
Genetic Effects Analysis of QTLs for Rice Grain Size Based on CSSL-Z403 and Its Dissected Single and Dual-Segment Substitution Lines

Abstract
:1. Introduction
2. Results
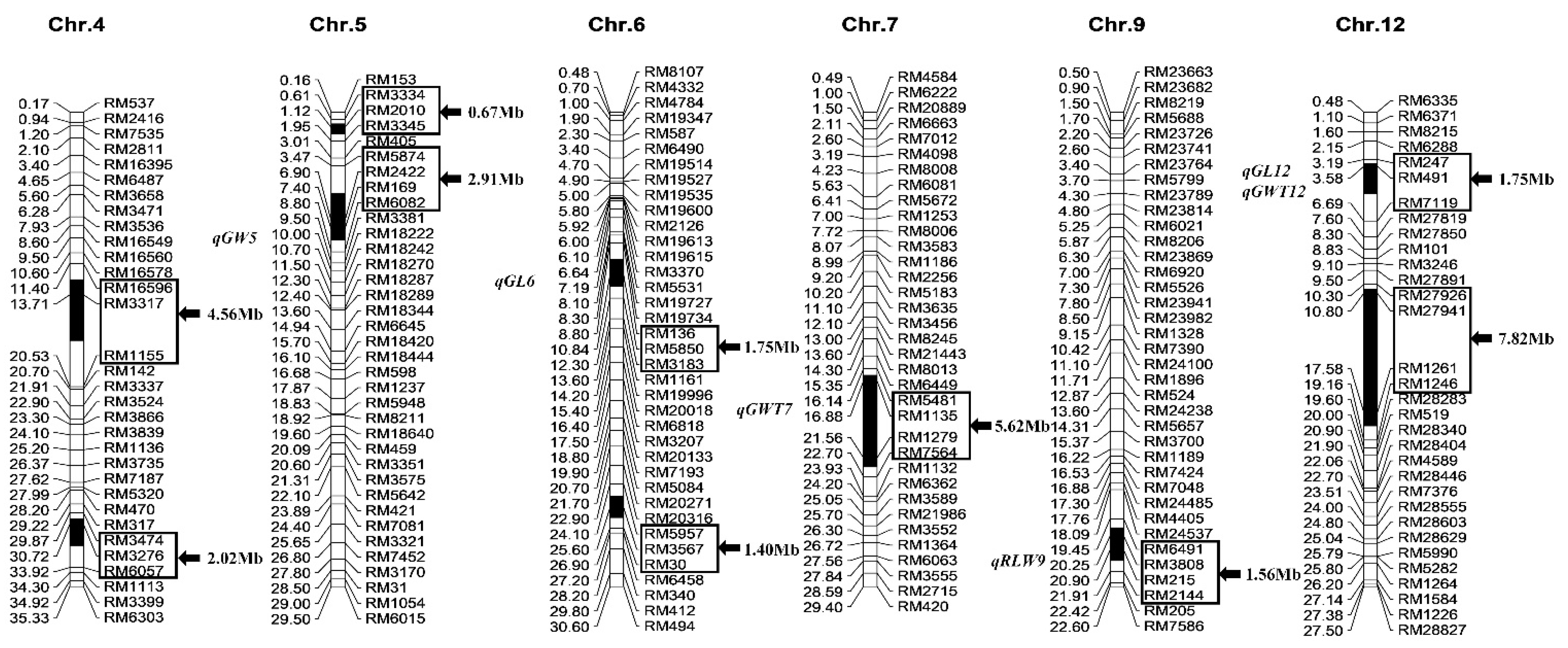
2.1. Identification of Substitution Segments in Z403
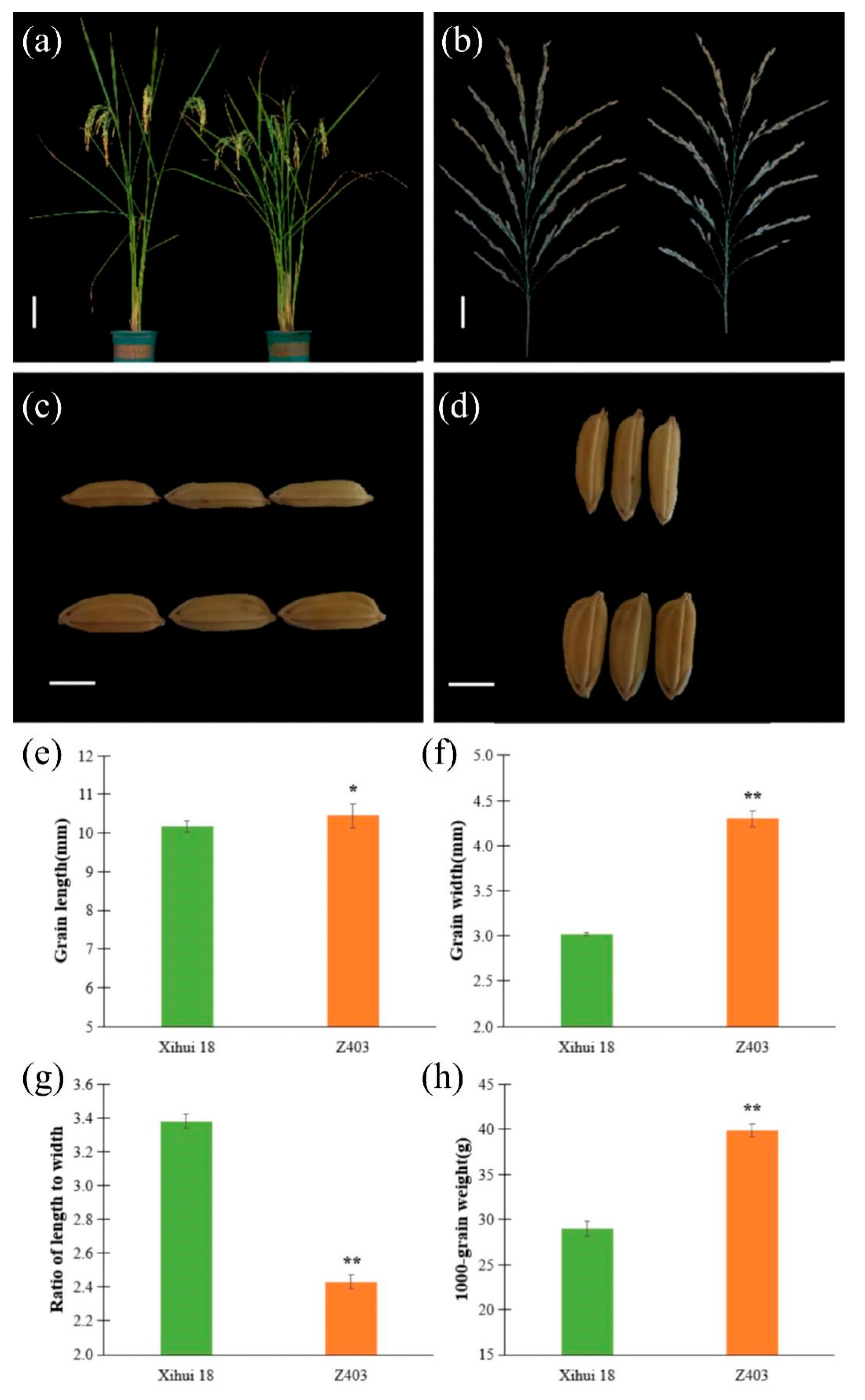
2.2. Analysis of Grain Size-Related Traits between Z403 and Xihui18
2.3. Identification of QTLs for Grain Size Using a Secondary F2 Population from Xihui18/Z403
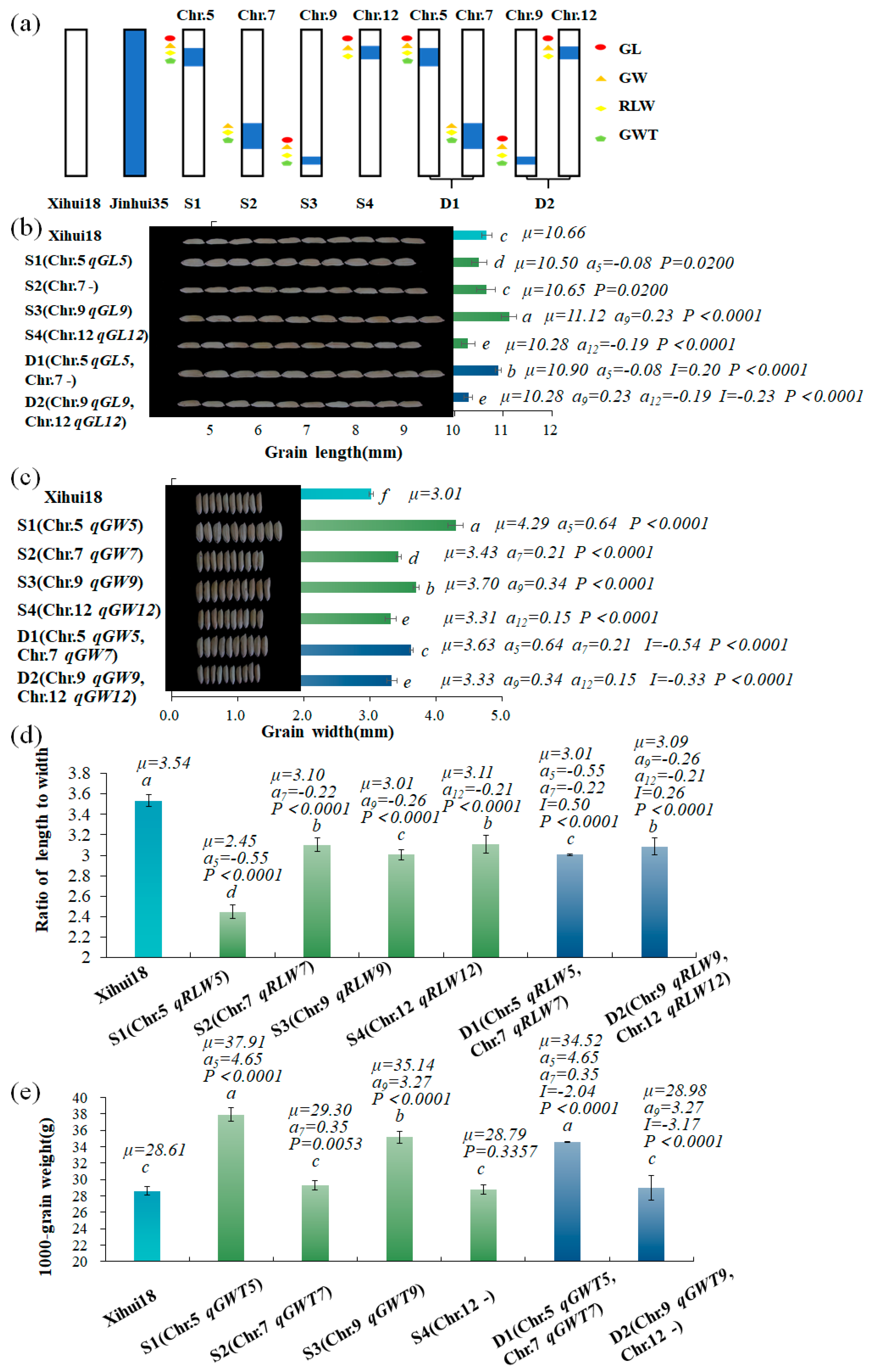
2.4. Verification and Analysis of Additive and Epistatic Effects of QTLs for Grain Size Using the Newly Developed SSSLs and DSSLs
2.4.1. Verification and Analysis of Additive Effects of QTLs for Grain Size by the SSSLs (S1–S4)
2.4.2. Analysis of Epistatic Effects of Two Non-allelic QTLs by DSSLs (D1–D2)
3. Discussion
3.1. The Developed SSSLs and DSSLs Dissected from Rice Large-Grain CSSL-Z403 Are Valuable Pre-Breeding Tools
3.2. qGL6, qGL7, qGW7, qGWT7, and qGL9 Might Be QTLs Newly Identified by Comparison with Previously Reported Genes
3.3. Identification of Epistatic Effects between Different Non-Allelic QTLs Is Essential for Breeding by Design
4. Materials and Methods
4.1. Experimental Materials
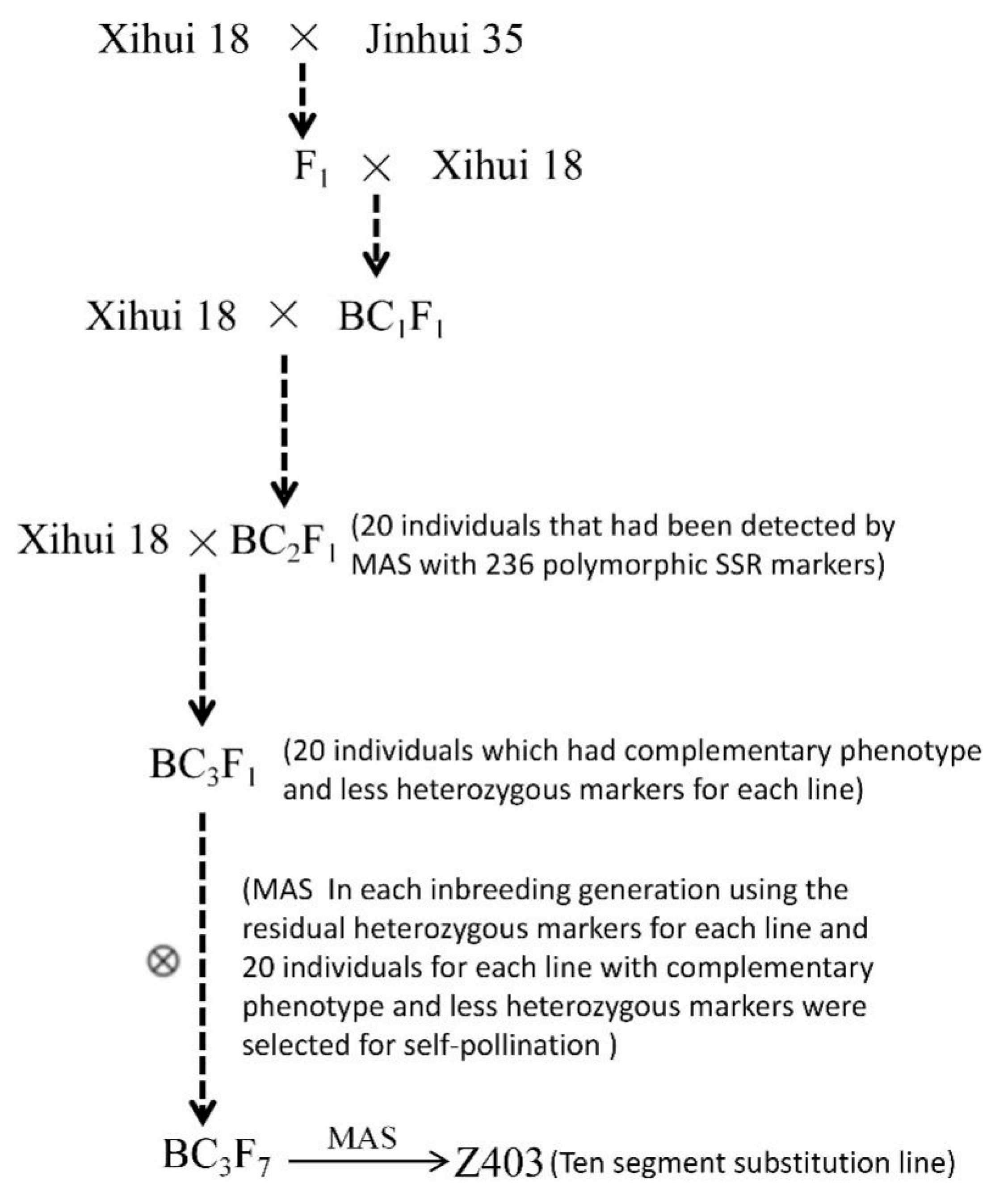
4.1.1. Development of Z403
4.1.2. Materials for QTL Mapping
4.2. Experimental Methods
4.2.1. Material Planting Methods
4.2.2. Assessment of Grain Size-Related Traits of All Materials
4.2.3. QTL Mapping
4.2.4. Development Method for Secondary SSSLs and DSSLs
4.3. Identification and Analysis of Additive Effect of QTLs for Grain Size by 4 SSSLs
