
Genome-Wide Characterization of B-Box Gene Family in Salvia miltiorrhiza
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of SmBBXs Genes
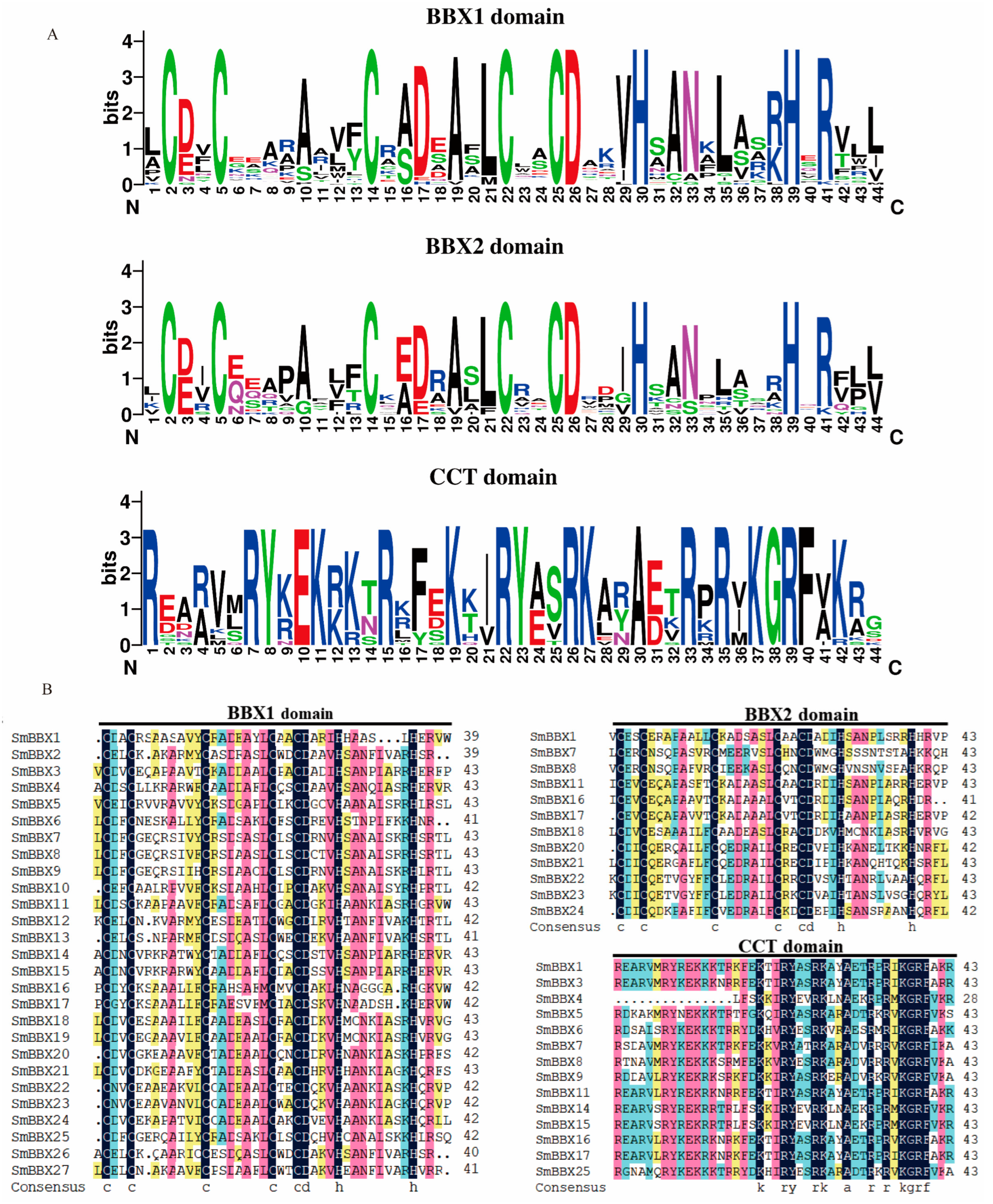
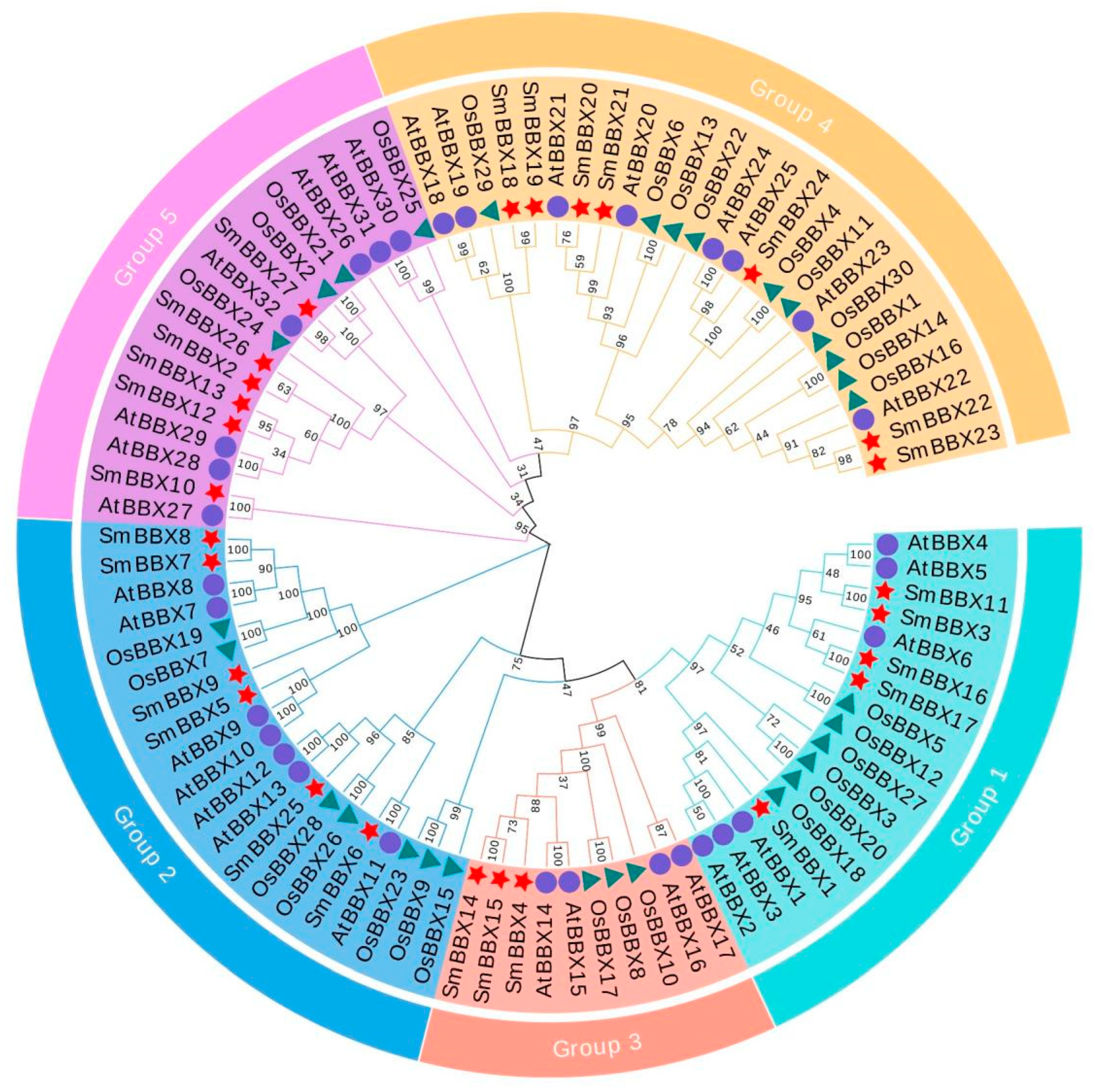
2.2. Protein Sequences and Phylogenetic Analyses of SmBBXs
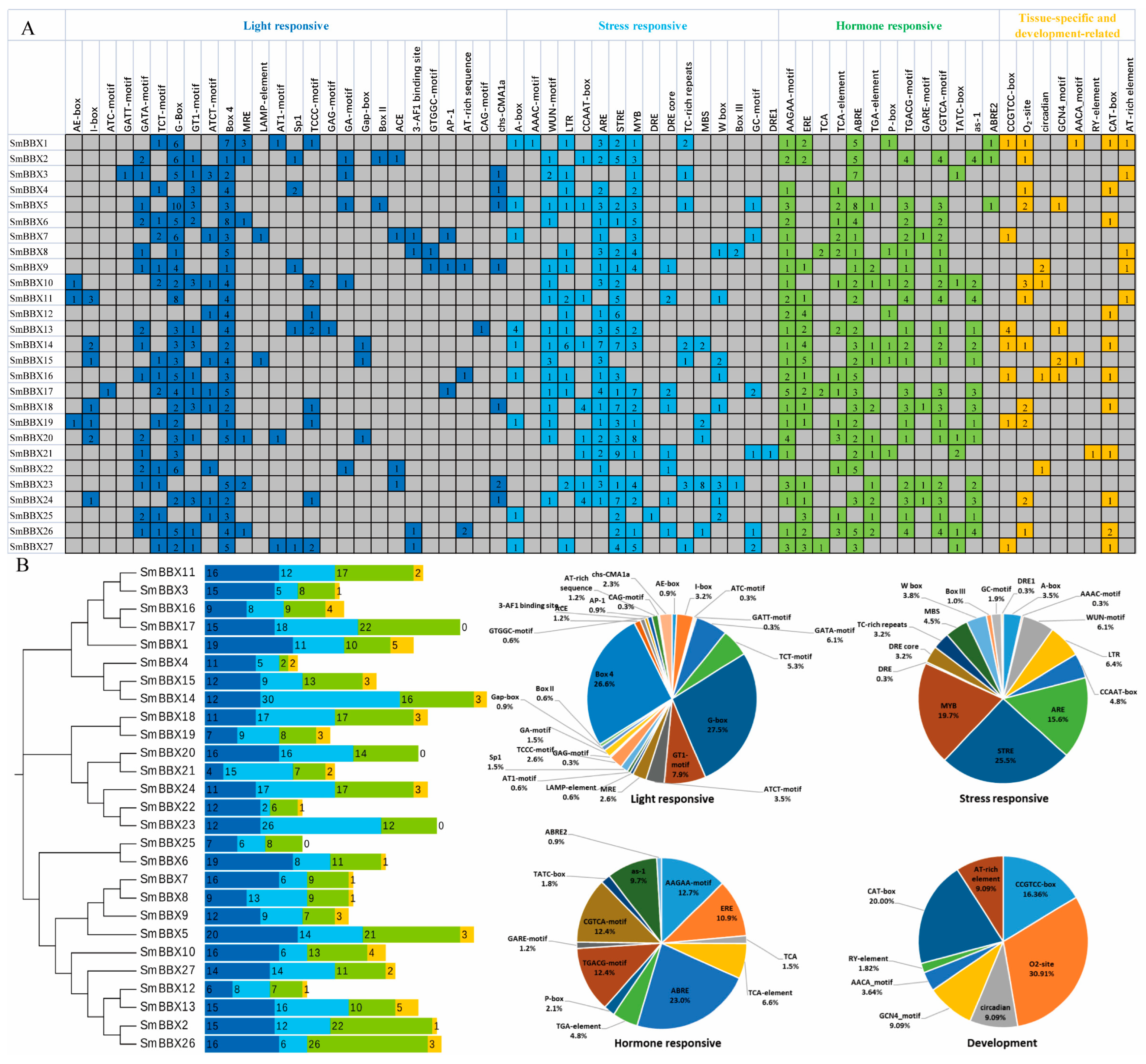
2.3. Analysis of cis-Elements in SmBBXs Promoter Region
2.4. Calculation of Ka/Ks Values for Homologous SmBBXs Gene Pairs
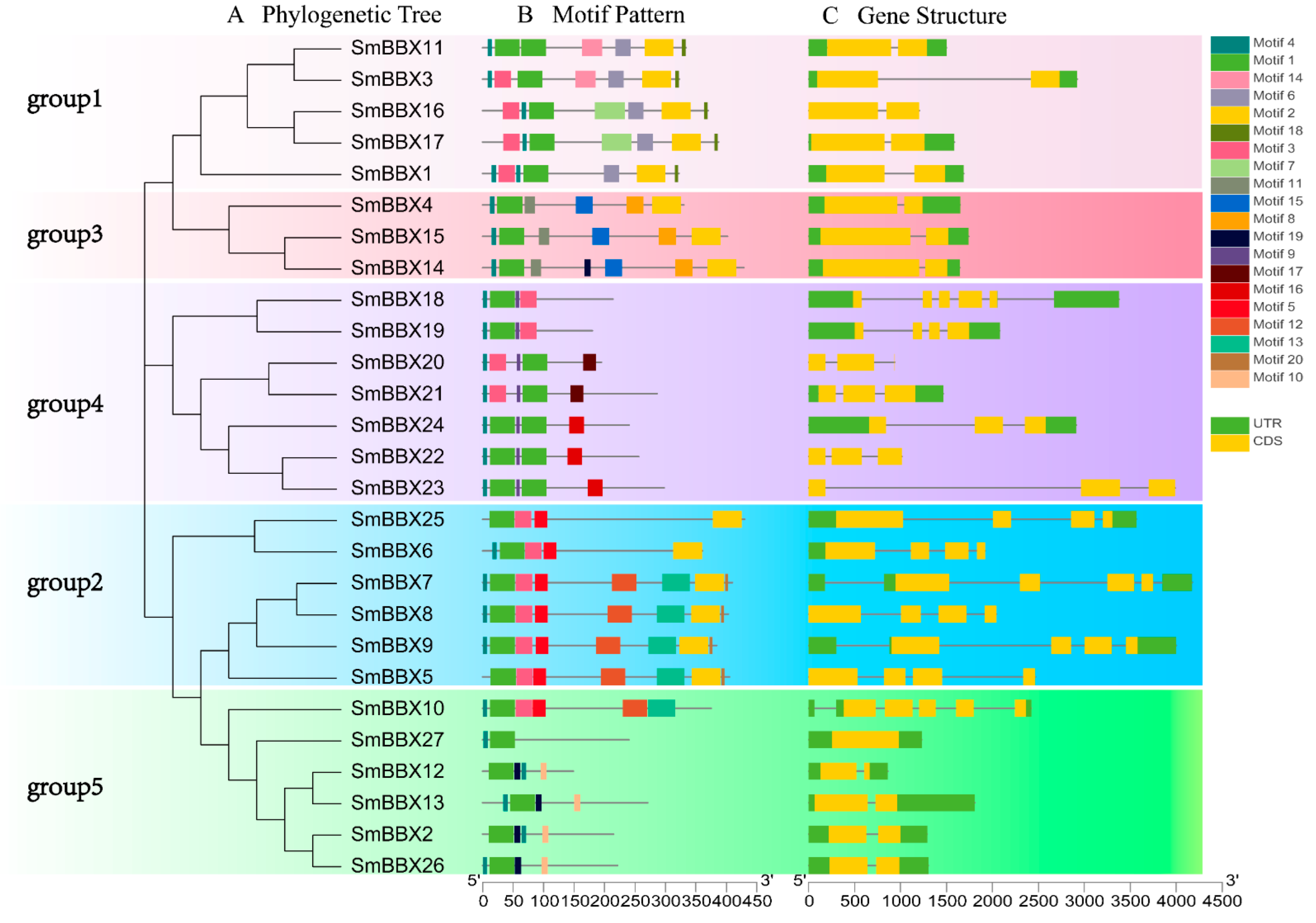
2.5. Gene Structure, Motifs, and Sequence Analysis of SmBBXs
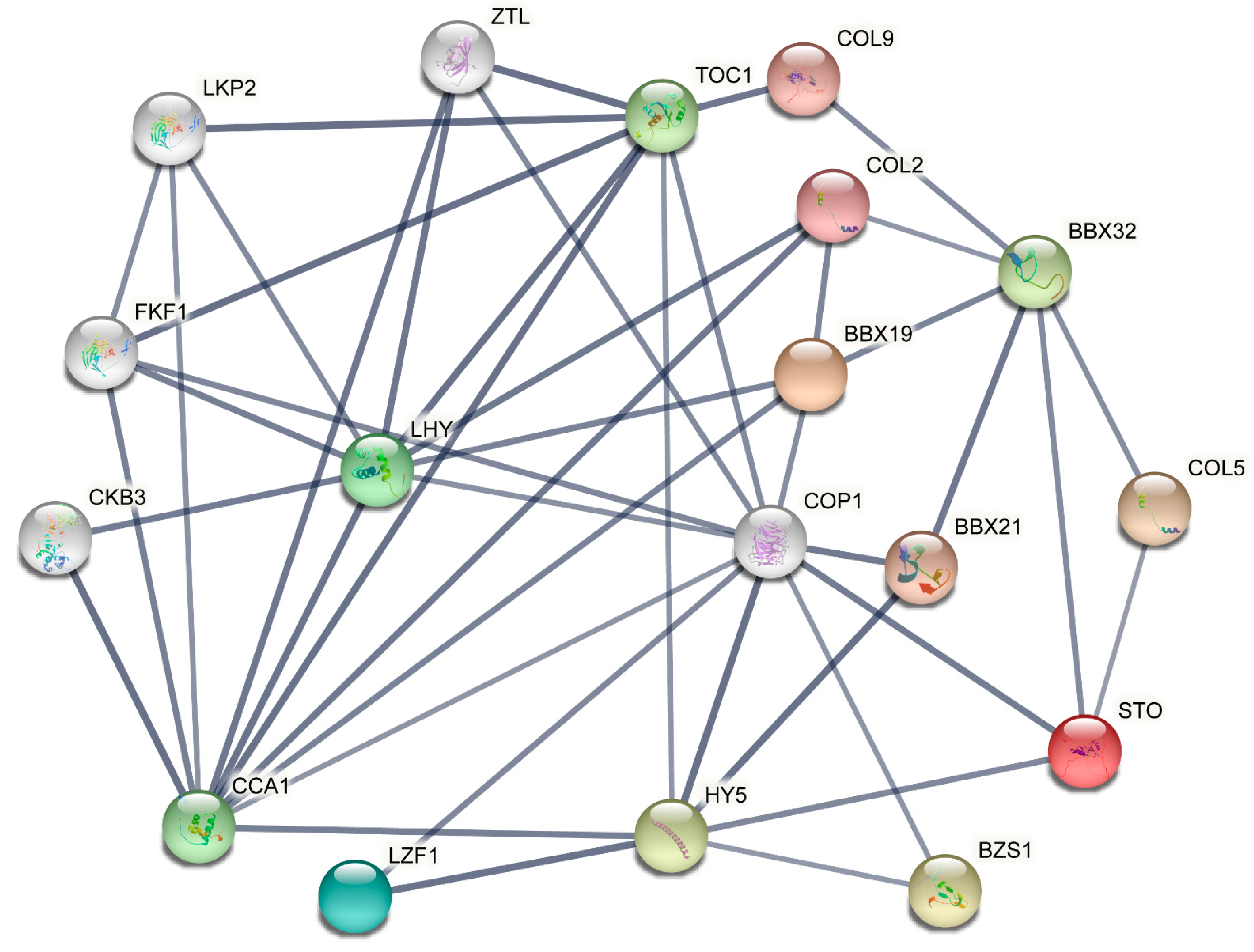
2.6. Analysis of Protein–Protein Interactions in SmBBXs
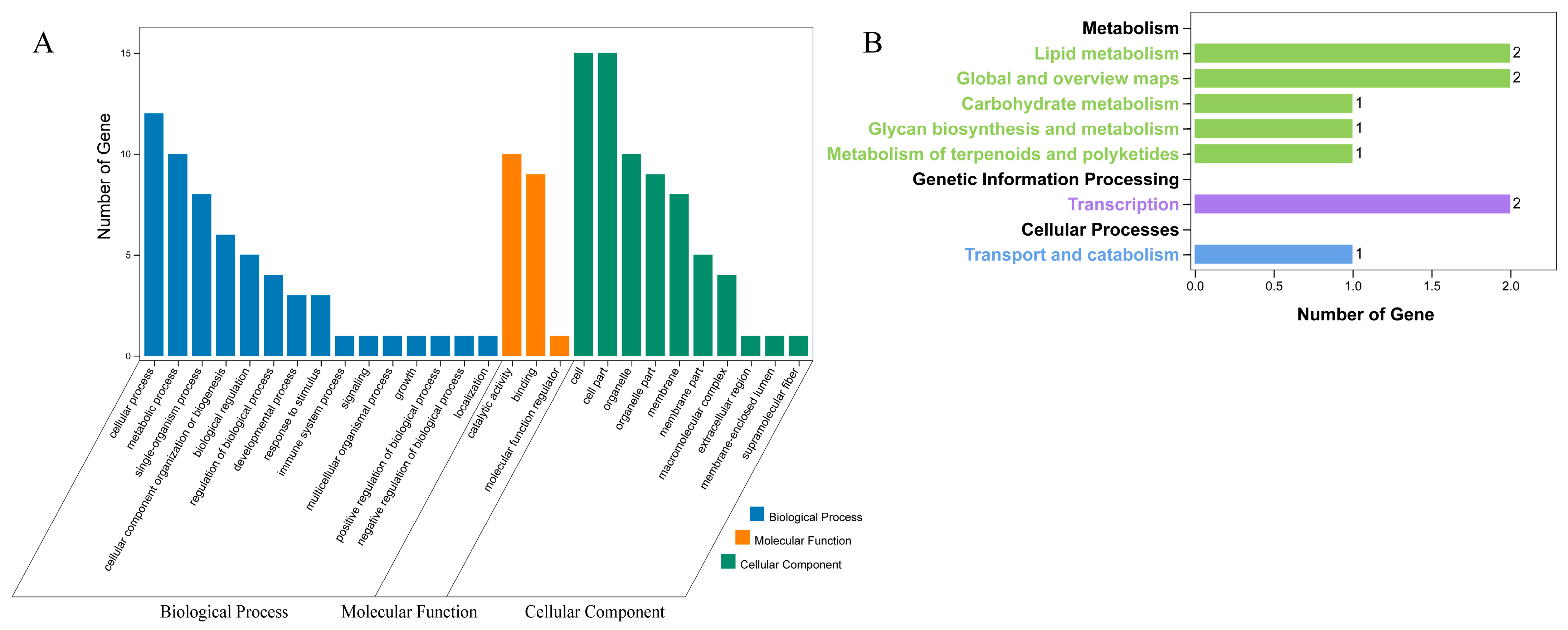
2.7. Analysis of GO and KEGG in SmBBXs
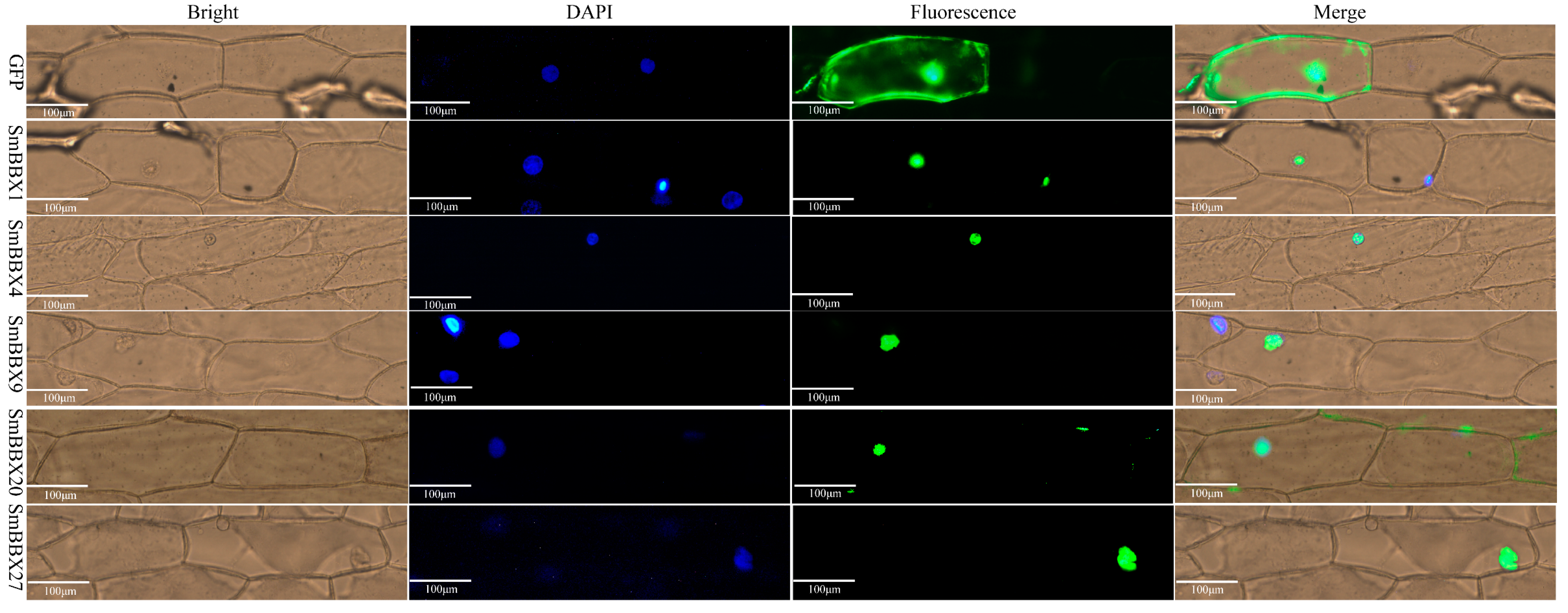
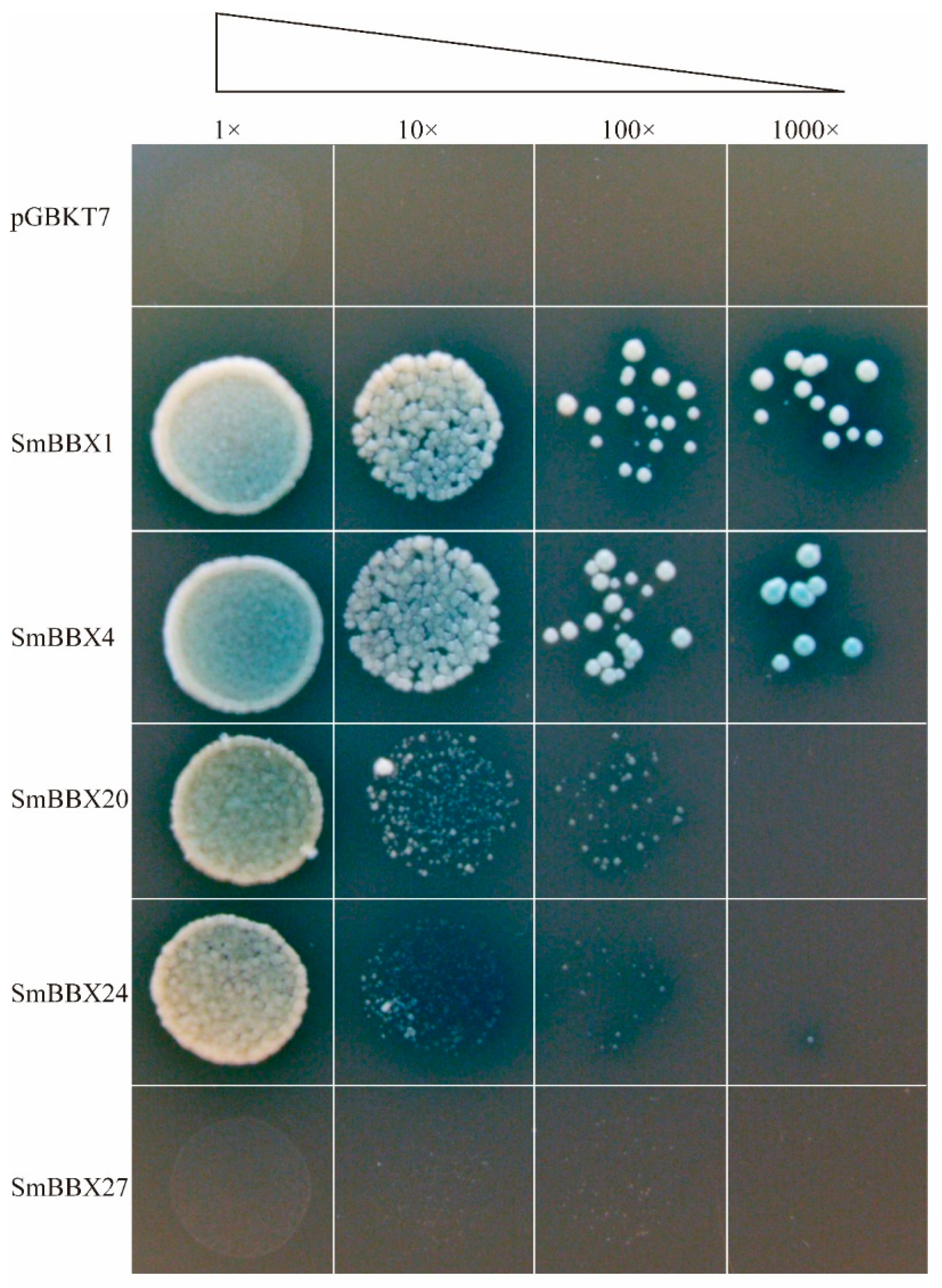
2.8. Subcellular Localization and Transcription Activation of SmBBX Genes
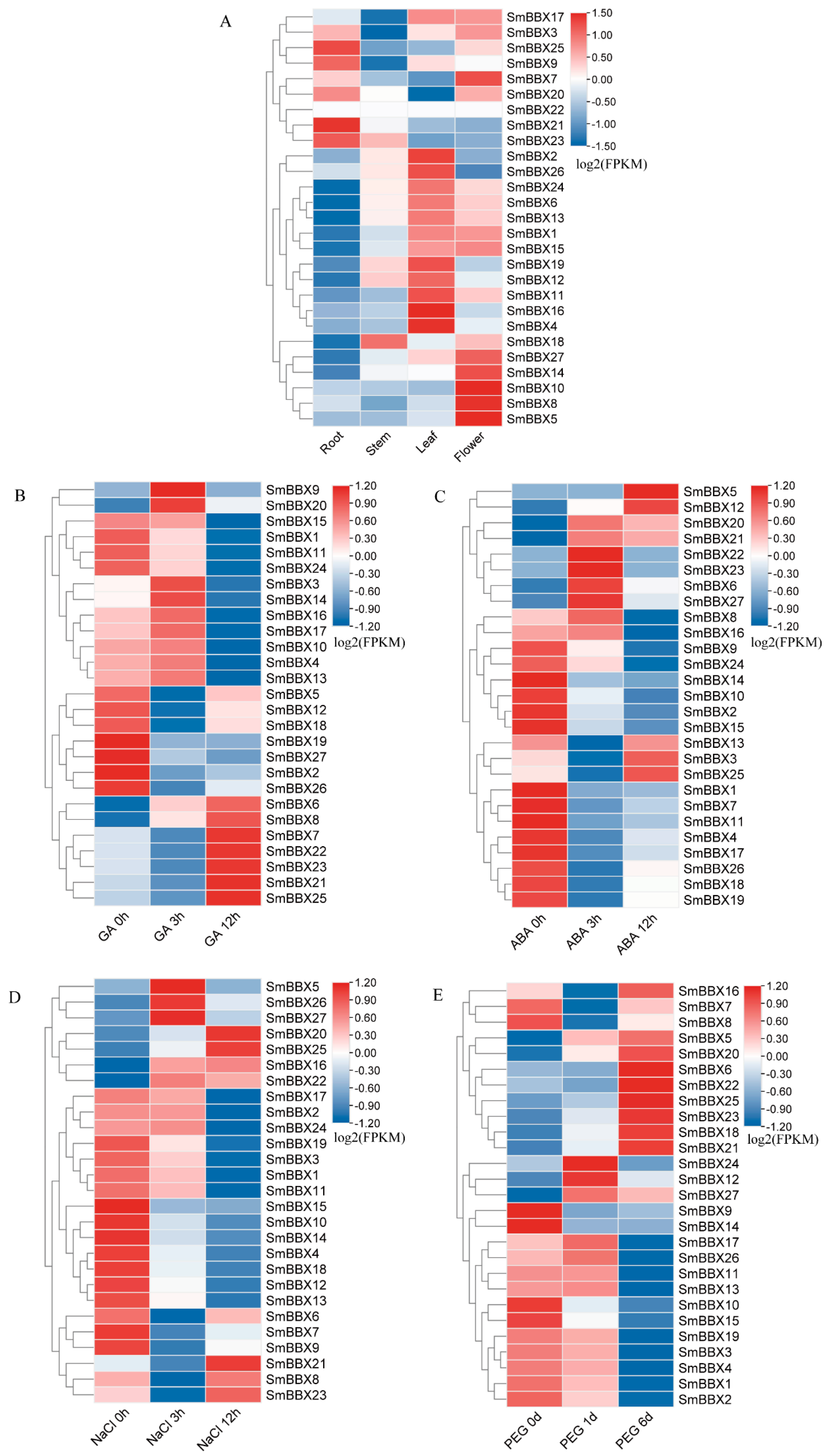
2.9. Expression Profiles of SmBBXs Genes
3. Discussion
4. Materials and Methods
4.1. Genome-Wide Identification of BBX Genes in S. miltiorrhiza
4.2. Sequence Alignment and Phylogenetic Analysis
4.3. Protein Properties, Exon and Intron Distribution, and Conserved Motif Analysis
4.4. Cis-Element Analysis for BBX Gene Promoters
4.5. Calculation of Ka/Ks Values for Homologous SmBBXs Gene Pairs
4.6. Gene Ontology (GO) and KEGG Annotation
4.7. Protein Interaction Network Analysis
4.8. Subcellular Localization of BBX Proteins
4.9. Transactivation Activity Assay of SmBBXs Genes
4.10. Expression Analysis of SmBBXs Genes Based on Transcriptome Sequencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khanna, R.; Kronmiller, B.; Maszle, D.R.; Coupland, G.; Holm, M.; Mizuno, T.; Wu, S.H. The Arabidopsis B-box zinc finger family. Plant Cell 2009, 21, 3416–3420. [Google Scholar] [CrossRef] [Green Version]
- Lyu, G.; Li, D.; Li, S. Bioinformatics analysis of BBX family genes and its response to UV-B in Arabidopsis thaliana. Plant Signal. Behav. 2020, 15, 1782647. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, X.; Weng, X.; Wang, L.; Xie, W. The rice B-box zinc finger gene family: Genomic identification, characterization, expression profiling and diurnal analysis. PLoS ONE 2012, 7, e48242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, R.; Chen, J.; Huang, B.; Huang, Z.; Zhang, Z. The BBX gene family in Moso bamboo (Phyllostachys edulis): Identification, characterization and expression profiles. BMC Genom. 