
Cervical Secretion Methylation Is Associated with the Pregnancy Outcome of Frozen-Thawed Embryo Transfer
 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
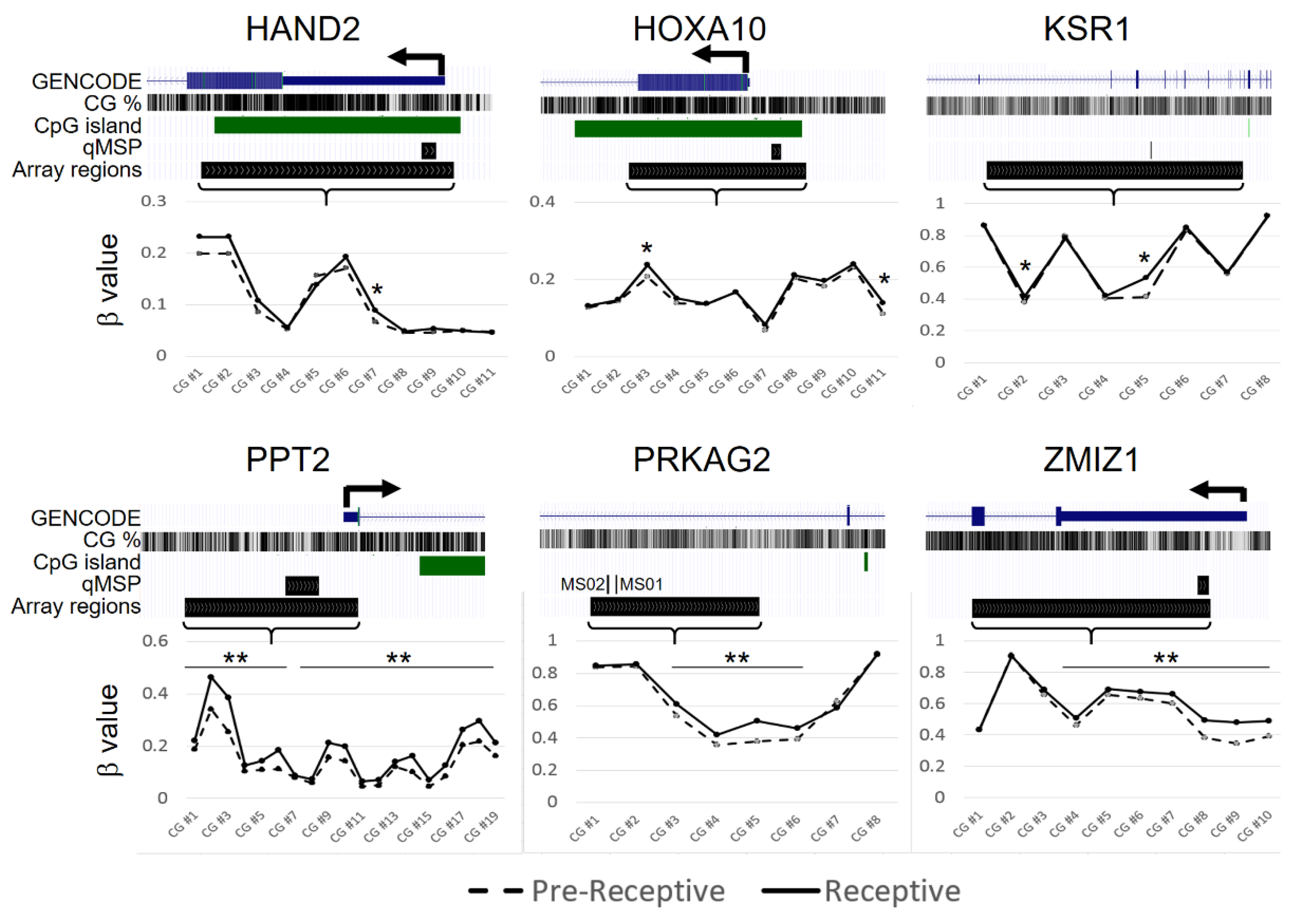
2.1. Candidate Gene Selection
2.2. CpG-Level Differential Methylation Analysis of Candidate Genes between Receptive Endometrium Compared to the Pre-Receptive Endometrium
2.3. DNA Methylation Analysis of Candiadate Genes via Methylation Array
2.4. Candidate Genes’ DNA Methylation Analysis via Quantitative Methylation-Specific Polymerase Chain Reaction (qMSP)
2.5. Machine Learning Approach for Differential Methylation Analysis of Candidate Genes
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Patients and Samples
4.3. DNA Extraction, Bisulphite Conversion, and DNA Methylation Measurement
4.4. Statistical Analyses and Machine Learning
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Geyter, C.; Wyns, C.; Calhaz-Jorge, C.; De Mouzon, J.; Ferraretti, A.P.; Kupka, M.; Andersen, A.N.; Nygren, K.G.; Goossens, V. 20 years of the European IVF-monitoring Consortium registry: What have we learned? A comparison with registries from two other regions. Hum. Reprod. 2020, 35, 2832–2849. [Google Scholar] [CrossRef]
- Munné, S.; Kaplan, B.; Frattarelli, J.L.; Child, T.; Nakhuda, G.; Shamma, F.N.; Silverberg, K.; Kalista, T.; Handyside, A.H.; Katz-Jaffe, M.; et al. Preimplantation genetic testing for aneuploidy versus morphology as selection criteria for single frozen-thawed embryo transfer in good-prognosis patients: A multicenter randomized clinical trial. Fertil. Steril. 2019, 112, 1071–1079.e7. [Google Scholar] [CrossRef]
- Yan, J.; Qin, Y.; Zhao, H.; Sun, Y.; Gong, F.; Li, R.; Sun, X.; Ling, X.; Li, H.; Hao, C.; et al. Live Birth with or without Preimplantation Genetic Testing for Aneuploidy. N. Engl. J. Med. 2021, 385, 2047–2058. [Google Scholar] [CrossRef]
- Norwitz, E.R.; Schust, D.J.; Fisher, S.J. Implantation and the survival of early pregnancy. N. Engl. J. Med. 2001, 345, 1400–1408. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Laufer, N. Repeated implantation failure: Clinical approach. Fertil. Steril. 2012, 97, 1039–1043. [Google Scholar] [CrossRef]
- Achache, H.; Revel, A. Endometrial receptivity markers, the journey to successful embryo implantation. Hum. Reprod. Updat. 2006, 12, 731–746. [Google Scholar] [CrossRef] [Green Version]
- Haouzi, D.; Dechaud, H.; Assou, S.; De Vos, J.; Hamamah, S. Insights into human endometrial receptivity from transcriptomic and proteomic data. Reprod. Biomed. Online 2012, 24, 23–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navot, D.; Scott, R.T.; Droesch, K.; Veeck, L.L.; Liu, H.-C.; Rosenwaks, Z. The window of embryo transfer and the efficiency of human conception in vitro. Fertil. Steril. 1991, 55, 114–118. [Google Scholar] [CrossRef]