4.4. Analysis of Epistatic Effect between Two Non-Allelic QTLs for Grain Size in the DSSLs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ray, D.; Mueller, N.; West, P.; Foley, J. Yield trends are insufficient to double global crop production by 2050. PLoS ONE 2013, 8, e66428. [Google Scholar]
- Vromant, N.; Duong, L.T.; Ollevier, F. Effect of fish on the yield and yield components of rice in integrated concurrent rice–fish systems. J. Agric. Sci. 2002, 138, 63–71. [Google Scholar] [CrossRef]
- Yoshida, S.; Ikegami, M.; Kuze, J.; Sawada, K.; Hashimoto, Z.; Ishii, T.; Nakamura, C.; Kamijima, O. QTL Analysis for Plant and Grain Characters of Sake-brewing Rice Using a Doubled Haploid Population. Breed. Sci. 2002, 52, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Rozbicki, J.; Ceglińska, A.; Gozdowski, D.; Jakubczak, M.; Cacak-Pietrzak, G.; Mądry, W.; Golba, J.; Piechociński, M.; Sobczyński, G.; Studnicki, M.; et al. Influence of the cultivar, environment and management on the grain yield and bread-making quality in winter wheat. J. Cereal Sci. 2015, 61, 126–132. [Google Scholar] [CrossRef]
- Balakrishnan, D.; Surapaneni, M.; Mesapogu, S.; Neelamraju, S. Development and use of chromosome segment substitution lines as a genetic resource for crop improvement. Theor. Appl. Genet. 2019, 132, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Wang, H.; Zhang, Q.; Zhou, K.; Li, M.; Li, R.; Xiang, S.; Zhang, T.; Ling, Y.; Yang, Z.; et al. Identification and Pyramiding of QTLs for Rice Grain Size Based on Short-Wide Grain CSSL-Z563 and Fine-Mapping of qGL3–2. Rice 2021, 14, 35. [Google Scholar] [CrossRef]
