
The Alpha-1 Subunit of the Na+/K+-ATPase (ATP1A1) Is a Host Factor Involved in the Attachment of Porcine Epidemic Diarrhea Virus
 ,
,
Abstract
:1. Introduction
2. Results
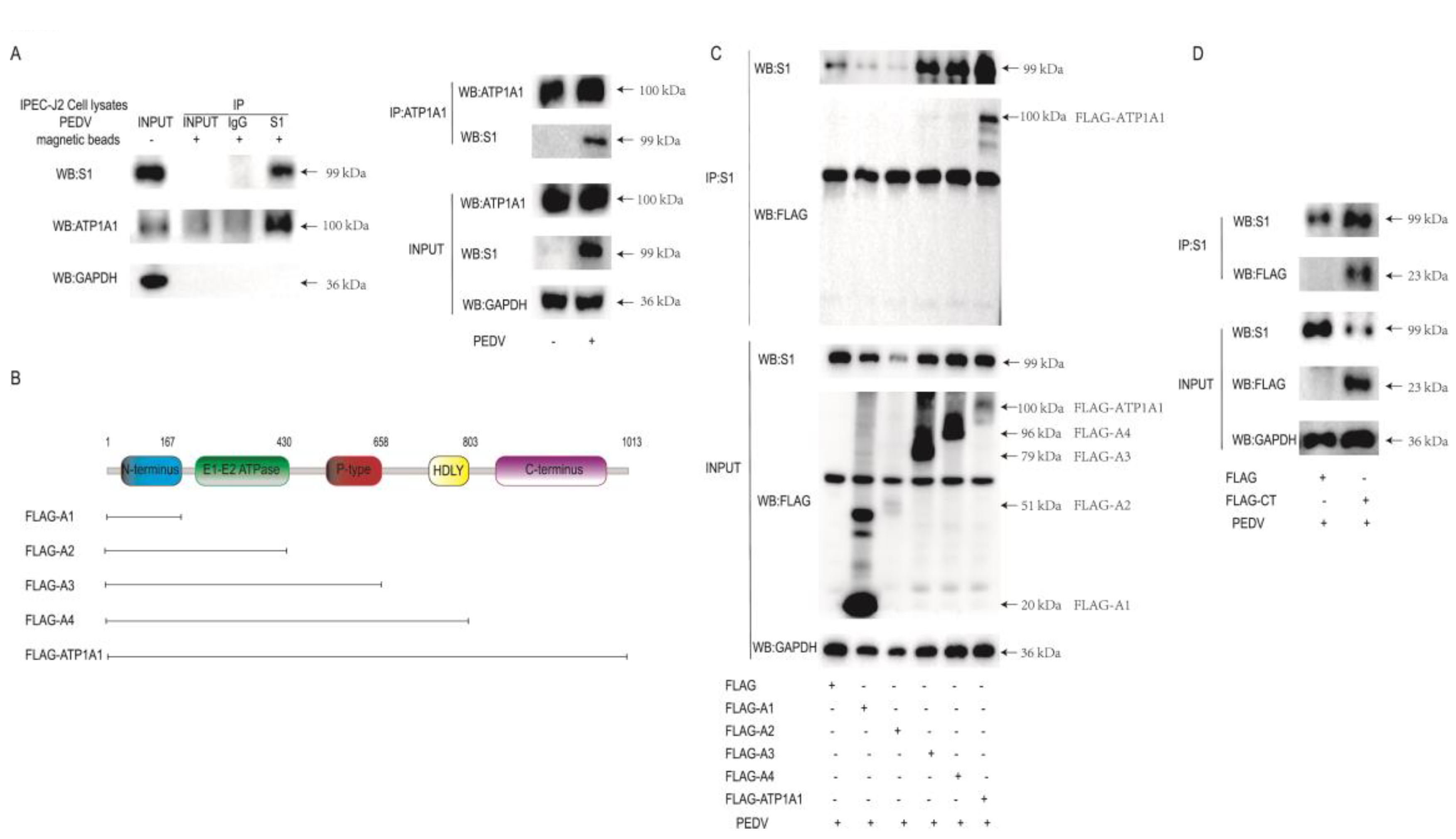
2.1. ATP1A1 CT Structural Domain Is Required for Interaction with PEDV S1
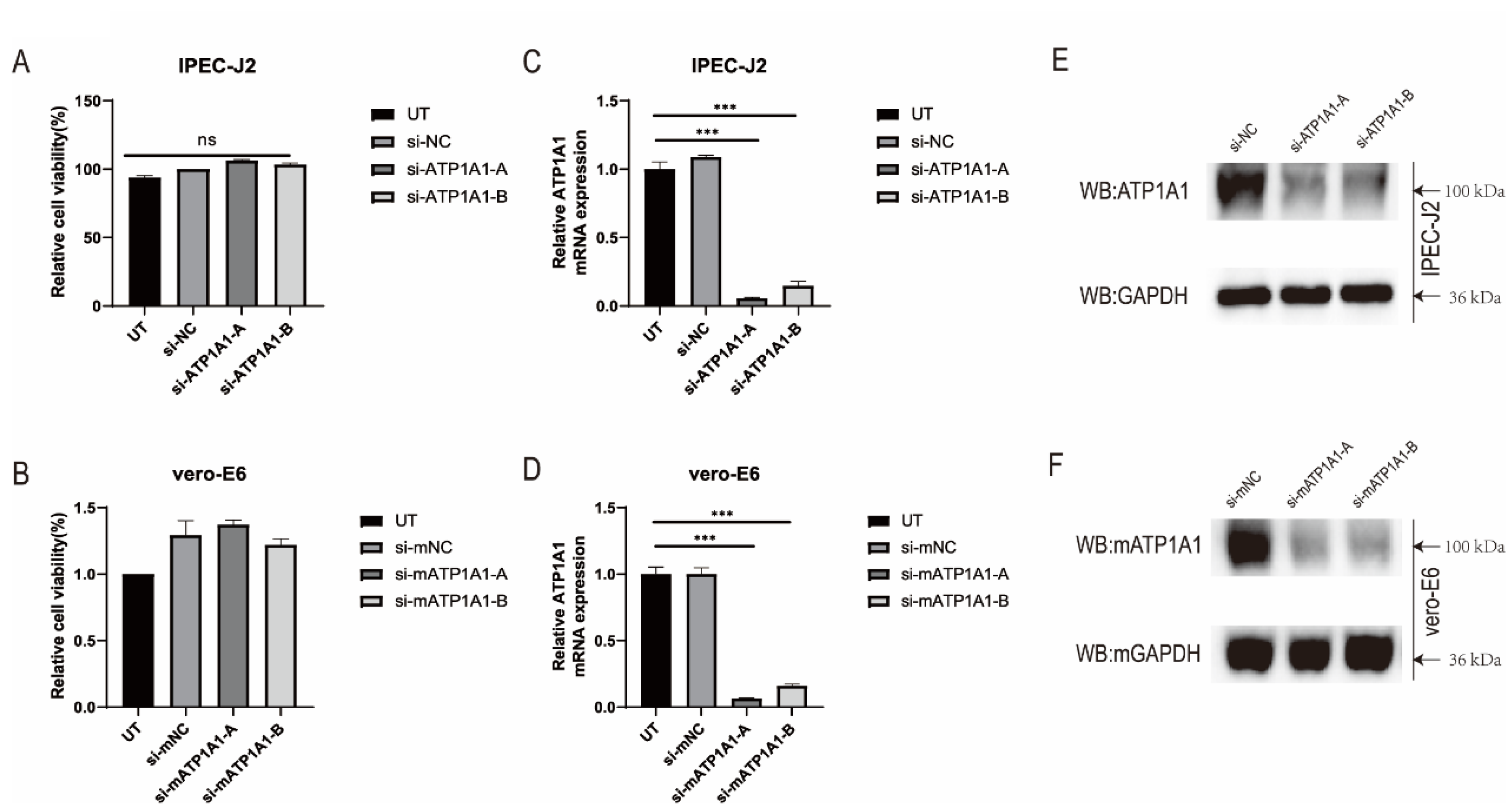
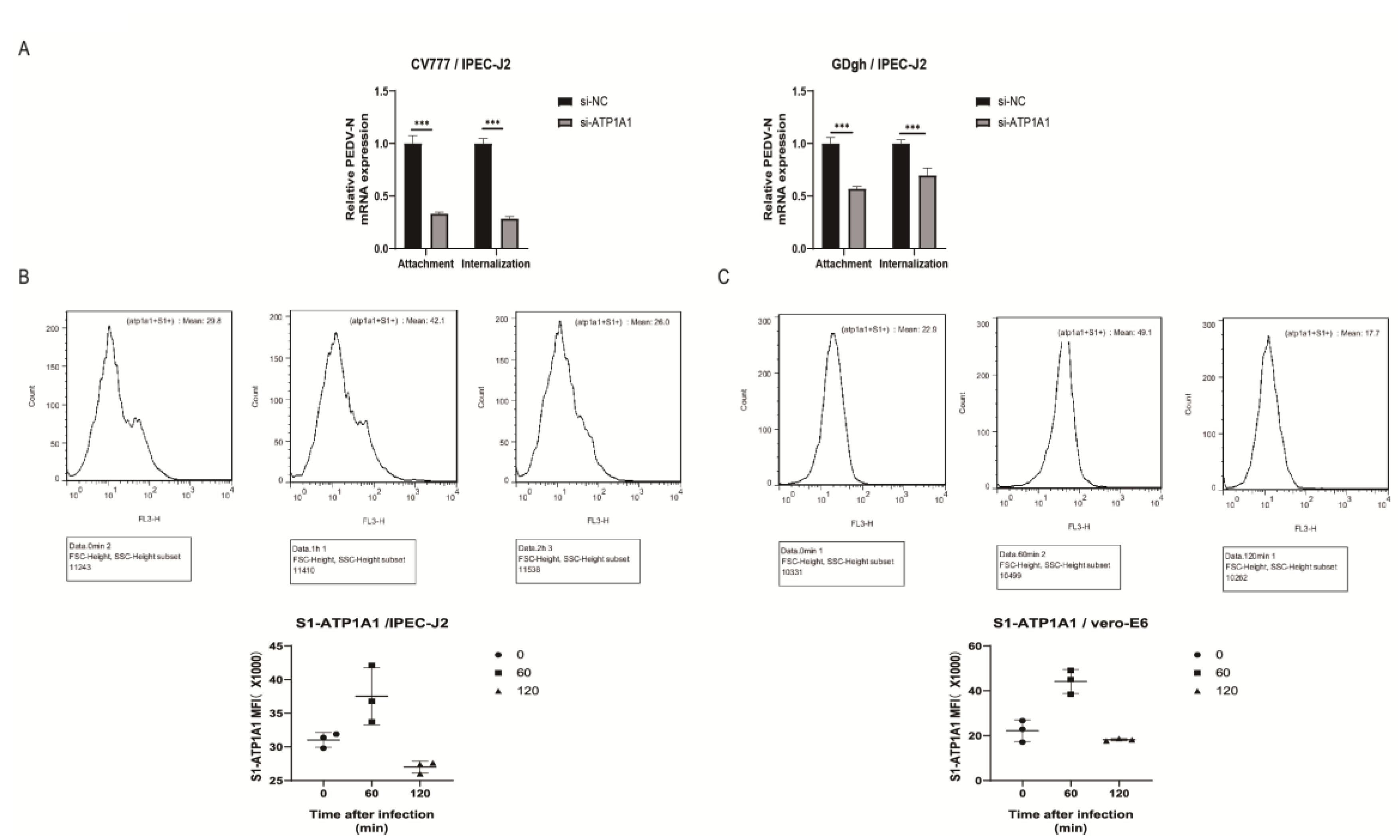
2.2. Knockdown of ATP1A1 Expression by siRNAs Transfection
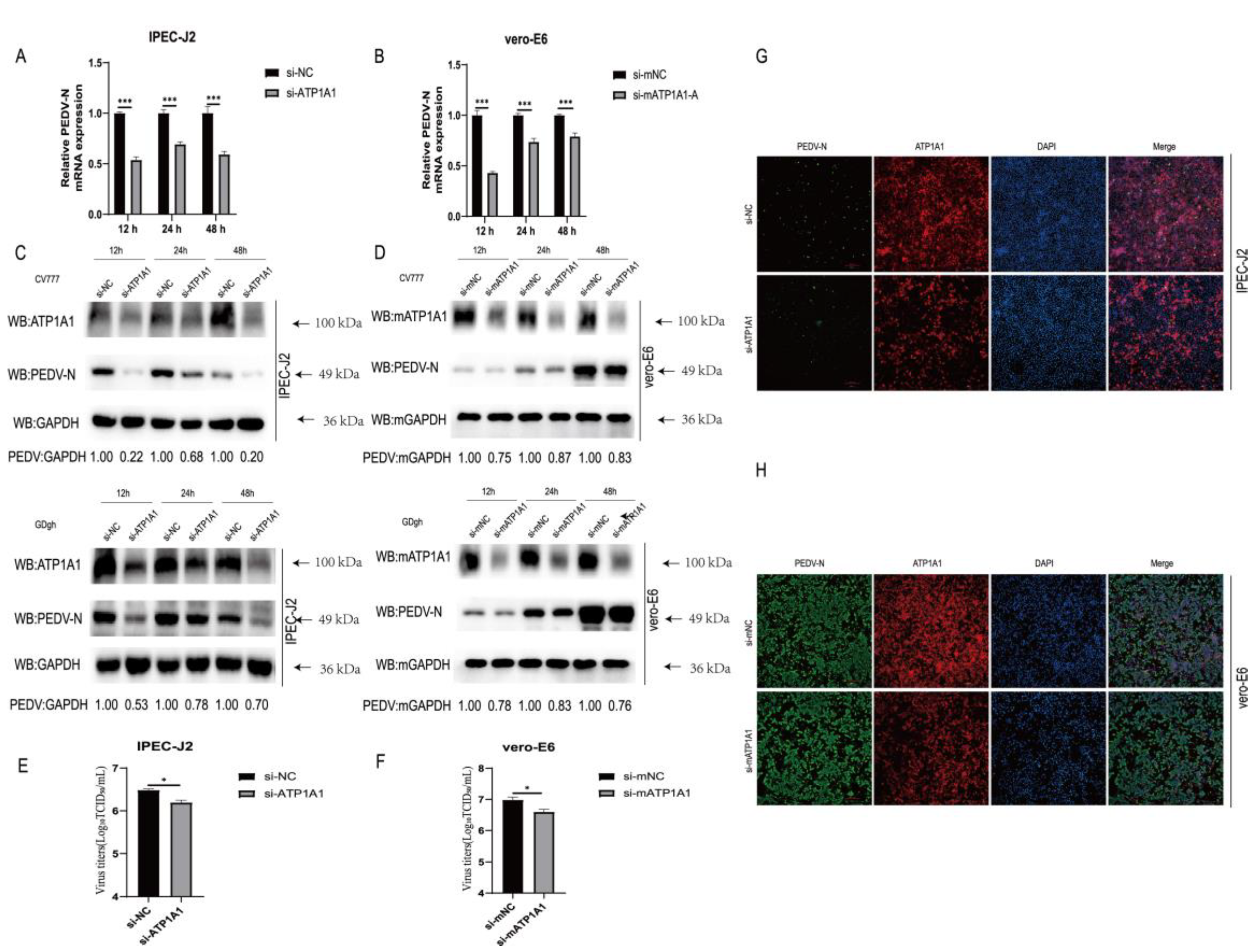
2.3. Knockdown of Endogenous ATP1A1 Expression Suppresses PEDV Infection
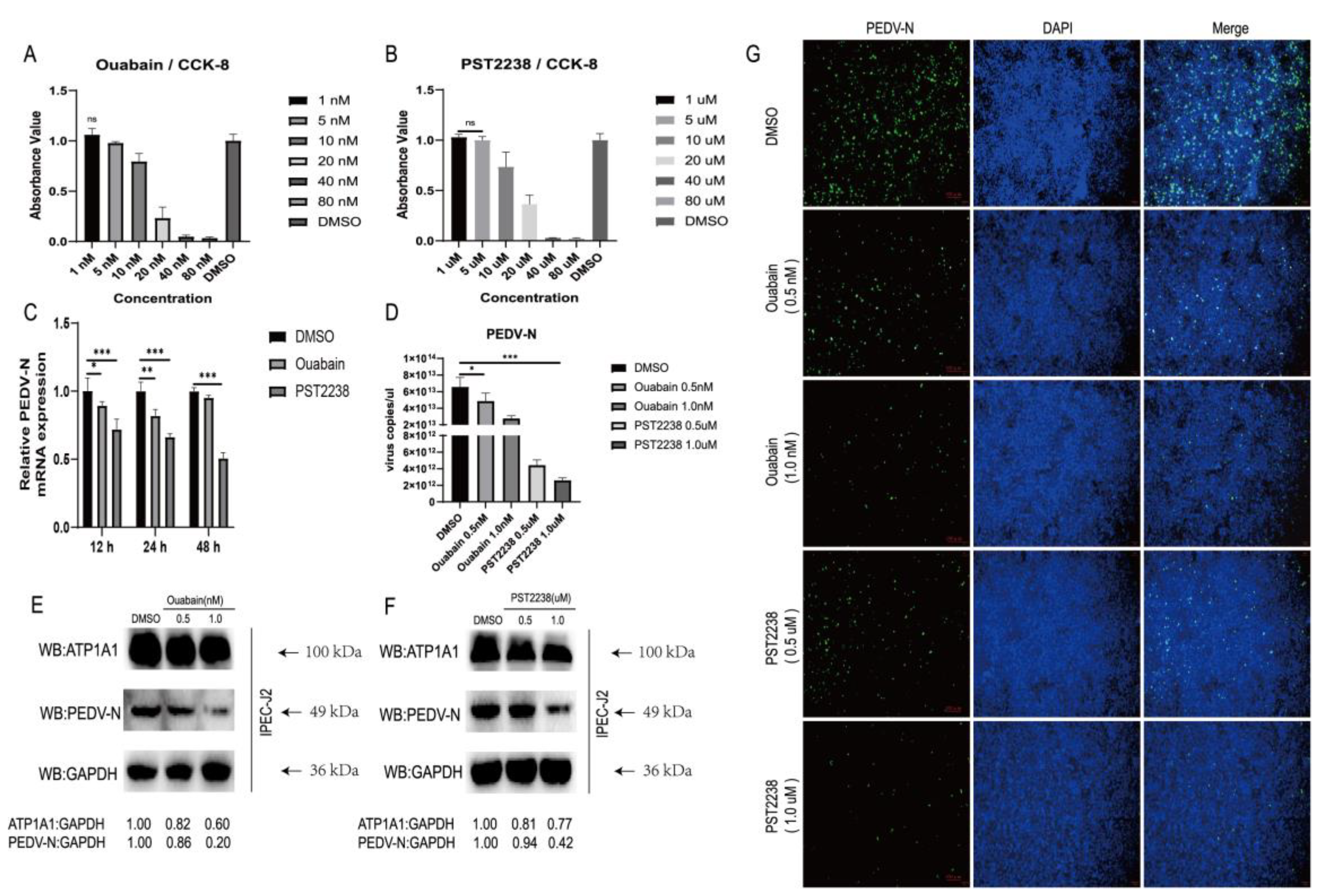
2.4. NKA Inhibitors Promote Degradation of ATP1A1 and Effectively Reduce PEDV Infection
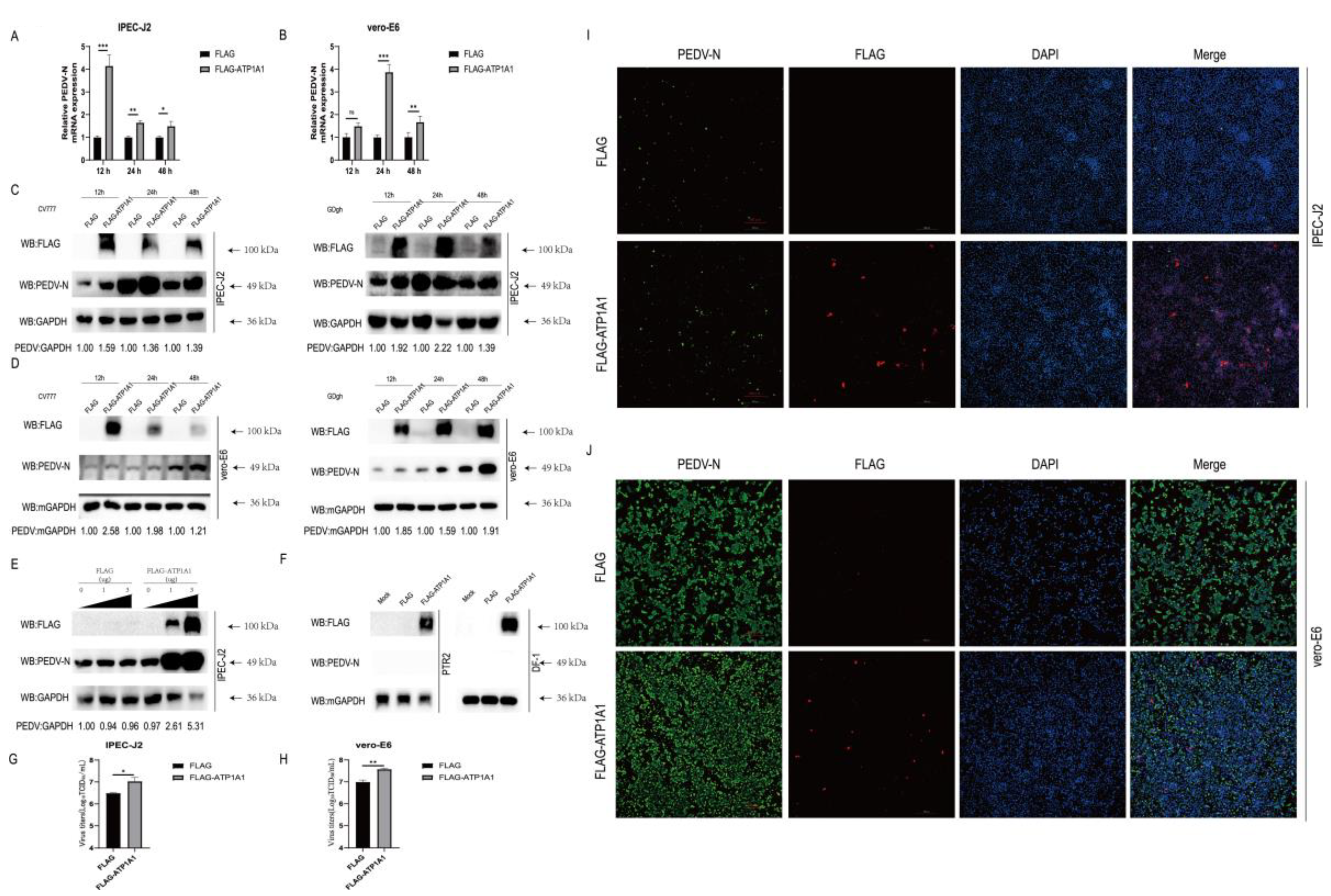
2.5. Overexpression of ATP1A1 Promotes PEDV Infection
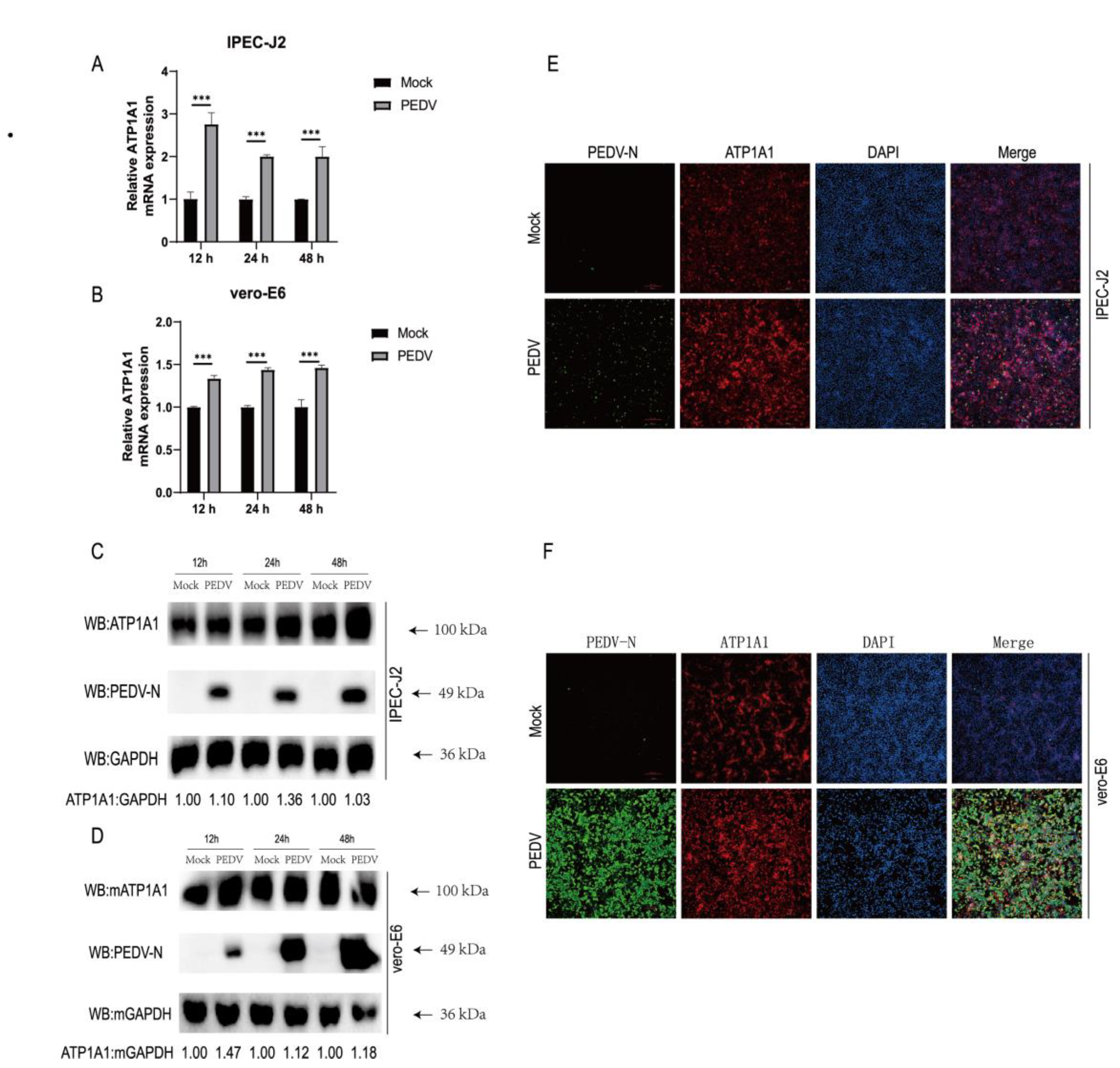
2.6. PEDV Infection Upregulates ATP1A1 Protein Expression in Target Cells
2.7. Knockdown of ATP1A1 Affects the Attachment of PEDV
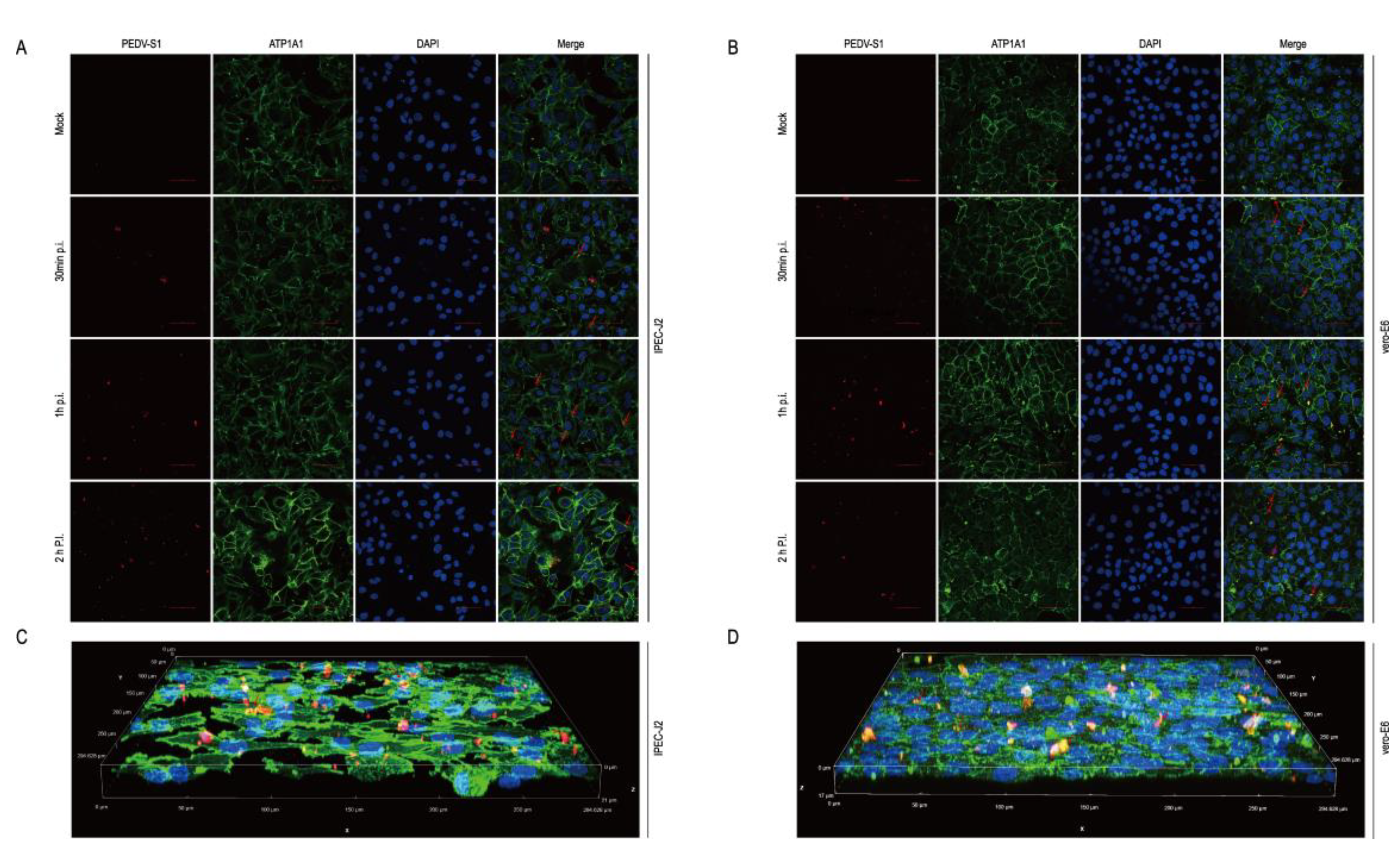
2.8. The Host Protein ATP1A1 Co-Localizes with the PEDV S1 Protein Early in PEDV Infection
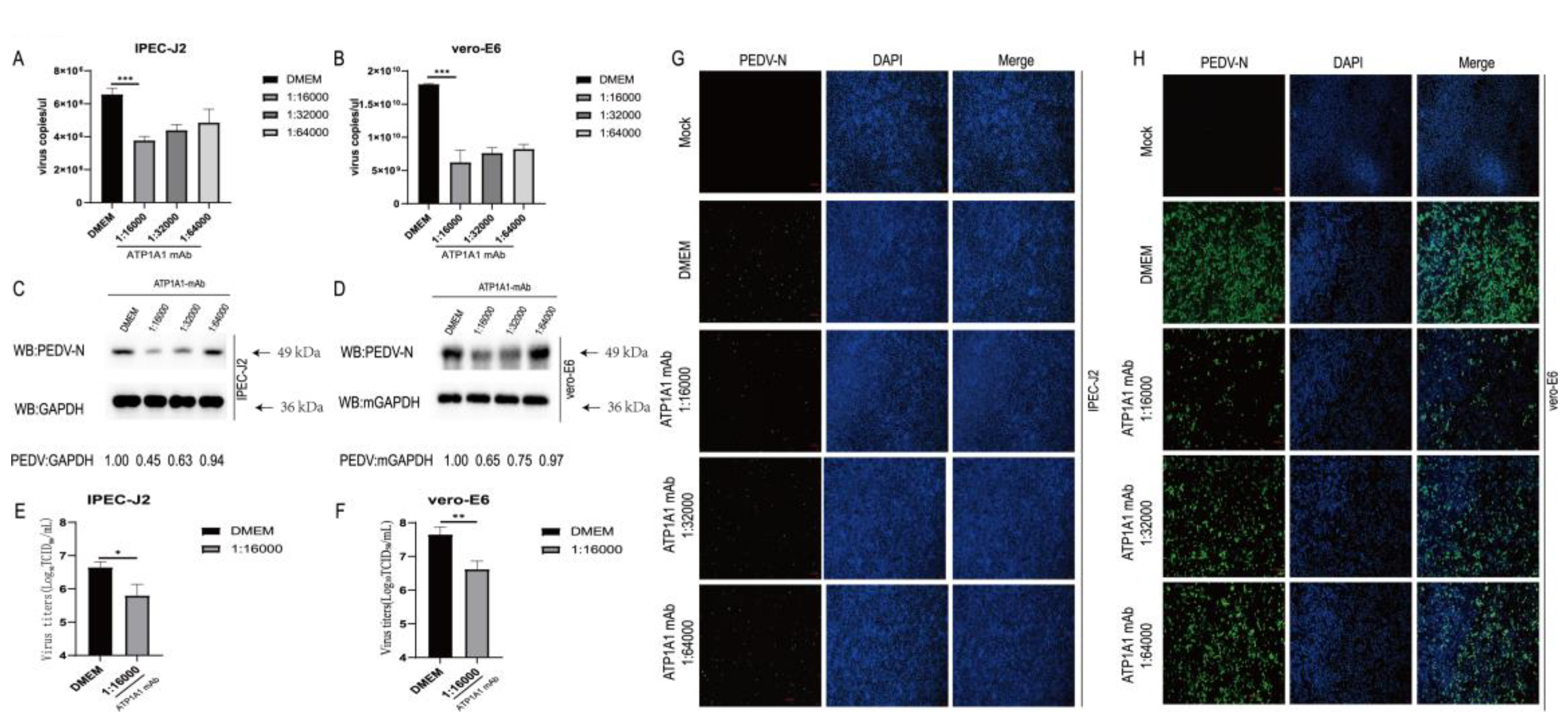
2.9. Monoclonal Antibody Pretreatment of ATP1A1 Effectively Inhibits PEDV Attachment
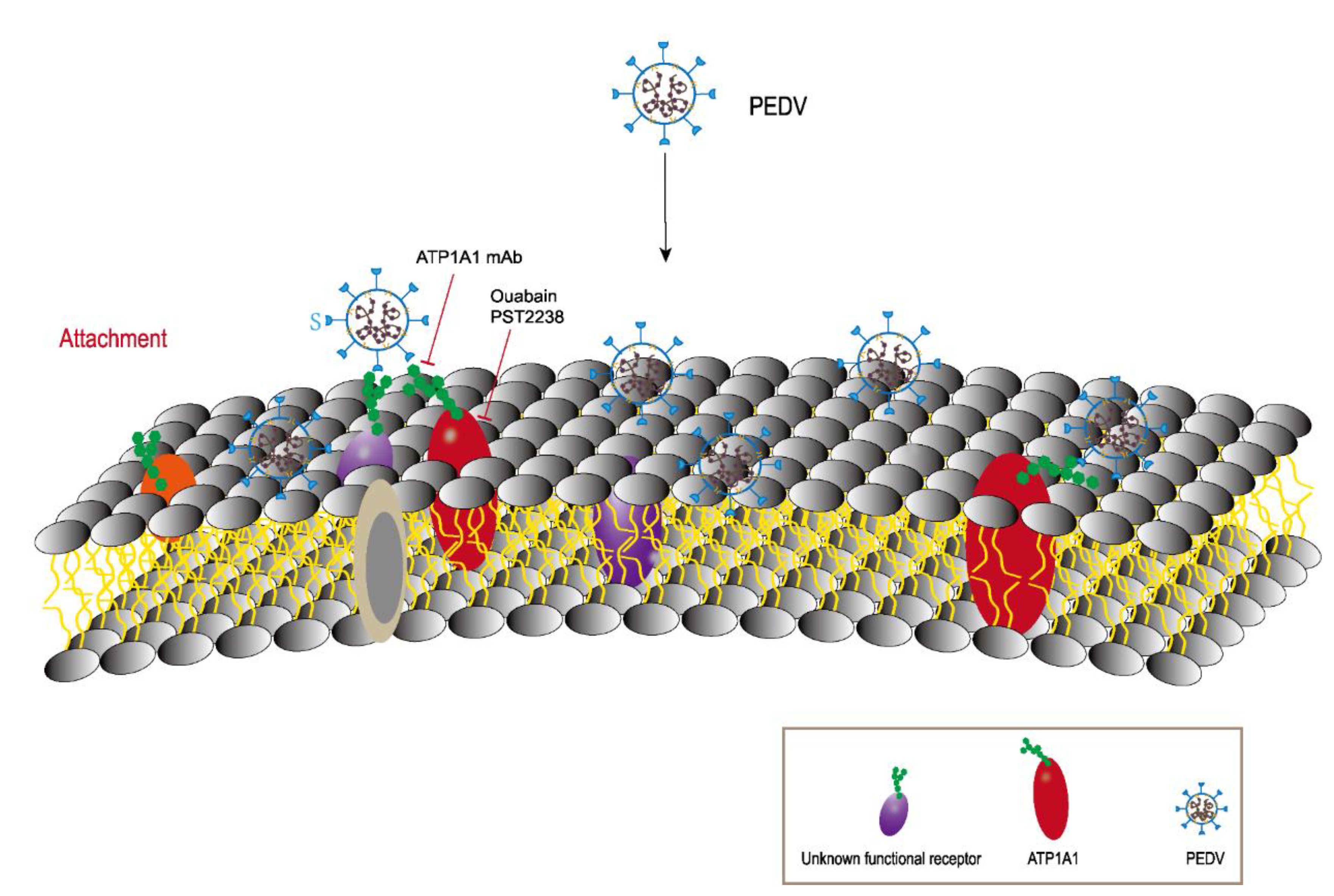
3. Discussion
4. Materials and Methods
4.1. Cells and Viruses
4.2. Antibodies, Inhibitors, and Reagents
4.3. Plasmid Constructs
4.4. Western Blot and IP
4.5. RNA Interference
4.6. Cell Viability Detection
4.7. Inhibitor Treatments
4.8. ATP1A1 mAb Inhibition Assay
4.9. Quantitative Real-Time PCR (RT-qPCR)
4.10. Immunofluorescence Assay and Confocal Microscopy
4.11. Flow Cytometry
4.12. TCID50 Assay
4.13. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.; Haas, M.; Liang, M.; Cai, T.; Tian, J.; Li, S.; Xie, Z. Ouabain Assembles Signaling Cascades through the Caveolar Na+/K+-ATPase. J. Biol. Chem. 2004, 279, 17250–17259. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, J.; Shi, D.; Shi, H.; Zhang, X.; Yuan, J.; Jiang, S.; Feng, L. Erratum to: Immunogenicity and antigenic relationships among spike proteins of porcine epidemic diarrhea virus subtypes G1 and G2. Arch. Virol. 2016, 161, 549. [Google Scholar] [CrossRef] [Green Version]
- Lee, C. Porcine epidemic diarrhea virus: An emerging and re-emerging epizootic swine virus. Virol. J. 2015, 12, 193. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Ge, X.; Chen, D.; Li, J.; Cai, Y.; Deng, J.; Zhou, L.; Guo, X.; Han, J.; Yang, H. The S Gene Is Necessary but Not Sufficient for the Virulence of Porcine Epidemic Diarrhea Virus Novel Variant Strain BJ2011C. J. Virol. 2018, 92, e00603-18. [Google Scholar] [CrossRef] [Green Version]
