

Smooth Muscle Heterogeneity and Plasticity in Health and Aortic Aneurysmal Disease
Abstract
:1. Introduction
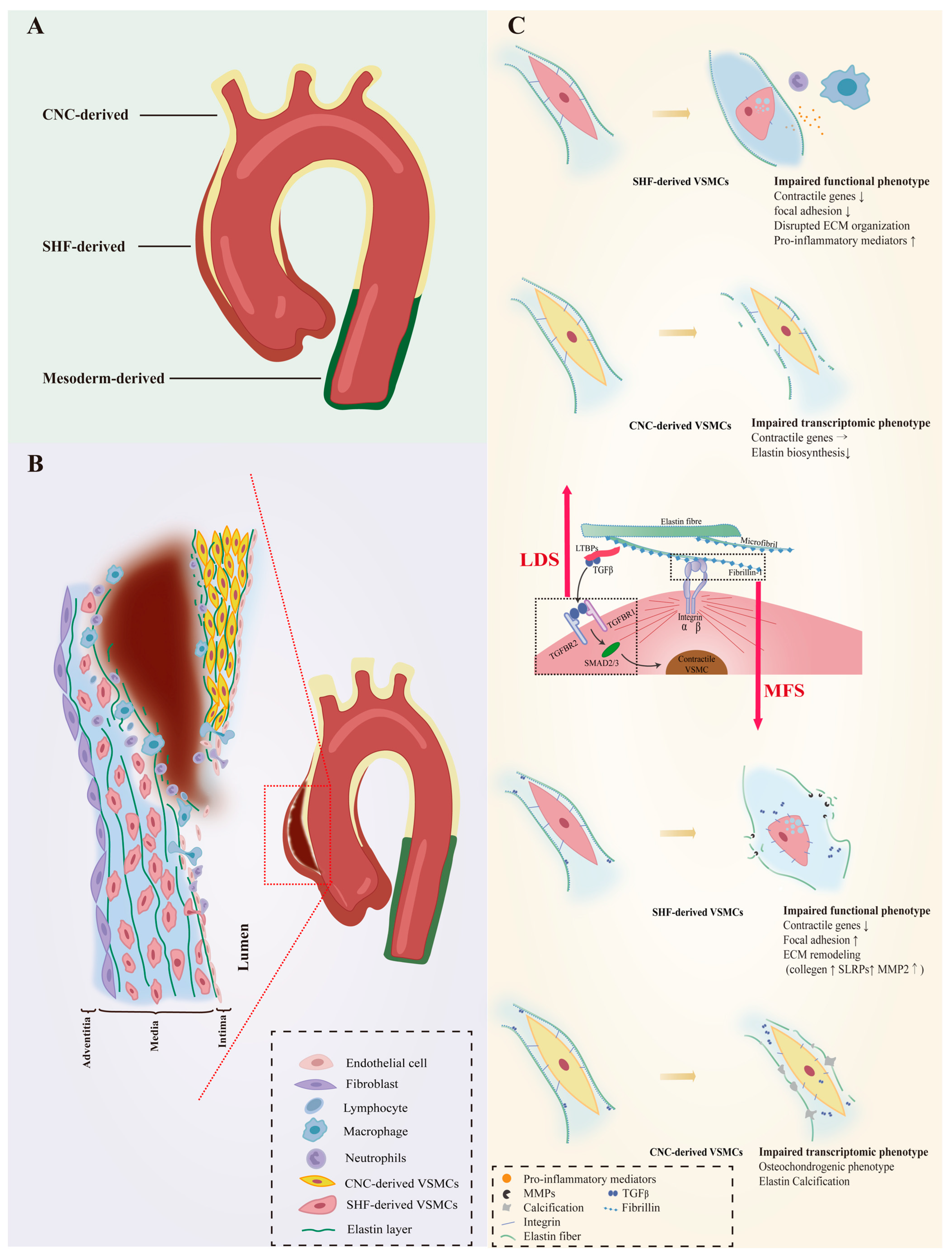
2. VSMC Heterogeneity in Normal Aorta
3. VSMC Phenotypic Diversity in Hereditary AAs
3.1. VSMC Phenotypic Diversity in LDS
3.2. VSMC Phenotypic Diversity in MFS
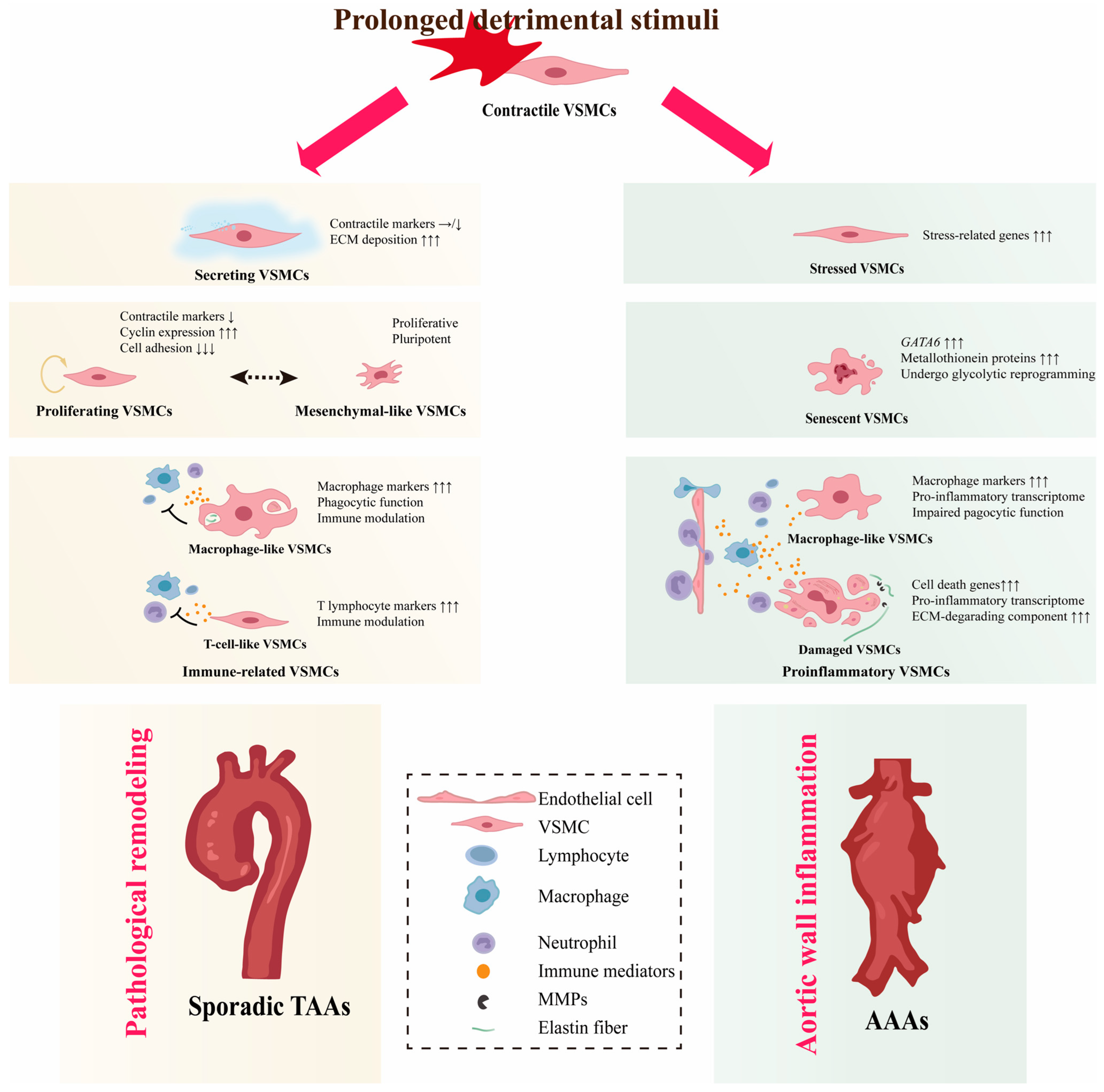
4. VSMC Phenotypic Diversity in Non-Hereditary AAs
4.1. VSMC Diversity in TAAs
4.1.1. Secreting VSMCs
4.1.2. Proliferating VSMCs
4.1.3. Immune-Related VSMCs
4.2. VSMC Diversity in AAAs
4.2.1. Proinflammatory VSMCs
4.2.2. Senescent VSMCs
4.2.3. Stressed VSMCs
5. Role of VSMC Phenotypic Diversity in AAs
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, Y.H.; LeMaire, S.A.; Webb, N.R.; Cassis, L.A.; Daugherty, A.; Lu, H.S. Aortic Aneurysms and Dissections Series. Arterioscler. Thromb. Vasc. Biol. 2020, 40, e37–e46. [Google Scholar] [CrossRef] [PubMed]
- Pinard, A.; Jones, G.T.; Milewicz, D.M. Genetics of Thoracic and Abdominal Aortic Diseases. Circ. Res. 2019, 124, 588–606. [Google Scholar] [CrossRef] [PubMed]
- Arnaud, P.; Hanna, N.; Benarroch, L.; Aubart, M.; Bal, L.; Bouvagnet, P.; Busa, T.; Dulac, Y.; Dupuis-Girod, S.; Edouard, T.; et al. Genetic diversity and pathogenic variants as possible predictors of severity in a French sample of nonsyndromic heritable thoracic aortic aneurysms and dissections (nshTAAD). Genet Med. 2019, 21, 2015–2024. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.; Badger, S.; McFarland, M.; Hughes, A. Abdominal Aortic Aneurysm Genetic Associations: Mostly False? A Systematic Review and Meta-analysis. Eur. J. Vasc. Endovasc. Surg. 2015, 51, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, C.X.; Malecki, C.; Robertson, E.; Hambly, B.; Jeremy, R. Molecular Mechanisms in Genetic Aortopathy–Signaling Pathways and Potential Interventions. Int. J. Mol. Sci. 2023, 24, 1795. [Google Scholar] [CrossRef]
- Davis, E.C. Smooth muscle cell to elastic lamina connections in developing mouse aorta. Role in aortic medial organization. Lab. Investig. 1993, 68, 89–99. [Google Scholar]
- Chamley-Campbell, J.H.; Campbell, G.R. What controls smooth muscle phenotype? Atherosclerosis 1981, 40, 347–357. [Google Scholar] [CrossRef]
- Pyun, J.-H.; Ahn, B.-Y.; Vuong, T.A.; Kim, S.W.; Jo, Y.; Jeon, J.; Baek, S.H.; Kim, J.; Park, S.; Bae, G.-U.; et al. Inducible Prmt1 ablation in adult vascular smooth muscle leads to contractile dysfunction and aortic dissection. Exp. Mol. Med. 2021, 53, 1569–1579. [Google Scholar] [CrossRef]
- Chiarini, A.; Onorati, F.; Marconi, M.; Pasquali, A.; Patuzzo, C.; Malashicheva, A.; Irtyega, O.; Faggian, G.; Pignatti, P.F.; Trabetti, E.; et al. Studies on sporadic non-syndromic thoracic aortic aneurysms: II. Alterations of extra-cellular matrix components and focal adhesion proteins. Eur. J. Prev. Cardiol. 2018, 25, 51–58. [Google Scholar] [CrossRef]
- Chiarini, A.; Onorati, F.; Marconi, M.; Pasquali, A.; Patuzzo, C.; Malashicheva, A.; Irtyega, O.; Faggian, G.; Pignatti, P.F.; Trabetti, E.; et al. Studies on sporadic non-syndromic thoracic aortic aneurysms: 1. Deregulation of Jagged/Notch 1 homeostasis and selection of synthetic/secretor phenotype smooth muscle cells. Eur. J. Prev. Cardiol. 2018, 25, 42–50. [Google Scholar] [CrossRef]
- Bochaton-Piallat, M.-L.; Bäck, M. Novel concepts for the role of smooth muscle cells in vascular disease: Towards a new smooth muscle cell classification. Cardiovasc. Res. 2018, 114, 477–480. [Google Scholar] [CrossRef]
- Stenmark, K.R.; Frid, M.G.; Graham, B.B.; Tuder, R.M. Dynamic and diverse changes in the functional properties of vascular smooth muscle cells in pulmonary hypertension. Cardiovasc. Res. 2018, 114, 551–564. [Google Scholar] [CrossRef]
- Frid, M.G.; Dempsey, E.C.; Durmowicz, A.G.; Stenmark, K.R. Smooth Muscle Cell Heterogeneity in Pulmonary and Systemic Vessels. Importance in vascular disease. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1203–1209. [Google Scholar] [CrossRef]
- Allahverdian, S.; Chaabane, C.; Boukais, K.; Francis, G.A.; Bochaton-Piallat, M.-L. Smooth muscle cell fate and plasticity in atherosclerosis. Cardiovasc. Res. 2018, 114, 540–550. [Google Scholar] [CrossRef] [Green Version]
- Gomez, D.; Owens, G.K. Smooth muscle cell phenotypic switching in atherosclerosis. Cardiovasc. Res. 2012, 95, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.H.; Campbell, G.R. Smooth Muscle Phenotypic Modulation—A Personal Experience. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1784–1789. [Google Scholar] [CrossRef] [Green Version]
- Jauhiainen, S.; Kiema, M.; Hedman, M.; Laakkonen, J.P. Large Vessel Cell Heterogeneity and Plasticity: Focus in Aortic Aneurysms. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 811–818. [Google Scholar] [CrossRef]
- Liu, M.; Gomez, D. Smooth Muscle Cell Phenotypic Diversity. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1715–1723. [Google Scholar] [CrossRef]