2021, 22, 533. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, H.; Huang, J.; Shi, T.; Meng, Z.; Chen, Q.; Deng, J. Genome-wide analysis of BBX gene family in Tartary buckwheat (Fagopyrum tataricum). PeerJ 2021, 9, e11939. [Google Scholar] [CrossRef]
- Zheng, L.W.; Ma, S.J.; Zhou, T.; Yue, C.P.; Hua, Y.P.; Huang, J.Y. Genome-wide identification of Brassicaceae B-BOX genes and molecular characterization of their transcriptional responses to various nutrient stresses in allotetraploid rapeseed. BMC Plant Biol. 2021, 21, 288. [Google Scholar] [CrossRef]
- Bu, X.; Wang, X.; Yan, J.; Zhang, Y.; Zhou, S.; Sun, X.; Yang, Y.; Ahammed, G.J.; Liu, Y.; Qi, M.; et al. Genome-Wide Characterization of B-Box Gene Family and Its Roles in Responses to Light Quality and Cold Stress in Tomato. Front. Plant Sci. 2021, 12, 698525. [Google Scholar] [CrossRef]
- Gangappa, S.N.; Botto, J.F. The BBX family of plant transcription factors. Trends Plant Sci. 2014, 19, 460–470. [Google Scholar] [CrossRef]
- Cao, J.; Yuan, J.; Zhang, Y.; Chen, C.; Zhang, B.; Shi, X.; Niu, R. Multi-layered roles of BBX proteins in plant growth and development. Stress Biol. 2023, 3, 1. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, H.J.; Qu, D.; Zhu, Z.Z.; Yang, Y.Z.; Zhao, Z.Y. The MdBBX22-miR858-MdMYB9/11/12 module regulates proanthocyanidin biosynthesis in apple peel. Plant Biotechnol. J. 2022, 20, 1683–1700. [Google Scholar] [CrossRef]
- An, J.P.; Wang, X.F.; Espley, R.V.; Lin-Wang, K.; Bi, S.Q.; You, C.X.; Hao, Y.J. An Apple B-Box Protein MdBBX37 Modulates Anthocyanin Biosynthesis and Hypocotyl Elongation Synergistically with MdMYBs and MdHY5. Plant Cell Physiol. 2020, 61, 130–143. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Zhang, C.L.; Li, H.L.; Wang, G.L.; You, C.X. Apple SINA E3 ligase MdSINA3 negatively mediates JA-triggered leaf senescence by ubiquitinating and degrading the MdBBX37 protein. Plant J. Cell Mol. Biol. 2022, 111, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Luo, D.; Xiong, C.; Lin, A.; Zhang, C.; Sun, W.; Zhang, J.; Yang, C.; Lu, Y.; Li, H.; Ye, Z.; et al. SlBBX20 interacts with the COP9 signalosome subunit SlCSN5-2 to regulate anthocyanin biosynthesis by activating SlDFR expression in tomato. Hortic. Res. 2021, 8, 163. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Zhang, C.; Dong, H.; Liu, X.; Guo, H.; Tong, B.; Fang, F.; Zhao, Y.; Yu, Y.; Liu, Y.; et al. Activation and negative feedback regulation of SlHY5 transcription by the SlBBX20/21-SlHY5 transcription factor module in UV-B signaling. Plant Cell 2022, 34, 2038–2055. [Google Scholar] [CrossRef]
- Lin, F.; Jiang, Y.; Li, J.; Yan, T.; Fan, L.; Liang, J.; Chen, Z.J.; Xu, D.; Deng, X.W. B-BOX DOMAIN PROTEIN28 Negatively Regulates Photomorphogenesis by Repressing the Activity of Transcription Factor HY5 and Undergoes COP1-Mediated Degradation. Plant Cell 2018, 30, 2006–2019. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Liang, Y.; Yan, T.; Wang, X.; Zhou, H.; Chen, C.; Zhang, Y.; Zhang, B.; Zhang, S.; Liao, J.; et al. The photomorphogenic repressors BBX28 and BBX29 integrate light and brassinosteroid signaling to inhibit seedling development in Arabidopsis. Plant Cell 2022, 34, 2266–2285. [Google Scholar] [CrossRef]
- Veciana, N.; Martín, G.; Leivar, P.; Monte, E. BBX16 mediates the repression of seedling photomorphogenesis downstream of the GUN1/GLK1 module during retrograde signalling. New Phytol. 2022, 234, 93–106. [Google Scholar] [CrossRef]
- Yuan, L.; Yu, Y.; Liu, M.; Song, Y.; Li, H.; Sun, J.; Wang, Q.; Xie, Q.; Wang, L.; Xu, X. BBX19 fine-tunes the circadian rhythm by interacting with PSEUDO-RESPONSE REGULATOR proteins to facilitate their repressive effect on morning-phased clock genes. Plant Cell 2021, 33, 2602–2617. [Google Scholar] [CrossRef]
- Li, Y.; Shi, Y.; Li, M.; Fu, D.; Wu, S.; Li, J.; Gong, Z.; Liu, H.; Yang, S. The CRY2-COP1-HY5-BBX7/8 module regulates blue light-dependent cold acclimation in Arabidopsis. Plant Cell 2021, 33, 3555–3573. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, H.; Ping, Q.; Zhang, Z.; Guan, Z.; Fang, W.; Chen, S.; Chen, F.; Jiang, J.; Zhang, F. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2019, 38, 15–24. [Google Scholar] [CrossRef]
