
Dynamic Association of ESCRT-II Proteins with ESCRT-I and ESCRT-III Complexes during Phagocytosis of Entamoeba histolytica
 , , , and
, , , and
Abstract
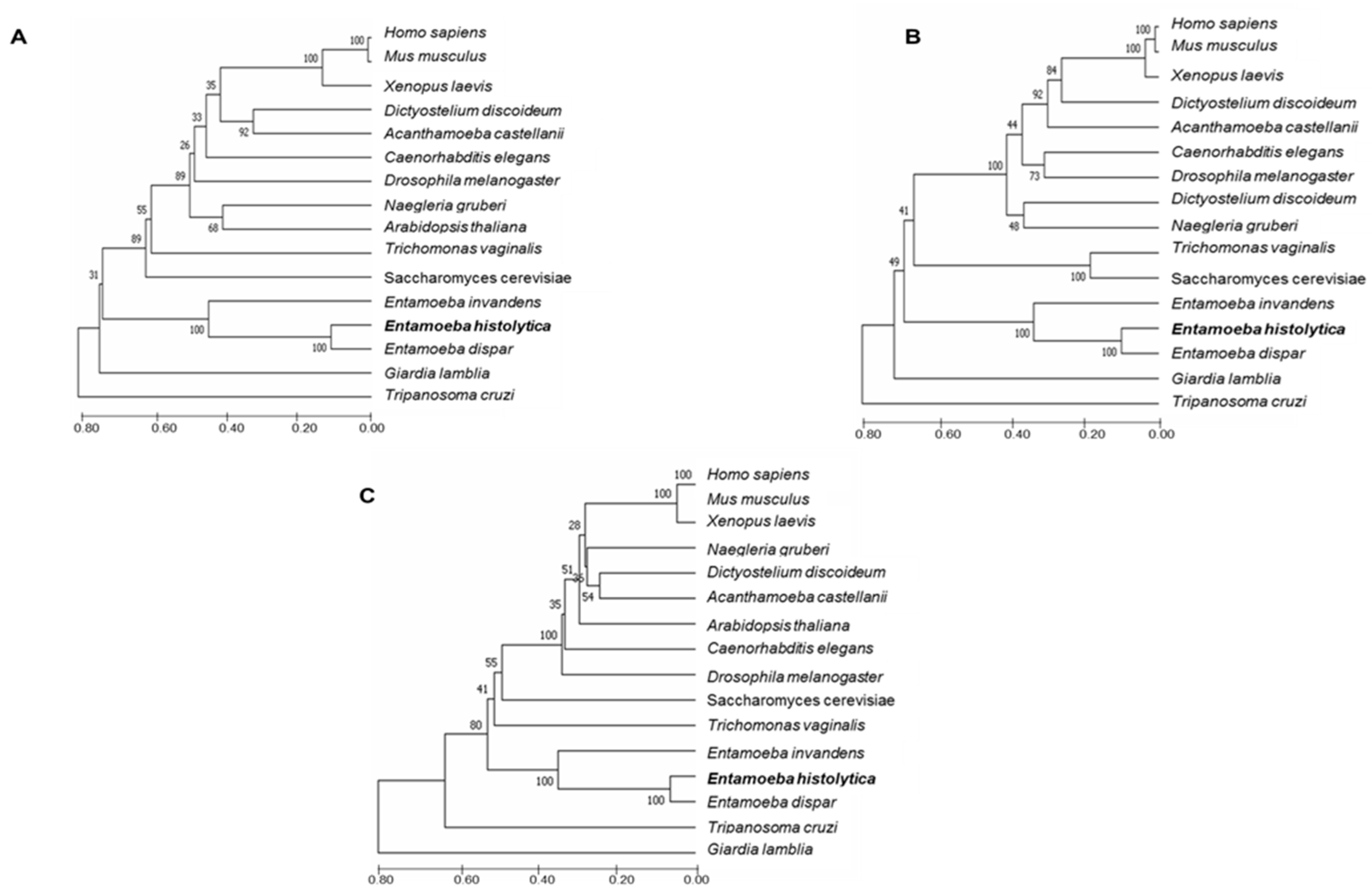
:1. Introduction
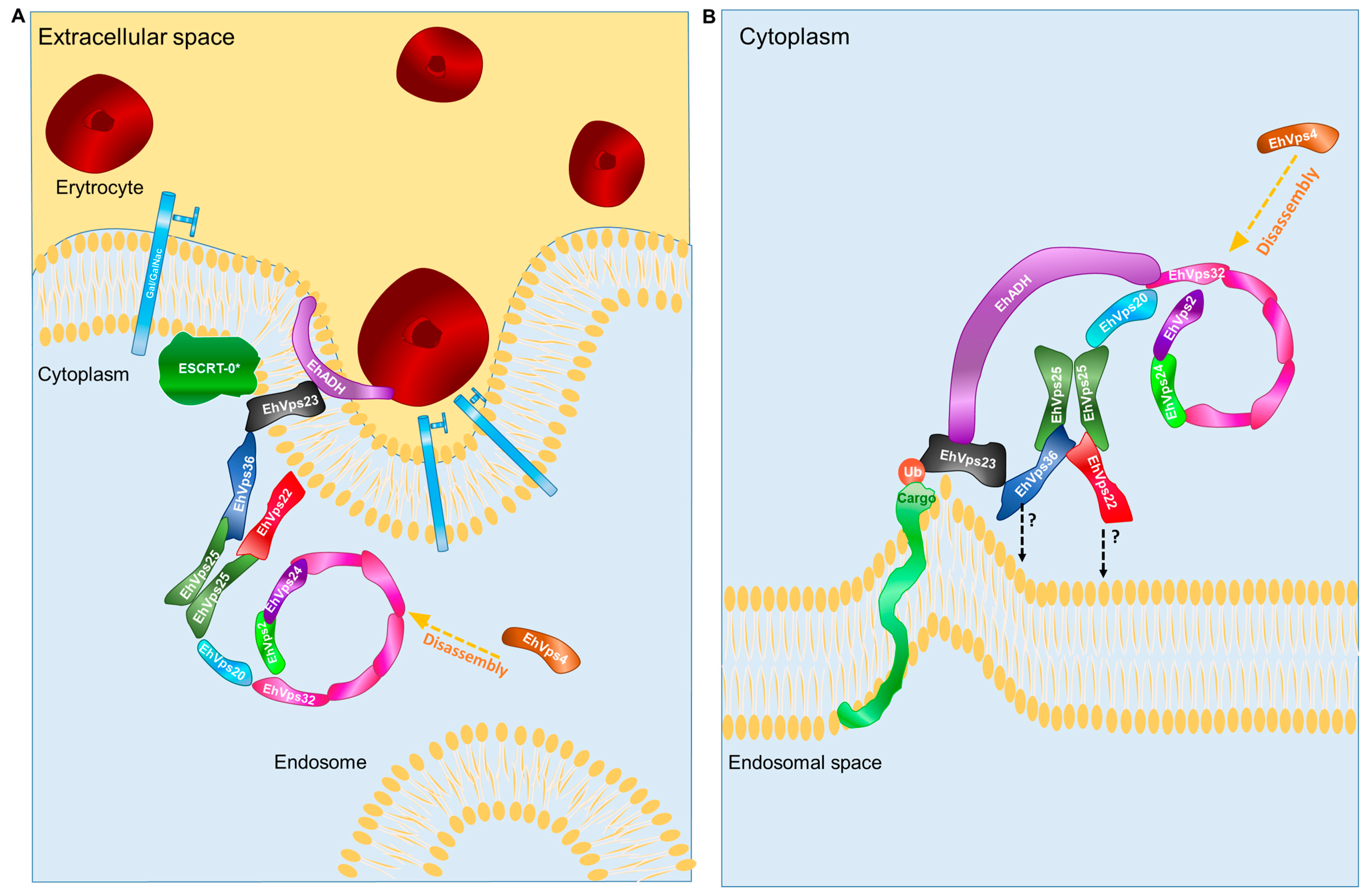
2. Results
2.1. EhVps22, EhVps25, and EhVps36 Possess Functional WH-WH Domains
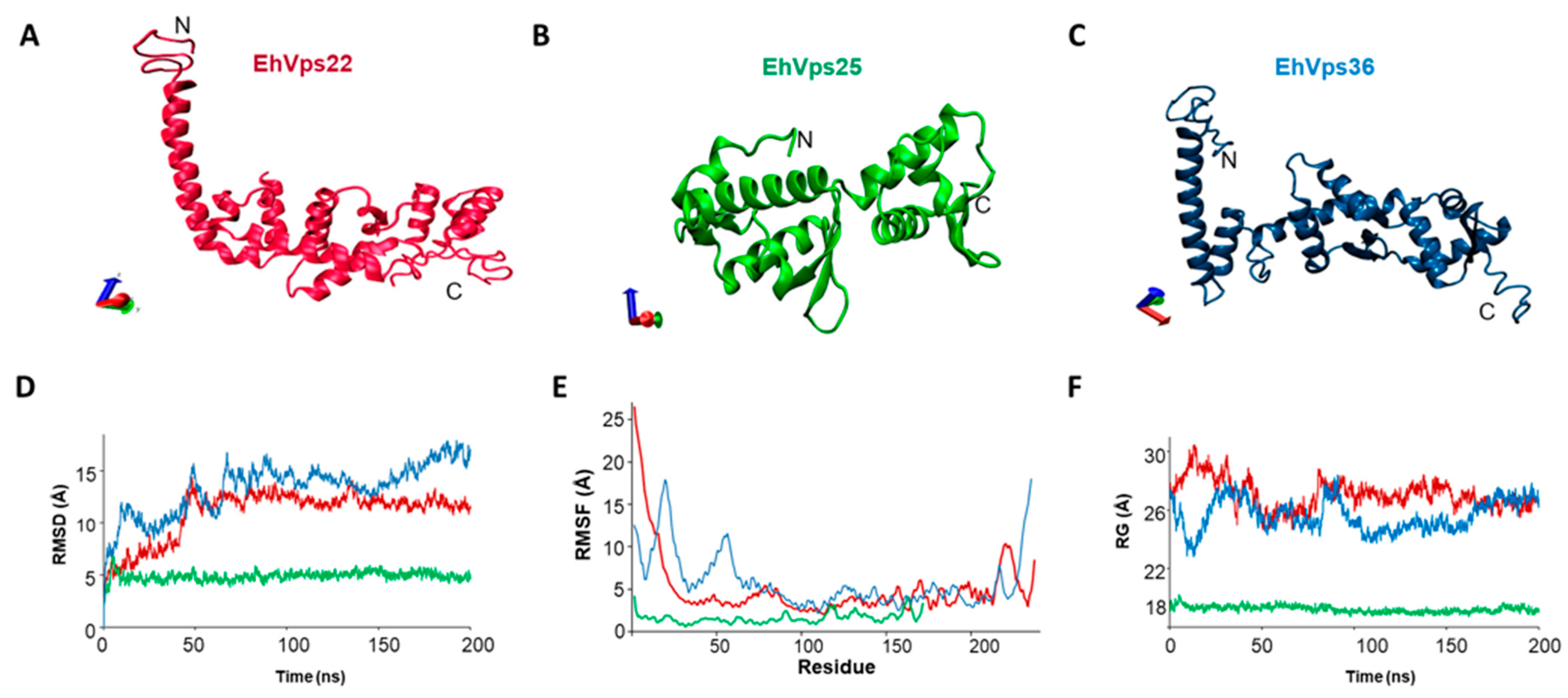
2.2. The Three-Dimensional Structures of EhVps22, EhVps25, and EhVps36 Proteins Present high HOMOLOGY with Their Orthologues
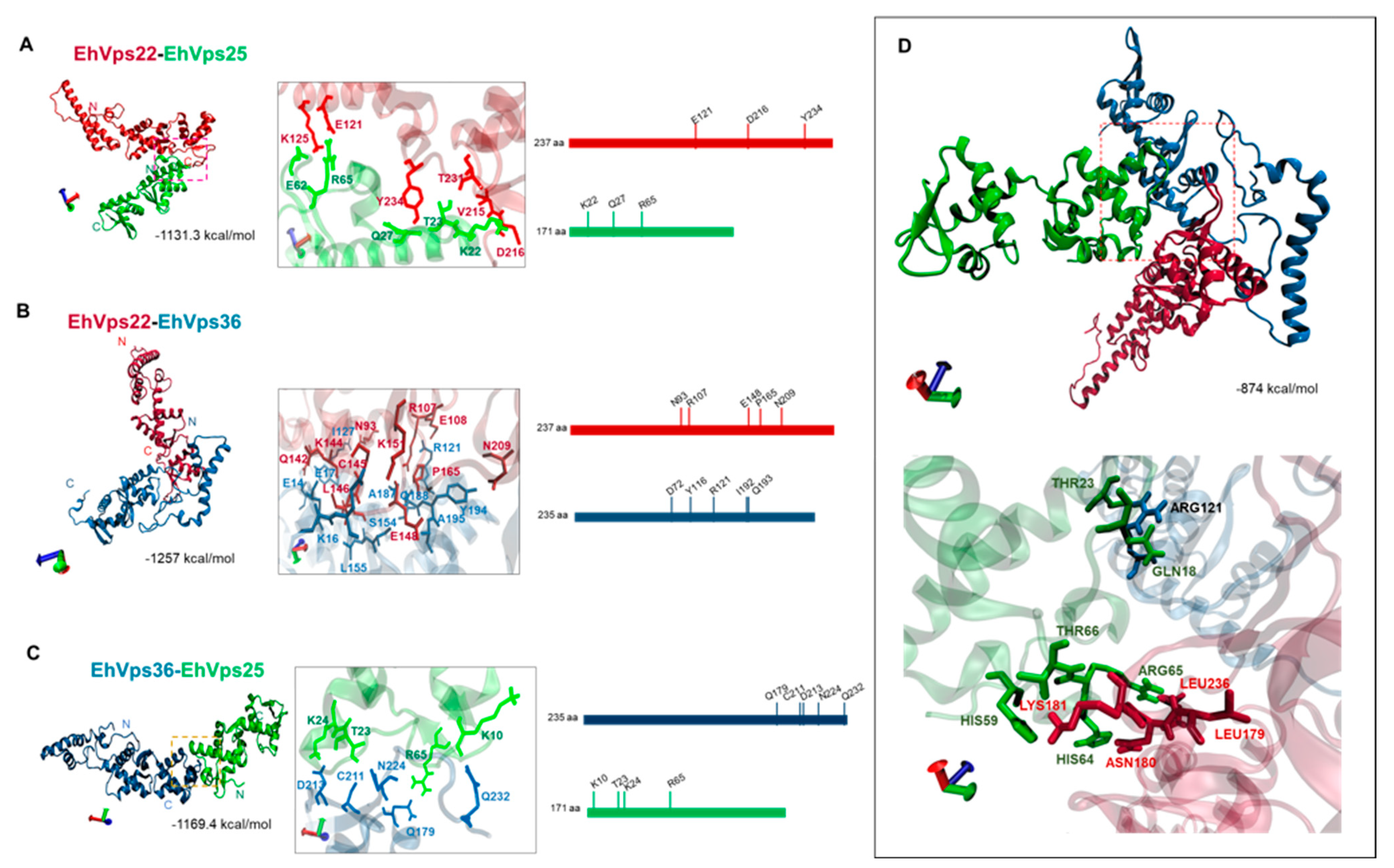
2.3. WH-WH Domains Key Regions for Interaction between EhVps22, EhVps25, and EhVps36 by In Silico Analyses
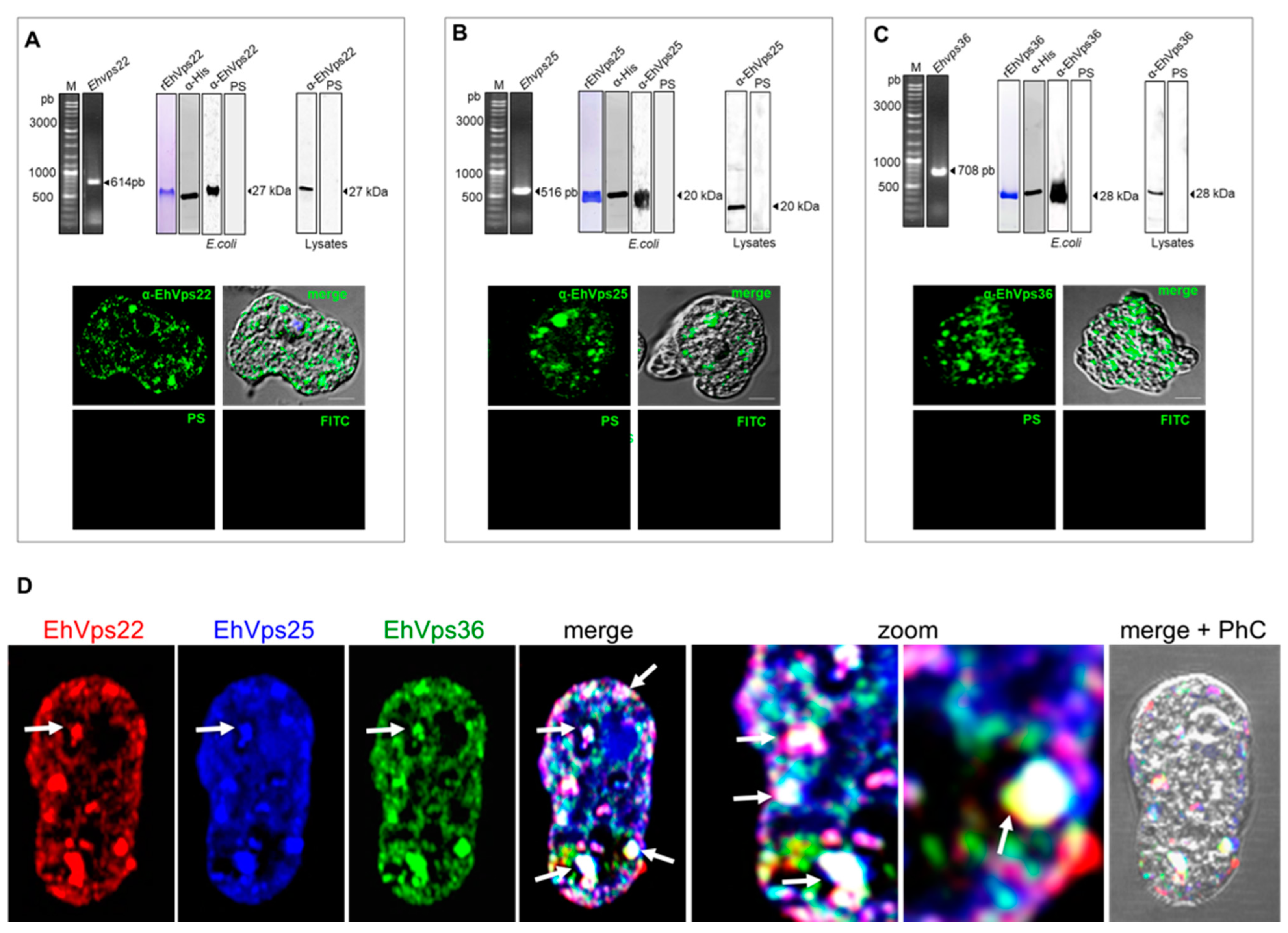
2.4. The ESCRT-II Proteins Are Located in the Cytoplasm of the Trophozoites, and Co-Localization Was Observed in Vesicles and Near the Plasma Membrane
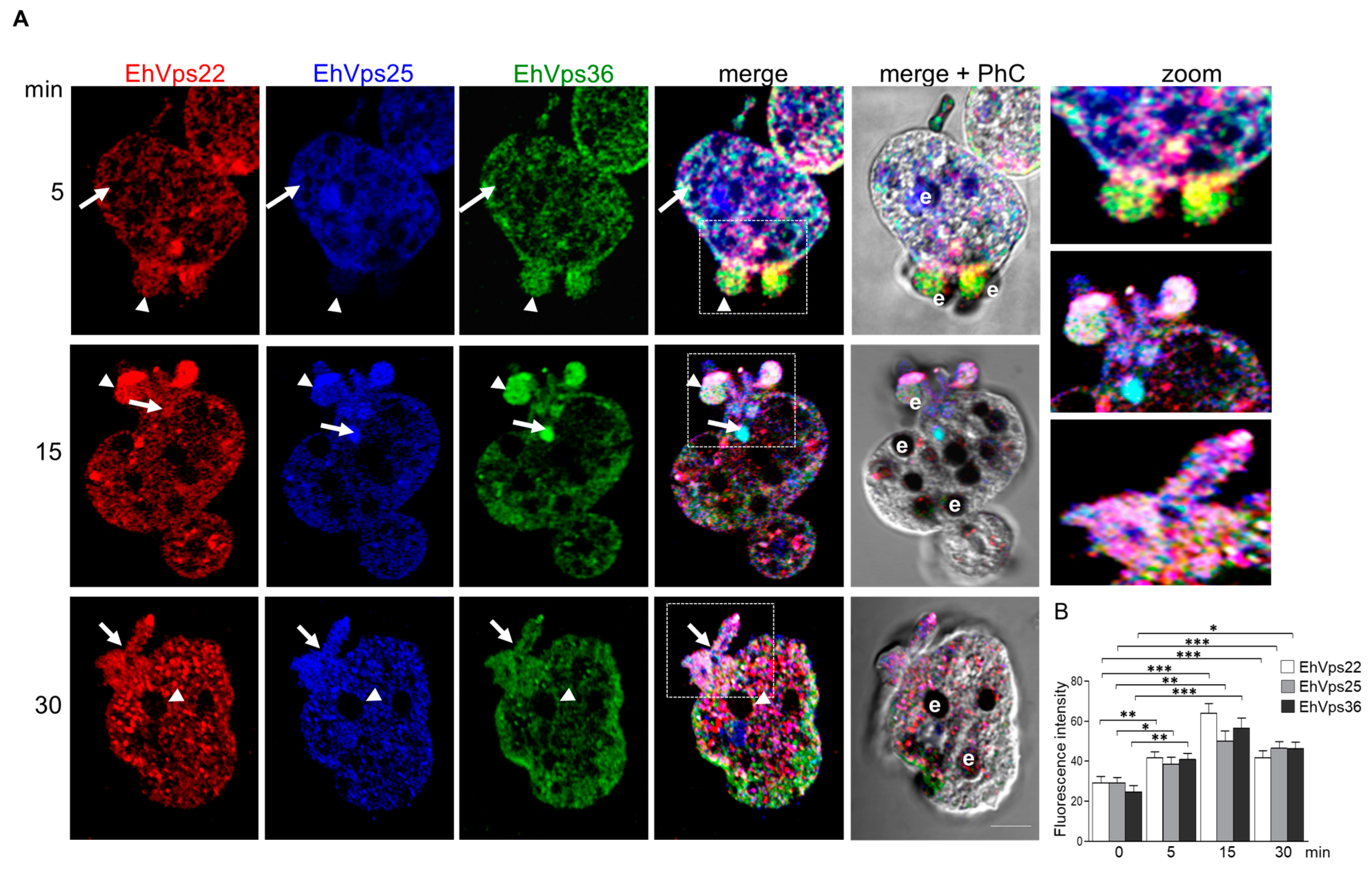
2.5. The ESCRT-II Proteins Change Their Location during Phagocytosis
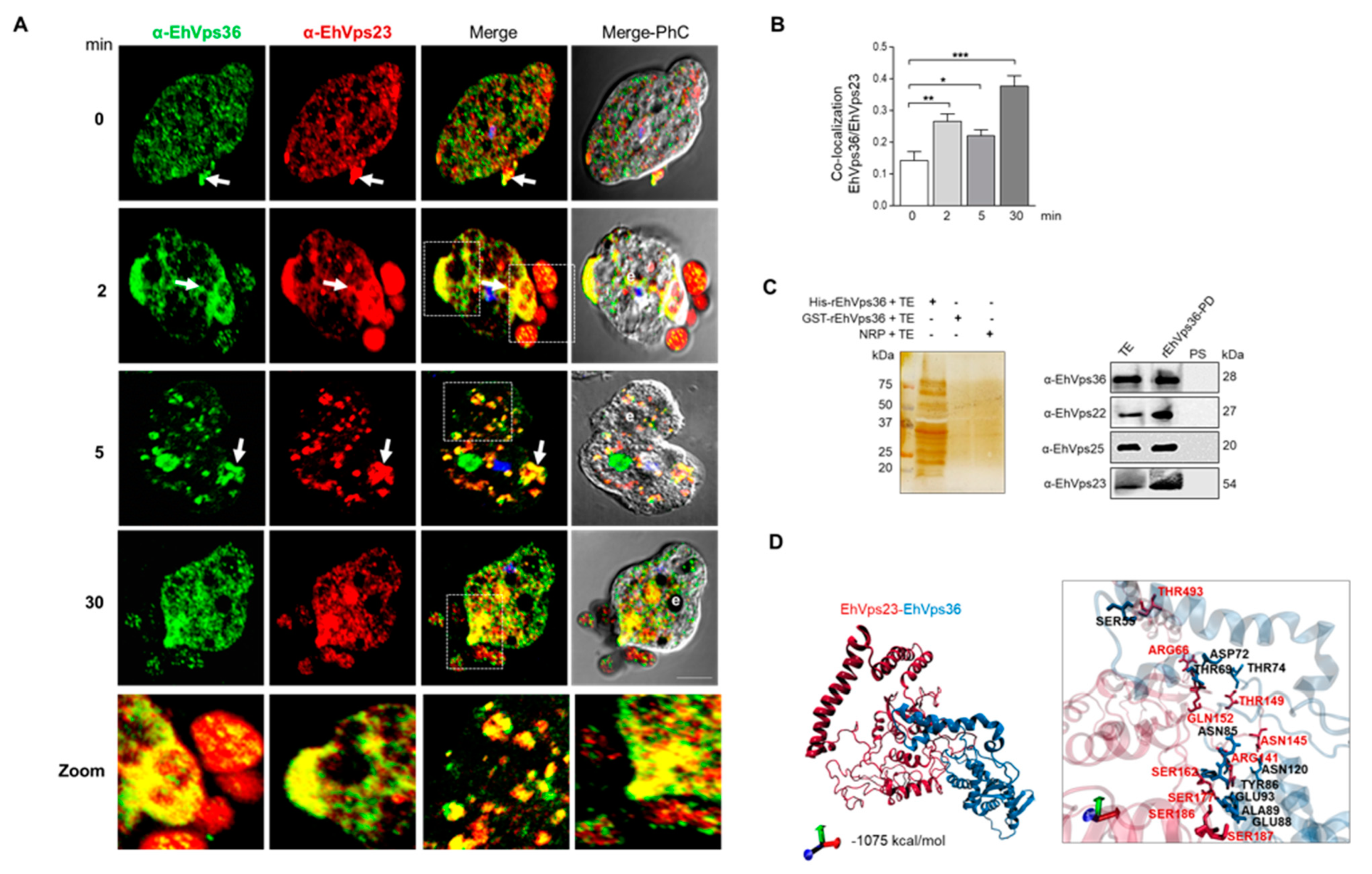
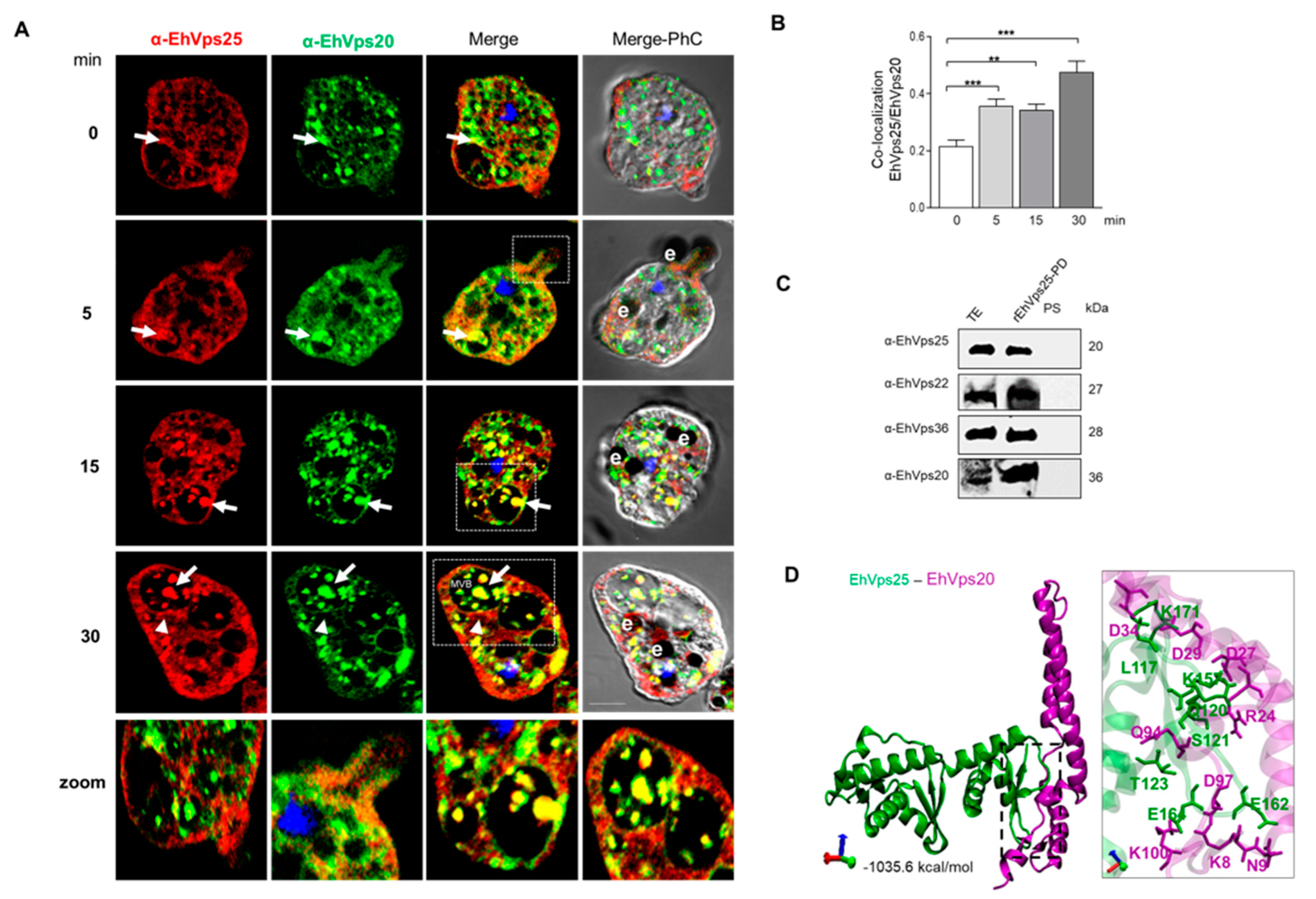
2.6. The ESCRT-II Proteins BIND ESCRT-I and ESCRT-III Complexes, and Their Interaction Increases during Phagocytosis
2.7. Mass Spectrometry Analysis Reveals the Presence of Proteins Involved in Phagocytosis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Recombinant Protein | Access Number | Protein | Function | Reference |
|---|---|---|---|---|
| EhVps22 | EHI_181240 | Rab B GTPase | Phagocytosis | Javier-Reyna et al., 2019 [34] |
| EHI_137860 | EhVps25 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_045320 | EhVps36 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_025270 | EhVps29 | Phagocytosis (Retromer complex) | ----------- | |
| EHI_030820 | ARP2/3 complex 20 kDa subunit | Phagocytic cups and cytoskeleton | Mrigya-Babuta et al., 2015 [38] | |
| EHI_182900 | Actin | Cytoskeleton | Parimita Rath et al., 2020 [38] | |
| EHI_049920 | Tubulin | Cytoskeleton (microtubule nucleation) | Gilchrist et al., 1999 [36] | |
| EHI_136160 | Calreticulin putative | Development of amoebic liver abscesses | Enrique-González et al., 2011 [32] | |
| EHI_012270 | Gal/GalNAc lectin heavy subunit | Adherense and cytolysis | Gilchrist et al., 1999 [36] | |
| EHI_052860 | Heat shock protein 70 | Amoebic pathogenicity by protection against oxidative and nitrosative stress | Gilchrist et al., 1999 [36] | |
| EHI_098290 | Hexokinase 1 | Flux control of glycolysis | Saucedo-Mendiola et al., 2014 [37] | |
| EhVps25 | EHI_181240 | Rab B GTPase | Phagocytosis | Javier-Reyna et al., 2019 [34] |
| EHI_131120 | EhVps22 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_045320 | EhVps36 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_066730 | EhVps20 | Phagocytosis (ESCRT-III) | Ávalos-Padilla et al., 2018 [22] | |
| EHI_169820 | EhVps32 | Phagocytosis (ESCRT-III) | Ávalos-Padilla et al., 2018 [22] | |
| EHI_030820 | ARP2/3 complex 20 kDa subunit | Phagocytic cups and cytoskeleton | Mrigya-Babuta et al., 2015 [38] | |