- Ali, M.L.; Sanchez, P.L.; Yu, S.; Lorieux, M.; Eizenga, G.C. Chromosome segment substitution lines: A powerful tool for the introgression of valuable genes from Oryza wild species into cultivated rice (O. sativa). Rice 2010, 3, 218–234. [Google Scholar]
- Shearman, J.R. A single segment substitution line population for identifying traits relevant to drought tolerance and avoidance. Genomics 2022, 114, 476–481. [Google Scholar] [CrossRef]
- Takai, T.; Nonoue, Y.; Yamamoto, S.; Yamanouchi, U.; Matsubara, K.; Liang, Z.W.; Lin, H.; Ono, N.; Uga, Y.; Yano, M. Development of chromosome segment substitution lines derived from backcross between indica donor rice cultivar ‘Nona Bokra’ and japonica recipient cultivar ‘Koshihikari’. Breed. Sci. 2007, 57, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Kanjoo, V.; Punyawaew, K.; Siangliw, J.L.; Jearakongman, S.; Vanavichit, A.; Toojinda, T. Evaluation of agronomic traits in chromosome segment substitution lines of KDML105 containing drought tolerance QTL under drought stress. Rice Sci. 2012, 19, 117–124. [Google Scholar] [CrossRef]
- Ebitani, T.; Takeuchi, Y.; Nonoue, Y.; Yamamoto, T.; Takeuchi, K.; Yano, M. Construction and Evaluation of Chromosome Segment Substitution Lines Carrying Overlapping Chromosome Segments of indica Rice Cultivar ‘Kasalath’ in a Genetic Background of japonica Elite Cultivar ‘Koshihikari’. Breed. Sci. 2005, 55, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wang, Z.; Xiang, S.; Lv, M.; Zhou, K.; Li, J.; Liang, P.; Li, M.; Li, R.; Ling, Y.; et al. Identification, pyramid, and candidate gene of QTL for yield-related traits based on rice CSSLs in indica Xihui18 background. Mol. Breed. 2022, 42, 19. [Google Scholar] [CrossRef]