- Majesky, M.W. Developmental Basis of Vascular Smooth Muscle Diversity. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 1248–1258. [Google Scholar] [CrossRef] [Green Version]
- Waldo, K.L.; Hutson, M.R.; Ward, C.C.; Zdanowicz, M.; Stadt, H.A.; Kumiski, D.; Abu-Issa, R.; Kirby, M.L. Secondary heart field contributes myocardium and smooth muscle to the arterial pole of the developing heart. Dev. Biol. 2005, 281, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Rowitch, D.H.; Soriano, P.; McMahon, A.P.; Sucov, H.M. Fate of the mammalian cardiac neural crest. Development 2000, 127, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Wasteson, P.; Johansson, B.R.; Jukkola, T.; Breuer, S.; Akyürek, L.M.; Partanen, J.; Lindahl, P. Developmental origin of smooth muscle cells in the descending aorta in mice. Development 2008, 135, 1823–1832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, H.; Rateri, D.L.; Moorleghen, J.J.; Majesky, M.W.; Daugherty, A. Smooth Muscle Cells Derived From Second Heart Field and Cardiac Neural Crest Reside in Spatially Distinct Domains in the Media of the Ascending Aorta—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 1722–1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkels, H.; Ehinger, E.; Vassallo, M.; Buscher, K.; Dinh, H.Q.; Kobiyama, K.; Hamers, A.A.; Cochain, C.; Vafadarnejad, E.; Saliba, A.-E.; et al. Atlas of the Immune Cell Repertoire in Mouse Atherosclerosis Defined by Single-Cell RNA-Sequencing and Mass Cytometry. Circ. Res. 2018, 122, 1675–1688. [Google Scholar] [CrossRef]
- Kalluri, A.S.; Vellarikkal, S.K.; Edelman, E.R.; Nguyen, L.; Subramanian, A.; Ellinor, P.T.; Regev, A.; Kathiresan, S.; Gupta, R.M. Single-Cell Analysis of the Normal Mouse Aorta Reveals Functionally Distinct Endothelial Cell Populations. Circulation 2019, 140, 147–163. [Google Scholar] [CrossRef]
- Dobnikar, L.; Taylor, A.L.; Chappell, J.; Oldach, P.; Harman, J.L.; Oerton, E.; Dzierzak, E.; Bennett, M.R.; Spivakov, M.; Jørgensen, H.F. Disease-relevant transcriptional signatures identified in individual smooth muscle cells from healthy mouse vessels. Nat. Commun. 2018, 9, 4567. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Zhang, J.; Gao, A.; Zhang, M.; Wang, Z.; Yu, F.; Guo, X.; Su, G.; Zhang, Y.; Zhang, C. An intersegmental single-cell profile reveals aortic heterogeneity and identifies a novel Malat1+ vascular smooth muscle subtype involved in abdominal aortic aneurysm formation. Signal Transduct. Target. Ther. 2022, 7, 125. [Google Scholar] [CrossRef]
- Upadhyay, G.; Yin, Y.; Yuan, H.; Li, X.; Derynck, R.; Glazer, R.I. Stem cell antigen-1 enhances tumorigenicity by disruption of growth differentiation factor-10 (GDF10)–dependent TGF-β signaling. Proc. Natl. Acad. Sci. USA 2011, 108, 7820–7825. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Z.; Torsney, E.; Afzal, A.R.; Davison, F.; Metzler, B.; Xu, Q. Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J. Clin. Investig. 2004, 113, 1258–1265. [Google Scholar] [CrossRef]
- Wang, G.; Jacquet, L.; Karamariti, E.; Xu, Q. Origin and differentiation of vascular smooth muscle cells. J. Physiol. 2015, 593, 3013–3030. [Google Scholar] [CrossRef] [Green Version]
- Sajeesh, S.; Broekelman, T.; Mecham, R.P.; Ramamurthi, A. Stem cell derived extracellular vesicles for vascular elastic matrix regenerative repair. Acta Biomater. 2020, 113, 267–278. [Google Scholar] [CrossRef]
- Sainz, J.; Zen, A.A.H.; Caligiuri, G.; Demerens, C.; Urbain, D.; Lemitre, M.; Lafont, A. Isolation of “Side Population” Progenitor Cells From Healthy Arteries of Adult Mice. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, T.; Freis, E.D. Pressure Pulse in Small Arteries. Circ. Res. 1962, 11, 838–842. [Google Scholar] [CrossRef]
- Leuprecht, A.; Perktold, K.; Kozerke, S.; Boesiger, P. Combined CFD and MRI study of blood flow in a human ascending aorta model. Biorheology 2002, 39, 425–429. [Google Scholar]
- Vincent, P.E.; Plata, A.M.; Hunt, A.A.E.; Weinberg, P.D.; Sherwin, S.J. Blood flow in the rabbit aortic arch and descending thoracic aorta. J. R. Soc. Interface 2011, 8, 1708–1719. [Google Scholar] [CrossRef] [Green Version]
- Stein, P.D.; Sabbah, H.N. Turbulent blood flow in the ascending aorta of humans with normal and diseased aortic valves. Circ. Res. 1976, 39, 58–65. [Google Scholar] [CrossRef]
- Chandran, K.B. Flow Dynamics in the Human Aorta. J. Biomech. Eng. 1993, 115, 611–616. [Google Scholar] [CrossRef]
- Cheung, C.; Bernardo, A.S.; Trotter, M.W.B.; Pedersen, R.A.; Sinha, S. Generation of human vascular smooth muscle subtypes provides insight into embryological origin–dependent disease susceptibility. Nat. Biotechnol. 2012, 30, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Gadson, P.F., Jr.; Dalton, M.L.; Patterson, E.; Svoboda, D.D.; Hutchinson, L.; Schram, D.; Rosenquist, T.H. Differential Response of Mesoderm- and Neural Crest-Derived Smooth Muscle to TGF-β1: Regulation of c-myb and α1 (I) Procollagen Genes. Exp. Cell Res. 1997, 230, 169–180. [Google Scholar] [CrossRef]