| EHI_167300 | Grainin 1 | Vesicular maturation and exocytosis and Programmed cell death | Tovar et al., 2000 [41] | |
| EhVps36 | EHI_181240 | Rab B GTPase | Phagocytosis | Javier-Reyna et al., 2019 [34] |
| EHI_135460 | EhVps23 | Phagocytosis (ESCRT-I) | Galindo-Olea et al., 2021 [15] | |
| EHI_131120 | EhVps22 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_137860 | EhVps25 | Phagocytosis (ESCRT-II) | López-Reyes et al., 2010 [18] | |
| EHI_169820 | EhVps32 | Phagocytosis (ESCRT-III) | Ávalos-Padilla et al., 2018 [22] | |
| EHI_023500 | Calmodulin, putative | Phagocytosis and actin dynamics | Rout et al., 2011 [40] | |
| EHI_120360 | Grainin | Associated with reduced virulence | Tovar et al., 2000 [41] | |
| EHI_201510 | Clathrin heavy chain, putative | Secretion | Rosalinda-Tovar et al., 2000 [41] | |
| EHI_120900 | Calcium-binding protein 1 (EhCBP1) | Phagocytic cups and cytoskeleton dynamics | Nickel et al., 2000 [39] |
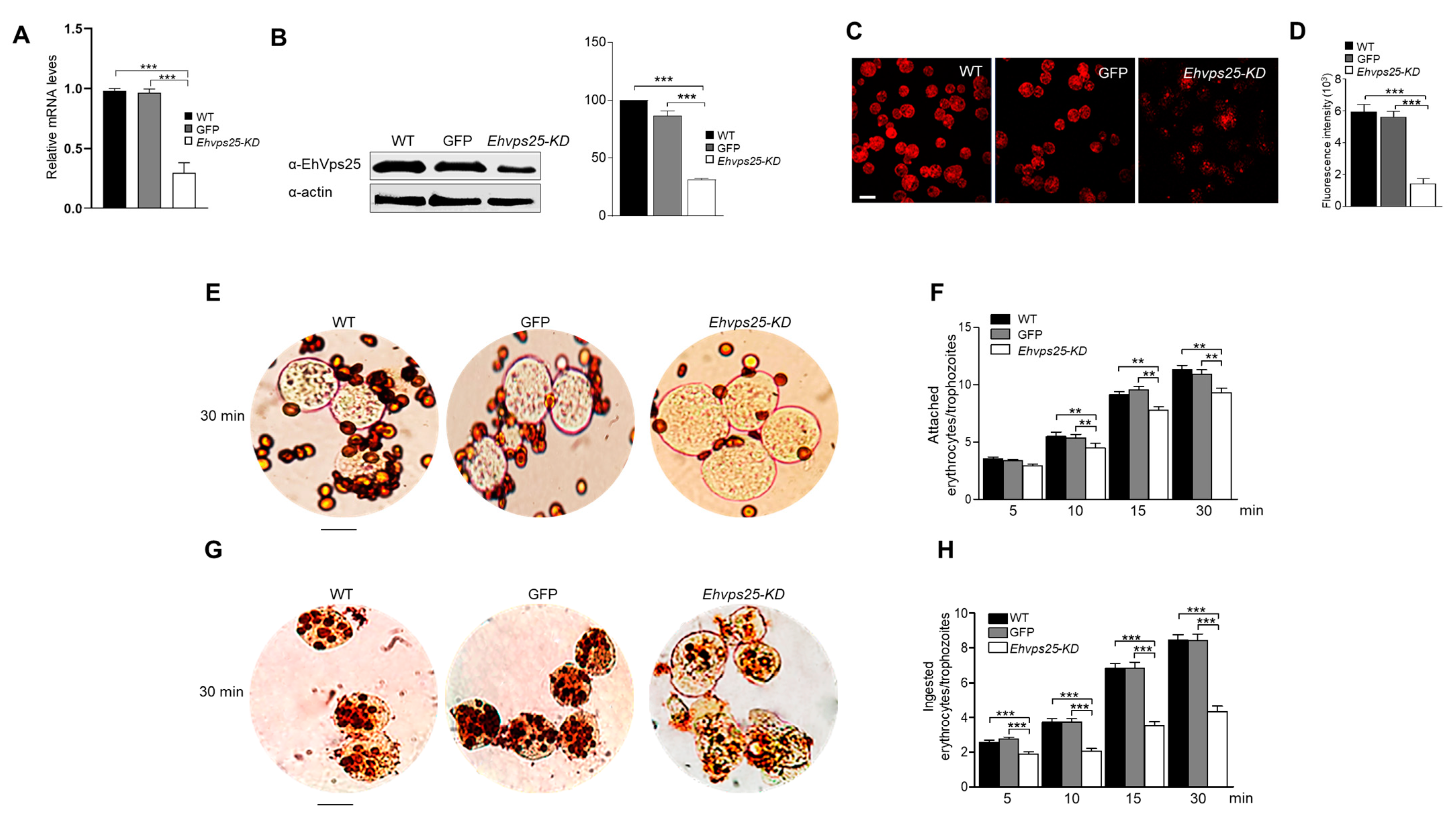
2.8. The ESCRT-II Complex Is Involved in Phagocytosis and Adherence
3. Discussion
4. Materials and Methods
4.1. ESCRT-II Searching and Phylogenetic Analysis
4.2. 3D Structure of EhVps22, EhVps25, EhVps36
4.3. Molecular Dynamics Simulation (MDS)
4.4. Trajectory Analysis
4.5. Protein–Protein Docking
4.6. Culture of Trophozoites
4.7. Cloning of the E. histolytica vps22, vps25, and vps36 Genes (Ehvps22, Ehvps25, and Ehvps36)
4.8. PCR and RT-qPCR Assays
4.9. Expression and Purification of Recombinant Proteins and Generation of Anti-EhVps22, EhVps25, and EhVps36 Antibodies
4.10. Western Blot Experiments
4.11. Phagocytosis Assays
4.12. Laser Confocal Microscopy Assays
4.13. Pull-Down Assays
4.14. Spectrometry Mass
4.15. Silencing Assay
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmidt, O.; Teis, D. The ESCRT machinery. Curr. Biol. 2012, 22, R116–R120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowers, K.; Lottridge, J.; Helliwell, S.B.; Goldthwaite, L.M.; Luzio, J.P.; Stevens, T.H. Protein–Protein Interactions of ESCRT Complexes in the Yeast Saccharomyces cerevisiae. Traffic 2004, 5, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Vietri, M.; Radulovic, M.; Stenmark, H. The many functions of ESCRTs. Nat. Rev. Mol. Cell Biol. 2020, 21, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Henne, W.M.; Buchkovich, N.J.; Emr, S.D. The ESCRT Pathway. Dev. Cell 2011, 21, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Katzmann, D.J.; Babst, M.; Emr, S.D. Ubiquitin-Dependent Sorting into the Multivesicular Body Pathway Requires the Function of a Conserved Endosomal Protein Sorting Complex, ESCRT-I. Cell 2001, 106, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Babst, M.; Katzmann, D.J.; Estepa-Sabal, E.J.; Meerloo, T.; Emr, S.D. Escrt-III: An endosome-associated heterooligomeric protein complex required for mvb sorting. Dev. Cell 2002, 3, 271–282. [Google Scholar] [CrossRef] [Green Version]
