
Population Structure and Genetic Diversity of Shanlan Landrace Rice for GWAS of Cooking and Eating Quality Traits

Abstract
:1. Introduction
2. Results
2.1. Identification of SNPs and InDels in Shanlan Landrace Rice by Next-Generation Sequencing
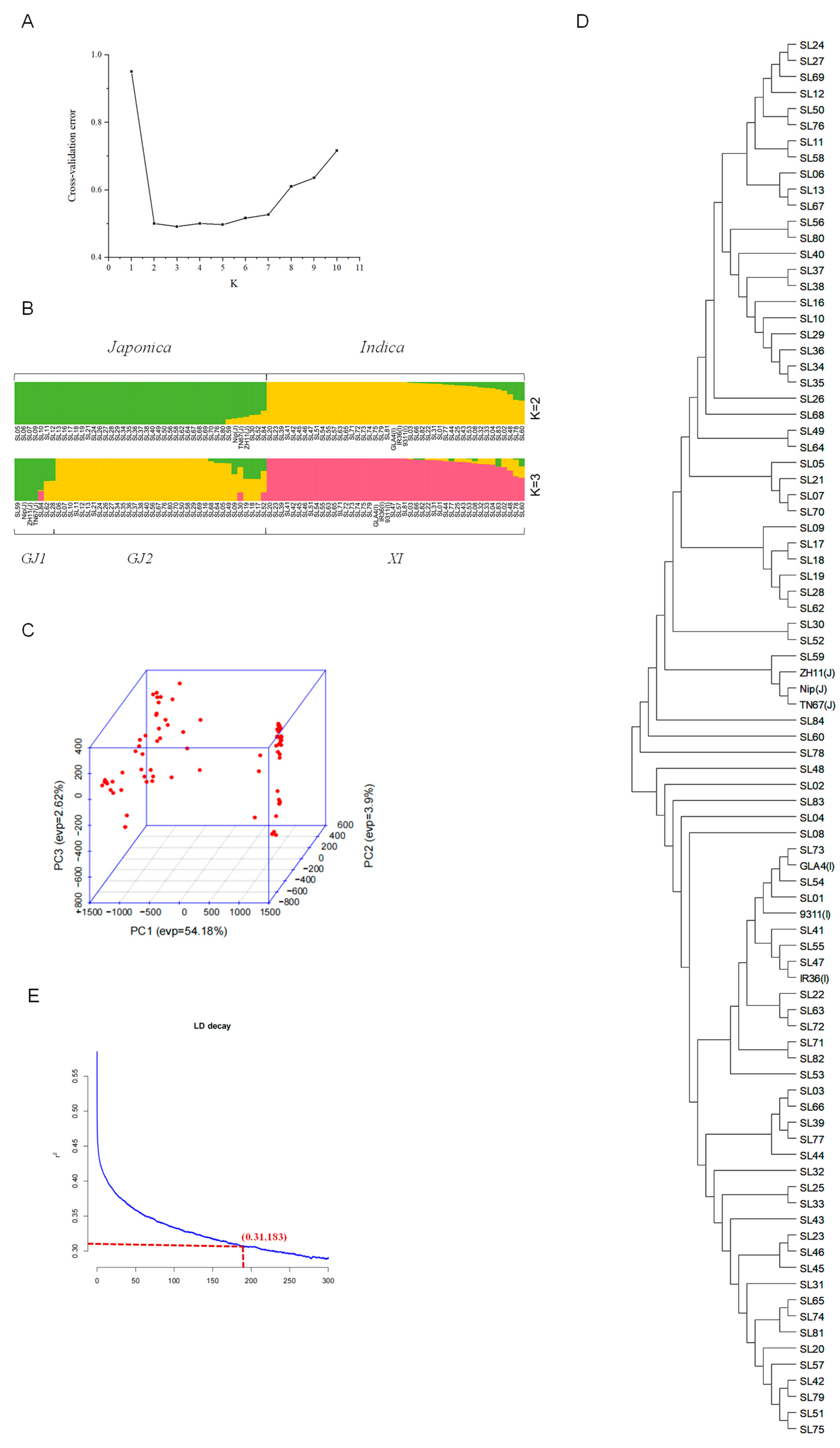
2.2. Genetic Population Structure
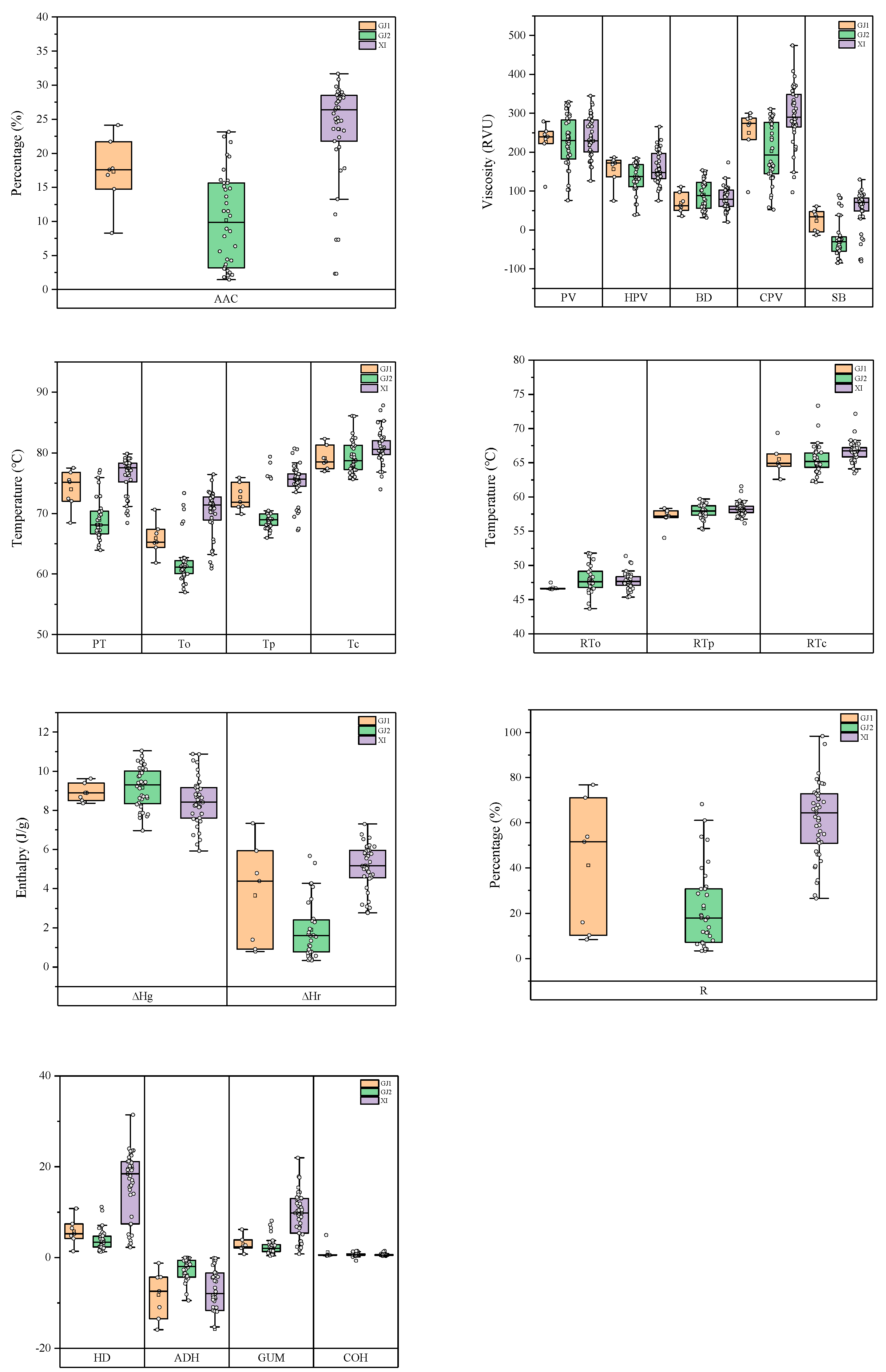
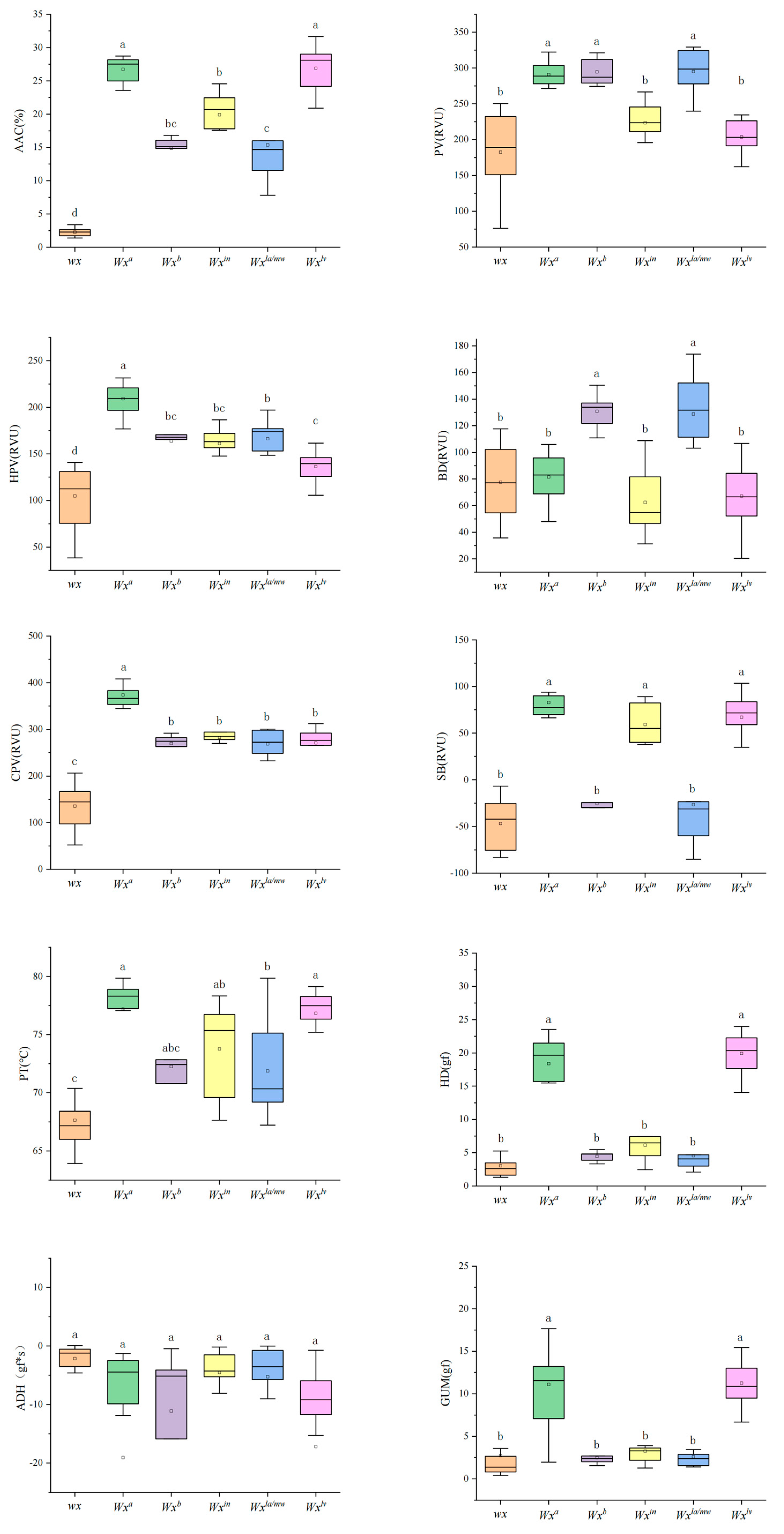
2.3. Phenotypic Variation in the Study Population
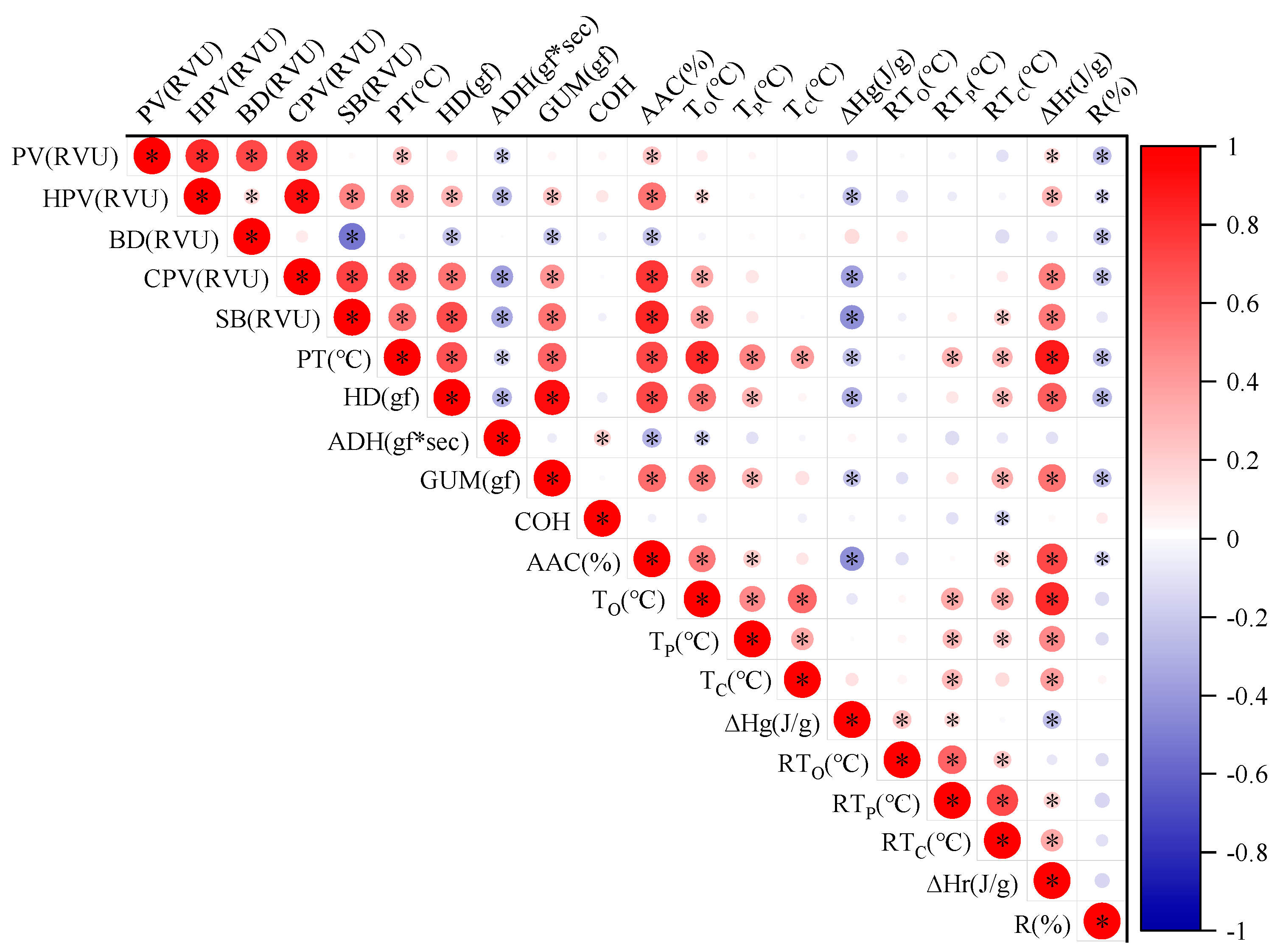
2.4. Correlation Analysis
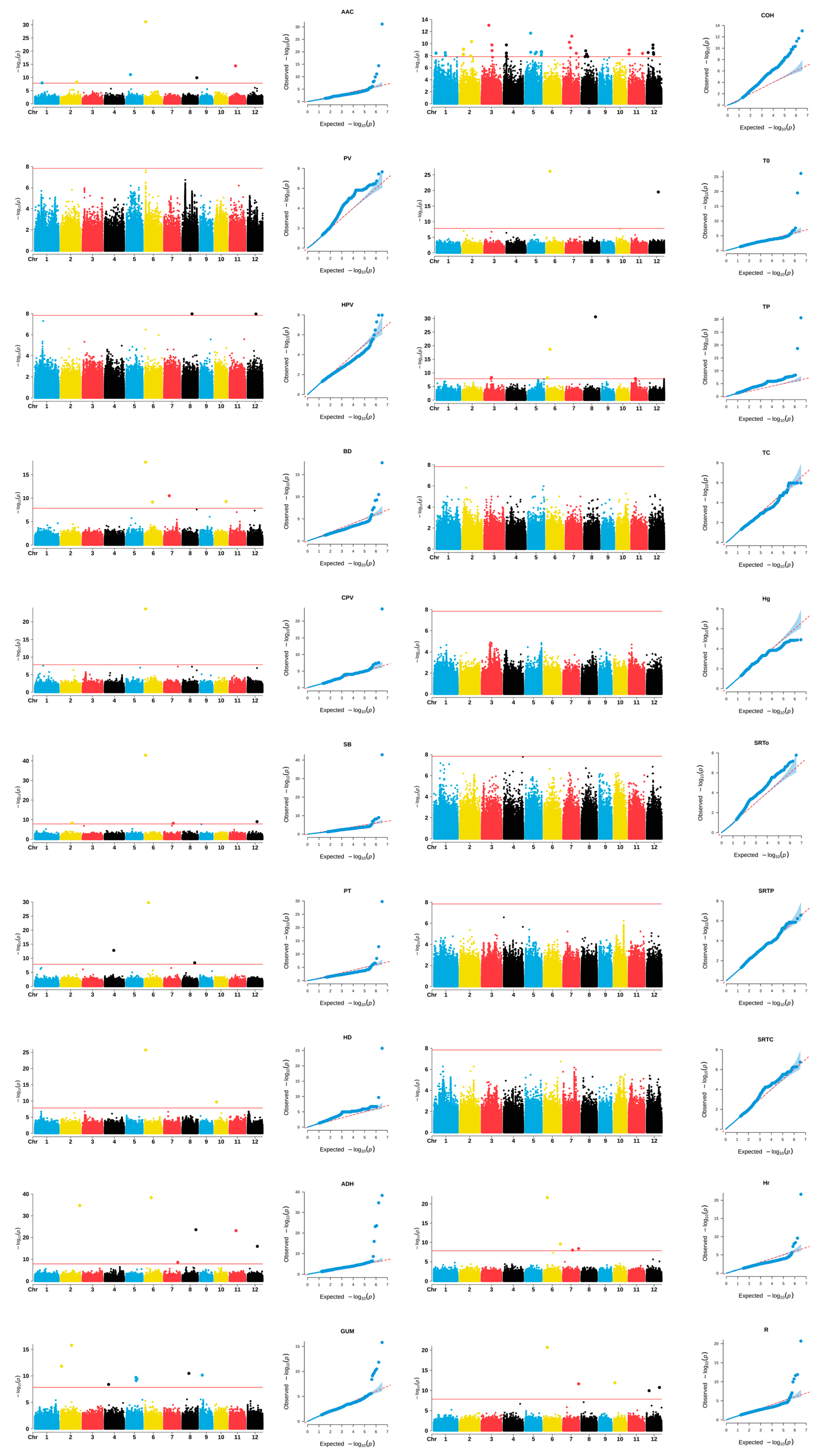
2.5. GWAS Results
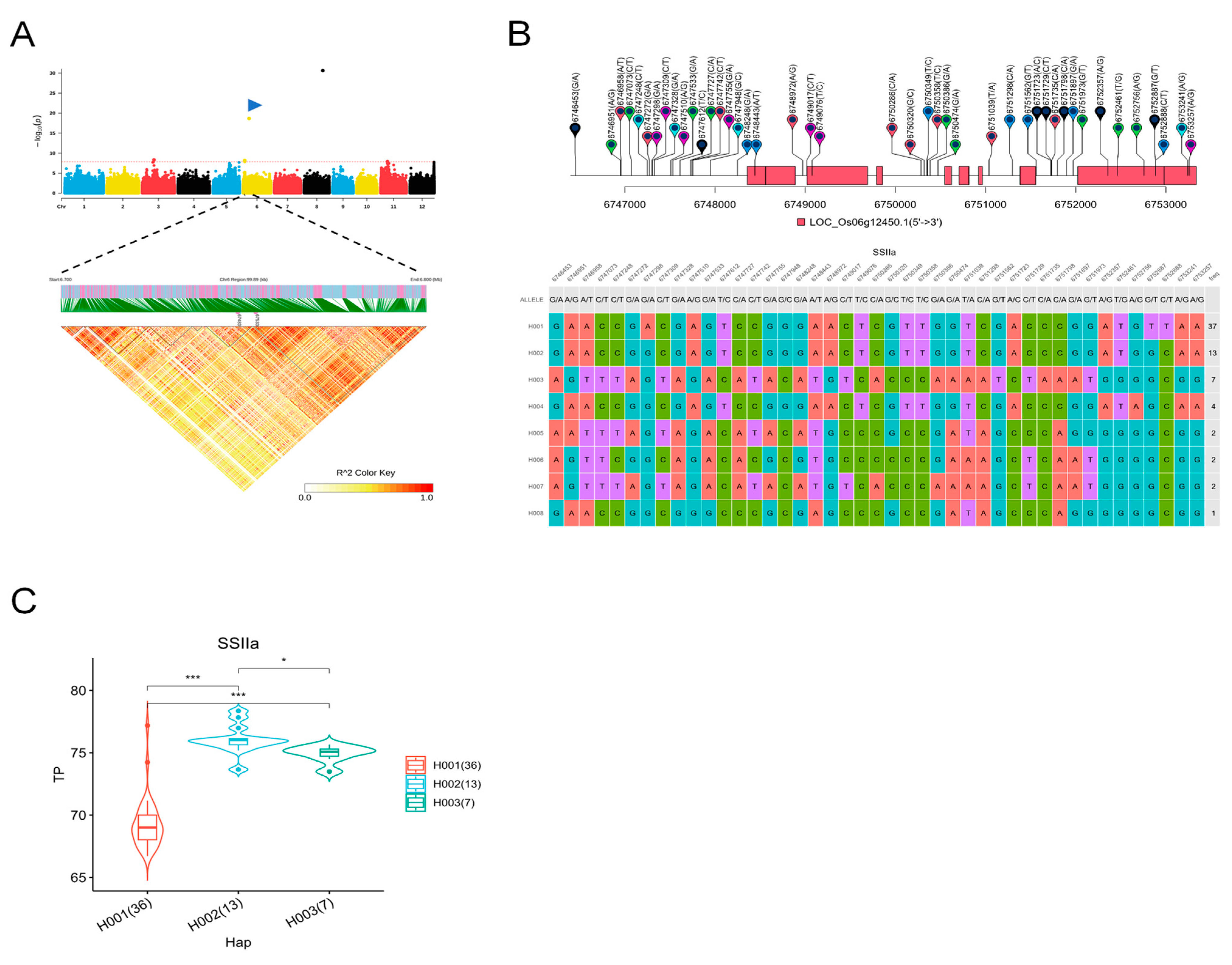
2.6. Identification of Favorable Alleles in the Wx and SSIIa Genes
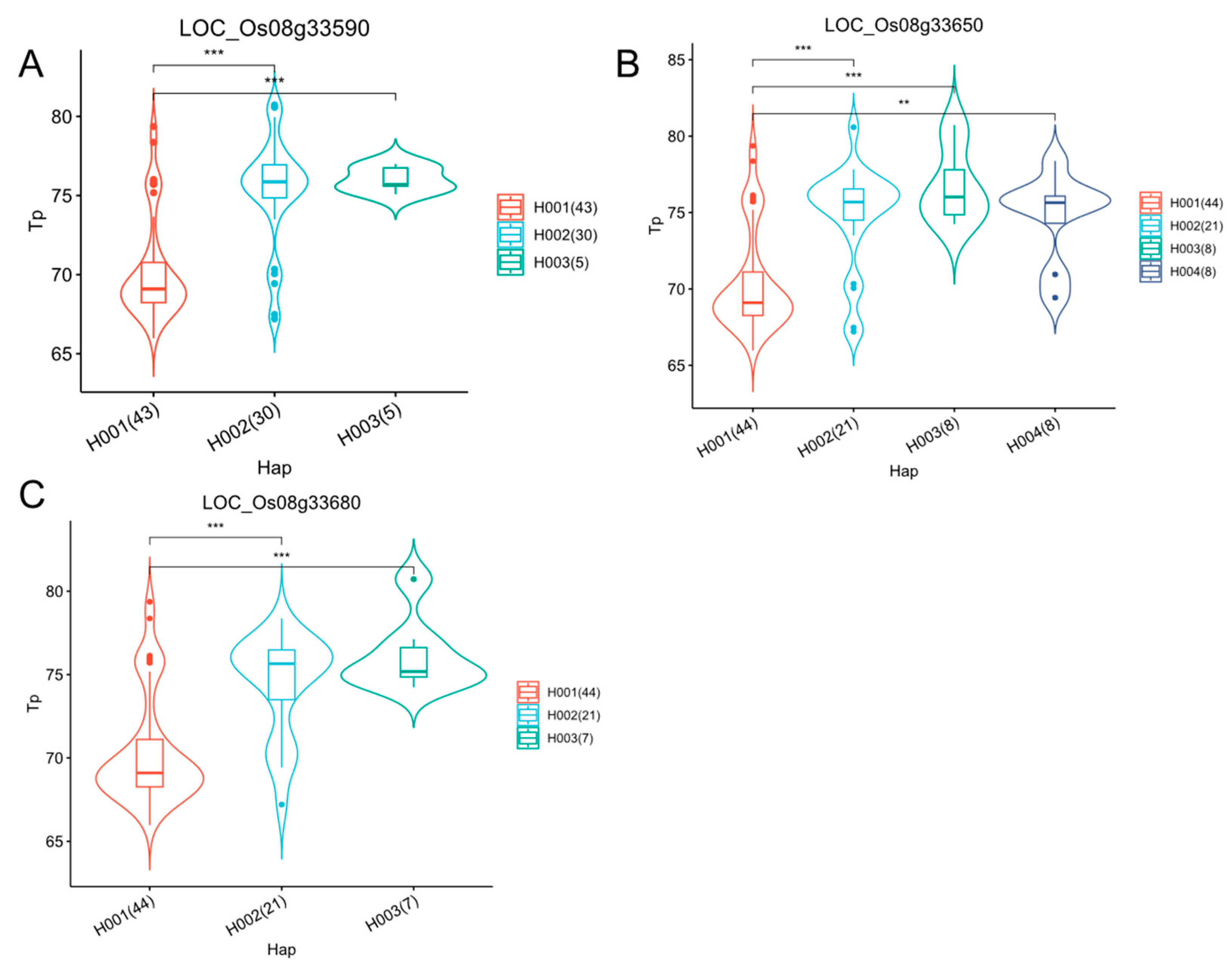
2.7. Identification of Candidate Genes
3. Discussion
3.1. Population Structure Analysis Indicates Shanlan Landrace Rice Is a Specific Rice Germplasm
3.2. Shanlan Landrace Rice Displayed Wide Diversity in Starch Physicochemical Properties
3.3. Shanlan Landrace Rice Contains Many Favorable Wx Alleles
3.4. New Loci Have Been Identified in Tp
4. Materials and Methods
4.1. Materials
4.2. DNA Extraction, Genotyping, and Linkage Disequilibrium
4.3. Population Structure, Phylogenetic Tree Construction, and Kinship Analysis
4.4. Analysis of Starch Physicochemical Properties
4.5. Genome-Wide Association Study
4.6. Candidate Gene Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, Z.X.; Lin, X.; Wang, L.; Li, C.F.; Liu, S.X. Effects of ultrasonic treatment on the cooking and fermentation properties of Shanlan rice. J. Cereal Sci. 2020, 95, 103003. [Google Scholar] [CrossRef]
- Zheng, C.M.; Huang, D.Y.; Chen, H. Affinity and hybridization of Shanlan upland rice with common rice. Chinese J. Trop. Crops 1998, 2, 74–81, (In Chinese with English Abstract). [Google Scholar]
- Yuan, N.N.; Wei, X.; Xue, D.Y.; Yang, Q.Y. The Origin and Evolution of Upland Rice in Li Ethnic Communities in Hainan Province. J. Plant Genet. Resour. 2013, 14, 202–207, (In Chinese with English Abstract). [Google Scholar]
- Yang, G.; Yang, Y.; Guan, Y.; Xu, Z.; Wang, J.; Yun, Y.; Yan, X.; Tang, Q. Genetic diversity of Shanlan upland rice (Oryza sativa L.) and association analysis of SSR markers linked to agronomic traits. Biomed Res. Int. 2021, 2021, 7588652. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Huang, Y.; Yang, X.; Su, M.; Xiong, H.; Dai, Y.; Wu, W.; Pei, X.; Yuan, Q. Genetic diversity and relationship of Shanlan upland rice were revealed based on 214 upland rice SSR markers. Plants 2023, 12, 2876. [Google Scholar] [CrossRef]