- Topouzis, S.; Majesky, M.W. Smooth Muscle Lineage Diversity in the Chick Embryo. Two types of aortic smooth muscle cell differ in growth and receptor-mediated transcriptional responses to transforming growth factor-beta. Dev. Biol. 1996, 178, 430–445. [Google Scholar] [CrossRef]
- Thieszen, S.L.; Dalton, M.; Gadson, P.F.; Patterson, E.; Rosenquist, T.H. Embryonic Lineage of Vascular Smooth Muscle Cells Determines Responses to Collagen Matrices and Integrin Receptor Expression. Exp. Cell Res. 1996, 227, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Oyama, N.; Gona, P.; Salton, C.J.; Chuang, M.L.; Jhaveri, R.R.; Blease, S.J.; Manning, A.R.; Lahiri, M.; Botnar, R.M.; Levy, D.; et al. Differential Impact of Age, Sex, and Hypertension on Aortic Atherosclerosis: The Framingham Heart Study. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neisius, U.; Gona, P.N.; Oyama-Manabe, N.; Chuang, M.L.; O’donnell, C.J.; Manning, W.J.; Tsao, C.W. Relation of MRI Aortic Wall Area and Plaque to Incident Cardiovascular Events: The Framingham Heart Study. Radiology 2022, 304, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Haimovici, H.; Maier, N. Fate of aortic homografts in canine atherosclerosis. 3. study of fresh abdominal and thoracic aortic implants into thoracic aorta: Role of tissue susceptibility in atherogenesis. Arch. Surg. 1964, 89, 961–969. [Google Scholar] [CrossRef]
- Leroux-Berger, M.; Queguiner, I.; Maciel, T.T.; Ho, A.; Relaix, F.; Kempf, H. Pathologic calcification of adult vascular smooth muscle cells differs on their crest or mesodermal embryonic origin. J. Bone Miner. Res. 2011, 26, 1543–1553. [Google Scholar] [CrossRef]
- Muyor, K.; Laget, J.; Cortijo, I.; Duranton, F.; Jover, B.; Argilés, À.; Gayrard, N. Vascular calcification in different arterial beds in ex vivo ring culture and in vivo rat model. Sci. Rep. 2022, 12, 11861. [Google Scholar] [CrossRef]
- Li, Y.; Fang, M.; Yang, J.; Yu, C.; Kuang, J.; Sun, T.; Fan, R. Analysis of the contribution of 129 candidate genes to thoracic aortic aneurysm or dissection of a mixed cohort of sporadic and familial cases in South China. Am. J. Transl. Res. 2021, 13, 4281–4295. [Google Scholar]
- Isselbacher, E.M.; Cardenas, C.L.; Lindsay, M.E. Hereditary Influence in Thoracic Aortic Aneurysm and Dissection. Circulation 2016, 133, 2516–2528. [Google Scholar] [CrossRef] [Green Version]
- Renard, M.; Francis, C.; Ghosh, R.; Scott, A.F.; Witmer, P.D.; Adès, L.C.; Andelfinger, G.U.; Arnaud, P.; Boileau, C.; Callewaert, B.L.; et al. Clinical Validity of Genes for Heritable Thoracic Aortic Aneurysm and Dissection. J. Am. Coll. Cardiol. 2018, 72, 605–615. [Google Scholar] [CrossRef]
- Sakai, L.Y.; Keene, D.R. Fibrillin protein pleiotropy: Acromelic dysplasias. Matrix Biol. 2019, 80, 6–13. [Google Scholar] [CrossRef]
- Prakash, S.K.; LeMaire, S.A.; Guo, D.-C.; Russell, L.; Regalado, E.S.; Golabbakhsh, H.; Johnson, R.J.; Safi, H.J.; Estrera, A.L.; Coselli, J.S.; et al. Rare Copy Number Variants Disrupt Genes Regulating Vascular Smooth Muscle Cell Adhesion and Contractility in Sporadic Thoracic Aortic Aneurysms and Dissections. Am. J. Hum. Genet. 2010, 87, 743–756. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Cai, L.; Jia, L.; Li, X.; Xi, X.; Zheng, S.; Liu, X.; Piao, C.; Liu, T.; Sun, Z.; et al. Wide mutation spectrum and frequent variant Ala27Thr of FBN1 identified in a large cohort of Chinese patients with sporadic TAAD. Sci. Rep. 2015, 5, 13115. [Google Scholar] [CrossRef]
- Li, Y.; Gao, S.; Han, Y.; Song, L.; Kong, Y.; Jiao, Y.; Huang, S.; Du, J.; Li, Y. Variants of Focal Adhesion Scaffold Genes Cause Thoracic Aortic Aneurysm. Circ. Res. 2021, 128, 8–23. [Google Scholar] [CrossRef]
- Liu, C.-L.; Liu, X.; Zhang, Y.; Liu, J.; Yang, C.; Luo, S.; Liu, T.; Wang, Y.; Lindholt, J.S.; Diederichsen, A.; et al. Eosinophils Protect Mice From Angiotensin-II Perfusion–Induced Abdominal Aortic Aneurysm. Circ. Res. 2021, 128, 188–202. [Google Scholar] [CrossRef]
- Benjamins, J.W.; Yeung, M.W.; van de Vegte, Y.J.; Said, M.A.; van der Linden, T.; Ties, D.; Juarez-Orozco, L.E.; Verweij, N.; van der Harst, P. Genomic insights in ascending aortic size and distensibility. Ebiomedicine 2022, 75, 103783. [Google Scholar] [CrossRef]
- Milewicz, D.M.; Braverman, A.C.; De Backer, J.; Morris, S.A.; Boileau, C.; Maumenee, I.H.; Jondeau, G.; Evangelista, A.; Pyeritz, R.E. Marfan syndrome. Nat. Rev. Dis. Primers 2021, 7, 64. [Google Scholar] [CrossRef]
- Van Laer, L.; Dietz, H.; Loeys, B. Loeys-Dietz syndrome. Adv. Exp. Med. Biol. 2014, 802, 95–105. [Google Scholar] [CrossRef]
- Gouda, P.; Kay, R.; Habib, M.; Aziz, A.; Aziza, E.; Welsh, R. Clinical features and complications of Loeys-Dietz syndrome: A systematic review. Int. J. Cardiol. 2022, 362, 158–167. [Google Scholar] [CrossRef]