- Li, J.; Yang, H.X.; Xu, G.Y.; Deng, K.L.; Yu, J.J.; Xiang, S.Q.; Zhou, K.; Zhang, Q.; Li, R.; Li, M.; et al. QTL analysis of Z414, a chromosome segment substitution line with short/wide grains, and substitution mapping of qGL11 in rice. Rice 2022, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Takeuchi, Y.; Miura, K.; Yamaguchi, T.; Ando, T.; Ebitani, T.; Higashitani, A.; Yamaya, T.; Yano, M.; Sato, T. Fine mapping of a major quantitative trait locus, qLG-9, that controls seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 769–778. [Google Scholar] [CrossRef]
- Wu, G.; Deng, H.; Yu, M.; Cai, Y.; Zhou, D.; Tan, J.; Yu, J.; Luo, X.; Tong, S.; Wang, P.; et al. Genetic analysis of rice seed recovery under low-temperature conditions using a new CSSL population with a high-density genetic map in rice. Mol. Breed. 2020, 40, 109. [Google Scholar] [CrossRef]
- Furuta, T.; Uehara, K.; Angeles-Shim, R.B.; Shim, J.; Ashikari, M.; Takashi, T. Development and evaluation of chromosome segment substitution lines (CSSLs) carrying chromosome segments derived from Oryza rufipogon in the genetic background of Oryza sativa L. Breed. Sci. 2014, 63, 468–475. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-Q.; Cui, G.-K.; Guan, C.-R.; Wang, J.; Liang, G.-H. QTL Detection for Rice Grain Shape Using Chromosome Single Segment Substitution Lines. Rice Sci. 2011, 18, 273–278. [Google Scholar] [CrossRef]
- Shim, R.A.; Angeles, E.R.; Ashikari, M.; Takashi, T. Development and evaluation of Oryza glaberrima Steud. chromosome segment substitution lines (CSSLs) in the background of O. sativa L. cv. Koshihikari. Breed. Sci. 2010, 60, 613–619. [Google Scholar] [CrossRef] [Green Version]
