1. Introduction
Torpor is a fundamental metabolic state of energy conservation. Hibernation, defined by prolonged torpor lasting days to two or more weeks, interrupted by brief 12–24 h episodes of euthermic metabolism, represents some of the most extreme examples of mammalian energy conservation. Evidence suggests that hibernation is an extension of sleep that involves the modulation of thermogenesis [
1]. In ground squirrels, suppression of thermogenesis is sufficient to account for the initial fall in metabolic rate, after which a decline in core body temperature suppresses metabolic rate further through thermodynamic influence on metabolic processes [
2]. Adenosine, a ubiquitous neuromodulator present in most tissues, modulates thermogenesis in the CNS [
3,
4]. The purine binds to four subtypes of G-protein coupled receptors, designated the adenosine A
1, A
2A, A
2B and A
3 receptors (A
1AR, A
2aAR, A
2bAR, and A
3AR, respectively). In the brain, A
1AR and A
2AAR are widely expressed with only a small amount of A
3AR, whereas A
2BAR is only present in peripheral tissues. A
1AR and A
3AR receptors propagate their signal through G
i/o G-proteins and are neuroinhibitory. In contrast, the A
2AAR and A
2BAR interact with G
s proteins and are neuroexcitatory.
Stimulation of A
1AR within the CNS is necessary for the onset of torpor in hibernating Arctic Ground Squirrels (AGS;
Urocitellus parryii); however, the response is regulated by a process entrained to a circannual rhythm that governs seasonal sensitivity to the A
1AR agonist N
6-cyclohexyladenosine (CHA). Evidence from our laboratory suggests that the winter season enhances the influence of CHA on thermogenesis within the median preoptic nucleus and the rostral raphe pallidus to inhibit the premotor sympathetic neurons within the rPA that drive thermogenesis [