- Schoenhoff, F.S.; Alejo, D.E.; Black, J.H.; Crawford, T.C.; Dietz, H.C.; Grimm, J.C.; Magruder, J.T.; Patel, N.D.; Vricella, L.A.; Young, A.; et al. Management of the aortic arch in patients with Loeys–Dietz syndrome. J. Thorac. Cardiovasc. Surg. 2020, 160, 1166–1175. [Google Scholar] [CrossRef]
- Meester, J.A.N.; Verstraeten, A.; Schepers, D.; Alaerts, M.; Van Laer, L.; Loeys, B.L. Differences in manifestations of Marfan syndrome, Ehlers-Danlos syndrome, and Loeys-Dietz syndrome. Ann. Cardiothorac. Surg. 2017, 6, 582–594. [Google Scholar] [CrossRef] [Green Version]
- MacFarlane, E.G.; Parker, S.J.; Shin, J.Y.; Ziegler, S.G.; Creamer, T.J.; Bagirzadeh, R.; Bedja, D.; Chen, Y.; Calderon, J.F.; Weissler, K.; et al. Lineage-specific events underlie aortic root aneurysm pathogenesis in Loeys-Dietz syndrome. J. Clin. Investig. 2019, 129, 659–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Feng, H.; Yang, Y.; Huang, T.; Qiu, P.; Zhang, C.; Olsen, T.R.; Zhang, J.; Chen, Y.E.; Mizrak, D.; et al. hiPSC Modeling of Lineage-Specific Smooth Muscle Cell Defects Caused by TGFBR1 A230T Variant, and Its Therapeutic Implications for Loeys-Dietz Syndrome. Circulation 2021, 144, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Mack, C.P. Signaling Mechanisms That Regulate Smooth Muscle Cell Differentiation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 1495–1505. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Zhou, D.; Jiang, L.; Qiu, P.; Milewicz, D.M.; Chen, Y.E.; Yang, B. In Vitro Lineage-Specific Differentiation of Vascular Smooth Muscle Cells in Response to SMAD3 Deficiency: Implications for SMAD3-Related Thoracic Aortic Aneurysm. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1651–1663. [Google Scholar] [CrossRef] [PubMed]
- Bramel, E.E.; Creamer, T.J.; Saqib, M.; Nunez, W.A.C.; Bagirzadeh, R.; Roker, L.A.; Goff, L.A.; MacFarlane, E.G. Postnatal Smad3 Inactivation in Murine Smooth Muscle Cells Elicits a Temporally and Regionally Distinct Transcriptional Response. Front. Cardiovasc. Med. 2022, 9, 826495. [Google Scholar] [CrossRef]
- Milewicz, D.M.; Trybus, K.M.; Guo, D.-C.; Sweeney, H.L.; Regalado, E.; Kamm, K.; Stull, J.T. Altered Smooth Muscle Cell Force Generation as a Driver of Thoracic Aortic Aneurysms and Dissections. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Seike, Y.; Matsuda, H.; Ishibashi-Ueda, H.; Morisaki, H.; Morisaki, T.; Minatoya, K.; Ogino, H. Surgical Outcome and Histological Differences between Individuals with TGFBR1 and TGFBR2 Mutations in Loeys-Dietz Syndrome. Ann. Thorac. Cardiovasc. Surg. 2021, 27, 56–63. [Google Scholar] [CrossRef]
- Jaffe, M.; Sesti, C.; Washington, I.M.; Du, L.; Dronadula, N.; Chin, M.T.; Stolz, D.B.; Davis, E.C.; Dichek, D.A. Transforming Growth Factor-β Signaling in Myogenic Cells Regulates Vascular Morphogenesis, Differentiation, and Matrix Synthesis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, e1–e11. [Google Scholar] [CrossRef] [Green Version]
- Angelov, S.N.; Hu, J.H.; Wei, H.; Airhart, N.; Shi, M.; Dichek, D.A. TGF-β (Transforming Growth Factor-β) Signaling Protects the Thoracic and Abdominal Aorta From Angiotensin II-Induced Pathology by Distinct Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2102–2113. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Q.; Jiao, Y.; Qin, L.; Ali, R.; Zhou, J.; Ferruzzi, J.; Kim, R.W.; Geirsson, A.; Dietz, H.C.; et al. Tgfbr2 disruption in postnatal smooth muscle impairs aortic wall homeostasis. J. Clin. Investig. 2014, 124, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Gittenbergerdegroot, A.; Azhar, M.; Molin, D. Transforming Growth Factor β–SMAD2 Signaling and Aortic Arch Development. Trends Cardiovasc. Med. 2006, 16, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Molin, D.G.; Poelmann, R.E.; DeRuiter, M.C.; Azhar, M.; Doetschman, T.; Groot, A.C.G.-D. Transforming Growth Factor β–SMAD2 Signaling Regulates Aortic Arch Innervation and Development. Circ. Res. 2004, 95, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Langlois, D.; Hneino, M.; Bouazza, L.; Parlakian, A.; Sasaki, T.; Bricca, G.; Li, J.Y. Conditional inactivation of TGF-β type II receptor in smooth muscle cells and epicardium causes lethal aortic and cardiac defects. Transgenic Res. 2010, 19, 1069–1082. [Google Scholar] [CrossRef] [PubMed]
- Wurdak, H.; Ittner, L.M.; Lang, K.S.; Leveen, P.; Suter, U.; Fischer, J.A.; Karlsson, S.; Born, W.; Sommer, L. Inactivation of TGFβ signaling in neural crest stem cells leads to multiple defects reminiscent of DiGeorge syndrome. Genes Dev. 2005, 19, 530–535. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, B.; Ito, Y.; Makita, T.; Sasaki, T.; Chai, Y.; Sucov, H.M. Cardiovascular malformations with normal smooth muscle differentiation in neural crest-specific type II TGFβ receptor (Tgfbr2) mutant mice. Dev. Biol. 2006, 289, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Angelov, S.; Yildirim, I.A.; Wei, H.; Hu, J.H.; Majesky, M.W.; Brozovich, F.V.; Kim, F.; Dichek, D.A. Loss of Transforming Growth Factor Beta Signaling in Aortic Smooth Muscle Cells Causes Endothelial Dysfunction and Aortic Hypercontractility. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 1956–1971. [Google Scholar] [CrossRef]