- Lessey, B.A. Assessment of endometrial receptivity. Fertil. Steril. 2011, 96, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Gimeno, P.; Horcajadas, J.A.; Martinez-Conejero, J.A.; Esteban, F.J.; Alama, P.; Pellicer, A.; Simon, C. A genomic diagnostic tool for human endometrial receptivity based on the transcriptomic signature. Fertil. Steril. 2011, 95, 50–60.e15. [Google Scholar] [CrossRef]
- Sebastian-Leon, P.; Garrido, N.; Remohí, J.; Pellicer, A.; Gimeno, P.D. Asynchronous and pathological windows of implantation: Two causes of recurrent implantation failure. Hum. Reprod. 2018, 33, 626–635. [Google Scholar] [CrossRef]
- Caplakova, V.; Babusikova, E.; Blahovcova, E.; Balharek, T.; Zelieskova, M.; Hatok, J. DNA Methylation Machinery in the Endometrium and Endometrial Cancer. Anticancer. Res. 2016, 36, 4407–4420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houshdaran, S.; Zelenko, Z.; Irwin, J.C.; Giudice, L.C. Human endometrial DNA methylome is cycle-dependent and is associated with gene expression regulation. Mol. Endocrinol. 2014, 28, 1118–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, S.K.; Farquhar, C.; Mitchell, M.; Ponnampalam, A.P. Epigenetic regulation of endometrium during the menstrual cycle. Mol. Hum. Reprod. 2010, 16, 297–310. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef] [Green Version]
- Jirtle, R.L.; Skinner, M.K. Environmental epigenomics and disease susceptibility. Nat. Rev. Genet. 2007, 8, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Das, S.K. Epigenetic regulations through DNA methylation and hydroxymethylation: Clues for early pregnancy in decidualization. Biomol. Concepts 2014, 5, 95–107. [Google Scholar] [CrossRef]
- Kong, S.; Zhou, C.; Bao, H.; Ni, Z.; Liu, M.; He, B.; Huang, L.; Sun, Y.; Wang, H.; Lu, J. Epigenetic control of embryo-uterine crosstalk at peri-implantation. Cell. Mol. Life Sci. 2019, 76, 4813–4828. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Huang, X.; Mor, G.; Liao, A. Epigenetic modifications working in the decidualization and endometrial receptivity. Cell. Mol. Life Sci. 2019, 77, 2091–2101. [Google Scholar] [CrossRef]
- Lokk, K.; Modhukur, V.; Rajashekar, B.; Märtens, K.; Mägi, R.; Kolde, R.; Koltšina, M.; Nilsson, T.K.; Vilo, J.; Salumets, A.; et al. DNA methylome profiling of human tissues identifies global and tissue-specific methylation patterns. Genome Biol. 2014, 15, r54. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Bird, A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002, 16, 6–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saare, M.; Modhukur, V.; Suhorutshenko, M.; Rajashekar, B.; Rekker, K.; Sõritsa, D.; Karro, H.; Soplepmann, P.; Sõritsa, A.; Lindgren, C.M.; et al. The influence of menstrual cycle and endometriosis on endometrial methylome. Clin. Epigenetics 2016, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargett, C.E.; Schwab, K.E.; Deane, J.A. Endometrial stem/progenitor cells: The first 10 years. Hum. Reprod. Update 2016, 22, 137–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syed, S.M.; Tanwar, P.S. Axin2(+) endometrial stem cells: The source of endometrial regeneration and cancer. Mol. Cell. Oncol. 2020, 7, 1729681. [Google Scholar] [CrossRef] [PubMed]
- Jin, S. Bipotent stem cells support the cyclical regeneration of endometrial epithelium of the murine uterus. Proc. Natl. Acad. Sci. USA 2019, 116, 6848–6857. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.; Tan, S.; Buckett, W.; Dahan, M.H. Intra-patient variability in the endometrial receptivity assay (ERA) test. J. Assist. Reprod. Genet. 2018, 35, 929–930. [Google Scholar] [CrossRef] [PubMed]
- Hambartsoumian, E. Leukemia Inhibitory Factor (LIF) in Cervical Mucus of Infertile Women. Relationship with Endometrial LIF Secretion. Fertil. Steril. 1977, 1001, S18. [Google Scholar] [CrossRef]
- Gargiulo, A.R.; Fichorova, R.N.; Politch, J.A.; Hill, J.A.; Anderson, D.J. Detection of implantation-related cytokines in cervicovaginal secretions and peripheral blood of fertile women during ovulatory menstrual cycles. Fertil. Steril. 2004, 82 (Suppl. 3), 1226–1234. [Google Scholar] [CrossRef] [PubMed]
- Zanotta, N.; Monasta, L.; Skerk, K.; Luppi, S.; Martinelli, M.; Ricci, G.; Comar, M. Cervico-vaginal secretion cytokine profile: A non-invasive approach to study the endometrial receptivity in IVF cycles. Am. J. Reprod. Immunol. 2018, 81, e13064. [Google Scholar] [CrossRef]
- Huang, R.-L.; Su, P.-H.; Liao, Y.-P.; Wu, T.-I.; Hsu, Y.-T.; Lin, W.-Y.; Wang, H.-C.; Weng, Y.-C.; Ou, Y.-C.; Huang, T.H.-M.; et al. Integrated Epigenomics Analysis Reveals a DNA Methylation Panel for Endometrial Cancer Detection Using Cervical Scrapings. Clin. Cancer Res. 2017, 23, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, P.-L.; Huang, R.-L.; Wu, T.-I.; Liao, C.-C.; Chen, C.-W.; Su, P.-H.; Wang, H.-C.; Weng, Y.-C.; Lai, H.-C. Combined genetic mutations and DNA-methylated genes as biomarkers for endometrial cancer detection from cervical scrapings. Clin. Epigenetics 2019, 11, 170. [Google Scholar] [CrossRef]
