
Genome-Wide Identification and Phylogenetic and Expression Analyses of the PLATZ Gene Family in Medicago sativa L.
 , ,
, ,
Abstract
:1. Introduction
2. Results
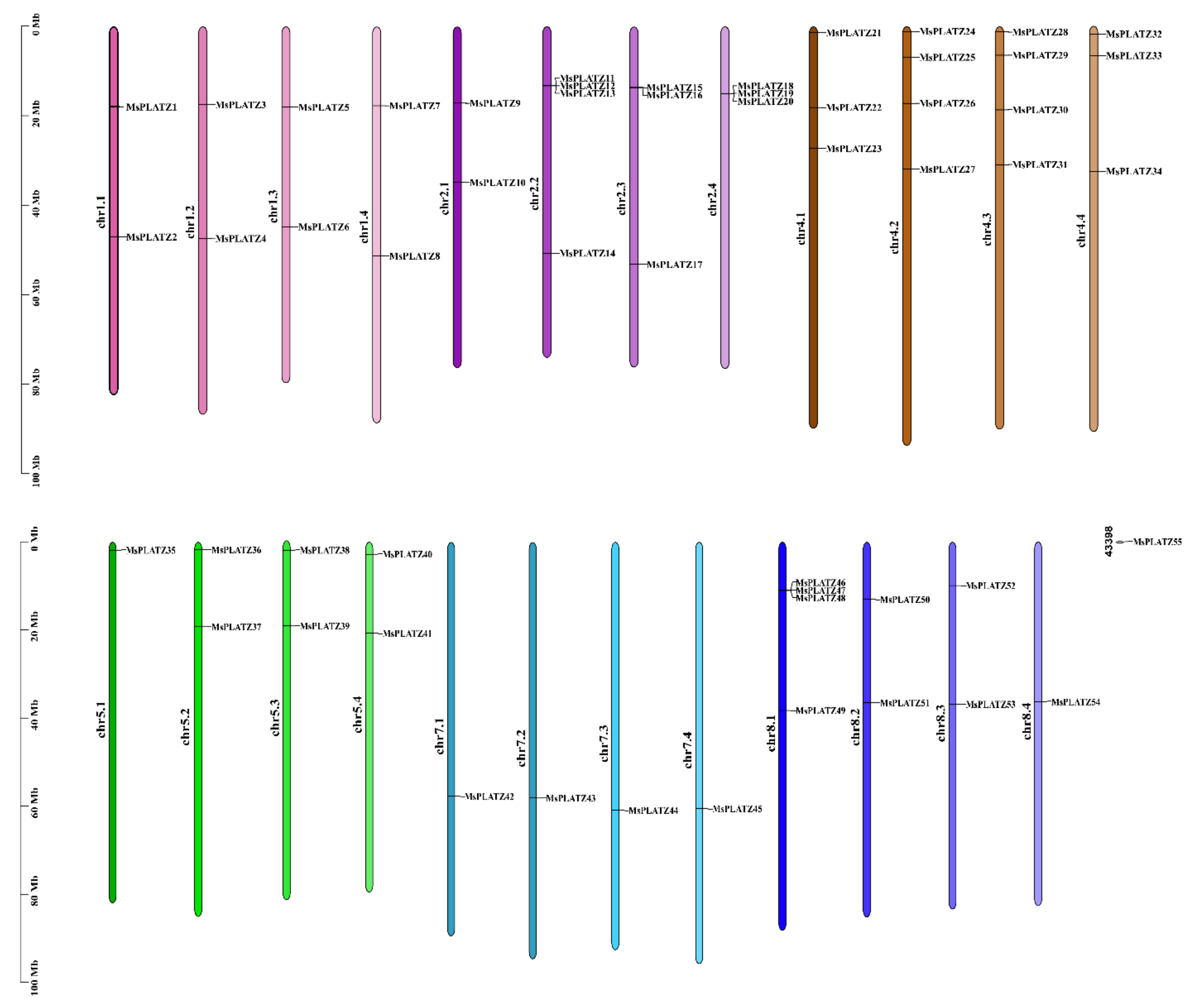
2.1. Identification of PLATZ Genes in the Alfalfa Genome
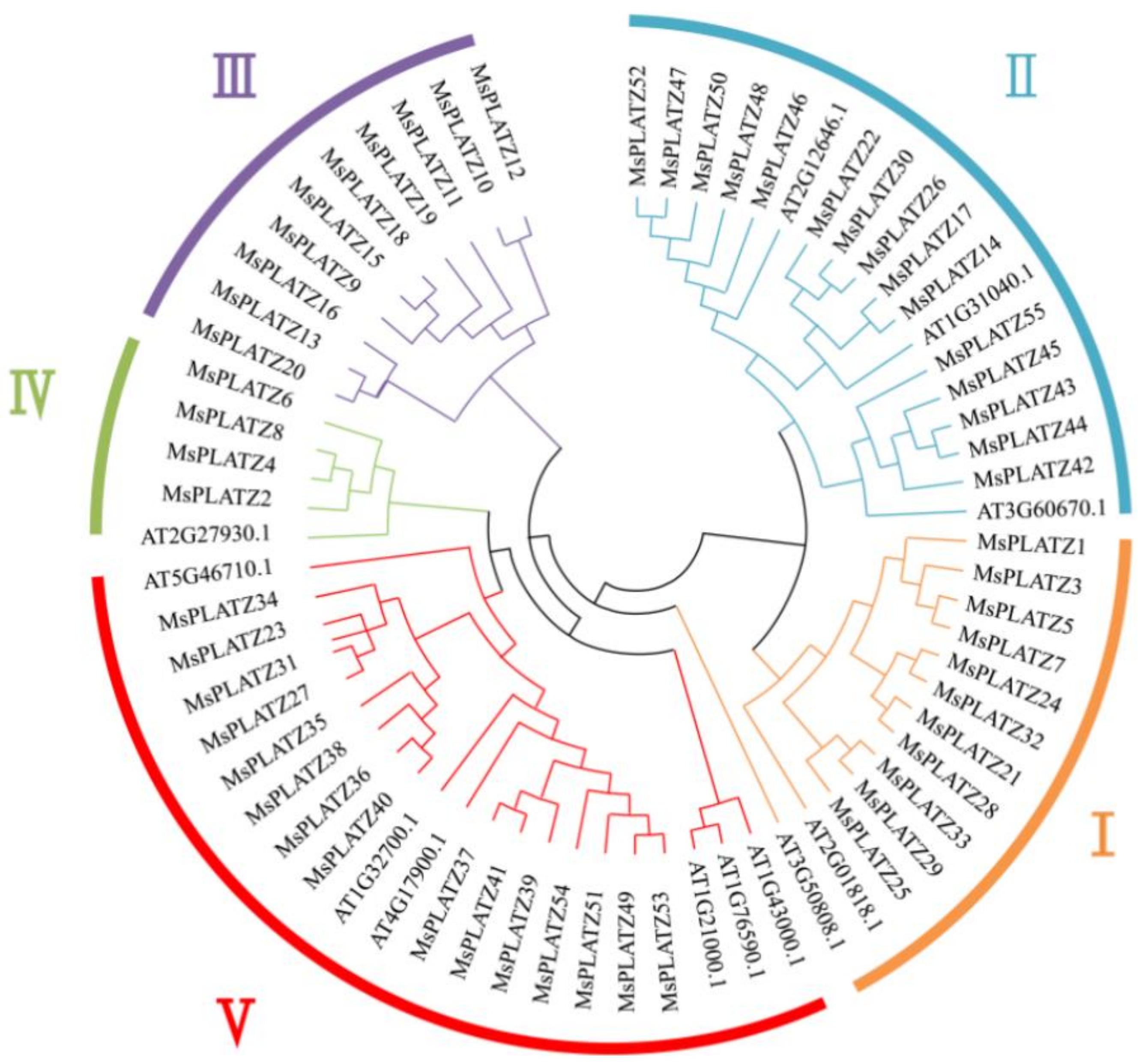
2.2. Phylogenetic Analysis and Classification of MsPLATZ Proteins
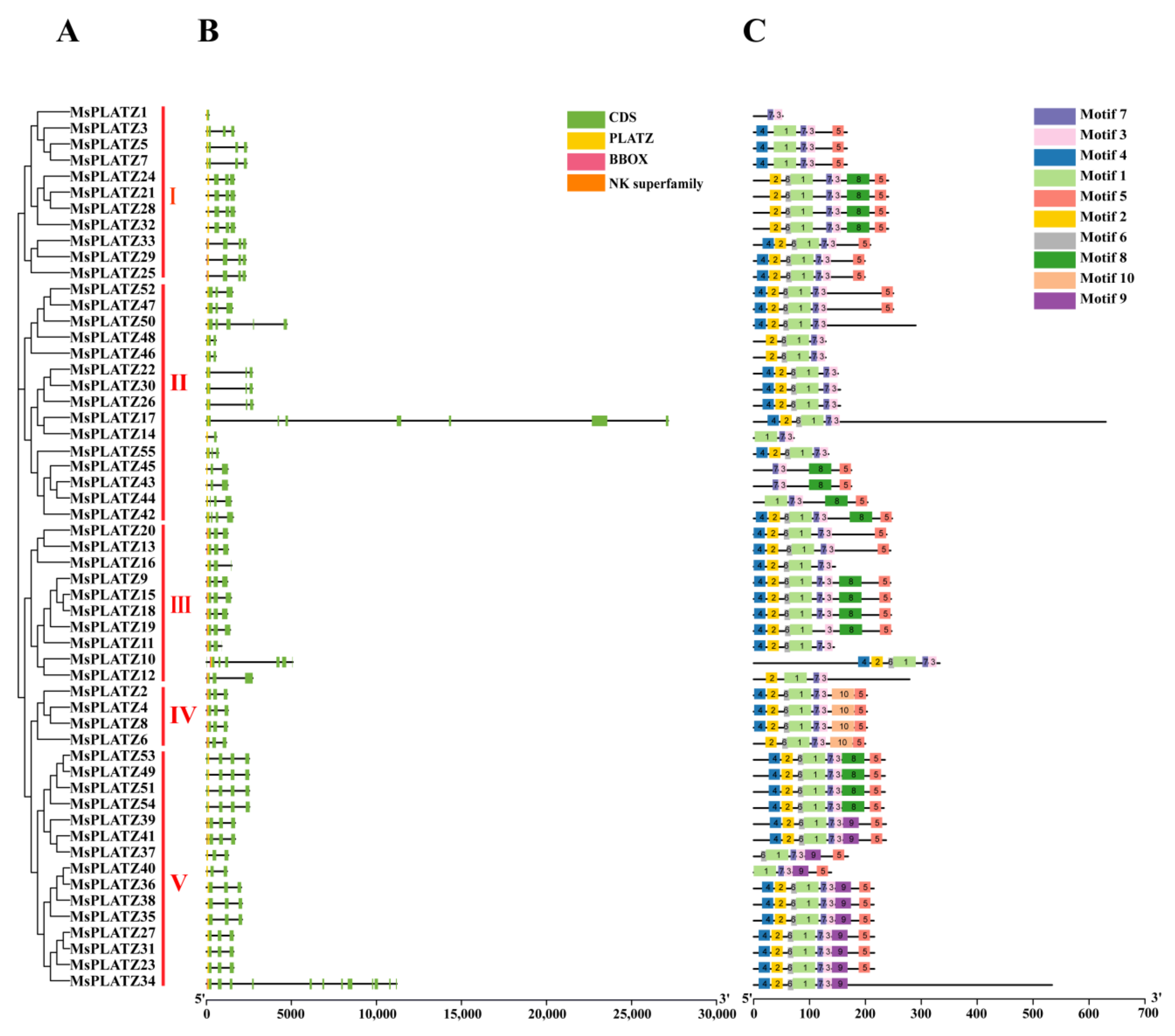
2.3. Gene structure and Conserved Motif Analysis of MsPLATZ Genes
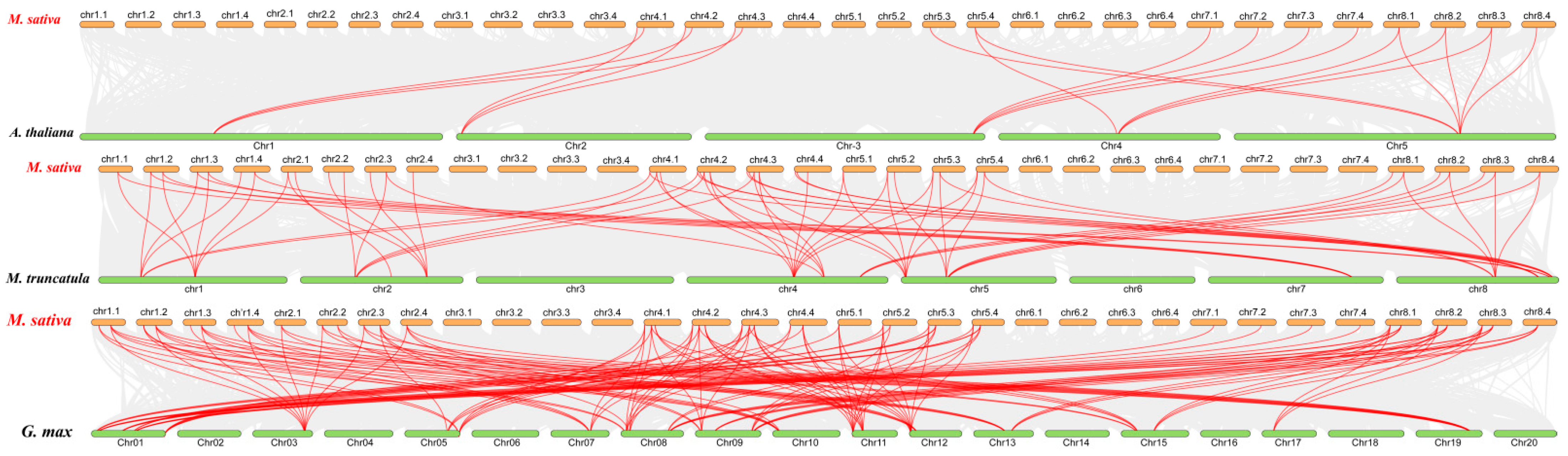
2.4. Gene Duplication Events and Synteny Analysis of MsPLATZ Genes
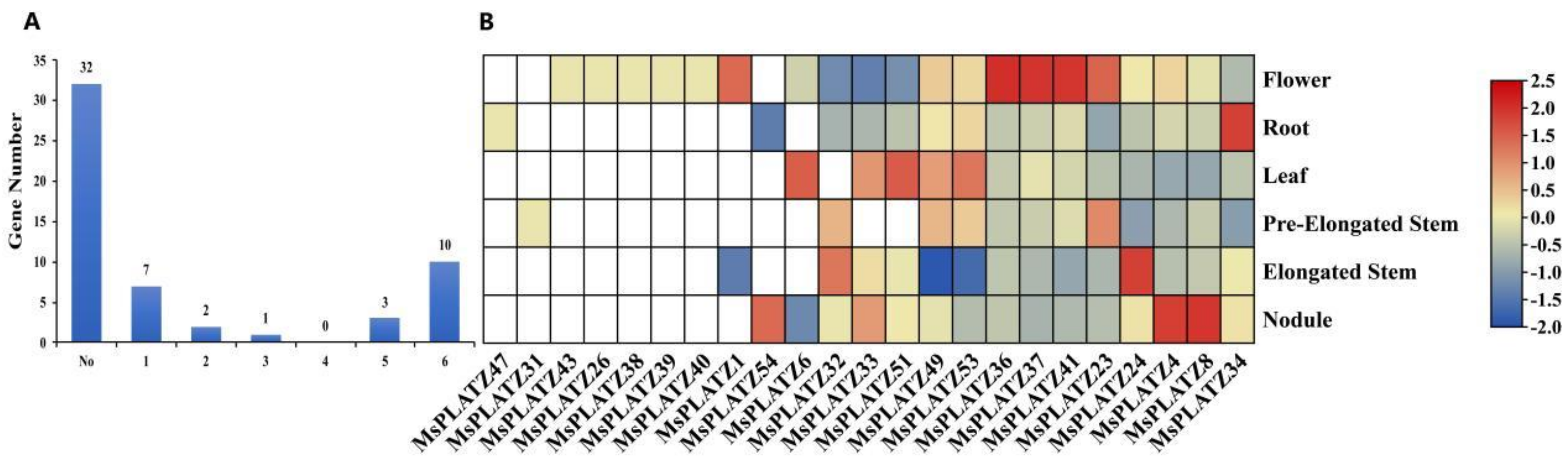
2.5. Expression Patterns of MsPLATZ Genes in Different Tissues of Medicago sativa
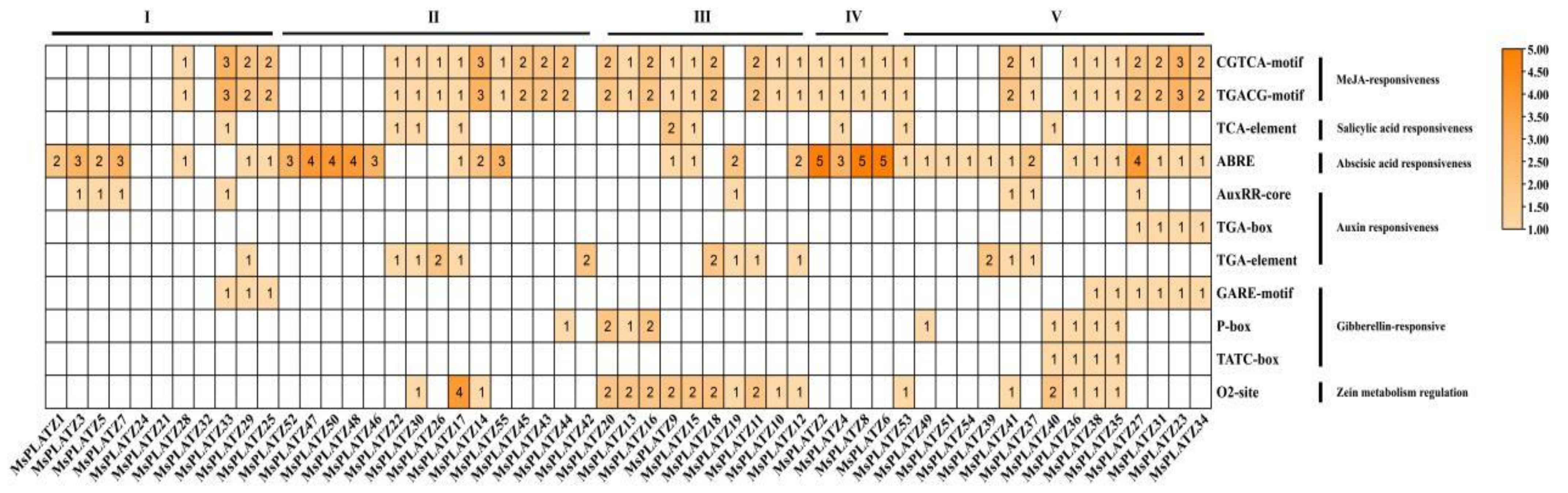
2.6. Analysis of Promoter Cis-Acting Elements of MsPLATZ Genes
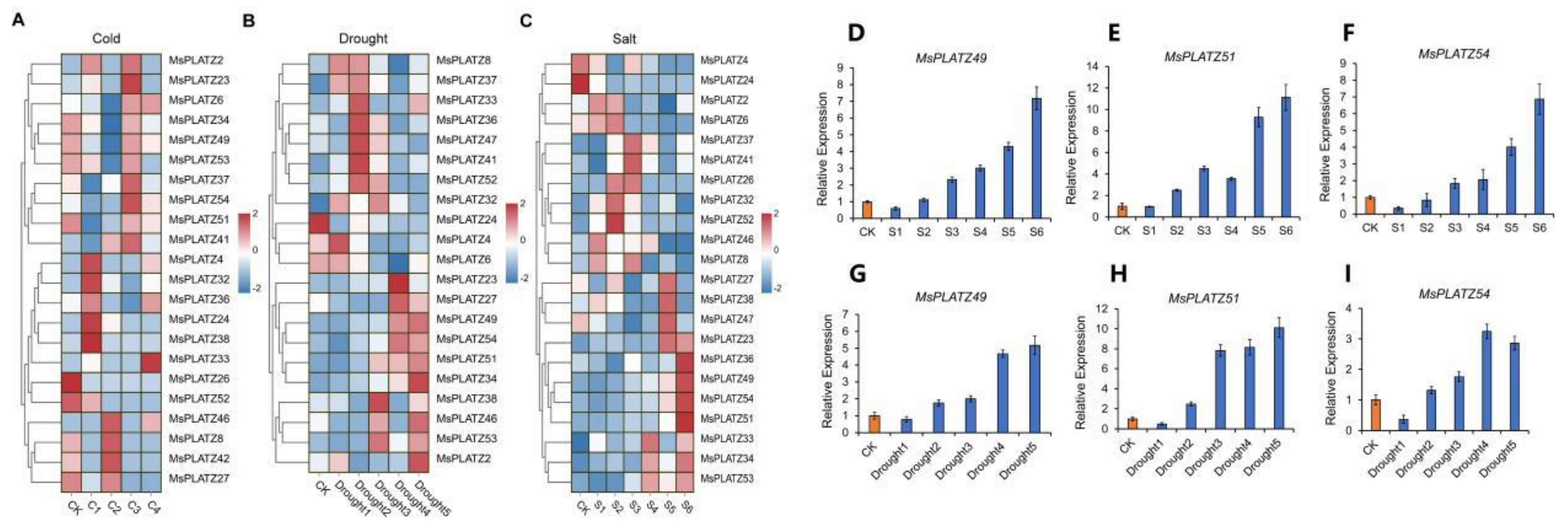
2.7. Differential Expression of MsPLATZ Genes under Different Abiotic Stress Treatments
3. Discussion
4. Materials and Methods
4.1. Identification of MsPLATZ Genes in the Medicago sativa Genome
4.2. Phylogenetic Analysis and Intron–Exon Structure Determination
4.3. Gene Duplication, Synteny Analysis and Cis-Element Analysis
4.4. Plant Materials and Treatments
4.5. Transcriptome Data Collection and Analysis
4.6. Expression Analyses of MsPLATZ Genes by qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Honma, T.; Goto, K. Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.B.; Foley, R.C.; Oñate-Sánchez, L. Transcription factors in plant defense and stress responses. Curr. Opin. Plant Biol. 2002, 5, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP family of plant transcription factors. Biol. Chem. 1998, 379, 633–646. [Google Scholar]
- Martin, C.; Paz-Ares, J. MYB transcription factors in plants. Trends Genet. 1997, 13, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bakshi, M.; Oelmuller, R. WRKY transcription factors: Jack of many trades in plants. Plant Signal. Behav. 2014, 9, e27700. [Google Scholar] [CrossRef] [Green Version]
- Takatsuji, H. Zinc-finger transcription factors in plants. Cell Mol. Life Sci. 1998, 54, 582–596. [Google Scholar] [CrossRef]