- Xie, W.-B.; Li, Z.; Shi, N.; Guo, X.; Tang, J.; Ju, W.; Han, J.; Liu, T.; Bottinger, E.P.; Chai, Y.; et al. Smad2 and Myocardin-Related Transcription Factor B Cooperatively Regulate Vascular Smooth Muscle Differentiation From Neural Crest Cells. Circ. Res. 2013, 113, e76–e86. [Google Scholar] [CrossRef]
- Huang, W.-Y.; Xie, W.; Guo, X.; Li, F.; Jose, P.A.; Chen, S.-Y.; Jiang, H.; Chen, Y.; Yu, T.; Zhao, X.; et al. Smad2 and PEA3 cooperatively regulate transcription of response gene to complement 32 in TGF-β-induced smooth muscle cell differentiation of neural crest cells. Am. J. Physiol. Cell Physiol. 2011, 301, C499–C506. [Google Scholar] [CrossRef]
- Inamoto, S.; Kwartler, C.S.; Lafont, A.L.; Liang, Y.Y.; Fadulu, V.T.; Duraisamy, S.; Willing, M.; Estrera, A.; Safi, H.; Hannibal, M.C.; et al. TGFBR2 mutations alter smooth muscle cell phenotype and predispose to thoracic aortic aneurysms and dissections. Cardiovasc. Res. 2010, 88, 520–529. [Google Scholar] [CrossRef] [Green Version]
- Ferruzzi, J.; Murtada, S.-I.; Li, G.; Jiao, Y.; Uman, S.; Ting, M.Y.L.; Tellides, G.; Humphrey, J.D. Pharmacologically Improved Contractility Protects against Aortic Dissection in Mice with Disrupted Transforming Growth Factor-β Signaling Despite Compromised Extracellular Matrix Properties. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.Y.; Angelov, S.N.; Zhu, J.; Bi, L.; Sanford, N.; Yildirim, I.A.; Dichek, D.A. Blockade of TGF-β (Transforming Growth Factor Beta) Signaling by Deletion of Tgfbr2 in Smooth Muscle Cells of 11-Month-Old Mice Alters Aortic Structure and Causes Vasomotor Dysfunction—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 764–771. [Google Scholar] [CrossRef]
- Krishnamurthy, V.K.; Evans, A.N.; Wansapura, J.P.; Osinska, H.; Maddy, K.E.; Biechler, S.V.; Narmoneva, D.A.; Goodwin, R.L.; Hinton, R.B. Asymmetric Cell–Matrix and Biomechanical Abnormalities in Elastin Insufficiency Induced Aortopathy. Ann. Biomed. Eng. 2014, 42, 2014–2028. [Google Scholar] [CrossRef]
- Regalado, E.S.; Morris, S.A.; Braverman, A.C.; Hostetler, E.M.; De Backer, J.; Li, R.; Pyeritz, R.E.; Yetman, A.T.; Cervi, E.; Shalhub, S.; et al. Comparative Risks of Initial Aortic Events Associated with Genetic Thoracic Aortic Disease. J. Am. Coll. Cardiol. 2022, 80, 857–869. [Google Scholar] [CrossRef]
- Safar, M.E.; Boudier, H.S. Vascular Development, Pulse Pressure, and the Mechanisms of Hypertension. Hypertension 2005, 46, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Fhayli, W.; Boyer, M.; Ghandour, Z.; Jacob, M.; Andrieu, J.; Starcher, B.; Estève, E.; Faury, G. Chronic administration of minoxidil protects elastic fibers and stimulates their neosynthesis with improvement of the aorta mechanics in mice. Cell. Signal. 2019, 62, 109333. [Google Scholar] [CrossRef]
- Pannu, H.; Fadulu, V.T.; Chang, J.; Lafont, A.; Hasham, S.N.; Sparks, E.; Giampietro, P.F.; Zaleski, C.; Estrera, A.L.; Safi, H.J.; et al. Mutations in Transforming Growth Factor-β Receptor Type II Cause Familial Thoracic Aortic Aneurysms and Dissections. Circulation 2005, 112, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Tran-Fadulu, V.; Pannu, H.; Kim, D.H.; Vick, G.W., 3rd; Lonsford, C.M.; Lafont, A.L.; Boccalandro, C.; Smart, S.; Peterson, K.L.; Hain, J.Z.; et al. Analysis of multigenerational families with thoracic aortic aneurysms and dissections due to TGFBR1 or TGFBR2 mutations. J. Med. Genet. 2009, 46, 607–613. [Google Scholar] [CrossRef]
- Jondeau, G.; Ropers, J.; Regalado, E.; Braverman, A.; Evangelista, A.; Teixedo, G.; De Backer, J.; Muiño-Mosquera, L.; Naudion, S.; Zordan, C.; et al. International Registry of Patients Carrying TGFBR1 or TGFBR2 Mutations: Results of the MAC (Montalcino Aortic Consortium). Circ. Cardiovasc. Genet. 2016, 9, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Robertson, I.; Jensen, S.; Handford, P. TB domain proteins: Evolutionary insights into the multifaceted roles of fibrillins and LTBPs. Biochem. J. 2011, 433, 263–276. [Google Scholar] [CrossRef] [Green Version]
- Bunton, T.E.; Biery, N.J.; Myers, L.; Gayraud, B.; Ramirez, F.; Dietz, H.C. Phenotypic Alteration of Vascular Smooth Muscle Cells Precedes Elastolysis in a Mouse Model of Marfan Syndrome. Circ. Res. 2001, 88, 37–43. [Google Scholar] [CrossRef]
- Iosef, C.; Pedroza, A.J.; Cui, J.Z.; Dalal, A.R.; Arakawa, M.; Tashima, Y.; Koyano, T.K.; Burdon, G.; Churovich, S.M.P.; Orrick, J.O.; et al. Quantitative proteomics reveal lineage-specific protein profiles in iPSC-derived Marfan syndrome smooth muscle cells. Sci. Rep. 2020, 10, 20392. [Google Scholar] [CrossRef] [PubMed]
- Elango, J.; Hou, C.; Bao, B.; Wang, S.; de Val, J.E.M.S.; Wenhui, W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function. Polymers 2022, 14, 876. [Google Scholar] [CrossRef] [PubMed]