- Li, F.; Zhang, M.; Zhang, Y.; Liu, T.; Qu, X. GnRH analogues may increase endometrial Hoxa10 promoter methylation and affect endometrial receptivity. Mol. Med. Rep. 2014, 11, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Kulp, J.L.; Mamillapalli, R.; Taylor, H.S. Aberrant HOXA10 Methylation in Patients With Common Gynecologic Disorders: Implications for Reproductive Outcomes. Reprod. Sci. 2016, 23, 455–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Tan, Y.J.; Wang, M.; Chen, Y.F.; Li, X.Y. DNA Methylation Inhibitor 5-Aza-2’-Deoxycytidine Modulates Endometrial Receptivity Through Upregulating HOXA10 Expression. Reprod. Sci. 2018, 26, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Nazarenko, T.A.; Kalinina, E.A.; Knyazeva, E.A.; Kiselev, V.I.; Smolnikova, V.Y.; Sukhikh, G.T. The role of abnormal hypermethylation of the HOXA10 and HOXA11 promoters in implantation failures in IVF programs. Gynecol. Endocrinol. 2019, 35 (Suppl. 1), 31–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Kannan, A.; DeMayo, F.J.; Lydon, J.P.; Cooke, P.S.; Yamagishi, H.; Srivastava, D.; Bagchi, M.K.; Bagchi, I.C. The antiproliferative action of progesterone in uterine epithelium is mediated by Hand2. Science 2011, 331, 912–916. [Google Scholar] [CrossRef] [Green Version]
- Bhurke, A.S.; Bagchi, I.C.; Bagchi, M.K. Progesterone-Regulated Endometrial Factors Controlling Implantation. Am. J. Reprod. Immunol. 2016, 75, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Šućurović, S.; Nikolić, T.; Brosens, J.J.; Mulac-Jeričević, B. Analysis of heart and neural crest derivatives-expressed protein 2 (HAND2)-progesterone interactions in peri-implantation endometriumdagger. Biol. Reprod. 2020, 102, 1111–1121. [Google Scholar] [CrossRef]
- Kukushkina, V.; Modhukur, V.; Suhorutšenko, M.; Peters, M.; Mägi, R.; Rahmioglu, N.; Velthut-Meikas, A.; Altmäe, S.; Esteban, F.J.; Vilo, J.; et al. DNA methylation changes in endometrium and correlation with gene expression during the transition from pre-receptive to receptive phase. Sci. Rep. 2017, 7, 3916. [Google Scholar] [CrossRef]
- Chen, C.-W.; Huang, R.-L.; Do, A.Q.; Wang, H.-C.; Lee, Y.-X.; Wang, C.-W.; Hsieh, C.-C.; Tzeng, C.-R.; Hu, Y.-M.; Chen, C.-H.; et al. Genome-wide analysis of cervical secretions obtained during embryo transfer reveals the association between deoxyribonucleic acid methylation and pregnancy outcomes. FS Sci. 2022, 3, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Reel, P.S.; Reel, S.; Pearson, E.; Trucco, E.; Jefferson, E. Using machine learning approaches for multi-omics data analysis: A review. Biotechnol. Adv. 2021, 49, 107739. [Google Scholar] [CrossRef]