- Bao, J.S.; Deng, B.W.; Zhang, L. Molecular and genetic bases of rice cooking and eating quality: An updated review. Cereal Chem. 2023, 100, 1220–1233. [Google Scholar] [CrossRef]
- Cai, X.L.; Wang, Z.Y.; Xing, Y.; Zhang, J.L.; Hong, M.M. Aberrant splicing of intron 1 leads to the heterogeneous 5′ UTR and decreased expression of waxy gene in rice cultivars of intermediate amylose content: Aberrant splicing of rice waxy gene. Plant J. 1998, 14, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Hoai, T.T.; Matsusaka, H.; Toyosawa, Y.; Suu, T.D.; Satoh, H.; Kumamaru, T. Influence of single-nucleotide polymorphisms in the gene encoding granule-bound starch synthase I on amylose content in Vietnamese rice cultivars. Breed. Sci. 2014, 64, 142–148. [Google Scholar] [CrossRef]
- Liu, L.; Ma, X.; Liu, S.; Zhu, C.; Jiang, L.; Wang, Y.; Shen, Y.; Ren, Y.; Dong, H.; Chen, L.; et al. Identification and characterization of a novel Waxy allele from a Yunnan rice landrace. Plant Mol. Biol. 2009, 71, 609–626. [Google Scholar] [CrossRef] [PubMed]
- Mikami, I.; Aikawa, M.; Hirano, H.Y.; Sano, Y. Altered tissue-specific expression at the Wx gene of the opaque mutants in rice. Euphytica 1999, 105, 91–97. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, Y.; Sakai, M.; Imbe, T. Molecular characterization of Wx-mq, a novel mutant gene for low amylose content in endosperm of rice (Oryza sativa L.). Breed. Sci. 2002, 52, 131–135. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, J.; Chen, S.; Fan, X.; Li, Q.; Lu, Y.; Wang, M.; Yu, H.; Yi, C.; Tang, S.; et al. Wxlv, the ancestral allele of rice waxy gene. Mol. Plant 2019, 12, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, Y.; Chen, S.; Liu, X.; Zhu, J.; Zhou, L.; Lu, Y.; Li, Q.; Fan, X.; Tang, S.; et al. A rare Waxy allele coordinately improves rice eating and cooking quality and grain transparency. J. Integr. Plant Biol. 2021, 63, 889–901. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, J.; Fan, F.; Zhu, J.; Chen, T.; Wang, C.; Zheng, T.; Zhang, J.; Zhong, W.; Xu, J. Development of AS-PCR marker based on a key mutation confirmed by resequencing of Wx-mp in Milky Princess and its application in japonica soft rice (Oryza sativa L.) breeding. Plant Breed. 2013, 132, 595–603. [Google Scholar] [CrossRef]