- Babst, M.; Sato, T.K.; Banta, L.M.; Emr, S.D. Endosomal transport function in yeast requires a novel AAA-type ATPase, Vps4p. EMBO J. 1997, 16, 1820–1831. [Google Scholar] [CrossRef]
- Wunder, C.; Wollert, T.; Hurley, J.H.; Lippincott-Schwartz, J. Membrane scission by the ESCRT-III complex. Nature 2009, 458, 172–177. [Google Scholar] [CrossRef] [Green Version]
- Gill, D.J.; Teo, H.; Sun, J.; Perisic, O.; Veprintsev, D.B.; Emr, S.D.; Williams, R.L. Structural insight into the ESCRT-I/-II link and its role in MVB trafficking. EMBO J. 2007, 26, 600–612. [Google Scholar] [CrossRef] [Green Version]
- Teo, H.; Gill, D.J.; Sun, J.; Perisic, O.; Veprintsev, D.B.; Vallis, Y.; Emr, S.D.; Williams, R.L. ESCRT-I Core and ESCRT-II GLUE Domain Structures Reveal Role for GLUE in Linking to ESCRT-I and Membranes. Cell 2006, 125, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Teo, H.; Perisic, O.; González, B.; Williams, R.L. ESCRT-II, an Endosome-Associated Complex Required for Protein Sorting: Crystal Structure and Interactions with ESCRT-III and Membranes. Dev. Cell 2004, 7, 559–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odorizzi, G.; Katzmann, D.J.; Babst, M.; Audhya, A.; Emr, S.D. Bro1 is an endosome-associated protein that functions in the MVB pathway in Saccharomyces cerevisiae. J. Cell Sci. 2003, 116, 1893–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, H.; Chevallier, J.; Mayran, N.; Le Blanc, I.; Ferguson, C.; Fauré, J.; Blanc, N.S.; Matile, S.; Dubochet, J.; Sadoul, R.; et al. Role of LBPA and Alix in Multivesicular Liposome Formation and Endosome Organization. Science 2004, 303, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Saha, N.; Dutta, S.; Datta, S.P.; Sarkar, S. The minimal ESCRT machinery of Giardia lamblia has altered inter-subunit interactions within the ESCRT-II and ESCRT-III complexes. Eur. J. Cell Biol. 2018, 97, 44–62. [Google Scholar] [CrossRef] [PubMed]
- Ausencio, G.; Rosario, J.-R.; Guillermina, G.-R.; Cecilia, B.; Bibiana, C.-M.; Lizbeth, S.-V.; Esther, O. EhVps23, an ESCRT-I Member, Is a Key Factor in Secretion, Motility, Phagocytosis and Tissue Invasion by Entamoeba histolytica. Front. Cell. Infect. Microbiol. 2022, 12, 835654. [Google Scholar] [CrossRef]
- Avalos-Padilla, Y.; Betanzos, A.; Javier-Reyna, R.; García-Rivera, G.; Chávez-Munguía, B.; Lagunes-Guillén, A.; Ortega, J.; Orozco, E. EhVps32 Is a Vacuole-Associated Protein Involved in Pinocytosis and Phagocytosis of Entamoeaba histolytica. PLoS Pathog. 2015, 11, e1005079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Cuevas, N.A.; Weber, C.; Hon, C.-C.; Guillen, N. Gene expression profiling in Entamoeba histolytica identifies key components in iron uptake and metabolism. PLoS ONE 2014, 9, e107102. [Google Scholar] [CrossRef]