5]. The mechanism underlying seasonal sensitivity to CHA is unknown but may involve changes in adenosinergic tone within thermoregulatory nuclei in the hypothalamus or changes at the level of the A
1AR.
Sleep and thermogenesis are modulated, in part, by A
1AR [
6] and A
2AR [
7]. Hibernation, sleep and thermogenesis show seasonal rhythms in ground squirrels [
1,
8] and humans [
9], but it is unknown if the properties of A
1AR agonists or the expression of A
1AR vary with season. In the current study, we establish for the first-time pharmacological characteristics of the adenosine agonist CHA and antagonist DPCPX in AGS brain tissue. In addition, we test the hypothesis that a seasonal shift in the potency of CHA, the A
1AR agonist most studied regarding A
1AR agonist-induced hibernation and torpor, is associated with the seasonal expression of hibernation in AGS.
2. Results
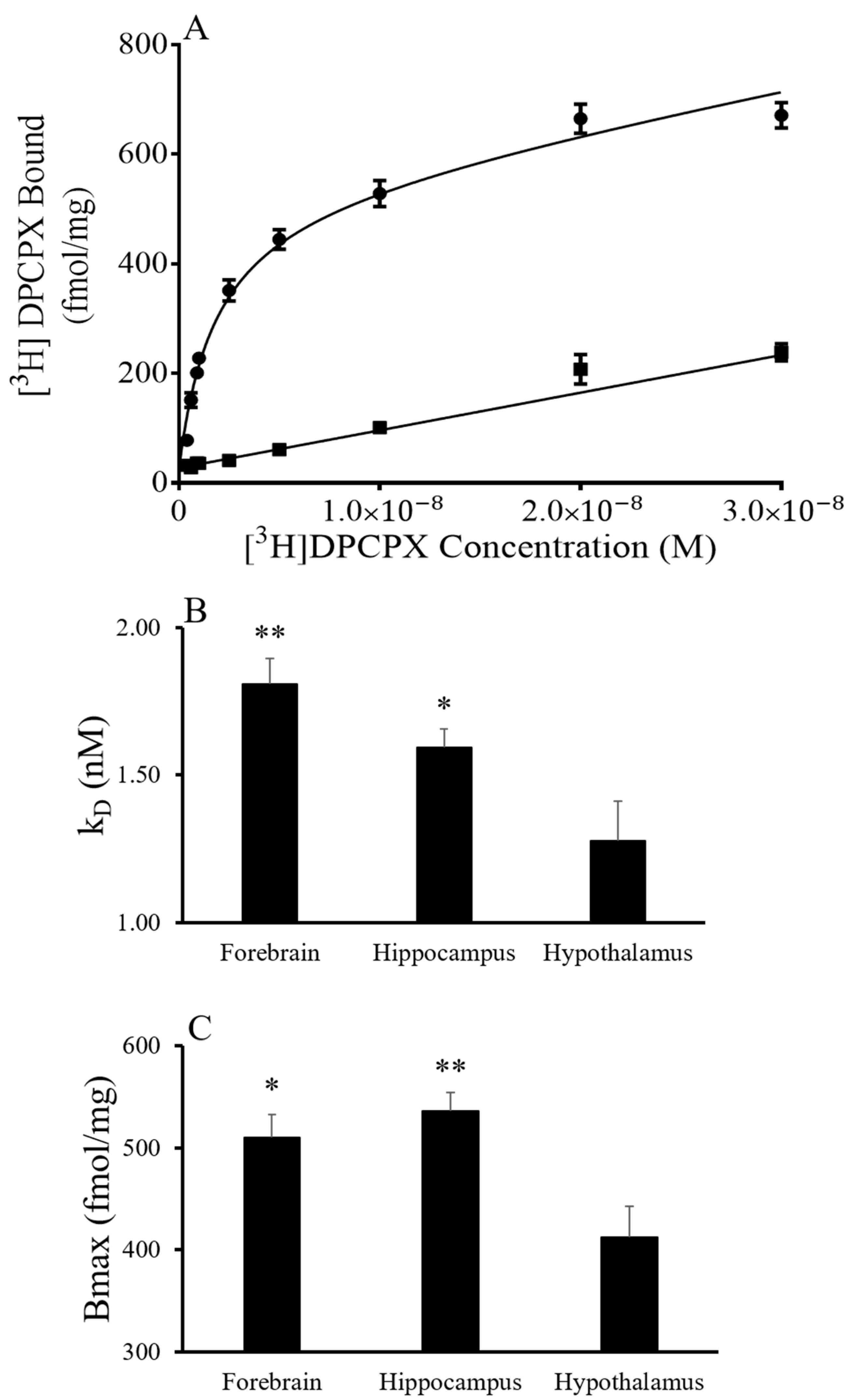
To characterize the A
1AR, saturation and displacement assays were conducted in the forebrain, hippocampus and hypothalamus of AGS euthanized during the summer and torpid state in the winter season. Saturation experiments using [
3H] DPCPX indicated a single site model and yielded K
D and Bmax values that showed an effect of brain region but not of the season (
Figure 1). The saturation curves for [
3H] DPCPX binding were similar in the forebrain, hypothalamus and hippocampus of tissue collected in summer and from torpid AGS in winter (
Figure 1). The results indicate that the hypothalamus has a lower K
D and Bmax than the forebrain and hippocampus independent of the season (