- Balakrishnan, D.; Surapaneni, M.; Yadavalli, V.R.; Addanki, K.R.; Mesapogu, S.; Beerelli, K.; Neelamraju, S. Detecting CSSLs and yield QTLs with additive, epistatic and QTL×environment interaction effects from Oryza sativa × O. nivara IRGC81832 cross. Sci. Rep. 2020, 10, 7766. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, P.K.; De Leon, T.; Singh, P.K.; Parco, A.; Cohn, M.A.; Sasaki, T. A Chromosome Segment Substitution Library of Weedy Rice for Genetic Dissection of Complex Agronomic and Domestication Traits. PLoS ONE 2015, 10, e0130650. [Google Scholar] [CrossRef] [Green Version]
- Bessho-Uehara, K.; Furuta, T.; Masuda, K.; Yamada, S.; Angeles-Shim, R.B.; Ashikari, M.; Takashi, T. Construction of rice chromosome segment substitution lines harboring Oryza barthii genome and evaluation of yield-related traits. Breed. Sci. 2017, 67, 408–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheema, K.K.; Bains, N.S.; Mangat, G.S.; Das, A.; Vikal, Y.; Brar, D.S.; Khush, G.S.; Singh, K. Development of high yielding IR64 × Oryza rufipogon (Griff.) introgression lines and identification of introgressed alien chromosome segments using SSR markers. Euphytica 2008, 160, 401–409. [Google Scholar] [CrossRef]
- Zhang, G. Target chromosome-segment substitution: A way to breeding by design in rice. Crop J. 2021, 9, 658–668. [Google Scholar] [CrossRef]
- Goswami, S.; Rao, V.S. Natural variation in crop plants: Its scope and utilization. J. Genet. 2021, 100, 29. [Google Scholar]