- López-Reyes, I.; García-Rivera, G.; Bañuelos, C.; Herranz, S.; Vincent, O.; López-Camarillo, C.; Marchat, L.A.; Orozco, E. Detection of the Endosomal Sorting Complex Required for Transport in Entamoeba histolytica and Characterization of the EhVps4 Protein. J. Biomed. Biotechnol. 2010, 2010, 890674. [Google Scholar] [CrossRef] [Green Version]
- Ren, X.; Hurley, J.H. VHS domains of ESCRT-0 cooperate in high-avidity binding to polyubiquitinated cargo. EMBO J. 2010, 29, 1045–1054. [Google Scholar] [CrossRef]
- Blanc, C.; Charette, S.J.; Mattei, S.; Aubry, L.; Smith, E.W.; Cosson, P.; Letourneur, F. Dictyostelium Tom1 Participates to an Ancestral ESCRT-0 Complex. Traffic 2009, 10, 161–171. [Google Scholar] [CrossRef]
- Ausencio, G.; Rosario, J.-R.; Guillermina, G.-R.; Cecilia, B.; Sarita, M.; Jaime, O.-L.; Bibiana, C.-M.; Lizbeth, S.-V.; Esther, O. EhVps23: A Component of ESCRT-I That Participates in Vesicular Trafficking and Phagocytosis of Entamoeba histolytica. Front. Cell. Infect. Microbiol. 2021, 11, 770759. [Google Scholar] [CrossRef]
- Yunuen, A.-P.; Yunuen, A.-P.; Roland, L.K.; Rosario, J.-R.; Guillermina, G.-R.; Reinhard, L.; Rumiana, D.; Esther, O. The Conserved ESCRT-III Machinery Participates in the Phagocytosis of Entamoeba histolytica. Front. Cell. Infect. Microbiol. 2018, 8, 53. [Google Scholar] [CrossRef]
- Arroyo, R.; Orozco, E. Localization and identification of an Entamoeba histolytica adhesin. Mol. Biochem. Parasitol. 1987, 23, 151–158. [Google Scholar] [CrossRef]
- Bañuelos, C.; García-Rivera, G.; López-Reyes, I.; Orozco, E. Functional characterization of EhADH112: An Entamoeba histolytica Bro1 domain-containing protein. Exp. Parasitol. 2005, 110, 292–297. [Google Scholar] [CrossRef]
- Williams, R.L.; Urbé, S. The emerging shape of the ESCRT machinery. Nat. Rev. Mol. Cell Biol. 2007, 8, 355–368. [Google Scholar] [CrossRef]
- Karimi, Z.; Falsafi-Zadeh, S.; Galehdari, H.; Jalali, A. Homology modeling and molecular dynamics simulation of odonthubuthus doriae (Od1) scorpion toxin in comparison to the BmK M1. Bioinformation 2012, 8, 474–478. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.; Akram, Z.; Iqbal, H.M.N.; Munn, A.L. The regulation of Endosomal Sorting Complex Required for Transport and accessory proteins in multivesicular body sorting and enveloped viral budding—An overview. Int. J. Biol. Macromol. 2019, 127, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cecilia, B.; Abigail, B.; Abigail, B.; Rosario, J.-R.; Ausencio, G.; Esther, O. Molecular interplays of the Entamoeba histolytica endosomal sorting complexes required for transport during phagocytosis. Front. Cell. Infect. Microbiol. 2022, 12, 855797. [Google Scholar] [CrossRef]
- Guiney, E.L.; Zhu, L.; Sardana, R.; Emr, S.D.; Baile, M.G. Methods for studying the regulation of membrane traffic by ubiquitin and the ESCRT pathway. Methods Enzymol. 2019, 619, 269–291. [Google Scholar] [CrossRef]
- Baños-Mateos, S.; Rojas, A.L.; Hierro, A. VPS29, a tweak tool of endosomal recycling. Curr. Opin. Cell Biol. 2019, 59, 81–87. [Google Scholar] [CrossRef]