- Glaab, E.; Rauschenberger, A.; Banzi, R.; Gerardi, C.; Garcia, P.; Demotes, J. Biomarker discovery studies for patient stratification using machine learning analysis of omics data: A scoping review. BMJ Open 2021, 11, e053674. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wang, J.; Liu, L.; Chen, X.; Xu, H.; Li, T.C.; Wang, C.C.; Zhang, S. Effects of high progesterone level on the day of human chorionic gonadotrophin administration in in vitro fertilization cycles on epigenetic modification of endometrium in the peri-implantation period. Fertil. Steril. 2017, 108, 269–276.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Y.; Hu, L.; Zhang, T.; Wang, M.; Xu, H.; Li, T.C.; Sun, Y.; Wang, C.C. Effects of high progesterone in in-vitro fertilization cycle on DNA methylation and gene expression of adhesion molecules on endometrium during implantation window. J. Assist. Reprod. Genet. 2019, 37, 33–43. [Google Scholar] [CrossRef]
- Enciso, M.; Aizpurua, J.; Rodríguez-Estrada, B.; Jurado, I.; Ferrández-Rives, M.; Rodríguez, E.; Pérez-Larrea, E.; Climent, A.B.; Marron, K.; Sarasa, J. The precise determination of the window of implantation significantly improves ART outcomes. Sci. Rep. 2021, 11, 13420. [Google Scholar] [CrossRef]
- Shi, C.; Shen, H.; Fan, L.-J.; Guan, J.; Zheng, X.-B.; Chen, X.; Liang, R.; Zhang, X.-W.; Cui, Q.-H.; Sun, K.-K.; et al. Endometrial MicroRNA Signature during the Window of Implantation Changed in Patients with Repeated Implantation Failure. Chin. Med. J. 2017, 130, 566–573. [Google Scholar] [CrossRef]
- Haouzi, D.; Entezami, F.; Torre, A.; Innocenti, C.; Antoine, Y.; Mauries, C.; Vincens, C.; Bringer-Deutsch, S.; Gala, A.; Ferrieres-Hoa, A.; et al. Customized Frozen Embryo Transfer after Identification of the Receptivity Window with a Transcriptomic Approach Improves the Implantation and Live Birth Rates in Patients with Repeated Implantation Failure. Reprod. Sci. 2020, 28, 69–78. [Google Scholar] [CrossRef]
- Lensen, S.; Lantsberg, D.; Gardner, D.K.; Sophian, A.D.; Wandafiana, N.; Kamath, M.S. The role of timing in frozen embryo transfer. Fertil. Steril. 2022, 118, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Doyle, N.; Jahandideh, S.; Hill, M.J.; Widra, E.A.; Levy, M.; Devine, K. A Randomized Controlled Trial Comparing Live Birth from Single Euploid Frozen Blastocyst Transfer Using Standardized Timing Versus Timing by Endometrial Receptivity Analysis. Fertil. Steril. 2021, 116, e101. [Google Scholar] [CrossRef]
- Cozzolino, M.; Diáz-Gimeno, P.; Pellicer, A.; Garrido, N. Use of the endometrial receptivity array to guide personalized embryo transfer after a failed transfer attempt was associated with a lower cumulative and per transfer live birth rate during donor and autologous cycles. Fertil. Steril. 2022, 118, 724–736. [Google Scholar] [CrossRef] [PubMed]