p = 0.0017, two-way ANOVA, the main effect of the region for K
D;
p = 0.0022, the main effect of the region for Bmax).
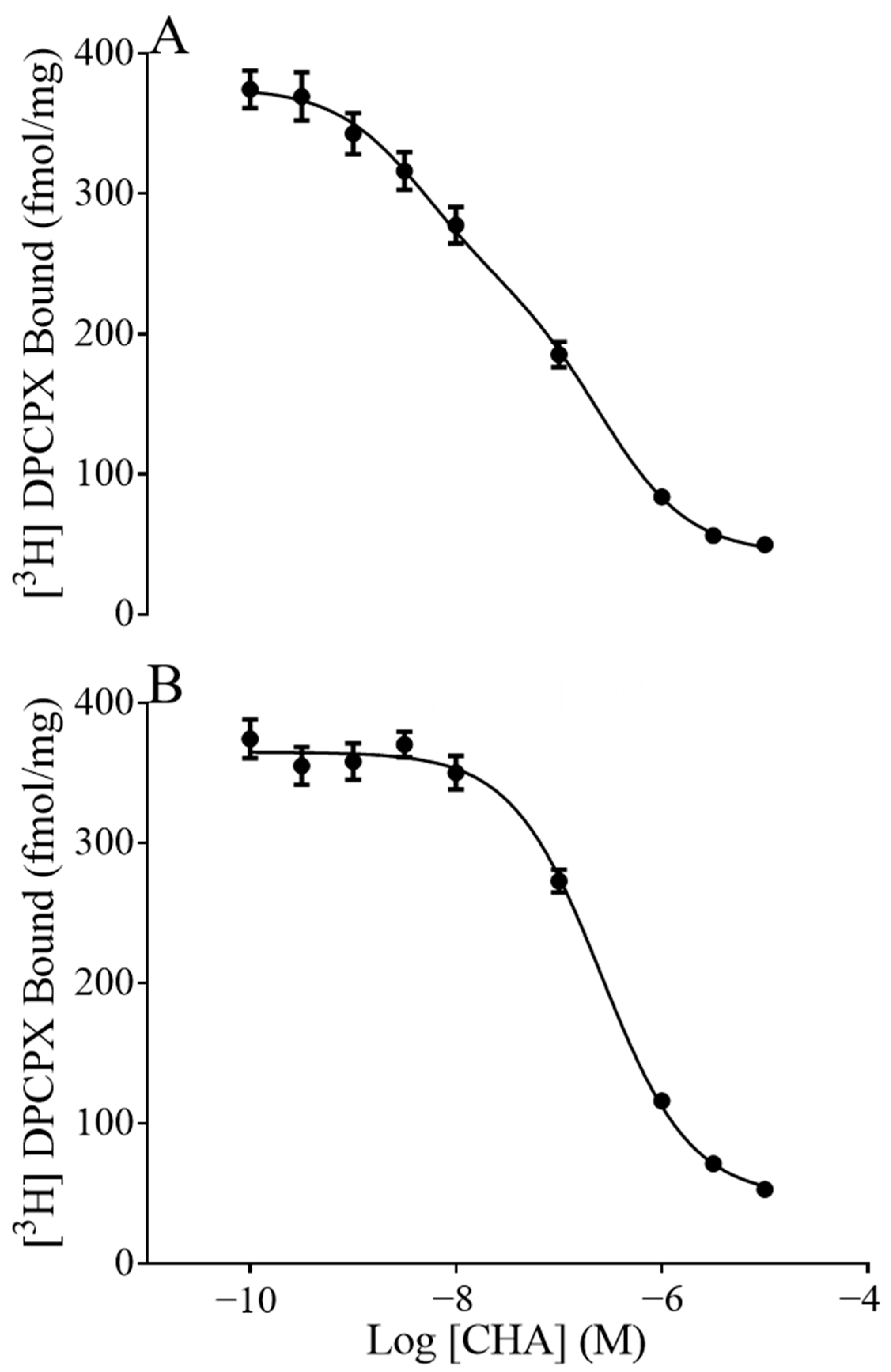
To ask if the fraction of receptors in the high or low affinity states was associated with seasonal sensitivity to CHA, we displaced [
3H]DPCPX with CHA. As expected, displacement assays fit a two-site model indicating the presence of a low and high affinity site (pki
Hi,Pki
Lo). Furthermore, in the presence of GTP, only the pki
Lo site was detected, as high concentrations of GTP promote the disassociation of G-proteins from GPCRs (
Figure 2). As expected, the pki
Lo established by the two-site model did not differ from the GTP shift pki
Lo, arguing that the GTP shift did indeed isolate the low affinity site. Here we found that the fraction of receptors in the high affinity state (Fraction High) was greater during torpor than during summer (
p = 0.0071, two-way ANOVA, main effect of season,
Table 1). In addition, there was an effect of region on the affinity of CHA at the low affinity site where the pki
Lo in the hippocampus was significantly less than the pki
Lo in the forebrain and hypothalamus, regardless of season (
p < 0.0001, two-way ANOVA, the main effect of region, with post-hoc Tukey
p < 0.0001 hippocampus vs. forebrain;
p < 0.0001 hippocampus vs. hypothalamus). The ratio of dissociation constants at the high and low affinity sites (pki
Hi/pki
Lo) was also greater in hippocamps than in other regions (
p < 0.001 two-way ANOVA, main effect of region). Post-hoc analysis showed that the hippocampus differed from the hypothalamus (
p < 0.001, Tukey) and trended towards being different from the forebrain (
p = 0.094).
