Genet. Evol. 2015, 33, 182–188. [Google Scholar] [CrossRef]
- Cuy-Chaparro, L.; Ricaurte-Contreras, L.A.; Bohorquez, M.D.; Arevalo-Pinzon, G.; Barreto-Santamaria, A.; Pabon, L.; Reyes, C.; Moreno-Perez, D.A.; Patarroyo, M.A. Identification of Babesia bovis MSA-1 functionally constraint regions capable of binding to bovine erythrocytes. Vet. Parasitol. 2022, 312, 109834. [Google Scholar] [CrossRef]
- Cuy-Chaparro, L.; Bohorquez, M.D.; Arevalo-Pinzon, G.; Castaneda-Ramirez, J.J.; Suarez, C.F.; Pabon, L.; Ordonez, D.; Gallego-Lopez, G.M.; Suarez, C.E.; Moreno-Perez, D.A.; et al. Babesia Bovis Ligand-Receptor Interaction: AMA-1 Contains Small Regions Governing Bovine Erythrocyte Binding. Int. J. Mol. Sci. 2021, 22, 714. [Google Scholar] [CrossRef]
- Jorge, S.; Dellagostin, O. The development of veterinary vaccines: A review of traditional methods and modern biotechnology approaches. Biotechnol. Res. Innov. 2017, 1, 6–13. [Google Scholar] [CrossRef]
- Rappuoli, R.; Covacci, A. Reverse vaccinology and genomics. Science 2003, 302, 602. [Google Scholar] [CrossRef] [PubMed]
- Rappuoli, R. Reverse vaccinology, a genome-based approach to vaccine development. Vaccine 2001, 19, 2688–2691. [Google Scholar] [CrossRef] [PubMed]
- Bambini, S.; Rappuoli, R. The use of genomics in microbial vaccine development. Drug Discov. Today 2009, 14, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Zauner, W.; Lingnau, K.; Mattner, F.; von Gabain, A.; Buschle, M. Defined synthetic vaccines. Biol Chem 2001, 382, 581–595. [Google Scholar] [CrossRef]
- De Brito, R.C.F.; Cardoso, J.M.O.; Reis, L.E.S.; Vieira, J.F.; Mathias, F.A.S.; Roatt, B.M.; Aguiar-Soares, R.; Ruiz, J.C.; Resende, D.M.; Reis, A.B. Peptide Vaccines for Leishmaniasis. Front. Immunol. 2018, 9, 1043. [Google Scholar] [CrossRef] [Green Version]
- Azmi, F.; Ahmad Fuaad, A.A.; Skwarczynski, M.; Toth, I. Recent progress in adjuvant discovery for peptide-based subunit vaccines. Hum. Vaccines Immunother. 2014, 10, 778–796. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Kim, D.S. Peptide Immunotherapy in Vaccine Development: From Epitope to Adjuvant. Adv. Protein Chem. Struct. Biol. 2015, 99, 1–14. [Google Scholar]
- Mosqueda, J.; McElwain, T.F.; Stiller, D.; Palmer, G.H. Babesia bovis merozoite surface antigen 1 and rhoptry-associated protein 1 are expressed in sporozoites, and specific antibodies inhibit sporozoite attachment to erythrocytes. Infect. Immun. 2002, 70, 1599–1603. [Google Scholar] [CrossRef] [Green Version]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuy-Chaparro, L.; Reyes, C.; Díaz-Guiot, E.V.; Moreno-Pérez, D.A.; Patarroyo, M.A. Developing Anti-Babesia bovis Blood Stage Vaccines: A New Perspective Regarding Synthetic Vaccines. Int. J. Mol. Sci. 2023, 24, 5219. https://doi.org/10.3390/ijms24065219
Cuy-Chaparro L, Reyes C, Díaz-Guiot EV, Moreno-Pérez DA, Patarroyo MA. Developing Anti-Babesia bovis Blood Stage Vaccines: A New Perspective Regarding Synthetic Vaccines. International Journal of Molecular Sciences. 2023; 24(6):5219. https://doi.org/10.3390/ijms24065219
Chicago/Turabian StyleCuy-Chaparro, Laura, César Reyes, Eliana Vanessa Díaz-Guiot, Darwin Andrés Moreno-Pérez, and Manuel Alfonso Patarroyo. 2023. "Developing Anti-Babesia bovis Blood Stage Vaccines: A New Perspective Regarding Synthetic Vaccines" International Journal of Molecular Sciences 24, no. 6: 5219. https://doi.org/10.3390/ijms24065219