- Murphy-Ullrich, J.E.; Suto, M.J. Thrombospondin-1 regulation of latent TGF-β activation: A therapeutic target for fibrotic disease. Matrix Biol. 2018, 68–69, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Yamashiro, Y.; Thang, B.Q.; Shin, S.J.; Lino, C.A.; Nakamura, T.; Kim, J.; Sugiyama, K.; Tokunaga, C.; Sakamoto, H.; Osaka, M.; et al. Role of Thrombospondin-1 in Mechanotransduction and Development of Thoracic Aortic Aneurysm in Mouse and Humans. Circ. Res. 2018, 123, 660–672. [Google Scholar] [CrossRef]
- Liu, Z.; Morgan, S.; Ren, J.; Wang, Q.; Annis, D.S.; Mosher, D.F.; Zhang, J.; Sorenson, C.M.; Sheibani, N.; Liu, B.; et al. Thrombospondin-1 (TSP1) Contributes to the Development of Vascular Inflammation by Regulating Monocytic Cell Motility in Mouse Models of Abdominal Aortic Aneurysm. Circ. Res. 2015, 117, 129–141. [Google Scholar] [CrossRef]
- Li, H.; Xu, H.; Wen, H.; Wang, H.; Zhao, R.; Sun, Y.; Bai, C.; Ping, J.; Song, L.; Luo, M.; et al. Lysyl hydroxylase 1 (LH1) deficiency promotes angiotensin II (Ang II)-induced dissecting abdominal aortic aneurysm. Theranostics 2021, 11, 9587–9604. [Google Scholar] [CrossRef]
- Yang, H.; Zhou, T.; Sorenson, C.M.; Sheibani, N.; Liu, B. Myeloid-Derived TSP1 (Thrombospondin-1) Contributes to Abdominal Aortic Aneurysm Through Suppressing Tissue Inhibitor of Metalloproteinases-1. Arterioscler. Thromb. Vasc. Biol. 2020, 40, e350–e366. [Google Scholar] [CrossRef]
- Pedroza, A.J.; Tashima, Y.; Shad, R.; Cheng, P.; Wirka, R.; Churovich, S.; Nakamura, K.; Yokoyama, N.; Cui, J.Z.; Iosef, C.; et al. Single-Cell Transcriptomic Profiling of Vascular Smooth Muscle Cell Phenotype Modulation in Marfan Syndrome Aortic Aneurysm. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2195–2211. [Google Scholar] [CrossRef]
- Pedroza, A.J.; Dalal, A.R.; Shad, R.; Yokoyama, N.; Nakamura, K.; Cheng, P.; Wirka, R.C.; Mitchel, O.; Baiocchi, M.; Hiesinger, W.; et al. Embryologic Origin Influences Smooth Muscle Cell Phenotypic Modulation Signatures in Murine Marfan Syndrome Aortic Aneurysm. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 1154–1168. [Google Scholar] [CrossRef]
- Wanga, S.; Hibender, S.; Ridwan, Y.; van Roomen, C.; Vos, M.; van der Made, I.; van Vliet, N.; Franken, R.; van Riel, L.A.; Groenink, M.; et al. Aortic microcalcification is associated with elastin fragmentation in Marfan syndrome. J. Pathol. 2017, 243, 294–306. [Google Scholar] [CrossRef] [Green Version]
- Crosas-Molist, E.; Meirelles, T.; López-Luque, J.; Serra-Peinado, C.; Selva, J.; Caja, L.; Del Blanco, D.G.; Uriarte, J.J.; Bertran, E.; Mendizábal, Y.; et al. Vascular Smooth Muscle Cell Phenotypic Changes in Patients with Marfan Syndrome. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 960–972. [Google Scholar] [CrossRef] [Green Version]
- Dawson, A.; Li, Y.; Li, Y.; Ren, P.; Vasquez, H.G.; Zhang, C.; Rebello, K.R.; Ageedi, W.; Azares, A.R.; Mattar, A.B.; et al. Single-Cell Analysis of Aneurysmal Aortic Tissue in Patients with Marfan Syndrome Reveals Dysfunctional TGF-β Signaling. Genes 2021, 13, 95. [Google Scholar] [CrossRef]
- Pyeritz, R.E. Heritable thoracic aortic disorders. Curr. Opin. Cardiol. 2014, 29, 97–102. [Google Scholar] [CrossRef]
- Robertson, E.N.; Hambly, B.D.; Jeremy, R.W. Thoracic aortic dissection and heritability: Forensic implications. Forensic Sci. Med. Pathol. 2016, 12, 366–368. [Google Scholar] [CrossRef]
- Lu, H.; Daugherty, A. Aortic Aneurysms. Arterioscler. Thromb. Vasc. Biol. 2017, 37, e59–e65. [Google Scholar] [CrossRef] [Green Version]
- Kan, H.; Zhang, K.; Mao, A.; Geng, L.; Gao, M.; Feng, L.; You, Q.; Ma, X. Single-cell transcriptome analysis reveals cellular heterogeneity in the ascending aortas of normal and high-fat diet-fed mice. Exp. Mol. Med. 2021, 53, 1379–1389. [Google Scholar] [CrossRef]
- Zhang, K.; Kan, H.; Mao, A.; Geng, L.; Ma, X. Single-cell analysis of salt-induced hypertensive mouse aortae reveals cellular heterogeneity and state changes. Exp. Mol. Med. 2021, 53, 1866–1876. [Google Scholar] [CrossRef]
- Salmon, M.; Johnston, W.F.; Woo, A.; Pope, N.H.; Su, G.; Upchurch, G.R., Jr.; Owens, G.K.; Ailawadi, G. KLF4 Regulates Abdominal Aortic Aneurysm Morphology and Deletion Attenuates Aneurysm Formation. Circulation 2013, 128, S163–S174. [Google Scholar] [CrossRef] [Green Version]
- Shankman, L.S.; Gomez, D.; Cherepanova, O.A.; Salmon, M.; Alencar, G.F.; Haskins, R.M.; Swiatlowska, P.; Newman, A.A.C.; Greene, E.S.; Straub, A.C.; et al. KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nat. Med. 2015, 21, 628–637. [Google Scholar] [CrossRef] [Green Version]
- Cheng, J.; Gu, W.; Lan, T.; Deng, J.; Ni, Z.; Zhang, Z.; Hu, Y.; Sun, X.; Yang, Y.; Xu, Q. Single-cell RNA sequencing reveals cell type- and artery type-specific vascular remodelling in male spontaneously hypertensive rats. Cardiovasc. Res. 2021, 117, 1202–1216. [Google Scholar] [CrossRef]