- Belouzard, S.; Millet, J.K.; Licitra, B.N.; Whittaker, G.R. Mechanisms of Coronavirus Cell Entry Mediated by the Viral Spike Protein. Viruses 2012, 4, 1011–1033. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Ma, Y.; Yang, Y.; Zheng, Y.; Shang, J.; Zhou, Y.; Jiang, S.; Du, L.; Li, J.; Li, F. Cell Entry of Porcine Epidemic Diarrhea Coronavirus Is Activated by Lysosomal Proteases. J. Biol. Chem. 2016, 291, 24779–24786. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-E.; Park, E.-S.; Yu, J.-E.; Rho, J.; Paudel, S.; Hyun, B.-H.; Yang, D.-K.; Shin, H.-J. Development of transgenic mouse model expressing porcine aminopeptidase N and its susceptibility to porcine epidemic diarrhea virus. Virus Res. 2015, 197, 108–115. [Google Scholar] [CrossRef]
- Deng, F.; Ye, G.; Liu, Q.; Navid, M.T.; Zhong, X.; Li, Y.; Wan, C.; Xiao, S.; He, Q.; Fu, Z.F.; et al. Identification and Comparison of Receptor Binding Characteristics of the Spike Protein of Two Porcine Epidemic Diarrhea Virus Strains. Viruses 2016, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Ge, J.; Li, Y. Porcine aminopeptidase N is a functional receptor for the PEDV coronavirus. Virology 2007, 365, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.-M.; Wang, B.; Zhou, J.; Huang, Y.-W. Aminopeptidase-N-independent entry of porcine epidemic diarrhea virus into Vero or porcine small intestine epithelial cells. Virology 2018, 517, 16–23. [Google Scholar] [CrossRef]
- Luo, L.; Wang, S.; Zhu, L.; Fan, B.; Liu, T.; Wang, L.; Zhao, P.; Dang, Y.; Sun, P.; Chen, J.; et al. Aminopeptidase N-null neonatal piglets are protected from transmissible gastroenteritis virus but not porcine epidemic diarrhea virus. Sci. Rep. 2019, 9, 13186. [Google Scholar] [CrossRef] [Green Version]
- Karger, A.; Mettenleiter, T.C. Identification of cell surface molecules that interact with pseudorabies virus. J. Virol. 1996, 70, 2138–2145. [Google Scholar] [CrossRef] [Green Version]
- Tamura, M.; Natori, K.; Kobayashi, M.; Miyamura, T.; Takeda, N. Interaction of Recombinant Norwalk Virus Particles with the 105-Kilodalton Cellular Binding Protein, a Candidate Receptor Molecule for Virus Attachment. J. Virol. 2000, 74, 11589–11597. [Google Scholar] [CrossRef] [Green Version]