- Xia, P.; Hu, W.; Liang, T.; Yang, D.; Liang, Z. An attempt to establish an Agrobacterium-mediated transient expression system in medicinal plants. Protoplasma 2020, 257, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Song, J.; Luo, H.; Zhang, Y.; Li, Q.; Zhu, Y.; Xu, J.; Li, Y.; Song, C.; Wang, B.; et al. Analysis of the Genome Sequence of the Medicinal Plant Salvia miltiorrhiza. Mol. Plant 2016, 9, 949–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.-c.; Ji, A.-j.; Zhang, X.; Song, J.-y.; Chen, S.-l. Biosynthesis and Regulation of Active Compounds in Medicinal Model Plant Salvia miltiorrhiza. Chin. Herb. Med. 2016, 8, 3–11. [Google Scholar] [CrossRef]
- Kiełbowicz-Matuk, A.; Rey, P.; Rorat, T. Interplay between circadian rhythm, time of the day and osmotic stress constraints in the regulation of the expression of a Solanum Double B-box gene. Ann. Bot. 2014, 113, 831–842. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Q.; Guthrie, C.; Sarmast, M.K.; Dehesh, K. BBX19 interacts with CONSTANS to repress FLOWERING LOCUS T transcription, defining a flowering time checkpoint in Arabidopsis. Plant Cell 2014, 26, 3589–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imtiaz, M.; Yang, Y.; Liu, R.; Xu, Y.; Khan, M.A.; Wei, Q.; Gao, J.; Hong, B. Identification and functional characterization of the BBX24 promoter and gene from chrysanthemum in Arabidopsis. Plant Mol. Biol. 2015, 89, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huai, J.; Shang, F.; Xu, G.; Tang, W.; Jing, Y.; Lin, R. A PIF1/PIF3-HY5-BBX23 Transcription Factor Cascade Affects Photomorphogenesis. Plant Physiol. 2017, 174, 2487–2500. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zeng, J.; Deng, K.; Tu, X.; Zhao, X.; Tang, D.; Liu, X. DBB1a, involved in gibberellin homeostasis, functions as a negative regulator of blue light-mediated hypocotyl elongation in Arabidopsis. Planta 2011, 233, 13–23. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, X.; Aiwaili, P.; Mu, X.; Zhao, M.; Zhao, J.; Cheng, L.; Ma, C.; Gao, J.; Hong, B. A zinc finger protein BBX19 interacts with ABF3 to affect drought tolerance negatively in chrysanthemum. Plant J. Cell Mol. Biol. 2020, 103, 1783–1795. [Google Scholar] [CrossRef]
- Min, J.H.; Chung, J.S.; Lee, K.H.; Kim, C.S. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J. Integr. Plant Biol. 2015, 57, 313–324. [Google Scholar] [CrossRef]
