
Sleeve Gastrectomy-Induced Weight Loss Increases Insulin Clearance in Obese Mice

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
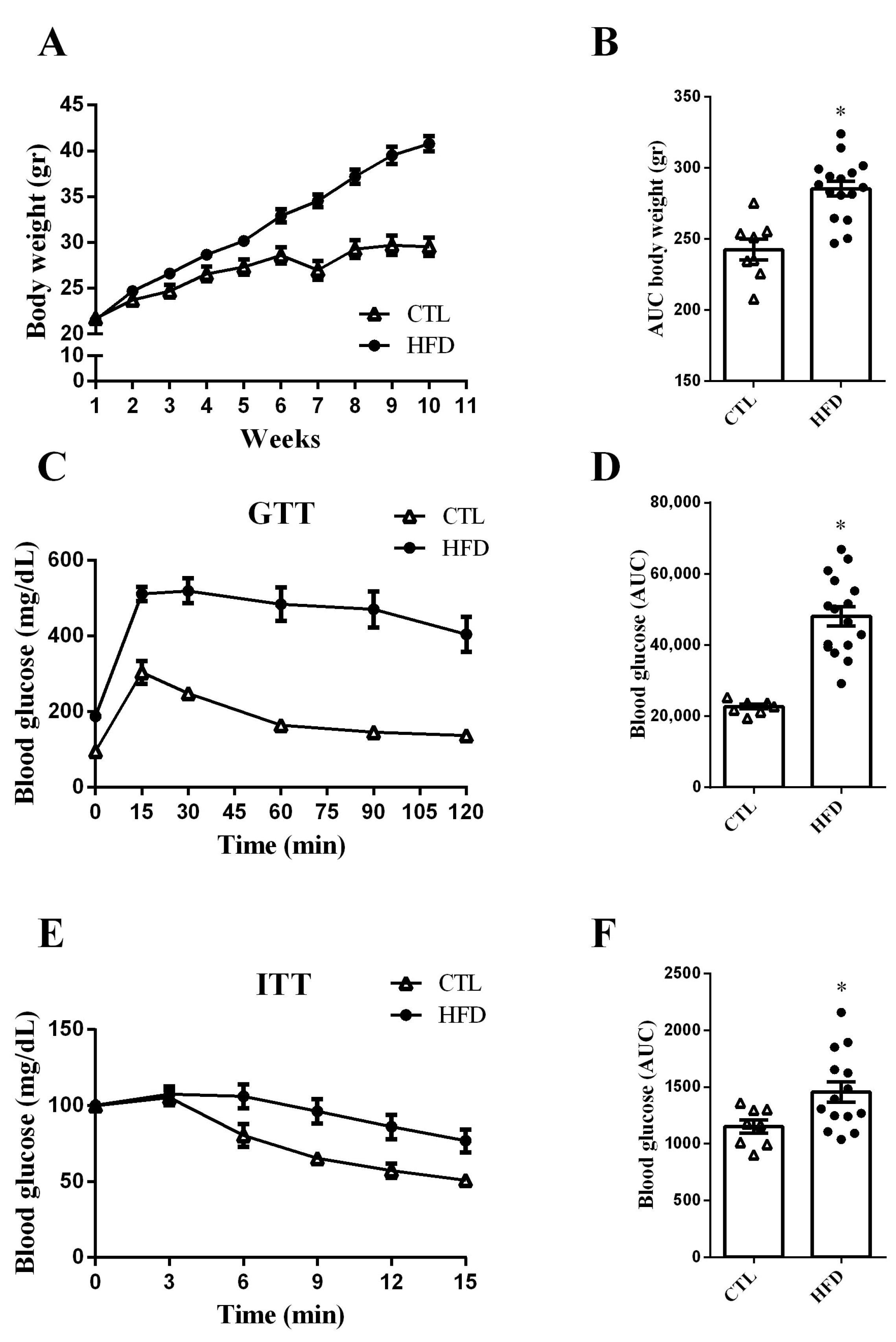
2.1. HFD Increases Body Weight Gain and Impairs Glucose Tolerance and Insulin Sensitivity in Mice
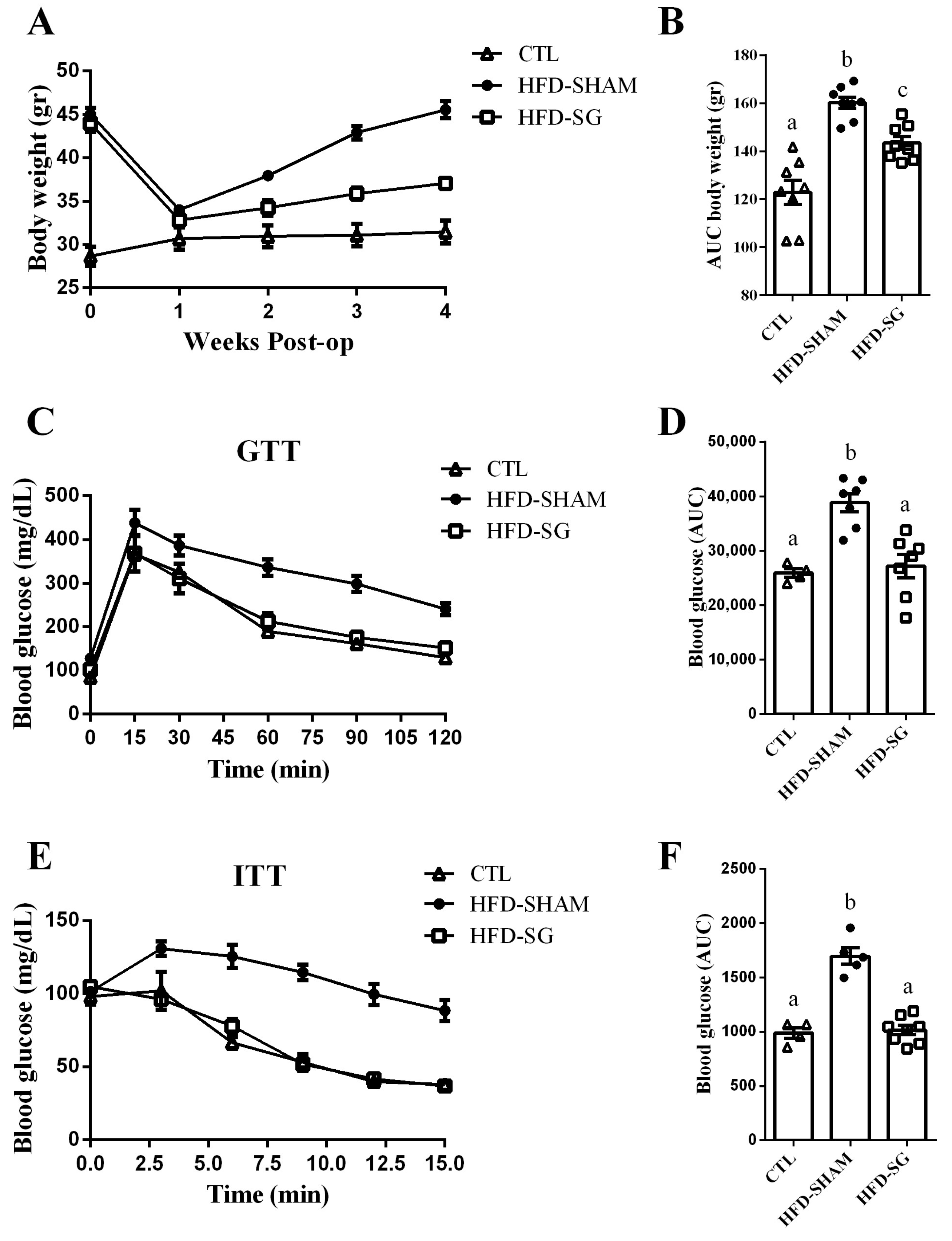
2.2. SG Reduces Body Weight Gain and Improves Glucose Tolerance and Insulin Sensitivity in HFD-SG Mice
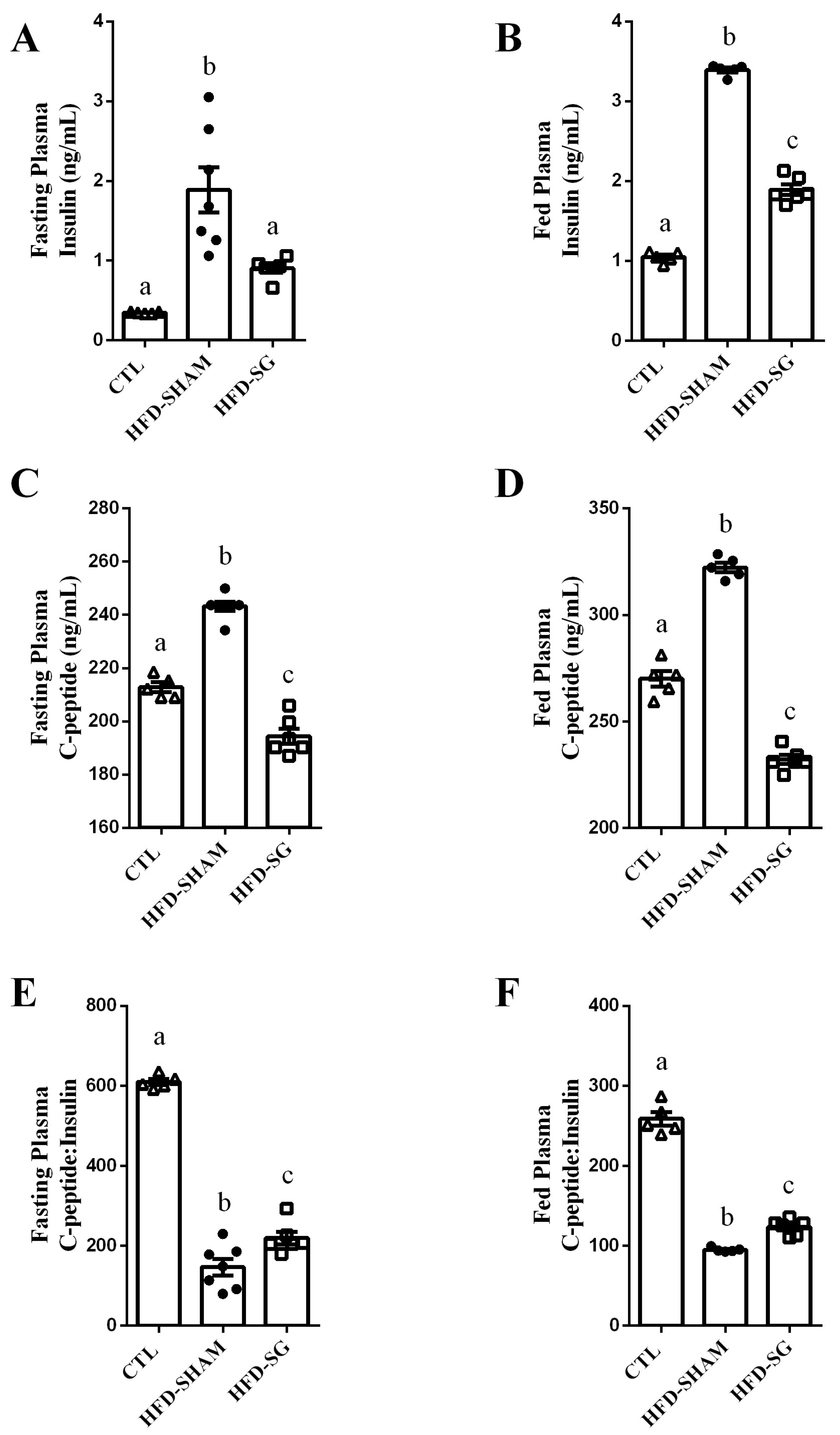
2.3. SG Reduces Fasting and Fed Insulin and C-Peptide Levels While Increasing Hepatic Insulin Clearance in HFD-SG Mice
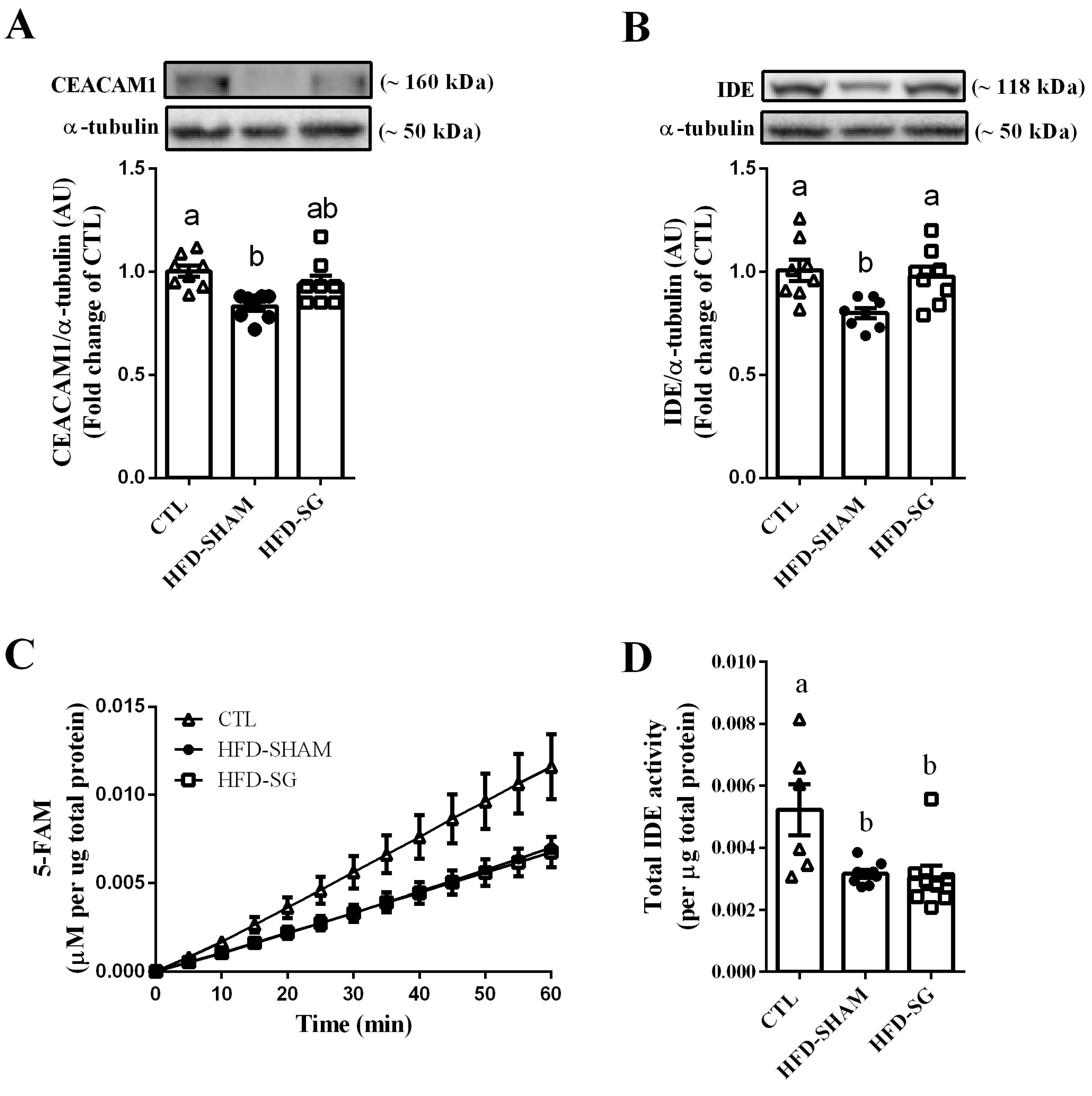
2.4. SG Increases Hepatic IDE Expression but Not IDE Activity in HFD-SG Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Sleeve Gastrectomy and Sham Operations
4.3. Intraperitoneal Glucose (ipGTT) and Insulin (ipITT) Tolerance Tests
4.4. Plasma Insulin and C-Peptide Measurements
4.5. Western Blot Analysis
4.6. IDE Activity Measurements
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrannini, E.; Mingrone, G. Impact of different bariatric surgical procedures on insulin action and beta-cell function in type 2 diabetes. Diabetes Care 2009, 32, 514–520. [Google Scholar] [CrossRef] [Green Version]
- Batterham, R.L.; Cummings, D.E. Mechanisms of Diabetes Improvement Following Bariatric/Metabolic Surgery. Diabetes Care 2016, 39, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfe, B.M.; Kvach, E.; Eckel, R.H. Treatment of Obesity: Weight Loss and Bariatric Surgery. Circ. Res. 2016, 118, 1844–1855. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, N.B.; Bojsen-Møller, K.N.; Dirksen, C.; Martinussen, C.; Svane, M.S.; Kristiansen, V.B.; Holst, J.J.; Madsbad, S. Sustained Improvements in Glucose Metabolism Late after Roux-En-Y Gastric Bypass Surgery in Patients with and Without Preoperative Diabetes. Sci. Rep. 2019, 9, 15154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, C.J.; Ingelfinger, J.R. Bariatric Surgery and Restoration of Insulin Sensitivity—It’s Weight Loss. N. Engl. J. Med. 2020, 383, 777–778. [Google Scholar] [CrossRef]
- Dirksen, C.; Jørgensen, N.B.; Bojsen-Møller, K.N.; Jacobsen, S.H.; Hansen, D.L.; Worm, D.; Holst, J.J.; Madsbad, S. Mechanisms of improved glycaemic control after Roux-en-Y gastric bypass. Diabetologia 2012, 55, 1890–1901. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, M.; Baldi, S.; Mari, A.; Colligiani, D.; Guarino, D.; Camastra, S.; Barsotti, E.; Berta, R.; Moriconi, D.; Bellini, R.; et al. Roux-en-Y gastric bypass and sleeve gastrectomy: Mechanisms of diabetes remission and role of gut hormones. J. Clin. Endocrinol. Metab. 2013, 98, 4391–4399. [Google Scholar] [CrossRef] [Green Version]
- Goldfine, A.B.; Patti, M.E. Diabetes improvement following Roux-en-Y gastric bypass: Understanding dynamic changes in insulin secretion and action. Diabetes 2014, 63, 1454–1456. [Google Scholar] [CrossRef] [Green Version]
- Bojsen-Møller, K.N. Mechanisms of improved glycaemic control after Roux-en-Y gastric bypass. Dan. Med. J. 2015, 62, B5057. [Google Scholar]
- Vaurs, C.; Brun, J.F.; Bérard, E.; Chalret du Rieu, M.; Hanaire, H.; Ritz, P. ß-cell pancreatic dysfunction plays a role in hyperglycemic peaks observed after gastric bypass surgery of obese patients. Surg. Obes. Relat. Dis. 2016, 12, 795–802. [Google Scholar] [CrossRef]