- Holguera, J.; Villar, E.; Muñoz-Barroso, I. Identification of cellular proteins that interact with Newcastle Disease Virus and human Respiratory Syncytial Virus by a two-dimensional virus overlay protein binding assay (VOPBA). Virus Res. 2014, 191, 138–142. [Google Scholar] [CrossRef]
- Chang, J.-S.; Chi, S.-C. GHSC70 Is Involved in the Cellular Entry of Nervous Necrosis Virus. J. Virol. 2015, 89, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Conner, S.D.; Schmid, S.L. Regulated portals of entry into the cell. Nature 2003, 422, 37–44. [Google Scholar] [CrossRef]
- Ostrom, R.S.; Gregorian, C.; Drenan, R.M.; Xiang, Y.; Regan, J.W.; Insel, P.A. Receptor Number and Caveolar Co-localization Determine Receptor Coupling Efficiency to Adenylyl Cyclase. J. Biol. Chem. 2001, 276, 42063–42069. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Cai, T. Na+- K+- ATPase- Mediated Signal Transduction: From Protein Interaction to Cellular Function. Mol. Interv. 2003, 3, 157–168. [Google Scholar] [CrossRef]
- Capendeguy, O.; Horisberger, J.-D. The role of the third extracellular loop of the Na+,K+-ATPase α subunit in a luminal gating mechanism. J. Physiol. 2005, 565, 207–218. [Google Scholar] [CrossRef]
- HA, K. Novel progesterone target genes identified by an improved differential display technique suggest that progestin- induced growth inhibition of breast cancer cells coincides with enhancement of differentiation. J. Biol. Chem. 1997, 272, 16637–16643. [Google Scholar]
- Mijatovic, T.; Roland, I.; Van Quaquebeke, E.; Nilsson, B.; Mathieu, A.; Van Vynckt, F.; Darro, F.; Blanco, G.; Facchini, V.; Kiss, R. The α1 subunit of the sodium pump could represent a novel target to combat non-small cell lung cancers. J. Pathol. 2007, 212, 170–179. [Google Scholar] [CrossRef]
- Wang, Y.; Zhan, Y.; Xu, R.; Shao, R.; Jiang, J.; Wang, Z. Src mediates extracellular signal-regulated kinase 1/2 activation and autophagic cell death induced by cardiac glycosides in human non-small cell lung cancer cell lines. Mol. Carcinog. 2014, 54, E26–E34. [Google Scholar] [CrossRef]
- Xu, Z.-W.; Wang, F.-M.; Gao, M.-J.; Chen, X.-Y.; Hu, W.-L.; Xu, R.-C. Targeting the Na+ /K+-ATPase alpha1 Subunit of Hepatoma HepG2 Cell Line to Induce Apoptosis and Cell Cycle Arresting. Biol. Pharm. Bull. 2010, 33, 743–751. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, L.; Xu, L.; Wang, P.; Jiang, Y.; Yong, P.; Zhang, C.; Zhang, H.; Meng, Z.; Yang, P. Na+/K+-ATPase α1 subunit, a novel therapeutic target for hepatocellular carcinoma. Oncotarget 2015, 6, 28183–28193. [Google Scholar] [CrossRef] [Green Version]