- Job, N.; Yadukrishnan, P.; Bursch, K.; Datta, S.; Johansson, H. Two B-Box Proteins Regulate Photomorphogenesis by Oppositely Modulating HY5 through their Diverse C-Terminal Domains. Plant Physiol. 2018, 176, 2963–2976. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Tao, R.; Yin, L.; Ni, J.; Yang, Q.; Yan, X.; Yang, F.; Guo, X.; Li, H.; Teng, Y. Two B-box proteins, PpBBX18 and PpBBX21, antagonistically regulate anthocyanin biosynthesis via competitive association with Pyrus pyrifolia ELONGATED HYPOCOTYL 5 in the peel of pear fruit. Plant J. Cell Mol. Biol. 2019, 100, 1208–1223. [Google Scholar] [CrossRef]
- Li, C.; Pei, J.; Yan, X.; Cui, X.; Tsuruta, M.; Liu, Y.; Lian, C. A poplar B-box protein PtrBBX23 modulates the accumulation of anthocyanins and proanthocyanidins in response to high light. Plant Cell Environ. 2021, 44, 3015–3033. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, Y.; Xie, C.; Zu, Y.; Zhan, F.; Mei, X.; Xia, Y.; Li, Y. Effects of UV-B radiation on the infectivity of Magnaporthe oryzae and rice disease-resistant physiology in Yuanyang terraces. Photochem. Photobiol. Sci. 2018, 17, 8–17. [Google Scholar] [CrossRef]
- Hatier, J.H.; Clearwater, M.J.; Gould, K.S. The functional significance of black-pigmented leaves: Photosynthesis, photoprotection and productivity in Ophiopogon planiscapus ‘Nigrescens’. PLoS ONE 2013, 8, e67850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yan, Y.; Wan, P.; Dong, W.; Huang, K.; Ran, L.; Mi, J.; Lu, L.; Zeng, X.; Cao, Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef]
- Song, Z.; Yan, T.; Liu, J.; Bian, Y.; Heng, Y.; Lin, F.; Jiang, Y.; Wang Deng, X.; Xu, D. BBX28/BBX29, HY5 and BBX30/31 form a feedback loop to fine-tune photomorphogenic development. Plant J. 2020, 104, 377–390. [Google Scholar] [CrossRef]
- Gangappa, S.N.; Botto, J.F. The Multifaceted Roles of HY5 in Plant Growth and Development. Mol. Plant 2016, 9, 1353–1365. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.; Ravindran, N.; Singh, D.; Rahul, P.V.; Datta, S. Role of Arabidopsis BBX proteins in light signaling. J. Plant Biochem. Biotechnol. 2020, 29, 623–635. [Google Scholar] [CrossRef]
- Bursch, K.; Toledo-Ortiz, G.; Pireyre, M.; Lohr, M.; Braatz, C.; Johansson, H. Identification of BBX proteins as rate-limiting cofactors of HY5. Nat. Plants 2020, 6, 921–928. [Google Scholar] [CrossRef]
- Poole, R.L. The TAIR database. Methods Mol. Biol. 2007, 406, 179–212. [Google Scholar] [CrossRef] [PubMed]