- Huang, Y.; Dong, H.; Mou, C.; Wang, P.; Hao, Q.; Zhang, M.; Wu, H.; Zhang, F.; Ma, T.; Miao, R.; et al. Ribonuclease H-like gene SMALL GRAIN2 regulates grain size in rice through brassinosteroid signaling pathway. J. Integr. Plant Biol. 2022, 64, 1883–1900. [Google Scholar] [CrossRef]
- Zhang, L.; Qi, Y.; Wu, M.; Zhao, L.; Zhao, Z.; Lei, C.; Hao, Y.; Yu, X.; Sun, Y.; Zhang, X.; et al. Mitochondrion-targeted PENTATRICOPEPTIDE REPEAT5 is required for cis-splicing of nad4 intron 3 and endosperm development in rice. Crop J. 2021, 9, 282–296. [Google Scholar] [CrossRef]
- Yuan, H.; Qin, P.; Hu, L.; Zhan, S.; Wang, S.; Gao, P.; Li, J.; Jin, M.; Xu, Z.; Gao, Q.; et al. OsSPL18 controls grain weight and grain number in rice. J. Genet. Genom. 2019, 46, 41–51. [Google Scholar] [CrossRef]
- Abu-Zaitoon, Y.M.; Bennett, K.; Normanly, J.; Nonhebel, H.M. A large increase in IAA during development of rice grains correlates with the expression of tryptophan aminotransferase OsTAR1 and a grain-specific YUCCA. Physiol. Plant. 2012, 146, 487–499. [Google Scholar] [CrossRef]
- Ando, T.; Yamamoto, T.; Shimizu, T.; Ma, X.F.; Shomura, A.; Takeuchi, Y.; Lin, S.Y.; Yano, M. Genetic dissection and pyramiding of quantitative traits for panicle architecture by using chromosomal segment substitution lines in rice. Theor. Appl. Genet. 2008, 116, 881–890. [Google Scholar] [CrossRef]
- Carlborg, O.; Haley, C.S. Epistasis: Too often neglected in complex trait studies? Nat. Rev. Genet. 2004, 5, 618–625. [Google Scholar] [CrossRef]