- Burkard, C.; Verheije, M.H.; Haagmans, B.L.; van Kuppeveld, F.J.; Rottier, P.J.M.; Bosch, B.-J.; de Haan, C.A.M. ATP1A1-Mediated Src Signaling Inhibits Coronavirus Entry into Host Cells. J. Virol. 2015, 89, 4434–4448. [Google Scholar] [CrossRef] [Green Version]
- Lingemann, M.; McCarty, T.; Liu, X.; Buchholz, U.J.; Surman, S.; Martin, S.E.; Collins, P.L.; Munir, S. The alpha-1 subunit of the Na+,K+-ATPase (ATP1A1) is required for macropinocytic entry of respiratory syncytial virus (RSV) in human respiratory epithelial cells. PLOS Pathog. 2019, 15, e1007963. [Google Scholar] [CrossRef] [Green Version]
- Mukhopadhyay, R.; Venkatadri, R.; Katsnelson, J.; Arav-Boger, R. Digitoxin Suppresses Human Cytomegalovirus Replication via Na +, K +/ATPase α1 Subunit-Dependent AMP-Activated Protein Kinase and Autophagy Activation. J. Virol. 2018, 92, e01861-17. [Google Scholar] [CrossRef] [Green Version]
- Upham, J.P.; Pickett, D.; Irimura, T.; Anders, E.M.; Reading, P.C. Macrophage Receptors for Influenza A Virus: Role of the Macrophage Galactose-Type Lectin and Mannose Receptor in Viral Entry. J. Virol. 2010, 84, 3730–3737. [Google Scholar] [CrossRef] [Green Version]
- Wu, E.; Fernandez, J.; Fleck, S.K.; Von Seggern, D.J.; Huang, S.; Nemerow, G.R. A 50-kDa Membrane Protein Mediates Sialic Acid-Independent Binding and Infection of Conjunctival Cells by Adenovirus Type 37. Virology 2001, 279, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Cherniavsky-Lev, M.; Golani, O.; Karlish, S.J.; Garty, H. Ouabain-induced Internalization and Lysosomal Degradation of the Na+/K+-ATPase. J. Biol. Chem. 2014, 289, 1049–1059. [Google Scholar] [CrossRef] [Green Version]
- Wicht, O.; Li, W.; Willems, L.; Meuleman, T.J.; Wubbolts, R.W.; van Kuppeveld, F.J.M.; Rottier, P.J.M.; Bosch, B.J. Proteolytic Activation of the Porcine Epidemic Diarrhea Coronavirus Spike Fusion Protein by Trypsin in Cell Culture. J. Virol. 2014, 88, 7952–7961. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Saif, L.J.; Wang, Q. Porcine epidemic diarrhea virus (PEDV): An update on etiology, transmission, pathogenesis, and prevention and control. Virus Res. 2020, 286, 198045. [Google Scholar] [CrossRef]