- Nantel, A.; Quatrano, R.S. Characterization of three rice basic/leucine zipper factors, including two inhibitors of Em BP-1 DNA binding activity. J. Biol. Chem. 1996, 271, 31296–31305. [Google Scholar] [CrossRef] [Green Version]
- Nagano, Y.; Furuhashi, H.; Inaba, T.; Sasaki, Y. A novel class of plant-specific zinc-dependent DNA-binding protein that binds to A/T-rich DNA sequences. Nucleic Acids Res. 2001, 29, 4097–4105. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Hou, Q.; Si, L.; Huang, X.; Luo, J.; Lu, D.; Zhu, J.; Shangguan, Y.; Miao, J.; Xie, Y.; et al. The PLATZ transcription factor GL6 affects grain length and number in rice. Plant Physiol. 2019, 180, 2077–2090. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.R.; Xue, H.W. The rice PLATZ protein SHORT GRAIN6 determines grain size by regulating spikelet hull cell division. J. Integr. Plant Biol. 2020, 62, 847–864. [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Ye, J.; Zheng, X.; Xiang, X.; Li, C.; Fu, M.; Wang, Q.; Zhang, Z.; Wu, Y. The Maize imprinted gene Floury3 encodes a PLATZ protein required for t RNA and 5S rRNA transcription through interaction with RNA polymerase III. Plant Cell 2017, 29, 2661–2675. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Kim, J.; Jun, S.E.; Park, S.; Timilsina, R.; Kwon, D.S.; Kim, Y.; Park, S.J.; Hwang, J.Y.; Nam, H.G.; et al. ORESARA15, a PLATZ transcription factor, mediates leaf growth and senescence in Arabidopsis. New Phytol. 2018, 220, 609–623. [Google Scholar] [CrossRef] [Green Version]
- Hande, A.S.; Katageri, I.S.; Jadhav, M.P.; Adiger, S.; Gamanagatti, S.; Padmalatha, K.V.; Dhandapani, G.; Kanakachari, M.; Kumar, P.A.; Reddy, V.S. Transcript profiling of genes expressed during fibre development in diploid cotton (Gossypium arboreum L.). BMC Genom. 2017, 18, 675. [Google Scholar]
- Chao, Q.; Gao, Z.F.; Zhang, D.; Zhao, B.G.; Dong, F.Q.; Fu, C.X.; Liu, L.J.; Wang, B.C. The developmental dynamics of the Populus stem transcriptome. Plant Biotechnol. J. 2019, 17, 206–219. [Google Scholar] [CrossRef] [Green Version]