- Doust, A.N.; Lukens, L.; Olsen, K.M.; Mauro-Herrera, M.; Meyer, A.; Rogers, K. Beyond the single gene: How epistasis and gene-by-environment effects influence crop domestication. Proc. Nat. Acad. Sci. USA 2014, 11, 6178–6183. [Google Scholar]
- Leng, Y.J.; Wang, S.L.; Wang, R.A.; Tao, T.; Jia, S.W.; Song, T.; Xu, L.N.; Cai, X.L.; Jin, S.K.; Gao, J.P. Multi-Environmental Genetic Analysis of Grain Size Traits Based on Chromosome Segment Substitution Line in Rice (Oryza sativa L.). Phyton 2022, 91, 943–958. [Google Scholar] [CrossRef]
- Zou, T.; Zhao, H.W.; Li, X.H.; Zheng, M.M.; Zhang, S.D.; Sun, L.L.; He, L.; Pan, X.P.; Liu, Z.Q.; Fu, X.L. QTLs detection and pyramiding for stigma exsertion rate in wild rice species by using the single-segment substitution lines. Mol. Breed. 2020, 40, 74. [Google Scholar] [CrossRef]
- Paterson, A.H.; Damon, S.; Hewitt, J.D.; Zamir, D.; Rabinowitch, H.D.; Lincoln, S.E.; Lander, E.S.; Tanksley, S.D. Mendelian factors underlying quantitative traits in tomato: Comparison across species, generations, and environments. Genetics 1991, 127, 181–197. [Google Scholar] [CrossRef] [PubMed]
- McCouch, S.R.; Kochert, G.; Yu, Z.H.; Wang, Z.Y.; Khush, G.S.; Coffman, W.R.; Tanksley, S.D. Molecular mapping of rice chromosomes. Theor. Appl. Genet. 1988, 76, 815–829. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.M.; Tan, Y.; Zheng, L.Y.; Zhou, K.; He, G.H.; Ling, Y.H.; Zhang, L.H.; Xu, S.Z. Identification of rice chromosome segment substitution line Z322-1-10 and mapping QTLs for agronomic traits from the F3population. Cereal Res. Commun. 2016, 44, 370–380. [Google Scholar] [CrossRef] [Green Version]