- Dimitrov, D.S. Virus entry: Molecular mechanisms and biomedical applications. Nat. Rev. Genet. 2004, 2, 109–122. [Google Scholar] [CrossRef]
- Huan, C.-C.; Wang, Y.; Ni, B.; Wang, R.; Huang, L.; Ren, X.-F.; Tong, G.-Z.; Ding, C.; Fan, H.-J.; Mao, X. Porcine epidemic diarrhea virus uses cell-surface heparan sulfate as an attachment factor. Arch. Virol. 2015, 160, 1621–1628. [Google Scholar] [CrossRef]
- Wanitchang, A.; Saenboonrueng, J.; Srisutthisamphan, K.; Jongkaewwattana, A. Characterization of influenza A virus pseudotyped with the spike protein of porcine epidemic diarrhea virus. Arch. Virol. 2018, 163, 3255–3264. [Google Scholar] [CrossRef]
- Contreras, R.G. Relationship between Na+,K+-ATPase and cell attachment. J. Cell Sci. 1999, 112, 4223–4232. [Google Scholar] [CrossRef]
- Jeon, J.H.; Lee, C. Cellular cholesterol is required for porcine nidovirus infection. Arch. Virol. 2017, 162, 3753–3767. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, Y.; Yang, Q. Transferrin receptor 1 levels at the cell surface influence the susceptibility of newborn piglets to PEDV infection. PLOS Pathog. 2020, 16, e1008682. [Google Scholar] [CrossRef]
- Luo, X.; Guo, L.; Zhang, J.; Xu, Y.; Gu, W.; Feng, L.; Wang, Y. Tight Junction Protein Occludin Is a Porcine Epidemic Diarrhea Virus Entry Factor. J. Virol. 2017, 91, e00202-17. [Google Scholar] [CrossRef] [Green Version]
- Davis, P.J.; Lin, H.-Y.; Hercbergs, A.; Keating, K.A.; Mousa, S.A. Coronaviruses and Integrin αvβ3: Does Thyroid Hormone Modify the Relationship? Endocr. Res. 2020, 45, 210–215. [Google Scholar] [CrossRef]
- Li, C.; Su, M.; Yin, B.; Guo, D.; Wei, S.; Kong, F.; Feng, L.; Wu, R.; Sun, D. Integrin αvβ3 enhances replication of porcine epidemic diarrhea virus on Vero E6 and porcine intestinal epithelial cells. Vet. Microbiol. 2019, 237, 108400. [Google Scholar] [CrossRef]
- Shan, Z.; Yin, J.; Wang, Z.; Chen, P.; Li, Y.; Tang, L. Identification of the functional domain of the porcine epidemic diarrhoea virus receptor. J. Gen. Virol. 2015, 96, 2656–2660. [Google Scholar] [CrossRef]
- Sun, Y.-G.; Li, R.; Jiang, L.; Qiao, S.; Zhi, Y.; Chen, X.-X.; Xie, S.; Wu, J.; Li, X.; Deng, R.; et al. Characterization of the interaction between recombinant porcine aminopeptidase N and spike glycoprotein of porcine epidemic diarrhea virus. Int. J. Biol. Macromol. 2018, 117, 704–712. [Google Scholar] [CrossRef]