- Chandran, A.K.N.; Jung, K.H. Resources for systems biology in rice. J. Plant Biol. 2014, 57, 80–92. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.; Tosatto, S.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool:identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Li, K.B. ClustalW-MPI: ClustalW analysis using distributed and parallel computing. Bioinformatics 2003, 19, 1585–1586. [Google Scholar] [CrossRef] [Green Version]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Moreira, D. Orthologous Gene. In Encyclopedia of Astrobiology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucleic Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | CDS Length (bp) | AA | Mw (Da) | PI | Instability Index | Aliphatic Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| SmBBX1 | 969 | 322 | 35,644.8 | 5.96 | 42.37 | 64.04 | −0.592 | Nucleus |
| SmBBX2 | 648 | 215 | 23,113.5 | 4.61 | 78.87 | 60.00 | −0.529 | Nucleus |
| SmBBX3 | 972 | 323 | 35,792.9 | 5.28 | 62.17 | 58.02 | −0.629 | Nucleus |
| SmBBX4 | 993 | 330 | 37,849.6 | 5.65 | 63.70 | 63.00 | −0.758 | Nucleus |
| SmBBX5 | 1218 | 405 | 44,061.6 | 5.26 | 52.62 | 70.20 | −0.323 | Nucleus |
| SmBBX6 | 1086 | 361 | 40,530.3 | 5.62 | 60.45 | 62.69 | −0.638 | Nucleus |
| SmBBX7 | 1233 | 410 | 45,004.3 | 5.84 | 58.08 | 54.76 | −0.567 | Nucleus |
| SmBBX8 | 1212 | 403 | 44,222.4 | 5.71 | 57.96 | 57.42 | −0.574 | Nucleus |
| SmBBX9 | 1155 | 384 | 42,913.5 | 5.31 | 55.86 | 54.66 | −0.754 | Nucleus |
| SmBBX10 | 1128 | 375 | 42,417.0 | 5.78 | 53.29 | 60.59 | −0.776 | Nucleus |
| SmBBX11 | 1005 | 334 | 36,544.8 | 5.54 | 63.69 | 63.47 | −0.473 | Nucleus |
| SmBBX12 | 450 | 149 | 16,197.1 | 5.34 | 56.97 | 56.31 | −0.555 | Nucleus |
| SmBBX13 | 816 | 271 | 30,326.2 | 4.60 | 64.64 | 59.08 | −0.788 | Nucleus |
| SmBBX14 | 1290 | 429 | 47,868.3 | 5.37 | 51.67 | 67.53 | −0.691 | Nucleus |
| SmBBX15 | 1209 | 402 | 45,540.8 | 5.84 | 62.90 | 60.27 | −0.816 | Nucleus |
| SmBBX16 | 1113 | 370 | 39,824.7 | 6.01 | 35.11 | 69.00 | −0.282 | Nucleus |
| SmBBX17 | 1164 | 387 | 41,921.0 | 6.79 | 45.31 | 63.62 | −0.323 | Nucleus |
| SmBBX18 | 645 | 214 | 23,350.0 | 5.65 | 46.37 | 58.79 | −0.686 | Nucleus |
| SmBBX19 | 543 | 180 | 19,742.3 | 5.81 | 39.20 | 72.72 | −0.422 | Nucleus |
| SmBBX20 | 588 | 195 | 21,500.9 | 5.51 | 49.53 | 65.64 | −0.581 | Nucleus |
| SmBBX21 | 861 | 286 | 31,512.2 | 6.69 | 63.30 | 65.91 | −0.435 | Nucleus |