- Li, Y.; Ren, P.; Dawson, A.; Vasquez, H.G.; Ageedi, W.; Zhang, C.; Luo, W.; Chen, R.; Li, Y.; Kim, S.; et al. Single-Cell Transcriptome Analysis Reveals Dynamic Cell Populations and Differential Gene Expression Patterns in Control and Aneurysmal Human Aortic Tissue. Circulation 2020, 142, 1374–1388. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Lu, Z.; Gu, R.; Xuan, X.; Zhang, R.; Hu, J.; Dong, H. Deciphering the Intercellular Communication Between Immune Cells and Altered Vascular Smooth Muscle Cell Phenotypes in Aortic Aneurysm From Single-Cell Transcriptome Data. Front. Cardiovasc. Med. 2022, 9, 936287. [Google Scholar] [CrossRef]
- Song, W.; Qin, L.; Chen, Y.; Chen, J.; Wei, L. Single-cell transcriptome analysis identifies Versican(+) myofibroblast as a hallmark for thoracic aortic aneurysm marked by activation of PI3K-AKT signaling pathway. Biochem. Biophys. Res. Commun. 2022, 643, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, Y.; Chakraborty, A.; Li, Y.; Rebello, K.R.; Ren, P.; Luo, W.; Zhang, L.; Lu, H.S.; Cassis, L.A.; et al. Aortic Stress Activates an Adaptive Program in Thoracic Aortic Smooth Muscle Cells That Maintains Aortic Strength and Protects against Aneurysm and Dissection in Mice. Arterioscler. Thromb. Vasc. Biol. 2023, 43, 234–252. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-Y.; Qin, L.; Li, G.; Malagon-Lopez, J.; Wang, Z.; Bergaya, S.; Gujja, S.; Caulk, A.W.; Murtada, S.-I.; Zhang, X.; et al. Smooth Muscle Cell Reprogramming in Aortic Aneurysms. Cell Stem Cell 2020, 26, 542–557.e511. [Google Scholar] [CrossRef] [PubMed]
- Clément, M.; Chappell, J.; Raffort, J.; Lareyre, F.; Vandestienne, M.; Taylor, A.L.; Finigan, A.; Harrison, J.; Bennett, M.R.; Bruneval, P.; et al. Vascular Smooth Muscle Cell Plasticity and Autophagy in Dissecting Aortic Aneurysms. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1149–1159. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-B.; Jin, H.-Z.; Zhao, J.-L.; Wang, C.; Ma, W.-R.; Xing, J.; Zhang, X.-B.; Zhang, Y.-Y.; Dai, H.-D.; Zhao, N.-S.; et al. Single-cell transcriptomic analysis reveals differential cell subpopulations and distinct phenotype transition in normal and dissected ascending aorta. Mol. Med. 2022, 28, 158. [Google Scholar] [CrossRef]
- Zhao, G.; Lu, H.; Chang, Z.; Zhao, Y.; Zhu, T.; Chang, L.; Guo, Y.; Garcia-Barrio, M.T.; Chen, Y.E.; Zhang, J. Single-cell RNA sequencing reveals the cellular heterogeneity of aneurysmal infrarenal abdominal aorta. Cardiovasc. Res. 2020, 117, 1402–1416. [Google Scholar] [CrossRef]
- Chou, E.L.; Chaffin, M.; Simonson, B.; Pirruccello, J.P.; Akkad, A.-D.; Nekoui, M.; Cardenas, C.L.L.; Bedi, K.C.; Nash, C.; Juric, D.; et al. Aortic Cellular Diversity and Quantitative Genome-Wide Association Study Trait Prioritization Through Single-Nuclear RNA Sequencing of the Aneurysmal Human Aorta. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 1355–1374. [Google Scholar] [CrossRef]
- Wight, T.N.; Kang, I.; Evanko, S.P.; Harten, I.A.; Chang, M.Y.; Pearce, O.M.T.; Allen, C.E.; Frevert, C.W. Versican—A Critical Extracellular Matrix Regulator of Immunity and Inflammation. Front. Immunol. 2020, 11, 512. [Google Scholar] [CrossRef] [Green Version]
- Wirka, R.C.; Wagh, D.; Paik, D.T.; Pjanic, M.; Nguyen, T.; Miller, C.L.; Kundu, R.; Nagao, M.; Coller, J.; Koyano, T.K.; et al. Atheroprotective roles of smooth muscle cell phenotypic modulation and the TCF21 disease gene as revealed by single-cell analysis. Nat. Med. 2019, 25, 1280–1289. [Google Scholar] [CrossRef]
- Pan, H.; Xue, C.; Auerbach, B.J.; Fan, J.; Bashore, A.C.; Cui, J.; Yang, D.Y.; Trignano, S.B.; Liu, W.; Shi, J.; et al. Single-Cell Genomics Reveals a Novel Cell State During Smooth Muscle Cell Phenotypic Switching and Potential Therapeutic Targets for Atherosclerosis in Mouse and Human. Circulation 2020, 142, 2060–2075. [Google Scholar] [CrossRef]
- Pedroza, A.J.; Shad, R.; Dalal, A.R.; Yokoyama, N.; Nakamura, K.; Hiesinger, W.; Fischbein, M.P. Acute Induced Pressure Overload Rapidly Incites Thoracic Aortic Aneurysmal Smooth Muscle Cell Phenotype. Hypertension 2022, 79, e86–e89. [Google Scholar] [CrossRef]
- Ma, Q.; Yang, Q.; Xu, J.; Zhang, X.; Kim, D.; Liu, Z.; Da, Q.; Mao, X.; Zhou, Y.; Cai, Y.; et al. ATIC-Associated De Novo Purine Synthesis Is Critically Involved in Proliferative Arterial Disease. Circulation 2022, 146, 1444–1460. [Google Scholar] [CrossRef]
- Majesky, M.W.; Horita, H.; Ostriker, A.; Lu, S.; Regan, J.N.; Bagchi, A.; Dong, X.R.; Poczobutt, J.; Nemenoff, R.A.; Weiser-Evans, M.C.; et al. Differentiated Smooth Muscle Cells Generate a Subpopulation of Resident Vascular Progenitor Cells in the Adventitia Regulated by Klf4. Circ. Res. 2017, 120, 296–311. [Google Scholar] [CrossRef]
- Shikatani, E.A.; Chandy, M.; Besla, R.; Li, C.C.; Momen, A.; El-Mounayri, O.; Robbins, C.S.; Husain, M. c-Myb Regulates Proliferation and Differentiation of Adventitial Sca1 + Vascular Smooth Muscle Cell Progenitors by Transactivation of Myocardin. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1367–1376. [Google Scholar] [CrossRef] [Green Version]