- Skou, J.C. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochim. Biophys. Acta 1957, 23, 394–401. [Google Scholar] [CrossRef]
- Aizman, O.; Aperia, A. Na,K-ATPase as a Signal Transducer. Ann. N. Y. Acad. Sci. 2003, 986, 489–496. [Google Scholar] [CrossRef]
- Rajasekaran, A.K.; Rajasekaran, S.A. Role of Na-K-ATPase in the assembly of tight junctions. Am. J. Physiol. Physiol. 2003, 285, F388–F396. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, S.A. Na,K-ATPase β-Subunit Is Required for Epithelial Polarization, Suppression of Invasion, and Cell Motility. Mol. Biol. Cell 2001, 12, 279–295. [Google Scholar] [CrossRef]
- Cui, X.; Xie, Z. Protein Interaction and Na/K-ATPase-Mediated Signal Transduction. Molecules 2017, 22, 990. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Zhang, Z.; Xie, J.X.; Li, X.; Tian, J.; Cai, T.; Cui, H.; Ding, H.; Shapiro, J.I.; Xie, Z. Na/K-ATPase Mimetic pNaKtide Peptide Inhibits the Growth of Human Cancer Cells. J. Biol. Chem. 2011, 286, 32394–32403. [Google Scholar] [CrossRef] [Green Version]
- Aizman, O.; Uhlén, P.; Lal, M.; Brismar, H.; Aperia, A. Ouabain, a steroid hormone that signals with slow calcium oscillations. Proc. Natl. Acad. Sci. USA 2001, 98, 13420–13424. [Google Scholar] [CrossRef] [Green Version]
- Dodson, A.W.; Taylor, T.J.; Knipe, D.M.; Coen, D.M. Inhibitors of the sodium potassium ATPase that impair herpes simplex virus replication identified via a chemical screening approach. Virology 2007, 366, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Su, C.-T.; Hsu, J.T.-A.; Hsieh, H.-P.; Lin, P.-H.; Chen, T.-C.; Kao, C.-L.; Lee, C.-N.; Chang, S.-Y. Anti-HSV activity of digitoxin and its possible mechanisms. Antivir. Res. 2008, 79, 62–70. [Google Scholar] [CrossRef]
- Grosso, F.; Stoilov, P.; Lingwood, C.; Brown, M.; Cochrane, A. Suppression of Adenovirus Replication by Cardiotonic Steroids. J. Virol. 2017, 91, e01623-16. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, P.; Torielli, L.; Ferrandi, M.; Padoani, G.; Duzzi, L.; Florio, M.; Conti, F.; Melloni, P.; Vesci, L.; Corsico, N.; et al. PST2238: A new antihypertensive compound that antagonizes the long-term pressor effect of ouabain. J. Pharmacol. Exp. Ther. 1998, 285, 83–94. [Google Scholar]
- Schmidt, N.; Lareau, C.A.; Keshishian, H.; Ganskih, S.; Schneider, C.; Hennig, T.; Melanson, R.; Werner, S.; Wei, Y.; Zimmer, M.; et al. The SARS-CoV-2 RNA–protein interactome in infected human cells. Nat. Microbiol. 2020, 6, 339–353. [Google Scholar] [CrossRef]