- Arian, S.E.; Hessami, K.; Khatibi, A.; To, A.K.; Shamshirsaz, A.A.; Gibbons, W. Endometrial Receptivity Array Before Frozen Embryo Transfer Cycles: A Systematic Review and Meta-analysis. Fertil. Steril. 2022, in press. [Google Scholar] [CrossRef] [PubMed]
- Alfer, J.; Popovici, R.M.; Fattahi, A.; Krieg, J.; Dittrich, R.; Beckmann, M.W.; Hartmann, A.; Bleisinger, N. Endometrial delay is found to be part of a normal individual dynamic transformation process. Arch. Gynecol. Obstet. 2021, 304, 1599–1609. [Google Scholar] [CrossRef]
- Seishima, R.; Leung, C.; Yada, S.; Murad, K.B.A.; Tan, L.T.; Hajamohideen, A.; Tan, S.H.; Itoh, H.; Murakami, K.; Ishida, Y.; et al. Neonatal Wnt-dependent Lgr5 positive stem cells are essential for uterine gland development. Nat. Commun. 2019, 10, 5378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vilella, F.; Alama, P.; Moreno, I.; Mignardi, M.; Isakova, A.; Pan, W.; Simon, C.; Quake, S.R. Single-cell transcriptomic atlas of the human endometrium during the menstrual cycle. Nat. Med. 2020, 26, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Bastu, E.; Demiral, I.; Gunel, T.; Ulgen, E.; Gumusoglu, E.; Hosseini, M.K.; Sezerman, U.; Buyru, F.; Yeh, J. Potential Marker Pathways in the Endometrium That May Cause Recurrent Implantation Failure. Reprod. Sci. 2018, 26, 879–890. [Google Scholar] [CrossRef]
- Dassen, H.; Punyadeera, C.; Kamps, R.; Klomp, J.; Dunselman, G.; Dijcks, F.; de Goeij, A.; Ederveen, A.; Groothuis, P. Progesterone regulation of implantation-related genes: New insights into the role of oestrogen. Cell. Mol. Life Sci. 2007, 64, 1009–1032. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schlenker, T.; Schoolcraft, W.B. Blastocyst score affects implantation and pregnancy outcome: Towards a single blastocyst transfer. Fertil. Steril. 2000, 73, 1155–1158. [Google Scholar] [CrossRef]
- Wu, T.-I.; Huang, R.-L.; Su, P.-H.; Mao, S.-P.; Wu, C.-H.; Lai, H.-C. Ovarian cancer detection by DNA methylation in cervical scrapings. Clin. Epigenetics 2019, 11, 166. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pregnant (P) | Non-Pregnant (nP) | p-Value | |
|---|---|---|---|
| Subject, n | 24 | 17 | |
| Age, years (range) | 35.81 ± 1.87 (32–39) | 35.97 ± 1.85 (32–38.8) | 0.558 |
| Infertility cause | 0.868 | ||
| Advanced Maternal Age | 2 (8.33) | 3 (21.43) | |
| Ovulatory | 6 (25) | 5 (29.41) | |
| Male factor | 6 (25) | 4 (23.53) | |
| Tubal factor | 2 (8.33) | 0 (0) | |
| Polycystic ovary syndrome | 0 (0) | 1 (5.88) | |
| Uterine | 1 (4.17) | 1 (5.88) | |
| Unexplained | 5 (20.83) | 2 (11.76) | |
| Endometriosis | 2 (8.33) | 1 (5.88) | |
| EM thickness (mm) during ET day | 9.57 ± 2.62 | 9.24 ±2.36 | 0.659 |
| Embryonic transfer number | 2.0 ± 0.44 | 1.87 ± 0.64 | 0.28 |
| Frozen embryonic transfer regimen | 0.748 | ||