- Nakada-Tsukui, K.; Saito-Nakano, Y.; Ali, V.; Nozaki, T. A Retromerlike Complex Is a Novel Rab7 Effector That Is Involved in the Transport of the Virulence Factor Cysteine Protease in the Enteric Protozoan Parasite Entamoeba histolytica. Mol. Biol. Cell 2005, 16, 5294–5303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, V.K.; Yadav, R.; Watanabe, N.; Tomar, P.; Mukherjee, M.; Gourinath, S.; Nakada-Tsukui, K.; Nozaki, T.; Datta, S. Structural and thermodynamic characterization of metal binding in Vps29 from Entamoeba histolytica: Implication in retromer function. Mol. Microbiol. 2017, 106, 562–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzmán-Medrano, R.; Castillo-Juárez, B.A.; García-Pérez, R.M.; Salas-Casas, A.; Orozco, E.; Rodríguez, M.A. Entamoeba histolytica: Alterations in EhRabB protein in a phagocytosis deficient mutant correlate with the Entamoeba dispar RabB sequence. Exp. Parasitol. 2005, 110, 259–264. [Google Scholar] [CrossRef]
- Javier-Reyna, R.; Montaño, S.; García-Rivera, G.; Rodríguez, M.A.; González-Robles, A.; Orozco, E. EhRabB mobilises the EhCPADH complex through the actin cytoskeleton during phagocytosis of Entamoeba histolytica. Cell. Microbiol. 2019, 21, e13071. [Google Scholar] [CrossRef]
- Verma, K.; Srivastava, V.K.; Datta, S. Rab GTPases take centre stage in understanding Entamoeba histolytica biology. Small GTPases 2020, 11, 320–333. [Google Scholar] [CrossRef]
- Gilchrist, C.A.; Petri, W.A. Virulence factors of Entamoeba histolytica. Curr. Opin. Microbiol. 1999, 2, 433–437. [Google Scholar] [CrossRef]
- Saucedo-Mendiola, M.L.; Salas-Pacheco, J.M.; Nájera, H.; Rojo-Domínguez, A.; Yépez-Mulia, L.; Avitia-Domínguez, C.; Téllez-Valencia, A. Discovery of Entamoeba histolytica hexokinase 1 inhibitors through homology modeling and virtual screening. J. Enzym. Inhib. Med. Chem. 2014, 29, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Babuta, M.; Mansuri, M.S.; Bhattacharya, S.; Bhattacharya, A. The Entamoeba histolytica, Arp2/3 Complex Is Recruited to Phagocytic Cups through an Atypical Kinase EhAK1. PLoS Pathog. 2015, 11, e1005310. [Google Scholar] [CrossRef]
- Nickel, R.; Jacobs, T.; Urban, B.; Scholze, H.; Bruhn, H.; Leippe, M. Two novel calcium-binding proteins from cytoplasmic granules of the protozoan parasite Entamoeba histolytica. FEBS Lett. 2000, 486, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Rout, A.K.; Padhan, N.; Barnwal, R.P.; Bhattacharya, A.; Chary, K.V.R. Calmodulin-like Protein from Entamoeba histolytica: Solution Structure and Calcium-Binding Properties of a Partially Folded Protein. Biochemistry 2011, 50, 181–193. [Google Scholar] [CrossRef]
- Tovar, R.; Murguía-López, M.L.; de Lourdes Muñoz, M.a. Immunolocalization of Clathrin During Electron-Dense Granule Secretion in Entamoeba histolytica. Arch. Med. Res. 2000, 31, S143–S144. [Google Scholar] [CrossRef] [PubMed]