- Biselli, C.; Cavalluzzo, D.; Perrini, R.; Gianinetti, A.; Bagnaresi, P.; Urso, S.; Orasen, G.; Desiderio, F.; Lupotto, E.; Cattivelli, L.; et al. Improvement of marker-based predictability of Apparent Amylose Content in japonica rice through GBSSI allele mining. Rice 2014, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Su, F.; Huang, S.; Mei, F.; Niu, X.; Ma, C.; Zhang, H.; Zhu, X.; Zhu, J.K.; Zhang, J. Novel Wx alleles generated by base editing for improvement of rice grain quality. J. Integr. Plant Biol. 2021, 63, 1632–1638. [Google Scholar] [CrossRef]
- Zhou, H.; Xia, D.; Zhao, D.; Li, Y.; Li, P.; Wu, B.; Gao, G.; Zhang, Q.; Wang, G.; Xiao, J.; et al. The origin of Wxla provides new insights into the improvement of grain quality in rice. J. Integr. Plant Biol. 2021, 63, 878–888. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Tseng, M.C.; Wu, Y.P.; Lin, M.Y.; Wei, F.J.; Hwu, K.K.; Hsing, Y.I.; Lin, Y.R. Genetic factors responsible for eating and cooking qualities of rice grains in a recombinant inbred population of an inter-subspecific cross. Mol. Breed. 2014, 34, 655–673. [Google Scholar] [CrossRef]
- Wang, L.Q.; Liu, W.J.; Xu, Y.; He, Y.Q.; Luo, L.J.; Xing, Y.Z.; Xu, C.G.; Zhang, Q. Genetic basis of 17 traits and viscosity parameters characterizing the eating and cooking quality of rice grain. Theor. Appl. Genet. 2007, 115, 463–476. [Google Scholar] [CrossRef]
- Wang, X.; Pang, Y.; Zhang, J.; Wu, Z.; Chen, K.; Ali, J.; Ye, G.; Xu, J.; Li, Z. Genome-wide and gene-based association mapping for rice eating and cooking characteristics and protein content. Sci. Rep. 2017, 7, 17203. [Google Scholar] [CrossRef]
- Fan, C.; Yu, X.; Xing, Y.; Xu, C.; Luo, L.; Zhang, Q. The main effects; epistatic effects and environmental interactions of QTLs on the cooking and eating quality of rice in a doubled-haploid line population. Theor. Appl. Genet. 2005, 110, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Teng, B.; Zhang, Y.; Du, S.; Wu, J.; Li, Z.; Luo, Z.; Yang, J. Crystalline, thermal and swelling properties of starches from single-segment substitution lines with different Wx alleles in rice (Oryza sativa L.). J. Sci. Food Agric. 2017, 97, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Hua, Y.; Luo, T.; Liu, C.; Zhang, B.; Zhang, X.; Liu, Y.; Zhu, Z.; Tao, Y.; Zhu, Z.; et al. Generating waxy rice starch with target type of amylopectin fine structure and gelatinization temperature by waxy gene editing. Carbohydr Polym. 