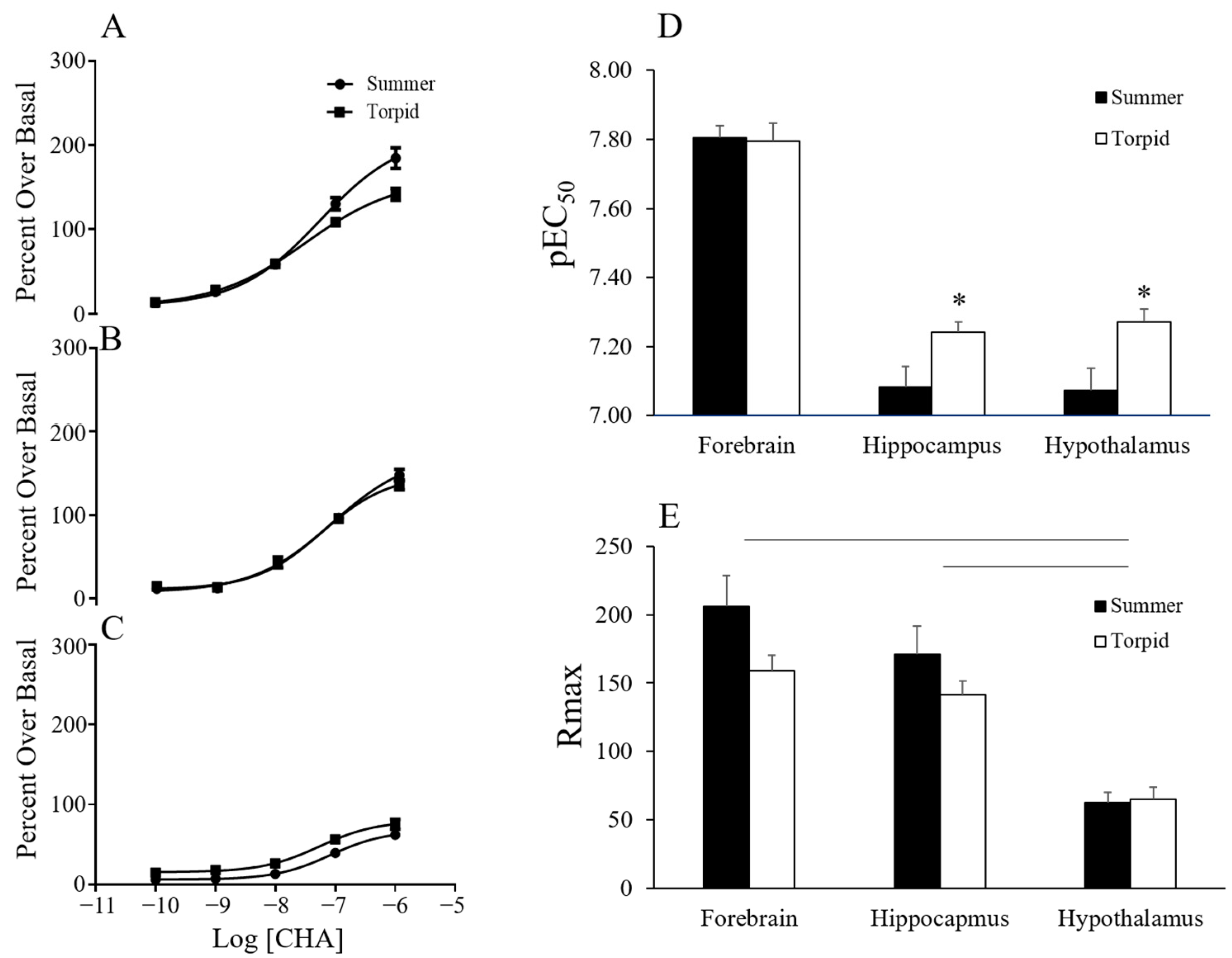
The functional response of CHA was investigated by agonist-induced
35S-GTPγS binding. Concentrations of CHA up to 1 µM stimulated
35S-GTPγS binding in a single-site manner. The resulting data revealed an effect of season and brain region on the pEC50 as well as an effect of region on the maximum signal (Rmax) (
Figure 3). CHA was more potent in tissue collected during torpor than during summer (
p < 0.005, two-way ANOVA, the main effect of the season). A trend towards an interaction between region and season (
p < 0.091) led us to perform further one-way analyses over the season that showed increased potency of CHA in torpor in the hypothalamus (
p < 0.05,
t-test) and in the hippocampus (
p < 0.05,
t-test) but not in the forebrain (
Figure 3). The Rmax was greatest in the forebrain and hippocampus and lowest in the hypothalamus (
p < 0.0001 two-way ANOVA, the main effect of region, with post-hoc Tukey
p < 0.0001 hippocampus vs. hypothalamus;
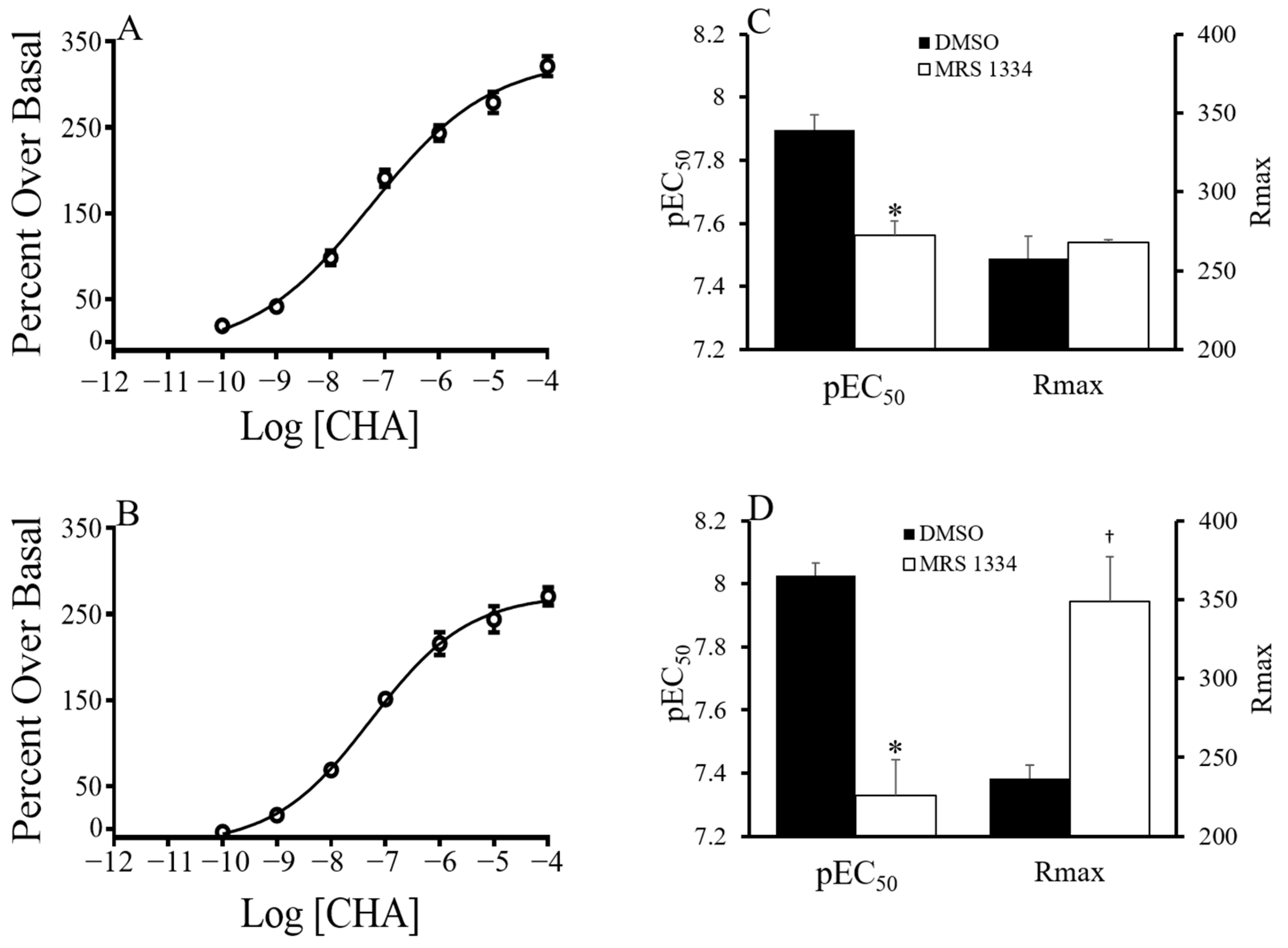
p < 0.0001 hypothalamus vs. forebrain). During the characterization of the functional response of CHA, a low hillslope was observed at concentrations greater than 1 µM. A low hillslope is indicative of negative cooperativity or a second lower affinity binding site. CHA has been shown to have a low affinity for the A
3AR in the rat brain [
10], and therefore A
3AR could be stimulated by high concentrations of CHA. We hypothesized that CHA at concentrations above 1 µM would stimulate the A
3AR resulting in a low hillslope. To eliminate A
3AR binding, we repeated the experiment in the presence of an A
3AR antagonist (MRS 1334; [
11]. In AGS, summer and torpid forebrain tissue, pre-blocked with 500 nM of MRS 1334, was stimulated with up to 100 µM CHA and
35S-GTPγS binding was measured (
Figure 4). The inclusion of MRS1334 did not have an effect on the hillslope of CHA-induced
35S-GTPγS binding in either season, providing evidence that the A
3AR stimulation was not contributing to the low hillslope. The potency of CHA, as indicated by the pEC
50, decreased in the presence of MRS1334, and the decrease was greater in tissue from torpid ASG than in tissue from summer AGS (
p < 0.0001, two-way ANOVA, the main effect of treatment;
p < 0.05 treatment x season). Interestingly, the Rmax only increased in the torpid tissue (
p < 0.05,
t-test torpid MRS1334 vs. vehicle).