- Solis, C.F.; Guillén, N. Silencing Genes by RNA Interference in the Protozoan Parasite Entamoeba histolytica. Methods Mol. Biol. 2008, 442, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Vayssié, L.; Vargas, M.; Weber, C.; Guillén, N. Double-stranded RNA mediates homology-dependant gene silencing of γ-tubulin in the human parasite Entamoeba histolytica. Mol. Biochem. Parasitol. 2004, 138, 21–28. [Google Scholar] [CrossRef]
- Christ, L.; Raiborg, C.; Wenzel, E.M.; Campsteijn, C.; Stenmark, H. Cellular Functions and Molecular Mechanisms of the ESCRT Membrane-Scission Machinery. Trends Biochem. Sci. 2017, 42, 42–56. [Google Scholar] [CrossRef]
- García-Rivera, G.; Rodríguez, M.A.; Ocádiz, R.; Martínez-López, M.C.; Arroyo, R.; González-Robles, A.; Orozco, E. Entamoeba histolytica: A novel cysteine protease and an adhesin form the 112 kDa surface protein. Mol. Microbiol. 1999, 33, 556–568. [Google Scholar] [CrossRef]
- Lozano, R.P.; Naghavi, M.P.; Lim, S.P.; Aboyans, V.P.; Abraham, J.M.P.H.; Adair, T.P.; Ahn, S.Y.M.P.H.; AlMazroa, M.A.M.D.; Anderson, H.R.P.; Anderson, L.M.P.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Curtiss, M.; Jones, C.; Babst, M. Efficient Cargo Sorting by ESCRT-I and the Subsequent Release of ESCRT-I from Multivesicular Bodies Requires the Subunit Mvb12. Mol. Biol. Cell 2007, 18, 636–645. [Google Scholar] [CrossRef]
- Slagsvold, T.; Aasland, R.; Hirano, S.; Bache, K.G.; Raiborg, C.; Trambaiolo, D.; Wakatsuki, S.; Stenmark, H. Eap45 in Mammalian ESCRT-II Binds Ubiquitin via a Phosphoinositide-interacting GLUE Domain. J. Biol. Chem. 2005, 280, 19600–19606. [Google Scholar] [CrossRef] [Green Version]
- Dimova, R. Giant Vesicles and Their Use in Assays for Assessing Membrane Phase State, Curvature, Mechanics, and Electrical Properties. Annu. Rev. Biophys. 2019, 48, 93–119. [Google Scholar] [CrossRef] [Green Version]
- Qiurong, X.; Qiurong, X.; Ahai, C.; Ahai, C.; Yunzhi, Z.; Yunzhi, Z.; Mingyue, Y.; Mingyue, Y.; Wei, X.; Wei, X.; et al. Component Interaction of ESCRT Complexes Is Essential for Endocytosis-Dependent Growth, Reproduction, DON Production and Full Virulence in Fusarium graminearum. Front. Microbiol. 2019, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Anand, G.; Sharma, S.; Parimita Rath, P.; Gourinath, S.; Bhattacharya, A. EhP3, a homolog of 14-3-3 family of protein participates in actin reorganization and phagocytosis in Entamoeba histolytica. PLoS Pathog. 2019, 15, e1007789. [Google Scholar] [CrossRef]
- Bharadwaj, R.; Sharma, S.; Arya, R.; Bhattacharya, S.; Bhattacharya, A. EhRho1 regulates phagocytosis by modulating actin dynamics through EhFormin1 and EhProfilin1 in Entamoeba histolytica. Cell. Microbiol. 2018, 20, e12851. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Somlata; Bhattacharya, S. A C2 domain protein kinase initiates phagocytosis in the protozoan parasite Entamoeba histolytica. Nat. Commun. 2011, 2, 230. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Hernández, M.; Javier-Reyna, R.; Sotto-Ortega, I.; García-Rivera, G.; Montaño, S.; Betanzos, A.; Zanatta, D.; Orozco, E. Protein Sumoylation Is Crucial for Phagocytosis in Entamoeba histolytica Trophozoites. Int. J. Mol. Sci. 2021, 22, 5709. [Google Scholar] [CrossRef]