- Kasirajan, L.; Hoang, N.V.; Furtado, A.; Botha, F.C.; Henry, R.J. Transcriptome analysis highlights key differentially expressed genes involved in cellulose and lignin biosynthesis of sugarcane genotypes varying in fiber content. Sci. Rep. 2018, 8, 11612. [Google Scholar] [CrossRef] [Green Version]
- González-Morales, S.I.; Chávez-Montes, R.A.; Hayano-Kanashiro, C.; Alejo-Jacuinde, G.; Rico-Cambron, T.Y.; de Folter, S.; Herrera-Estrella, L. Regulatory network analysis reveals novel regulators of seed desiccation tolerance in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E5232–E5241. [Google Scholar] [CrossRef] [Green Version]
- Zenda, T.; Liu, S.; Wang, X.; Liu, G.; Jin, H.; Dong, A.; Yang, Y.; Duan, H. Key maize drought-responsive genes and pathways revealed by comparative transcriptome and physiological analyses of contrasting inbred lines. Int. J. Mol. Sci. 2019, 20, 1268. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, R.; Huo, Y.; Liu, S.; Yang, G.; Huang, J.; Zheng, C.; Wu, C. Expression of cotton PLATZ1 in transgenic Arabidopsis reduces sensitivity to osmotic and salt stress for germination and seedling establishment associated with modification of the abscisic acid, gibberellin, and ethylene signalling pathways. BMC Plant Biol. 2018, 18, 218. [Google Scholar] [CrossRef] [Green Version]
- Yari, M.; Valizadeh, R.; Naserian, A.A.; Ghorbani, G.R.; Moghaddam, P.R.; Jonker, A.; Yu, P. Botanical traits, protein and carbohydrate fractions, ruminal degradability and energy contents of alfalfa hay harvested at three stages of maturity and in the afternoon and morning. Anim. Feed. Sci. Technol. 2012, 172, 162–170. [Google Scholar] [CrossRef]
- Radović, J.; Sokolović, D.; Marković, J. Alfalfa-most important perennial forage legume in animal husbandry. Biotechnol. Anim. Husb. 2009, 25, 465–475. [Google Scholar] [CrossRef]
- Chen, L.; He, F.; Long, R.; Zhang, F.; Li, M.; Wang, Z.; Kang, J.; Yang, Q. A global alfalfa diversity panel reveals genomic selection signatures in Chinese varieties and genomic associations with root development. Integr. Plant Biol. 2021, 63, 1937–1951. [Google Scholar] [CrossRef]