- Aknouch, I.; Sridhar, A.; Freeze, E.; Giugliano, F.P.; van Keulen, B.J.; Romijn, M.; Calitz, C.; García-Rodríguez, I.; Mulder, L.; Wildenberg, M.E.; et al. Human milk inhibits some enveloped virus infections, including SARS-CoV-2, in an intestinal model. Life Sci. Alliance 2022, 5, e202201432. [Google Scholar] [CrossRef]
- García-Dorival, I.; Wu, W.; Dowall, S.; Armstrong, S.; Touzelet, O.; Wastling, J.; Barr, J.N.; Matthews, D.; Carroll, M.; Hewson, R.; et al. Elucidation of the Ebola Virus VP24 Cellular Interactome and Disruption of Virus Biology through Targeted Inhibition of Host-Cell Protein Function. J. Proteome Res. 2014, 13, 5120–5135. [Google Scholar] [CrossRef]
- Krimmling, T.; Beineke, A.; Schwegmann-Weßels, C. Infection of porcine precision cut intestinal slices by transmissible gastroenteritis coronavirus demonstrates the importance of the spike protein for enterotropism of different virus strains. Vet. Microbiol. 2017, 205, 1–5. [Google Scholar] [CrossRef]
- Sánchez, C.M.; Izeta, A.; Sánchez-Morgado, J.M.; Alonso, S.; Sola, I.; Balasch, M.; Plana-Durán, J.; Enjuanes, L. Targeted recombination demonstrates that the spike gene of transmissible gastroenteritis coronavirus is a determinant of its enteric tropism and virulence. J. Virol. 1999, 73, 7607–7618. [Google Scholar] [CrossRef] [Green Version]
- Bosch, B.J.; van der Zee, R.; de Haan, C.A.; Rottier, P.J.M. The Coronavirus Spike Protein Is a Class I Virus Fusion Protein: Structural and Functional Characterization of the Fusion Core Complex. J. Virol. 2003, 77, 8801–8811. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Liu, Y.; Wang, S.; Zhang, L.; Liang, P.; Wang, L.; Dong, J.; Song, C. Molecular Characteristics and Pathogenicity of Porcine Epidemic Diarrhea Virus Isolated in Some Areas of China in 2015–2018. Front. Vet. Sci. 2020, 7, 607662. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, Y.; Gao, X.; Wang, Y.; Jia, X.; Xiao, Y.; Wang, T.; Li, X.; Tian, K. Development of a Neutralizing Monoclonal Antibody Against Porcine Epidemic Diarrhea Virus S1 Protein. Monoclon. Antibodies Immunodiagn. Immunother. 2016, 35, 37–40. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Liu, R.; Shuai, L.; Wang, X.; Luo, J.; Wang, C.; Chen, W.; Wang, X.; Ge, J.; et al. Metabotropic glutamate receptor subtype 2 is a cellular receptor for rabies virus. PLOS Pathog. 2018, 14, e1007189. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Reed, L.J. A simple method of estimating fifty peg cent endpoints. Am. J. Hyjiene 1938, 27, 493–497. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) |