- Kumari, R.; Gupta, P.; Tiwari, S. Ubc7/Ube2g2 ortholog in Entamoeba histolytica: Connection with the plasma membrane and phagocytosis. Parasitol. Res. 2018, 117, 1599–1611. [Google Scholar] [CrossRef]
- Mansuri, M.S.; Bhattacharya, S.; Bhattacharya, A. A novel alpha kinase EhAK1 phosphorylates actin and regulates phagocytosis in Entamoeba histolytica. PLoS Pathog. 2014, 10, e1004411. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Morgado, P.; Zhang, H.; Ehrenkaufer, G.; Manna, D.; Singh, U. Characterization of Extracellular Vesicles from Entamoeba histolytica Identifies Roles in Intercellular Communication That Regulates Parasite Growth and Development. Infect. Immun. 2020, 88, e00349-20. [Google Scholar] [CrossRef]
- Zhou, H.; Guan, Y.; Feng, M.; Fu, Y.; Tachibana, H.; Cheng, X. Evaluation on Elongation Factor 1 Alpha of Entamoeba histolytica Interaction with the Intermediate Subunit of the Gal/GalNAc Lectin and Actin in Phagocytosis. Pathogens 2020, 9, 702. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Amos, B.; Aurrecoechea, C.; Barba, M.; Barreto, A.; Basenko, E.Y.; Bażant, W.; Belnap, R.; Blevins, A.S.; Böhme, U.; Brestelli, J.; et al. VEuPathDB: The eukaryotic pathogen, vector and host bioinformatics resource center. Nucleic Acids Res. 2022, 50, D898–D911. [Google Scholar] [CrossRef]
- Wei, Z.; Chengxin, Z.; Yang, L.; Robin, P.; Eric, W.B.; Yang, Z. Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep. Methods 2021, 1, 100014. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y. Protein Structure and Function Prediction Using I-TASSER. Curr. Protoc. Bioinform. 2015, 52, 5.8.1–5.8.15. [Google Scholar] [CrossRef] [Green Version]
- Hurley, J.H. ESCRT complexes and the biogenesis of multivesicular bodies. Curr. Opin. Cell Biol. 2007, 20, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Huber, S.T.; Mostafavi, S.; Mortensen, S.A.; Sachse, C. Structure and assembly of ESCRT-III helical Vps24 filaments. Sci. Adv. 2020, 6, eaba4897. [Google Scholar] [CrossRef]
- Chen, V.B.; Arendall, W.B., III; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Lee, J.; Cheng, X.; Swails, J.M.; Yeom, M.S.; Eastman, P.K.; Lemkul, J.A.; Wei, S.; Buckner, J.; Jeong, J.C.; Qi, Y.; et al. CHARMM-GUI Input Generator for NAMD, GROMACS, AMBER, OpenMM, and CHARMM/OpenMM Simulations Using the CHARMM36 Additive Force Field. J. Chem. Theory Comput. 2016, 12, 405–413. [Google Scholar] [CrossRef]
- Huang, J.; MacKerell, A.D., Jr. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef] [Green Version]
- Feller, S.E.; Zhang, Y.; Pastor, R.W.; Brooks, B.R. Constant pressure molecular dynamics simulation: The Langevin piston method. J. Chem. Phys. 1995, 103, 4613–4621. [Google Scholar] [CrossRef]