It has been well established that GPCRs form dimers and higher order oligomers with GPCRs which can affect agonist signaling. If the effect of blocking A
3AR on Rmax or pEC
50 was due to A
1AR and A
3AR cross-talk, we asked if stimulating A
3AR would cause a change in Rmax or pEC
50. The A
3AR was stimulated with 132 nM 2-Chloro-N
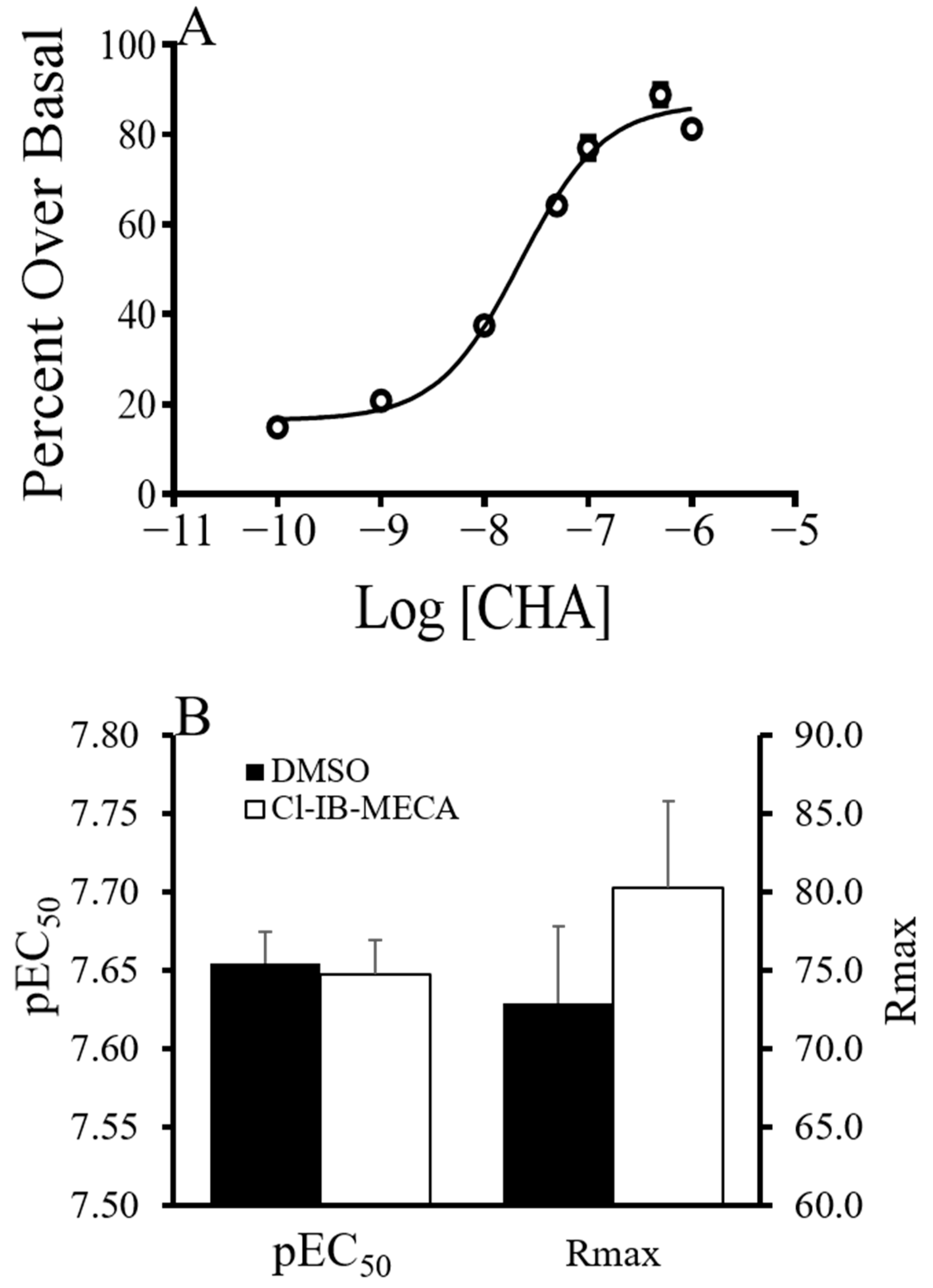
6-(3-iodobenzyl)-adenosine-5′-N-methyluronamide (Cl-IB–MECA). CHA-stimulated GDP/GTP exchange in the forebrain of summer AGS was then measured at concentrations up to 1.0 µM CHA using
35S-GTPγS binding assay (
Figure 5). There was no effect of Cl-IB-MECA on Rmax or pEC
50.