| SmBBX22 | 771 | 256 | 28,021.6 | 5.29 | 39.46 | 71.64 | −0.324 | Nucleus |
| SmBBX23 | 897 | 298 | 32,347.2 | 5.75 | 43.19 | 64.87 | −0.443 | Nucleus |
| SmBBX24 | 723 | 240 | 26,574.0 | 5.08 | 43.84 | 75.21 | −0.432 | Nucleus |
| SmBBX25 | 1293 | 430 | 46,981.8 | 6.19 | 56.66 | 62.93 | −0.607 | Nucleus |
| SmBBX26 | 666 | 221 | 24,120.2 | 5.72 | 67.91 | 67.15 | −0.524 | Nucleus |
| SmBBX27 | 723 | 240 | 25,808.4 | 8.92 | 64.57 | 72.00 | −0.198 | Nucleus |
| Name | AA(aa) | Domains | BBX1 | BBX2 | CCT | Structure |
|---|---|---|---|---|---|---|
| SmBBX1 | 322 | 2BBX + CCT | 20–59 | 59–102 | 257–300 |  |
| SmBBX2 | 215 | 1BBX | 4–42 | - | - |  |
| SmBBX3 | 323 | 1BBX + CCT | 49–92 | - | 266–309 |  |
| SmBBX4 | 330 | 1BBX + CCT | 16–59 | - | 297–324 |  |
| SmBBX5 | 405 | 1BBX + CCT | 5–48 | - | 347–390 |  |
| SmBBX6 | 361 | 1BBX + CCT | 20–60 | - | 317–360 |  |
| SmBBX7 | 410 | 2BBX + CCT | 4–47 | 47–90 | 353–396 |  |
| SmBBX8 | 403 | 2BBX + CCT | 4–47 | 47–90 | 346–389 |  |
| SmBBX9 | 384 | 1BBX + CCT | 4–47 | - | 327–370 |  |
| SmBBX10 | 375 | 1BBX | 5–47 | - | - |  |
| SmBBX11 | 334 | 2BBX + CCT | 12–55 | 55–98 | 270–313 |  |
| SmBBX12 | 149 | 1BBX | 3–45 | - | - |  |
| SmBBX13 | 271 | 1BBX | 39–80 | - | - |  |
| SmBBX14 | 429 | 1BBX + CCT | 19–62 | - | 373–415 |  |
| SmBBX15 | 402 | 1BBX + CCT | 19–62 | - | 347–389 |  |
| SmBBX16 | 371 | 2BBX + CCT | 26–68 | 68–108 | 298–340 |  |
| SmBBX17 | 387 | 2BBX + CCT | 27–69 | 70–112 | 315–357 |  |
| SmBBX18 | 214 | 2BBX | 4–47 | 56–96 | - |  |
| SmBBX19 | 180 | 1BBX | 4–47 | - | - |  |
| SmBBX20 | 195 | 2BBX | 5–47 | 58–100 | - |  |
| SmBBX21 | 286 | 2BBX | 4–47 | 57–100 | - |  |
| SmBBX22 | 256 | 2BBX | 5–47 | 56–99 | - |  |
| SmBBX23 | 298 | 2BBX | 5–47 | 56–99 | - |  |
| SmBBX24 | 240 | 2BBX | 5–47 | 57–98 | - |  |
| SmBBX25 | 430 | 1BBX + CCT | 4–46 | - | 382–424 |  |
| SmBBX26 | 221 | 1BBX | 4–43 | - | - |  |
| SmBBX27 | 240 | 1BBX | 5–45 | - | - |  |
| Duplicate Gene Pair | Ka | Ks | Ka/Ks | Purify Selection |
|---|---|---|---|---|
| SmBBX11/SmBBX3 | 0.1503 | 1.3306 | 0.1130 | Yes |
| SmBBX16/SmBBX17 | 0.2114 | 1.5300 | 0.1381 | Yes |
| SmBBX4/SmBBX14 | 0.4317 | 59.5177 | 0.0073 | Yes |
| SmBBX4/SmBBX15 | 0.4945 | 65.5893 | 0.0075 | Yes |
| SmBBX18/SmBBX19 | 0.2161 | 53.5979 | 0.0040 | Yes |
| SmBBX20/SmBBX21 | 0.2850 | 6.5822 | 0.0433 | Yes |
| SmBBX22/SmBBX23 | 0.3118 | 1.4688 | 0.2126 | Yes |