- Jafari, M.; Yari, M.; Ghabooli, M.; Sepehri, M.; Ghasemi, E.; Jonker, A. Inoculation and co-inoculation of alfalfa seedlings with root growth promoting microorganisms (Piriformospora indica, Glomus intraradices and Sinorhizobium meliloti) affect molecular structures, nutrient profiles and availability of hay for ruminants. Anim. Nutr. 2018, 4, 90–99. [Google Scholar] [CrossRef]
- Chen, H.; Zeng, Y.; Yang, Y.; Huang, L.; Tang, B.; Zhang, H.; Hao, F.; Liu, W.; Li, Y.; Liu, Y.; et al. Allele-aware chromosome-level genome assembly and efcient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat Commun. 2020, 11, 2494. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, R.; Liu, M.; Zhang, S.; Yan, K.; Yang, G.; Huang, J.; Zheng, C.; Wu, C. PLATZ2 negatively regulates salt tolerance in Arabidopsis seedlings by directly suppressing the expression of the CBL4/SOS3 and CBL10/SCaBP8 genes. J. Exp. Bot. 2020, 71, 5589–5602. [Google Scholar] [CrossRef]
- Wang, J.; Ji, C.; Li, Q.; Zhou, Y.; Wu, Y. Genome-wide analysis of the plantspecifc PLATZ proteins in maize and identifcation of their general role in interaction with RNA polymerase III complex. BMC Plant Biol. 2018, 18, 221. [Google Scholar] [CrossRef]
- Li, J.; Feng, S.; Zhang, Y.; Xu, L.; Luo, Y.; Yuan, Y.; Yang, Q.; Feng, B. Genome-wide identification and expression analysis of the plant-specific PLATZ gene family in Tartary buckwheat (Fagopyrum tataricum). BMC Plant Biol. 2022, 22, 160. [Google Scholar]
- Han, X.; Rong, H.; Tian, Y.; Qu, Y.; Xu, M.; Xu, L.A. Genome-Wide Identification of PLATZ Transcription Factors in Ginkgo biloba L. and Their Expression Characteristics During Seed Development. Front. Plant Sci. 2022, 13, 946194. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Cheng, M.; Li, M.; Guo, X.; Wu, Y.; Wang, J. Identification and Characterization of PLATZ Transcription Factors in Wheat. Int. J. Mol. Sci. 2020, 21, 8934. [Google Scholar] [CrossRef]
- Yamada, M.; Han, X.; Benfey, P.N. RGF1 controls root meristem size through ROS signalling. Nature 2020, 577, 85–88. [Google Scholar] [CrossRef]