4. Materials and Methods
4.1. Arctic Ground Squirrels
AGS tissue was obtained from a tissue bank (courtesy of B. Barnes, Fairbanks, AK, USA). All animal procedures were approved by the UAF Institutional Animal Care and Use Committee (protocol #06-44). AGS were captured near 66°38′ N, 149°38′ W under permit from the Alaska Department of Fish & Game. Animals were housed at 22 °C on an 18:6 day: night cycle (5/2011–8/2011) and at 2 °C on a 4:20 day: night cycle (8/2011-time of tissue collection; 1/2011). Hibernation was monitored using the “shavings added” method, where hibernation (torpor) is indicated when shavings placed on the back of the AGS remain undisturbed 24 h later [
21,
22]. All tissue was harvested from adult male AGS during the summer season or while torpid during the winter season. The summer season was defined as AGS that were captured after the previous hibernation season and kept in captivity for two months before tissue collection. Tissue from torpid AGS were collected during the winter season, after at least six to eleven torpor bouts and at least ten but not more than thirteen days in the current torpor bout.
Summer AGS were euthanized by decapitation under a surgical plane of anesthesia (isoflurane, 4% mixed with 100% medical grade oxygen, delivered at 1.5 L/min until unresponsive to a toe pinch). Torpid AGS were euthanized without being aroused from torpor and did not require anesthesia nor breathe at a rate sufficient to absorb the gas anesthesia. Immediately following euthanasia, the brain was removed, and the hippocampus, hypothalamus and remaining forebrain were isolated and frozen in liquid nitrogen. All tissue was stored at −80 °C until use.
4.2. Isolation of Plasma Membrane for 35S GTPγS and [3H]DPCPX Binding Experiments
AGS membranes from the forebrain, hippocampus, hypothalamus and brainstem were isolated as described previously with modifications [
23]. Briefly, tissue was homogenized on ice using an all glass Dounce homogenizer (10–15 strokes) in 20× volume homogenization buffer containing 10 mM HEPES, 2 IU/mL ADA, 640 mM sucrose and protease inhibitor tablets (Roche, Indianapolis, IN, USA) and then further homogenized by polytron for 10–15 s. The suspension was centrifuged at 1000×
g for 10 min at 4 °C. Resultant supernatant was centrifuged at 48,000×
g for 15 min at 4 °C. Pellets were resuspended in Resuspension buffer containing 10 mM HEPES, 2 IU/mL ADA and protease inhibitor tablets. The suspension was centrifuged at 48,000×
g for 15 min at 4 °C. Pellets of AGS hippocampus and hypothalamus were suspended in a solution containing 6 mM HEPES, 122 µM GDP and 2.4 IU/mL ADA, the forebrain was suspended in 6 mM HEPES, 77 µM GDP, and 0.5 IU/mL ADA and both were incubated at room temperature under gentle rocking for 60 min and then centrifuged at 48,000×
g for 30 min. Subsequent pellets were resuspended in a Resuspension buffer and stored at −80 °C until use.
4.3. [3H] DPCPX Binding
To ask if membrane expression of the A1AR increased in the winter season, we performed saturation experiments to determine the K
D and Bmax of [
3H] DPCPX binding to A
1AR were conducted on the membrane of the forebrain, hippocampus and hypothalamus of winter and summer AGS following the guidelines of (Hulme 2010) with modification [
22]. On the day of the experiment, aliquots of summer and torpid AGS were thawed on ice. The protein content of each animal was determined by protein analysis (Bio-Rad, Hercules, CA, USA) followed by centrifugation at 48,000×
g for 30 min at 4 °C. The pellet was then resuspended in a solution containing 50 mM HEPES and 2 IU/mL ADA. Saturation experiments were performed by incubating 100 µg/mL protein with nine concentrations of [
3H] DPCPX ranging between 0.4 and 30 nM in the presence of 50 mM HEPES and 2 IU/mL ADA. Non-specific binding was defined in the presence of 7 µM cyclopentyltheophylline (CPT). The solution was allowed to incubate for 90 min at room temperature, and the membrane bound ligand was isolated as described below.