2023, 306, 120595. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.X.; Qian, Q.; Liu, Q.Q.; Yan, M.X.; Liu, X.F.; Yan, C.J.; Liu, G.F.; Gao, Z.Y.; Tang, S.Z.; Zeng, D.L. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proc. Natl. Acad. Sci. USA 2009, 106, 21760–21765. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Han, Y.P.; Jiang, L.; Xu, C.W.; Lu, J.F.; Xu, M.L. Functional analysis of starch-synthesis genes in determining rice eating and cooking qualities. Mol. Breed. 2006, 18, 277–290. [Google Scholar] [CrossRef]
- Yang, F.; Chen, Y.; Tong, C.; Huang, Y.; Xu, F.; Li, K.; Corke, H.; Sun, M.; Bao, J. Association mapping of starch physicochemical properties with starch synthesis-related gene markers in nonwaxy rice (Oryza sativa L.). Mol. Breed. 2014, 34, 1747–1763. [Google Scholar] [CrossRef]
- Begum, H.; Spindel, J.E.; Lalusin, A.; Borromeo, T.; Gregorio, G.; Hernandez, J. Genome-wide association mapping for yield and other agronomic traits in an elite breeding population of tropical rice (Oryza sativa L.). PLoS ONE 2015, 10, 0119873. [Google Scholar] [CrossRef]
- Yano, K.; Yamamoto, E.; Aya, K.; Takeuchi, H.; Lo, P.C.; Hu, L. Genome-wide association study using whole-genome sequencing rapidly identifies new genes influencing agronomic traits in rice. Nat. Genet. 2016, 48, 927–934. [Google Scholar] [CrossRef]
- Reig-Valiente, J.L.; Marqués, L.; Talón, M.; Domingo, C. Genome-wide association study of agronomic traits in rice cultivated in temperate regions. BMC Genom. 2018, 19, 706. [Google Scholar] [CrossRef]
- Zhang, P.; Zhong, K.; Zhong, Z.; Tong, H. Genome-wide association study of important agronomic traits within a core collection of rice (Oryza sativa L.). BMC Plant Biol. 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Zhou, X.; and Huang, X. Genome-wide association studies in rice: How to solve the low power problems? Mol. Plant 2019, 12, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.K.; Chetia, S.K.; Dey, P.C.; Rahman, A.; Saikia, S.; Sharma, V.; Sharma, H.; Sen, P.; Modi, M.K. Genome-wide association studies for agronomical traits in winter rice accessions of Assam. Genomics 2021, 113, 1037–1047. [Google Scholar] [CrossRef] [PubMed]