- He, F.; Zhang, L.; Zhao, G.; Kang, J.; Long, R.; Li, M.; Yang, Q.; Chen, L. Genome-Wide Identification and Expression Analysis of the NAC Gene Family in Alfalfa Revealed Its Potential Roles in Response to Multiple Abiotic Stresses. Int. J. Mol. Sci. 2022, 23, 10015. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Kiełbowicz-Matuk, A. Involvement of plant C2H2 -type zinc fifinger transcription factors in stress responses. Plant Sci. Int. J. Exp. Plant Biol. 2012, 185–186, 78–85. [Google Scholar]
- He, F.; Wei, C.; Zhang, Y.; Long, R.; Li, M.; Wang, Z.; Yang, Q.; Kang, J.; Chen, L. Genome-wide association analysis coupled with transcriptome analysis reveals candidate genes related to salt stress in alfalfa (Medicago sativa L). Front. Plant Sci. 2022, 12, 3407. [Google Scholar] [CrossRef]
- He, F.; Long, R.; Wei, C.; Zhang, Y.; Li, M.; Kang, J.; Yang, Q.; Wang, Z.; Chen, L. Genome-wide identification, phylogeny and expression analysis of the SPL gene family and its important role in salt stress in Medicago sativa L. BMC Plant Biol. 2022, 22, 295. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exonintron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- O‘Rourke, J.A.; Fu, F.; Bucciarelli, B.; Yang, S.S.; Samac, D.A.; Lamb, J.F.; Monteros, M.J.; Graham, M.A.; Gronwald, J.W.; Krom, N.; et al. The Medicago sativa gene index 1.2: A web-accessible gene expression atlas for investigating expression differences between Medicago sativa subspecies. BMC Genom. 2015, 16, 502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Deng, H.; Ma, W.; Zhou, Q.; Liu, Z. Genome-wide identification of the MADS-box transcription factor family in autotetraploid cultivated alfalfa (Medicago sativa L.) and expression analysis under abiotic stress. BMC Genom. 2021, 22, 603. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Kenneth, J.L.; Thomas, D.S. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr Location | CDS Length (bp) | Protein Length (aa) | Mw (kDa) | pI | Subcellular Location |