- Mullally, J.A.; Febres, G.J.; Bessler, M.; Korner, J. Sleeve Gastrectomy and Roux-en-Y Gastric Bypass Achieve Similar Early Improvements in Beta-cell Function in Obese Patients with Type 2 Diabetes. Sci. Rep. 2019, 9, 1880. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, W.C.; Bennett, R.G.; Hamel, F.G. Insulin degradation: Progress and potential. Endocr. Rev. 1998, 19, 608–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najjar, S.M.; Perdomo, G. Hepatic Insulin Clearance: Mechanism and Physiology. Physiology 2019, 34, 198–215. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Hay, S.O.; Kang, D.; McBride, M.; Li, L.; Zhao, J.; Leissring, M.A. Deletion of insulin-degrading enzyme elicits antipodal, age-dependent effects on glucose and insulin tolerance. PLoS ONE 2011, 6, e20818. [Google Scholar] [CrossRef] [Green Version]
- Kurauti, M.A.; Costa-Júnior, J.M.; Ferreira, S.M.; Dos Santos, G.J.; Protzek, A.O.; Nardelli, T.R.; de Rezende, L.F.; Boschero, A.C. Acute exercise restores insulin clearance in diet-induced obese mice. J. Endocrinol. 2016, 229, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Vettorazzi, J.F.; Kurauti, M.A.; Soares, G.M.; Borck, P.C.; Ferreira, S.M.; Branco, R.C.S.; Michelone, L.S.L.; Boschero, A.C.; Junior, J.M.C.; Carneiro, E.M. Bile acid TUDCA improves insulin clearance by increasing the expression of insulin-degrading enzyme in the liver of obese mice. Sci. Rep. 2017, 7, 14876. [Google Scholar] [CrossRef] [Green Version]