| SmBBX25/SmBBX6 | 0.6270 | 3.3931 | 0.1848 | Yes |
| SmBBX7/SmBBX8 | 0.2402 | 0.7176 | 0.3348 | Yes |
| SmBBX5/SmBBX9 | 0.7136 | 52.7066 | 0.0135 | Yes |
| SmBBX10/SmBBX27 | 0.8913 | 60.0470 | 0.0148 | Yes |
| SmBBX12/SmBBX13 | 0.5215 | 2.7516 | 0.1895 | Yes |
| SmBBX2/SmBBX26 | 0.4775 | 1.7270 | 0.2765 | Yes |
| SmBBX13/SmBBX26 | 0.5752 | 49.5077 | 0.0116 | Yes |
| SmBBX11/SmBBX3 | 0.1503 | 1.3306 | 0.1130 | Yes |
| SmBBX16/SmBBX17 | 0.2114 | 1.5300 | 0.1381 | Yes |
| SmBBX4/SmBBX14 | 0.4317 | 59.5177 | 0.0073 | Yes |
| SmBBX4/SmBBX15 | 0.4945 | 65.5893 | 0.0075 | Yes |
| SmBBX18/SmBBX19 | 0.2161 | 53.5979 | 0.0040 | Yes |
| SmBBX20/SmBBX21 | 0.2850 | 6.5822 | 0.0433 | Yes |
| SmBBX22/SmBBX23 | 0.3118 | 1.4688 | 0.2126 | Yes |
| SmBBX25/SmBBX6 | 0.6270 | 3.3931 | 0.1848 | Yes |
| SmGene ID | AtGene ID | (At) Other Names |
|---|---|---|
| SmBBX1 | AT3G02380 | AtBBX3, COL2 |
| SmBBX5 | AT4G15250 | AtBBX9, COL4 |
| SmBBX6 | AT2G47890 | AtBBX11, ATCOL5 |
| SmBBX7 | AT3G07650 | AtBBX7, COL9 |
| SmBBX10 | AT1G68190 | AtBBX27 |
| SmBBX11 | AT5G24930 | AtBBX5 |
| SmBBX12 | AT4G27310 | AtBBX28 |
| SmBBX13 | AT5G54470 | AtBBX29 |
| SmBBX14 | AT1G25440 | AtBBX15 |
| SmBBX16 | AT5G57660 | AtBBX6, COL5 |
| SmBBX18 | AT4G38960 | AtBBX19 |
| SmBBX20 | AT4G39070 | AtBBX20, BZS1, STH7 |
| SmBBX21 | AT1G75540 | AtBBX21, STH2 |
| SmBBX23 | AT1G78600 | AtBBX22, LZF1, STH3 |
| SmBBX24 | AT1G06040 | AtBBX24, STO |
| SmBBX25 | AT2G33500 | AtBBX12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Tong, Y.; Ye, J.; Zhang, C.; Li, B.; Hu, S.; Xue, X.; Tian, Q.; Wang, Y.; Li, L.; et al. Genome-Wide Characterization of B-Box Gene Family in Salvia miltiorrhiza. Int. J. Mol. Sci. 2023, 24, 2146. https://doi.org/10.3390/ijms24032146
Li Y, Tong Y, Ye J, Zhang C, Li B, Hu S, Xue X, Tian Q, Wang Y, Li L, et al. Genome-Wide Characterization of B-Box Gene Family in Salvia miltiorrhiza. International Journal of Molecular Sciences. 2023; 24(3):2146. https://doi.org/10.3390/ijms24032146
Chicago/Turabian StyleLi, Yunyun, Yunli Tong, Jun Ye, Caijuan Zhang, Bin Li, Suying Hu, Xiaoshan Xue, Qian Tian, Yueyue Wang, Lin Li, and et al. 2023. "Genome-Wide Characterization of B-Box Gene Family in Salvia miltiorrhiza" International Journal of Molecular Sciences 24, no. 3: 2146. https://doi.org/10.3390/ijms24032146