|---|---|---|---|---|---|---|---|
| MsPLATZ1 | MS.gene026494 | Ms1.1: 17,863,747–17,863,902 | 156 | 52 | 5.83 | 9.64 | Nucleus |
| MsPLATZ2 | MS.gene051384 | Ms1.1: 46,909,227–46,910,461 | 609 | 203 | 22.92 | 8.83 | Nucleus |
| MsPLATZ3 | MS.gene040928 | Ms1.2: 17,427,194–17,428,828 | 501 | 167 | 19.37 | 8.97 | Nucleus |
| MsPLATZ4 | MS.gene029705 | Ms1.2: 47,307,863–47,309,143 | 609 | 203 | 22.95 | 8.83 | Nucleus |
| MsPLATZ5 | MS.gene95330 | Ms1.3: 17,964,734–17,967,118 | 501 | 167 | 19.36 | 8.97 | Nucleus |
| MsPLATZ6 | MS.gene034971 | Ms1.3: 44,693,845–44,695,016 | 600 | 200 | 22.61 | 8.96 | Nucleus |
| MsPLATZ7 | MS.gene83024 | Ms1.4: 17,595,429–17,597,805 | 501 | 167 | 19.36 | 8.97 | Nucleus |
| MsPLATZ8 | MS.gene47395 | Ms1.4: 51,168,875–51,170,109 | 609 | 203 | 22.92 | 8.7 | Nucleus |
| MsPLATZ9 | MS.gene35330 | Ms2.1: 17,012,467–17,013,708 | 735 | 245 | 28.58 | 9.24 | Nucleus |
| MsPLATZ10 | MS.gene99752 | Ms2.1: 34,752,509–34,757,591 | 999 | 333 | 36.91 | 4.69 | Chloroplast |
| MsPLATZ11 | MS.gene073125 | Ms2.2: 13,169,890–13,170,791 | 432 | 144 | 16.70 | 9.03 | Cytosol |
| MsPLATZ12 | MS.gene073126 | Ms2.2: 13,185,522–13,188,245 | 837 | 279 | 31.60 | 5.81 | Extracellular |
| MsPLATZ13 | MS.gene073129 | Ms2.2: 13,219,049–13,220,333 | 735 | 245 | 28.46 | 8.71 | Extracellular |
| MsPLATZ14 | MS.gene57445 | Ms2.2: 50,631,783–50,632,379 | 216 | 72 | 8.39 | 8.85 | Chloroplast |
| MsPLATZ15 | MS.gene91274 | Ms2.3: 13,498,110–13,499,564 | 738 | 246 | 28.68 | 9.26 | Nucleus |
| MsPLATZ16 | MS.gene91270 | Ms2.3: 13,545,547–13,547,034 | 438 | 146 | 16.93 | 9.21 | Extracellular |
| MsPLATZ17 | MS.gene58706 | Ms2.3: 53,047,725–53,074,883 | 1890 | 630 | 72.48 | 9 | Nucleus |
| MsPLATZ18 | MS.gene94621 | Ms2.4: 14,906,505–14,907,746 | 738 | 246 | 28.71 | 9.26 | Nucleus |
| MsPLATZ19 | MS.gene94620 | Ms2.4: 14,914,416–14,915,814 | 741 | 247 | 28.69 | 9.1 | Nucleus |
| MsPLATZ20 | MS.gene94618 | Ms2.4: 14,941,107–14,942,368 | 714 | 238 | 27.88 | 8.79 | Extracellular |
| MsPLATZ21 | MS.gene015620 | Ms4.1: 1,317,444–1,319,118 | 723 | 241 | 27.86 | 8.94 | Nucleus |