- Ding, L.; Sousa, K.M.; Jin, L.; Dong, B.; Kim, B.W.; Ramirez, R.; Xiao, Z.; Gu, Y.; Yang, Q.; Wang, J.; et al. Vertical sleeve gastrectomy activates GPBAR-1/TGR5 to sustain weight loss, improve fatty liver, and remit insulin resistance in mice. Hepatology 2016, 64, 760–773. [Google Scholar] [CrossRef] [Green Version]
- Douros, J.D.; Niu, J.; Sdao, S.; Gregg, T.; Fisher-Wellman, K.; Bharadwaj, M.; Molina, A.; Arumugam, R.; Martin, M.; Petretto, E.; et al. Sleeve gastrectomy rapidly enhances islet function independently of body weight. JCI Insight 2019, 4, e126688. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, S.N.; Harris, D.A.; Aliakbarian, H.; Luo, J.N.; Henke, M.T.; Subramaniam, R.; Vernon, A.H.; Tavakkoli, A.; Sheu, E.G.; Devlin, A.S. Bariatric surgery reveals a gut-restricted TGR5 agonist with anti-diabetic effects. Nat. Chem. Biol. 2021, 17, 20–29. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, E.; Yang, Q.; Jin, L.; Sousa, K.M.; Dong, B.; Wang, Y.; Tu, J.; Ma, X.; Tian, J.; et al. Vertical sleeve gastrectomy confers metabolic improvements by reducing intestinal bile acids and lipid absorption in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2019388118. [Google Scholar] [CrossRef]
- Templeman, N.M.; Flibotte, S.; Chik, J.H.L.; Sinha, S.; Lim, G.E.; Foster, L.J.; Nislow, C.; Johnson, J.D. Reduced Circulating Insulin Enhances Insulin Sensitivity in Old Mice and Extends Lifespan. Cell Rep. 2017, 20, 451–463. [Google Scholar] [CrossRef]
- Shanik, M.H.; Xu, Y.; Skrha, J.; Dankner, R.; Zick, Y.; Roth, J. Insulin resistance and hyperinsulinemia: Is hyperinsulinemia the cart or the horse? Diabetes Care 2008, 31 (Suppl. S2), S262–S268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalano, K.J.; Maddux, B.A.; Szary, J.; Youngren, J.F.; Goldfine, I.D.; Schaufele, F. Insulin resistance induced by hyperinsulinemia coincides with a persistent alteration at the insulin receptor tyrosine kinase domain. PLoS ONE 2014, 9, e108693. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Olefsky, J.M. Effect of experimental hyperinsulinemia on insulin binding and glucose transport in isolated rat adipocytes. Am. J. Physiol. 1978, 235, E53–E62. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.; Desai, K.S.; Steiner, G. Receptor and postreceptor insulin resistance induced by in vivo hyperinsulinemia. Can. J. Physiol. Pharmacol. 1983, 61, 802–807. [Google Scholar] [CrossRef]
- Piccinini, F.; Polidori, D.C.; Gower, B.A.; Fernandez, J.R.; Bergman, R.N. Dissection of hepatic versus extra-hepatic insulin clearance: Ethnic differences in childhood. Diabetes Obes. Metab. 2018, 20, 2869–2875. [Google Scholar] [CrossRef]
- Brancati, F.L.; Kao, W.H.; Folsom, A.R.; Watson, R.L.; Szklo, M. Incident type 2 diabetes mellitus in African American and white adults: The Atherosclerosis Risk in Communities Study. JAMA 2000, 283, 2253–2259. [Google Scholar] [CrossRef]
- Lee, C.C.; Haffner, S.M.; Wagenknecht, L.E.; Lorenzo, C.; Norris, J.M.; Bergman, R.N.; Stefanovski, D.; Anderson, A.M.; Rotter, J.I.; Goodarzi, M.O.; et al. Insulin clearance and the incidence of type 2 diabetes in Hispanics and African Americans: The IRAS Family Study. Diabetes Care 2013, 36, 901–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meistas, M.T.; Margolis, S.; Kowarski, A.A. Hyperinsulinemia of obesity is due to decreased clearance of insulin. Am. J. Physiol. 1983, 245, E155–E159. [Google Scholar] [CrossRef]