4.4. 35S-GTPγS Binding
To investigate the functional response of CHA activation of the A1AR in summer and torpid animals,
35S-GTPγS binding experiments were performed as described previously with modifications [
23]. On the day of the experiment, aliquots of summer and torpid AGS were thawed on ice. The protein content was then determined by protein analysis (Bio-Rad, Hercules, CA, USA) followed by centrifugation at 48,000×
g for 30 min at 4 °C. The pellet was then resuspended in Assay buffer containing 50 mM HEPES, 200 mM NaCl, 10 mM MgCl
2, 40 µM GDP, 100 µM Saponin, 1 IU/mL ADA and 1 mM DTT at pH 7.4. 100 µg per ml protein was incubated with 400 pM of
35S-GTPγS in a total volume of 100 µL for 90 min under gentle rocking at 37 °C. The non-specific activity was determined in the presence of 5 µM GTPγS. The constitutive activity was defined as binding in the absence of CHA. The reaction was terminated by rapid vacuum filtration, and then each well was washed three times with 200 µL of ice cold 50 mM HEPES. The plate was allowed to dry overnight. 40 µL of scintillation cocktail (PerkinElmer, Waltham, MA, USA) was added to each well, and
35S activity was determined in a 1450 Microbeta plus microplate scintillation counter (PerkinElmer, Waltham, MA, USA) utilizing a one-minute counting time. The effect of the A
3AR was determined by preincubating the membrane on ice with an A
3AR antagonist (MRS 1334, 500 nM) or agonist (Cl-IB-MECA, 132 nM) for at least one hour before conducting the
35S-GTPγS binding experiment.
To ask if the efficacy of CHA or the percentage of A1AR receptors in the high or low affinity state could explain the seasonal difference in response, KiLo was determined by displacing 1 nM [3H] DPCPX with nine concentrations of CHA ranging between 100 pM and 10 µM in the presence of 100 µg/mL protein, 50 mM HEPES, 2 IU/mL ADA and 1 mM GTP. KiHi was defined by displacing 1 nM [3H] DPCPX (PerkinElmer, Waltham, MA, USA) with CHA in the same manner as the KiLo experiments but without GTP. The solution was allowed to equilibrate for 90 min at room temperature, as indicated by kinetic experiments. Free and bound [3H] DPCPX was separated through an Inotech glass fiber filter pad (0.35 mM thickness/0.75 µM retention) (Inotech Bio. Sys., Derwood, MD, USA) by rapid filtration (0.5 mL per sec per well) with a cell harvester (Tomtec, Hamden, CT, USA). The filter was then allowed to dry overnight. The next morning each well was isolated and placed in a scintillation vial. Scintillation cocktail (PerkinElmer, Waltham, MA, USA) was added, and radioactivity was determined (1450 Microbeta Plus, PerkinElmer, Waltham, MA, USA) with a five-minute count per well. Unlabeled agonists and antagonists were obtained from Sigma (St. Louis, MO, USA).
4.5. Data Analysis
35S-GTPγS specific binding was determined by subtracting non-specific binding from overall binding. Specific binding was converted to percent over constitutive receptor activity. pEC50, Hill slope and span were determined using the function Log(agonist) vs. response—variable slope (four parameters) in Graphpad Prism 5 (v 5.04) (Graphpad Software, La Jolla, CA, USA).
[
3H] DPCPX bound was converted from cpm to fmol per mg protein, and the specific binding was calculated. A sum of squares F-test was used to determine if a one-site or two-site model was appropriate, and then the K
D and Bmax were calculated using the appropriate model using Graphpad Prism. Ki
Lo for the displacement of [
3H] DPCPX in the presence of GTP was calculated using the average K
D (one or two site—Fit Ki). Ki
Hi was calculated for the displacement of [
3H] DPCPX without GTP (one or two site—Fit Ki) using the average K
D and Ki
Lo. Fraction Hi is the fraction of all the sites that have a high affinity for the competitor. It is calculated by Graphpad prism via the below equations.
Data were analyzed by two-way ANOVA across brain region and season, followed by Tukey post-hoc comparisons or t-tests where indicated (R Studio). The significance threshold was defined as p < 0.05. Data are shown as mean ± SEM.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}