| MsPLATZ22 | MS.gene36510 | Ms4.1: 18,036,419–18,039,121 | 453 | 151 | 16.90 | 8.57 | Nucleus |
| MsPLATZ23 | MS.gene065915 | Ms4.1: 27,206,398–27,208,000 | 648 | 216 | 23.96 | 9.31 | Nucleus |
| MsPLATZ24 | MS.gene62657 | Ms4.2: 1,067,724–1,069,373 | 723 | 241 | 27.90 | 9.03 | Nucleus |
| MsPLATZ25 | MS.gene72172 | Ms4.2: 6,826,199–6,828,514 | 597 | 199 | 22.67 | 8.82 | Nucleus |
| MsPLATZ26 | MS.gene028280 | Ms4.2: 17,145,854–17,148,601 | 465 | 155 | 17.48 | 8.57 | Nucleus |
| MsPLATZ27 | MS.gene83177 | Ms4.2: 31,843,428–31,845,025 | 648 | 216 | 23.96 | 9.31 | Nucleus |
| MsPLATZ28 | MS.gene95852 | Ms4.3: 1,135,074–1,136,748 | 723 | 241 | 27.86 | 8.94 | Nucleus |
| MsPLATZ29 | MS.gene048775 | Ms4.3: 6,372,228–6,374,549 | 597 | 199 | 22.67 | 8.82 | Nucleus |
| MsPLATZ30 | MS.gene023191 | Ms4.3: 18,576,051–18,578,761 | 465 | 155 | 17.40 | 8.57 | Nucleus |
| MsPLATZ31 | MS.gene031722 | Ms4.3: 30,835,572–30,837,171 | 648 | 216 | 23.96 | 9.31 | Nucleus |
| MsPLATZ32 | MS.gene058564 | Ms4.4: 1,743,997–1,745,674 | 723 | 241 | 27.89 | 9.03 | Nucleus |
| MsPLATZ33 | MS.gene030429 | Ms4.4: 6,545,886–6,548,219 | 627 | 209 | 23.65 | 8.81 | Nucleus |
| MsPLATZ34 | MS.gene006587 | Ms4.4: 32,411,516–32,422,710 | 1602 | 534 | 58.31 | 8.69 | Nucleus |
| MsPLATZ35 | MS.gene058083 | Ms5.1: 1,754,805–1,756,904 | 645 | 215 | 24.06 | 9.07 | Nucleus |
| MsPLATZ36 | MS.gene21784 | Ms5.2: 1,651,210–1,653,257 | 645 | 215 | 24.16 | 9.15 | Nucleus |
| MsPLATZ37 | MS.gene64179 | Ms5.2: 18,922,507–18,923,800 | 507 | 169 | 19.24 | 9.91 | Nucleus |
| MsPLATZ38 | MS.gene40512 | Ms5.3: 2,100,515–2,102,615 | 645 | 215 | 24.06 | 9.07 | Nucleus |
| MsPLATZ39 | MS.gene036303 | Ms5.3: 19,070,783–19,072,470 | 711 | 237 | 27.11 | 9.44 | Nucleus |
| MsPLATZ40 | MS.gene024806 | Ms5.4: 2,681,147–2,682,369 | 417 | 139 | 15.60 | 9.72 | Nucleus |
| MsPLATZ41 | MS.gene027893 | Ms5.4: 20,364,134–20,365,827 | 711 | 237 | 27.11 | 9.44 | Nucleus |
| MsPLATZ42 | MS.gene96355 | Ms7.1: 56,837,237–56,838,808 | 744 | 248 | 27.69 | 8.99 | Extracellular |
| MsPLATZ43 | MS.gene20118 | Ms7.2: 57,201,793–57,203,057 | 525 | 175 | 18.96 | 9.22 | Nucleus |