- Biselli, C.; Volante, A.; Desiderio, F.; Tondelli, A.; Gianinetti, A.; Finocchiaro, F.; Taddei, F.; Gazza, L.; Sgrulletta, D.; Cattivelli, L.; et al. GWAS for starch-related parameters in japonica rice (Oryza sativa L.). Plants 2019, 8, 292. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Rashid, M.A.R.; Zhang, Y.; Zhao, Y.; Pan, Y. Genome wide association study on development and evolution of glutinous rice. BMC Genom. Data 2022, 23, 33. [Google Scholar] [CrossRef]
- Matsushima, R.; Maekawa, M.; Kusano, M.; Tomita, K.; Kondo, H.; Nishimura, H.; Crofts, N.; Fujita, N.; Sakamoto, W. Amyloplast membrane protein substandard starch grain6 controls starch grain size in rice endosperm. Plant Physiol. 2016, 170, 1445–1459. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wei, X.; Sang, T.; Zhao, Q.; Feng, Q.; Zhao, Y.; Li, C.; Zhu, C.; Lu, T.; Zhang, Z.; et al. Genome-wide association studies of 14 agronomic traits in rice landraces. Nat. Genet. 2010, 42, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; He, H.; Zou, Y.; Chen, W.; Yu, R.; Liu, X. Development and application of a set of breeder-friendly SNP markers for genetic analyses and molecular breeding of rice (Oryza sativa L.). Theor. Appl. Genet. 2011, 123, 869–879. [Google Scholar] [CrossRef]
- Xie, W.; Wang, G.; Yuan, M.; Yao, W.; Lyu, K.; Zhao, H.; Yang, M.; Li, P.; Zhang, X.; Yuan, J.; et al. Breeding signatures of rice improvement revealed by a genomic variation map from a large germplasm collection. Proc. Natl. Acad. Sci. USA 2015, 112, 5411–5419. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic variation in 3010 diverse accessions of Asian cultivated rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef]
- Toosang, S.; Jamjod, S.; Pusadee, T. Characterization of grain quality traits of Thai highland glutinous rice landraces. Chiang Mai J. Sci. 2024, 51, e2024007. [Google Scholar] [CrossRef]
- Feng, F.; Li, Y.; Qin, X.; Liao, Y.; Siddique, K.H.M. Changes in rice grain quality of indica and japonica type varieties released in China from 2000 to 2014. Front. Plant Sci. 2017, 8, 1863. [Google Scholar] [CrossRef]
- Shi, S.; Wang, E.; Li, C.; Cai, M.; Cheng, B.; Cao, C.; Jiang, Y. Use of protein content, amylose content; and RVA parameters to evaluate the taste quality of rice. Front. Nutr. 2022, 8, 758547. [Google Scholar] [CrossRef]