|---|---|
| ATP1A1-F a | CGGGGTACCATGGGGAAGGGGGTTGGAC |
| ATP1A1-CT-F a | CGGGGTACCATGCTGGATGACAACTTCGCCTC |
| ATP1A1-1R a | CGGGATATCCTAGGCTTGCTGAGGAACCATGTTTTTG |
| ATP1A1-2R a | CGGGATATCCTACACTGCCCTGTTACAAAGACCTG |
| ATP1A1-3R a | CGGGATATCCTAGGCATCCCTGGGGTTCACC |
| ATP1A1-4R a | CGGGATATCCTAGACGGTGCCCAGGGG |
| ATP1A1-R a | CGGGATATCCTAGTAGTAGGTTTCCTTCTCCACCC |
| PEDV-N-F b | ATGATCTGGTGGCTGCTGTC |
| PEDV-N-R b | CTTCGAAGTGGCCCTGGATT |
| Porcine-ATP1A1-F b | CTTGAGCCGAGGCTTAACAC |
| Porcine-ATP1A1-R b | GAATGCCATAGGCCAAGAAA |
| Monkey-ATP1A1-F b | CAGCAGTGGACCTATGAGCA |
| Monkey-ATP1A1-R b | CATTCCAGGGCAGTAGGAAA |
| Porcine-GAPDH-F b | GATGCTGGTGCTGAGTATGT |
| Porcine-GAPDH-R b | GGCAGAGATGATGACCCTTT |
| Monkey-GAPDH-F b | CGAGATCCCTCCAAAATCAA |
| Monkey-GAPDH-R b | TGACGATCTTGAGGCTGTTG |
| Target Genes | (5′-3′) (Sense) | (5′-3′) (Antisense) |
|---|---|---|
| si-NC si-ATP1A1-A si-ATP1A1-B si-mNC si-mATP1A1-A si-mATP1A1-B | UUCUCCGAACGUGUCACGUTT CAGGAAGAACUGCCUUGUGAATT GCAGCUGGAUGACAUCUUGAATT UUCUCCGAACGUGUCACGUTT GAUUCGAAAUGGUGAGAAATT GUGAAGGAGAUGAGAGAAATT | ACGUGACACGUUCGGAGAATT UUCACAAGGCAGUUCUUCCUGTT UUCAAGAUGUCAUCCAGCUGCTT ACGUGACACGUUCGGAGAATT UUUCUCACCAUUUCGAAUCT UUUCUCUCAUCUCCUUCACTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, M.; Liu, X.; Liang, T.; Ban, Y.; Liu, Y.; Zhang, L.; Xu, Z.; Song, C. The Alpha-1 Subunit of the Na+/K+-ATPase (ATP1A1) Is a Host Factor Involved in the Attachment of Porcine Epidemic Diarrhea Virus. Int. J. Mol. Sci. 2023, 24, 4000. https://doi.org/10.3390/ijms24044000
Xiong M, Liu X, Liang T, Ban Y, Liu Y, Zhang L, Xu Z, Song C. The Alpha-1 Subunit of the Na+/K+-ATPase (ATP1A1) Is a Host Factor Involved in the Attachment of Porcine Epidemic Diarrhea Virus. International Journal of Molecular Sciences. 2023; 24(4):4000. https://doi.org/10.3390/ijms24044000
Chicago/Turabian StyleXiong, Moukang, Xianhui Liu, Tairun Liang, Yanfang Ban, Yanling Liu, Leyi Zhang, Zheng Xu, and Changxu Song. 2023. "The Alpha-1 Subunit of the Na+/K+-ATPase (ATP1A1) Is a Host Factor Involved in the Attachment of Porcine Epidemic Diarrhea Virus" International Journal of Molecular Sciences 24, no. 4: 4000. https://doi.org/10.3390/ijms24044000