| MsPLATZ44 | MS.gene82045 | Ms7.3: 59,965,702–59,967,169 | 612 | 204 | 22.34 | 9.31 | Extracellular |
| MsPLATZ45 | MS.gene010738 | Ms7.4: 59,599,942–59,601,202 | 525 | 175 | 18.93 | 9.22 | Nucleus |
| MsPLATZ46 | MS.gene036665 | Ms8.1: 10,732,368–10,732,928 | 387 | 129 | 15.08 | 8.84 | Cytosol |
| MsPLATZ47 | MS.gene59920 | Ms8.1: 10,748,163–10,749,697 | 750 | 250 | 28.74 | 8.61 | Nucleus |
| MsPLATZ48 | MS.gene041385 | Ms8.1: 10,830,973–10,831,533 | 387 | 129 | 15.08 | 8.84 | Cytosol |
| MsPLATZ49 | MS.gene60992 | Ms8.1: 37,680,744–37,683,265 | 705 | 235 | 26.73 | 9.5 | Nucleus |
| MsPLATZ50 | MS.gene041387 | Ms8.2: 12,789,586–12,794,323 | 870 | 290 | 33.82 | 8.67 | Nucleus |
| MsPLATZ51 | MS.gene009009 | Ms8.2: 35,918,983–35,921,509 | 705 | 235 | 26.74 | 9.5 | Nucleus |
| MsPLATZ52 | MS.gene58786 | Ms8.3: 9,688,042–9,689,586 | 750 | 250 | 28.74 | 8.61 | Nucleus |
| MsPLATZ53 | MS.gene71193 | Ms8.3: 36,233,879–36,236,397 | 705 | 235 | 26.73 | 9.5 | Nucleus |
| MsPLATZ54 | MS.gene038444 | Ms8.4: 35,728,371–35,730,908 | 699 | 233 | 26.50 | 9.5 | Nucleus |
| MsPLATZ55 | MS.gene81453 | 43398: 41,531–42,243 | 402 | 134 | 15.69 | 9.1 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; He, F.; Zhao, G.; Li, M.; Long, R.; Kang, J.; Yang, Q.; Chen, L. Genome-Wide Identification and Phylogenetic and Expression Analyses of the PLATZ Gene Family in Medicago sativa L. Int. J. Mol. Sci. 2023, 24, 2388. https://doi.org/10.3390/ijms24032388
Li X, He F, Zhao G, Li M, Long R, Kang J, Yang Q, Chen L. Genome-Wide Identification and Phylogenetic and Expression Analyses of the PLATZ Gene Family in Medicago sativa L. International Journal of Molecular Sciences. 2023; 24(3):2388. https://doi.org/10.3390/ijms24032388
Chicago/Turabian StyleLi, Xianyang, Fei He, Guoqing Zhao, Mingna Li, Ruicai Long, Junmei Kang, Qingchuan Yang, and Lin Chen. 2023. "Genome-Wide Identification and Phylogenetic and Expression Analyses of the PLATZ Gene Family in Medicago sativa L." International Journal of Molecular Sciences 24, no. 3: 2388. https://doi.org/10.3390/ijms24032388