- Li, H.; Prakash, S.; Nicholson, T.M.; Fitzgerald, M.A.; Gilbert, R.G. Instrumental measurement of cooked rice texture by dynamic rheological testing and its relation to the fine structure of rice starch. Carbohydr. Polym. 2016, 146, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, R.P.O.; Domingo, C.J.; Sreenivasulu, N. Multivariate-based classification of predicting cooking quality ideotypes in rice (Oryza sativa L.) indica germplasm. Rice 2018, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Suzuki, K.; Iijima, K.; Ebana, K. Variation in cooking and eating quality traits in japanese rice germplasm accessions. Breed. Sci. 2016, 66, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Bao, J.; Corke, H.; Sun, M. Association analysis of markers derived from starch biosynthesis related genes with starch physicochemical properties in the USDA rice mini-core collection. Front. Plant Sci. 2017, 8, 424. [Google Scholar] [CrossRef]
- Zhao, C.; Zhao, L.; Zhao, Q.; Chen, T.; Yao, S.; Zhu, Z.; Zhou, L.; Nadaf, A.B.; Liang, W.; Lu, K.; et al. Genetic dissection of eating and cooking qualities in different subpopulations of cultivated rice (Oryza sativa L.) through association mapping. BMC Genet. 2020, 21, 119. [Google Scholar] [CrossRef]
- Misra, G.; Badoni, S.; Domingo, C.J.; Cuevas, R.P.O.; Llorente, C.; Mbanjo, E.G.N.; Sreenivasulu, N. Deciphering the genetic architecture of cooked rice texture. Front. Plant Sci. 2018, 9, 1405. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Song, S.; Hu, C.; Jing, C.; Xu, Q.; Li, X.; Zhang, M.; Hai, M.; Shen, J.; Zhang, Y.; et al. QTL detection and candidate gene identification for eating and cooking quality traits in rice (Oryza sativa L.) via a Genome-Wide Association Study. Int. J. Mol. Sci. 2024, 25, 630. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Wang, Y.; Wang, J.; Li, Y.; Zhang, Y.; Zhao, X.; Xu, J.; Li, Z.; Zhao, T.; Wang, W.; et al. The basic helix-loop-helix transcription factor gene, OsbHLH38, plays a key role in controlling rice salt tolerance. J. Integr. Plant Biol. 2023, 65, 1859–1873. [Google Scholar] [CrossRef]
- Guo, T.; Yu, H.; Qiu, J.; Li, J.; Han, B.; Lin, H. Advances in rice genetics and breeding by molecular design in China (in Chinese). Sci. Vitae 2019, 49, 1185–1212. [Google Scholar]
- Zeng, D.; Tian, Z.; Rao, Y.; Dong, G.; Yang, Y.; Huang, L.; Leng, Y.; Xu, J.; Sun, C.; Zhang, G.; et al. Rational design of high-yield and superior-quality rice. Nat. Plants 2017, 3, 17031. [Google Scholar] [CrossRef]