| Hormone replacement therapy | 9 (66.7) | 8 (75) | |
| Nature cycle | 15 (33.33) | 9 (25) |
| Pregnant (P) | Non-Pregnant (nP) | p-Value | |
|---|---|---|---|
| Subject, n | 36 | 36 | |
| Age, years (range) | 37.93 ± 4.1 (31–46) | 37.57 ± 4.0 (29.6–43.6) | 0.558 |
| Infertility cause | 0.791 | ||
| Advanced Maternal Age | 12 (33.33) | 7 (19.44) | |
| Ovulatory | 4 (11.11) | 7 (19.44) | |
| Male factor | 4 (11.11) | 6 (16.67) | |
| Tubal factor | 0 (0) | 1 (2.78) | |
| Polycystic ovary syndrome | 2 (5.56) | 2 (5.56) | |
| Recurrent pregnancy loss | 0 (0) | 1 (2.78) | |
| Unexplained | 4 (11.11) | 4 (11.11) | |
| Endometriosis | 2 (5.56) | 1 (2.78) | |
| Multiple | 8 (22.22) | 7 (19.44) | |
| EM thickness (mm) during ET day | 9.48 ± 2.78 | 9.98 ± 2.2 | 0.659 |
| Embryonic transfer number | 2.0± 0.73 | 2.12 ± 0.83 | 0.28 |
| Frozen embryonic transfer regimen | 0.296 | ||
| Hormone replacement therapy | 24 (66.7) | 27 (75) | |
| Nature cycle | 12 (33.33) | 9 (25) |
| AUC | Accuracy | F_Measure | Precision | Recall | |
|---|---|---|---|---|---|
| Logistic Regression | 0.81 | 0.86667 | 0.88889 | 0.88889 | 0.88889 |
| Naive Bayes | 0.57 | 0.53333 | 0.53333 | 0.66667 | 0.44444 |
| Support Vector Machines | 0.81 | 0.73333 | 0.75 | 0.85714 | 0.66667 |
| K-Nearest Neighbor | 0.57 | 0.6 | 0.625 | 0.71429 | 0.55556 |
| Decision Trees | 0.76 | 0.66667 | 0.61538 | 1 | 0.44444 |
| Random Forest | 0.65 | 0.6 | 0.625 | 0.71429 | 0.55556 |
| Bootstrap-Aggregating Trees | 0.68 | 0.6 | 0.625 | 0.71429 | 0.55556 |
| Light GBM | 0.81 | 0.73333 | 0.71429 | 1 | 0.55556 |
| XG Boost | 0.83 | 0.8 | 0.8 | 1 | 0.66667 |
| Multilayer Perceptron | 0.89 | 0.73333 | 0.75 | 0.85714 | 0.66667 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.-X.; Su, P.-H.; Do, A.Q.; Tzeng, C.-R.; Hu, Y.-M.; Chen, C.-H.; Chen, C.-W.; Liao, C.-C.; Chen, L.-Y.; Weng, Y.-C.; et al. Cervical Secretion Methylation Is Associated with the Pregnancy Outcome of Frozen-Thawed Embryo Transfer. Int. J. Mol. Sci. 2023, 24, 1726. https://doi.org/10.3390/ijms24021726
Lee Y-X, Su P-H, Do AQ, Tzeng C-R, Hu Y-M, Chen C-H, Chen C-W, Liao C-C, Chen L-Y, Weng Y-C, et al. Cervical Secretion Methylation Is Associated with the Pregnancy Outcome of Frozen-Thawed Embryo Transfer. International Journal of Molecular Sciences. 2023; 24(2):1726. https://doi.org/10.3390/ijms24021726
Chicago/Turabian StyleLee, Yi-Xuan, Po-Hsuan Su, Anh Q. Do, Chii-Ruei Tzeng, Yu-Ming Hu, Chi-Huang Chen, Chien-Wen Chen, Chi-Chun Liao, Lin-Yu Chen, Yu-Chun Weng, and et al. 2023. "Cervical Secretion Methylation Is Associated with the Pregnancy Outcome of Frozen-Thawed Embryo Transfer" International Journal of Molecular Sciences 24, no. 2: 1726. https://doi.org/10.3390/ijms24021726