- Martyna, G.J.; Klein, M.L.; Tuckerman, M. Nosé–Hoover chains: The canonical ensemble via continuous dynamics. J. Chem. Phys. 1992, 97, 2635–2643. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Hénin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef]
- Koukos, P.I.; Glykos, N.M. Grcarma: A fully automated task-oriented interface for the analysis of molecular dynamics trajectories. J. Comput. Chem. 2013, 34, 2310–2312. [Google Scholar] [CrossRef]
- Comeau, S.R.; Gatchell, D.W.; Vajda, S.; Camacho, C.J. ClusPro: An automated docking and discrimination method for the prediction of protein complexes. Bioinformatics 2004, 20, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Kozakov, D.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Vajda, S. How good is automated protein docking? Proteins Struct. Funct. Bioinform. 2013, 81, 2159–2166. [Google Scholar] [CrossRef] [Green Version]
- Mashiach, E.; Schneidman-Duhovny, D.; Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: A web server for fast interaction refinement in molecular docking. Nucleic Acids Res. 2008, 36, W229–W232. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Hutchinson, E.G.; Michie, A.D.; Wallace, A.C.; Jones, M.L.; Thornton, J.M. PDBsum: A web-based database of summaries and analyses of all PDB structures. Trends Biochem. Sci. 1997, 22, 488–490. [Google Scholar] [CrossRef]
- Novikoff, P.M.; Novikoff, A.B. Peroxisomes in Absorptive Cells of Mammalian Small Intestine. J. Cell Biol. 1972, 53, 532–560. [Google Scholar] [CrossRef] [Green Version]
- Diamond, L.S.; Harlow, D.R.; Cunnick, C.C. A new medium for the axenic cultivation of Entamoeba histolytica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 431–432. [Google Scholar] [CrossRef]
- Orozco, E.; Guarneros, G.; Martinez-Palomo, A.; Sanchez, T. Entamoeba histolytica. Phagocytosis as a virulence factor. J. Exp. Med. 1983, 158, 1511–1521. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Hernández, M.; Javier-Reyna, R.; Martínez-Valencia, D.; Montaño, S.; Orozco, E. Dynamic Association of ESCRT-II Proteins with ESCRT-I and ESCRT-III Complexes during Phagocytosis of Entamoeba histolytica. Int. J. Mol. Sci. 2023, 24, 5267. https://doi.org/10.3390/ijms24065267
Díaz-Hernández M, Javier-Reyna R, Martínez-Valencia D, Montaño S, Orozco E. Dynamic Association of ESCRT-II Proteins with ESCRT-I and ESCRT-III Complexes during Phagocytosis of Entamoeba histolytica. International Journal of Molecular Sciences. 2023; 24(6):5267. https://doi.org/10.3390/ijms24065267
Chicago/Turabian StyleDíaz-Hernández, Mitzi, Rosario Javier-Reyna, Diana Martínez-Valencia, Sarita Montaño, and Esther Orozco. 2023. "Dynamic Association of ESCRT-II Proteins with ESCRT-I and ESCRT-III Complexes during Phagocytosis of Entamoeba histolytica" International Journal of Molecular Sciences 24, no. 6: 5267. https://doi.org/10.3390/ijms24065267