- Jin, L.; Lu, Y.; Shao, Y.; Zhang, G.; Xiao, P.; Shen, S.; Corke, H.; Bao, J. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J. Cereal Sci. 2010, 51, 159–164. [Google Scholar] [CrossRef]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAM tools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Dong, S.S.; Xu, J.Y.; He, W.M.; Yang, T.L. PopLDdecay: A fast and effective tool for linkage disequilibrium decay analysis based on variant call format files. Bioinformatics 2019, 35, 1786–1788. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. Plink: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Martínez-Cortés, G.; Salazar-Flores, J.; Fernández-Rodríguez, L.G.; Rubi-Castellanos, R.; Rodríguez-Loya, C.; Velarde-Félix, J.S.; Muñoz-Valle, J.F.; Parra-Rojas, I.; Rangel-Villalobos, H. Admixture and population structure in Mexican-Mestizos based on paternal lineages. J. Hum. Genet. 2012, 57, 568–574. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, Y.; Zhang, Y.; Hu, Y.; Ying, Y.; Xu, F.; Bao, J. Variation in starch physicochemical properties of rice with different genic allele combinations in two environments. J. Cereal Sci. 2022, 108, 103575. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, Z. GAPIT Version 3: Boosting power and accuracy for genomic association and prediction. Genom. Proteom. Bioinform. 2021, 19, 629–640. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNPs | Number | InDels | Number |
|---|---|---|---|
| Total SNPs | 11,266,589 | Total Indels | 1,635,574 |
| SNPs in exon | 2,619,968 | Indels in exon | 158,603 |
| SNPs in intron | 1,725,345 | Indels in intron | 379,545 |
| SNPs in UTR5 | 84,864 | Indels in UTR5 | 42,070 |
| SNPs in UTR3 | 139,473 | Indels in UTR3 | 51,556 |
| SNPs in intergenic region | 2,399,456 | Indels in intergenic region | 527,164 |
| Upstream | 2,182,885 | Upstream | 486,752 |
| Downstream | 1,735,090 | Downstream | 390,218 |
| Non-synonymous SNPs | 1,412,309 | Splicing Indels | 1971 |
| Splicing SNPs | 12,979 | Stop-gain Indels | 6580 |
| Stop-gain SNPs | 77,011 | Stop-loss Indels | 343 |
| Stop-loss SNPs | 4254 | Frameshift Indels | 95,147 |
| Non frameshift Indels | 49,081 |
| SNP Position 1 | p-Value | maf | Effect | PVE (%) 2 | Candidate Gene | |
|---|---|---|---|---|---|---|
| AAC | S1_13164701 | 1.17 × 10−8 | 0.48 | 0.04 | 0.32 | |
| S2_29156368 | 5.70 × 10−9 | 0.05 | −0.04 | 3.87 | ||
| S2_34721769 | 1.80 × 10−35 | 0.07 | 24.4 | 0.26 | ||
| S5_7890154 | 8.14 × 10−12 | 0.1 | 0.04 | 1.38 | ||