- Paterson, A.H.; Brubaker, C.L.; Wendel, J.F. A rapid method for extraction of cotton (gossypium spp.) genomic DNA suitable for RFLP or PCR analysis. Plant Mol. Biol. Rep. 1993, 11, 122–127. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | QTL | Chr. | Linked Marker | Additive Effect | Var. (%) | p-Value |
|---|---|---|---|---|---|---|
| Grain length (mm) | qGL6 | 6 | RM5850 | 0.08 | 8.54 | 0.0469 |
| qGL12 | 12 | RM491 | 0.11 | 16.10 | 0.0137 | |
| Grain width (mm) | qGW5 | 5 | RM169 | 0.09 | 19.15 | 0.0152 |
| Ratio of length to width | qRLW9 | 9 | RM3808 | 0.07 | 8.02 | 0.0395 |
| 1000-grain weight (g) | qGWT7 | 7 | RM1135 | 3.99 | 19.49 | 0.0351 |
| qGWT12 | 12 | RM491 | 2.76 | 9.34 | 0.0459 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, G.; Deng, K.; Yu, J.; Li, Q.; Li, L.; Xiang, A.; Ling, Y.; Zhang, C.; Zhao, F. Genetic Effects Analysis of QTLs for Rice Grain Size Based on CSSL-Z403 and Its Dissected Single and Dual-Segment Substitution Lines. Int. J. Mol. Sci. 2023, 24, 12013. https://doi.org/10.3390/ijms241512013
Xu G, Deng K, Yu J, Li Q, Li L, Xiang A, Ling Y, Zhang C, Zhao F. Genetic Effects Analysis of QTLs for Rice Grain Size Based on CSSL-Z403 and Its Dissected Single and Dual-Segment Substitution Lines. International Journal of Molecular Sciences. 2023; 24(15):12013. https://doi.org/10.3390/ijms241512013
Chicago/Turabian StyleXu, Guangyi, Keli Deng, Jinjin Yu, Qiaolong Li, Lu Li, Aoni Xiang, Yinghua Ling, Changwei Zhang, and Fangming Zhao. 2023. "Genetic Effects Analysis of QTLs for Rice Grain Size Based on CSSL-Z403 and Its Dissected Single and Dual-Segment Substitution Lines" International Journal of Molecular Sciences 24, no. 15: 12013. https://doi.org/10.3390/ijms241512013