- Bronczek, G.A.; Vettorazzi, J.F.; Soares, G.M.; Kurauti, M.A.; Santos, C.; Bonfim, M.F.; Carneiro, E.M.; Balbo, S.L.; Boschero, A.C.; Costa Júnior, J.M. The Bile Acid TUDCA Improves Beta-Cell Mass and Reduces Insulin Degradation in Mice With Early-Stage of Type-1 Diabetes. Front. Physiol. 2019, 10, 561. [Google Scholar] [CrossRef]
- Leighton, E.; Sainsbury, C.A.; Jones, G.C. A Practical Review of C-Peptide Testing in Diabetes. Diabetes Ther. 2017, 8, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Kurauti, M.A.; Ferreira, S.M.; Soares, G.M.; Vettorazzi, J.F.; Carneiro, E.M.; Boschero, A.C.; Costa-Júnior, J.M. Hyperinsulinemia is associated with increasing insulin secretion but not with decreasing insulin clearance in an age-related metabolic dysfunction mice model. J. Cell. Physiol. 2019, 234, 9802–9809. [Google Scholar] [CrossRef] [PubMed]
- Lee, W. The CEACAM1 expression is decreased in the liver of severely obese patients with or without diabetes. Diagn. Pathol. 2011, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Ke, B.; Zhao, Z.; Ye, X.; Gao, Z.; Ye, J. Regulation of insulin degrading enzyme activity by obesity-associated factors and pioglitazone in liver of diet-induced obese mice. PLoS ONE 2014, 9, e95399. [Google Scholar] [CrossRef] [Green Version]
- Rezende, L.F.; Santos, G.J.; Santos-Silva, J.C.; Carneiro, E.M.; Boschero, A.C. Ciliary neurotrophic factor (CNTF) protects non-obese Swiss mice against type 2 diabetes by increasing beta cell mass and reducing insulin clearance. Diabetologia 2012, 55, 