| S6_1779126 | 7.35 × 10−32 | 0.28 | 0.05 | 45.93 | Wx | |
| S7_25202120 | 2.73 × 10−9 | 0.16 | −6.76 | 0.08 | ||
| S8_24746239 | 2.52 × 10−24 | 0.2 | 16.73 | 0.59 | ||
| S11_10994972 | 3.95 × 10−15 | 0.09 | 0.05 | 5.4 | ||
| S12_17949665 | 1.22 × 10−16 | 0.2 | 6.67 | 1.21 | ||
| BD | S6_1721569 | 2.09 × 10−18 | 0.12 | −19.66 | 34.23 | Wx |
| S10_20833372 | 5.27 × 10−10 | 0.45 | 12.72 | 6.09 | ||
| GUM | S2_1491276 | 1.48 × 10−12 | 0.24 | −1.67 | 10.54 | |
| S2_19767079 | 1.62 × 10−16 | 0.05 | 3.97 | 27.19 | ||
| S4_7283069 | 4.07 × 10−9 | 0.09 | 1.45 | 18.39 | ||
| S5_17739626 | 1.97 × 10−10 | 0.17 | 1.26 | 29.53 | ||
| S5_18190039 | 7.49 × 10−10 | 0.26 | −1 | 15.46 | ||
| S6_1784985 | 1.87 × 10−26 | 0.37 | 7.25 | 18.47 | Wx | |
| S8_11936414 | 3.32 × 10−11 | 0.24 | 1.43 | 4.22 | ||
| HPV | S7_28619594 | 4.13 × 10−9 | 0.11 | 0.97 | 0 | |
| S12_15381517 | 1.08 × 10−8 | 0.12 | 17.44 | 19.16 | ||
| R | S6_6745643 | 2.12 × 10−21 | 0.49 | 0.15 | 45.43 | SSIIa |
| S7_28619594 | 2.33 × 10−12 | 0.11 | 0.12 | 5.56 | ||
| S10_2389557 | 1.33 × 10−12 | 0.27 | 0.1 | 9.45 | ||
| S12_4781628 | 1.14 × 10−10 | 0.08 | 0.12 | 11.5 | ||
| SB | S6_1620142 | 1.31 × 10−43 | 0.5 | −53.22 | 62.6 | SSG6 |
| S7_16991812 | 6.79 × 10−9 | 0.15 | 21.03 | 6.77 | ||
| S12_17341408 | 1.03 × 10−9 | 0.11 | 19.38 | 3.9 | ||
| To | S12_16601981 | 3.15 × 10−20 | 0.07 | 3.34 | 22.08 | |
| Tp | S3_13207912 | 4.79 × 10−9 | 0.47 | 3.77 | 4.63 | |
| S6_6745643 | 2.15 × 10−19 | 0.5 | 2.89 | 45.91 | SSIIa | |
| S8_20995138 | 2.43 × 10−31 | 0.49 | −12.01 | 33.73 | ||
| S11_7696066 | 1.25 × 10−8 | 0.47 | −3.63 | 1.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Deng, B.; Peng, Y.; Gao, Y.; Hu, Y.; Bao, J. Population Structure and Genetic Diversity of Shanlan Landrace Rice for GWAS of Cooking and Eating Quality Traits. Int. J. Mol. Sci. 2024, 25, 3469. https://doi.org/10.3390/ijms25063469
Zhang L, Deng B, Peng Y, Gao Y, Hu Y, Bao J. Population Structure and Genetic Diversity of Shanlan Landrace Rice for GWAS of Cooking and Eating Quality Traits. International Journal of Molecular Sciences. 2024; 25(6):3469. https://doi.org/10.3390/ijms25063469
Chicago/Turabian StyleZhang, Lin, Bowen Deng, Yi Peng, Yan Gao, Yaqi Hu, and Jinsong Bao. 2024. "Population Structure and Genetic Diversity of Shanlan Landrace Rice for GWAS of Cooking and Eating Quality Traits" International Journal of Molecular Sciences 25, no. 6: 3469. https://doi.org/10.3390/ijms25063469