1495–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najjar, S.M.; Philippe, N.; Suzuki, Y.; Ignacio, G.A.; Formisano, P.; Accili, D.; Taylor, S.I. Insulin-stimulated phosphorylation of recombinant pp120/HA4, an endogenous substrate of the insulin receptor tyrosine kinase. Biochemistry 1995, 34, 9341–9349. [Google Scholar] [CrossRef]
- Ferraz-Bannitz, R.; Welendorf, C.R.; Coelho, P.O.; Salgado, W.; Nonino, C.B.; Beraldo, R.A.; Foss-Freitas, M.C. Bariatric surgery can acutely modulate ER-stress and inflammation on subcutaneous adipose tissue in non-diabetic patients with obesity. Diabetol. Metab. Syndr. 2021, 13, 19. [Google Scholar] [CrossRef]
- Billeter, A.T.; Scheurlen, K.M.; Israel, B.; Straub, B.K.; Schirmacher, P.; Kopf, S.; Nawroth, P.P.; Müller-Stich, B.P. Gastric Bypass Resolves Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Low-BMI Patients: A Prospective Cohort Study. Ann. Surg. 2022, 276, 814–821. [Google Scholar] [CrossRef]
- Li, H.; Wu, J.; Zhu, L.; Sha, L.; Yang, S.; Wei, J.; Ji, L.; Tang, X.; Mao, K.; Cao, L.; et al. Insulin degrading enzyme contributes to the pathology in a mixed model of Type 2 diabetes and Alzheimer’s disease: Possible mechanisms of IDE in T2D and AD. Biosci. Rep. 2018, 38, BSR20170862. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ding, Q.; Wang, Z. Nuclear respiratory factor 1 mediates the transcription initiation of insulin-degrading enzyme in a TATA box-binding protein-independent manner. PLoS ONE 2012, 7, e42035. [Google Scholar] [CrossRef] [Green Version]
- Nemati, R.; Lu, J.; Dokpuang, D.; Booth, M.; Plank, L.D.; Murphy, R. Increased Bile Acids and FGF19 After Sleeve Gastrectomy and Roux-en-Y Gastric Bypass Correlate with Improvement in Type 2 Diabetes in a Randomized Trial. Obes. Surg. 2018, 28, 2672–2686. [Google Scholar] [CrossRef] [PubMed]
- Guo, A.; Li, K.; Xiao, Q. Fibroblast growth factor 19 alleviates palmitic acid-induced mitochondrial dysfunction and oxidative stress via the AMPK/PGC-1α pathway in skeletal muscle. Biochem. Biophys. Res. Commun. 2020, 526, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Garibay, D.; Cummings, B.P. A Murine Model of Vertical Sleeve Gastrectomy. J. Vis. Exp. 2017, 130, 56534. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, G.M.; Lopes, L.E.S.; Balbo, S.L.; Marmentini, C.; Bronczek, G.A.; Kurauti, M.A.; Bonfleur, M.L.; Velloso, L.A.; Carneiro, E.M.; Boschero, A.C.; et al. Sleeve Gastrectomy-Induced Weight Loss Increases Insulin Clearance in Obese Mice. Int. J. Mol. Sci. 2023, 24, 1729. https://doi.org/10.3390/ijms24021729
Soares GM, Lopes LES, Balbo SL, Marmentini C, Bronczek GA, Kurauti MA, Bonfleur ML, Velloso LA, Carneiro EM, Boschero AC, et al. Sleeve Gastrectomy-Induced Weight Loss Increases Insulin Clearance in Obese Mice. International Journal of Molecular Sciences. 2023; 24(2):1729. https://doi.org/10.3390/ijms24021729
Chicago/Turabian StyleSoares, Gabriela Moreira, Luana Emanuelly Sinhori Lopes, Sandra Lucinei Balbo, Carine Marmentini, Gabriela Alves Bronczek, Mirian Ayumi Kurauti, Maria Lúcia Bonfleur, Licio Augusto Velloso, Everardo Magalhães Carneiro, Antonio Carlos Boschero, and et al. 2023. "Sleeve Gastrectomy-Induced Weight Loss Increases Insulin Clearance in Obese Mice" International Journal of Molecular Sciences 24, no. 2: 1729. https://doi.org/10.3390/ijms24021729