- Worssam, M.D.; Lambert, J.; Oc, S.; Taylor, J.C.K.; Taylor, A.L.; Dobnikar, L.; Chappell, J.; Harman, J.L.; Figg, N.L.; Finigan, A.; et al. Cellular mechanisms of oligoclonal vascular smooth muscle cell expansion in cardiovascular disease. Cardiovasc. Res. 2022, 119, 1279–1294. [Google Scholar] [CrossRef]
- Chappell, J.; Harman, J.L.; Narasimhan, V.M.; Yu, H.; Foote, K.; Simons, B.D.; Bennett, M.R.; Jørgensen, H.F. Extensive Proliferation of a Subset of Differentiated, yet Plastic, Medial Vascular Smooth Muscle Cells Contributes to Neointimal Formation in Mouse Injury and Atherosclerosis Models. Circ. Res. 2016, 119, 1313–1323. [Google Scholar] [CrossRef]
- Feil, S.; Fehrenbacher, B.; Lukowski, R.; Essmann, F.; Schulze-Osthoff, K.; Schaller, M.; Feil, R. Transdifferentiation of Vascular Smooth Muscle Cells to Macrophage-Like Cells During Atherogenesis. Circ. Res. 2014, 115, 662–667. [Google Scholar] [CrossRef]
- Harman, J.L.; Jørgensen, H.F. The role of smooth muscle cells in plaque stability: Therapeutic targeting potential. Br. J. Pharmacol. 2019, 176, 3741–3753. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, H.; Zhang, Q.; Han, X.; Zhang, Z.; Shen, L.; Wang, L.; Lui, K.O.; He, B.; Zhou, B. Smooth muscle-derived macrophage-like cells contribute to multiple cell lineages in the atherosclerotic plaque. Cell Discov. 2021, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Leone, O.; Corsini, A.; Pacini, D.; Corti, B.; Lorenzini, M.; Laus, V.; Foà, A.; Reggiani, M.L.B.; Di Marco, L.; Rapezzi, C. The complex interplay among atherosclerosis, inflammation, and degeneration in ascending thoracic aortic aneurysms. J. Thorac. Cardiovasc. Surg. 2019, 160, 1434–1443.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesi, G.; Anichini, C.; Tozzini, S.; Boddi, V.; Calamai, G.; Gori, F. Pathology of the thoracic aorta: A morphologic review of 338 surgical specimens over a 7-year period. Cardiovasc. Pathol. 2009, 18, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Negedu, M.N.; Duckworth, C.A.; Yu, L.-G. Galectin-2 in Health and Diseases. Int. J. Mol. Sci. 2022, 24, 341. [Google Scholar] [CrossRef]
- Minas, T.Z.; Candia, J.; Dorsey, T.H.; Baker, F.; Tang, W.; Kiely, M.; Smith, C.J.; Zhang, A.L.; Jordan, S.V.; Obadi, O.M.; et al. Serum proteomics links suppression of tumor immunity to ancestry and lethal prostate cancer. Nat. Commun. 2022, 13, 1759. [Google Scholar] [CrossRef]
- Huang, Y.; Wang, H.-C.; Zhao, J.; Wu, M.-H.; Shih, T.-C. Immunosuppressive Roles of Galectin-1 in the Tumor Microenvironment. Biomolecules 2021, 11, 1398. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, X.; Guo, Y.; Liu, Z.; Wei, S.; Yuan, Q.; Shang, H.; Sang, W.; Cui, S.; Xu, T.; et al. Macrophage ALDH2 (Aldehyde Dehydrogenase 2) Stabilizing Rac2 Is Required for Efferocytosis Internalization and Reduction of Atherosclerosis Development. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 700–716. [Google Scholar] [CrossRef]
- Kojima, Y.; Werner, N.; Ye, J.; Nanda, V.; Tsao, N.; Wang, Y.; Flores, A.M.; Miller, C.L.; Weissman, I.; Deng, H.; et al. Proefferocytic Therapy Promotes Transforming Growth Factor-β Signaling and Prevents Aneurysm Formation. Circulation 2018, 137, 750–753. [Google Scholar] [CrossRef]
- López-Otín, C.; Kroemer, G. Hallmarks of Health. Cell 2021, 184, 33–63. [Google Scholar] [CrossRef]
- Luo, W.; Wang, Y.; Zhang, L.; Ren, P.; Zhang, C.; Li, Y.; Azares, A.R.; Zhang, M.; Guo, J.; Ghaghada, K.B.; et al. Critical Role of Cytosolic DNA and Its Sensing Adaptor STING in Aortic Degeneration, Dissection, and Rupture. Circulation 2020, 141, 42–66. [Google Scholar] [CrossRef]
- Saito, T.; Hasegawa, Y.; Ishigaki, Y.; Yamada, T.; Gao, J.; Imai, J.; Uno, K.; Kaneko, K.; Ogihara, T.; Shimosawa, T.; et al. Importance of endothelial NF-κB signalling in vascular remodelling and aortic aneurysm formation. Cardiovasc. Res. 2013, 97, 106–114. [Google Scholar] [CrossRef]
- Rong, J.X.; Shapiro, M.; Trogan, E.; Fisher, E.A. Transdifferentiation of mouse aortic smooth muscle cells to a macrophage-like state after cholesterol loading. Proc. Natl. Acad. Sci. USA 2003, 100, 13531–13536. [Google Scholar] [CrossRef]
- Vengrenyuk, Y.; Nishi, H.; Long, X.; Ouimet, M.; Savji, N.; Martinez, F.O.; Cassella, C.P.; Moore, K.J.; Ramsey, S.A.; Miano, J.M.; et al. Cholesterol Loading Reprograms the MicroRNA-143/145–Myocardin Axis to Convert Aortic Smooth Muscle Cells to a Dysfunctional Macrophage-Like Phenotype. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Sanz, L.; Bernal, S.; Jimenez-Castilla, L.; Prieto, I.; La Manna, S.; Gomez-Lopez, S.; Blanco-Colio, L.M.; Egido, J.; Martin-Ventura, J.L.; Gomez-Guerrero, C. Fcγ receptor activation mediates vascular inflammation and abdominal aortic aneurysm development. Clin. Transl. Med. 2021, 11, e463. [Google Scholar] [CrossRef]
- Filiberto, A.C.; Spinosa, M.D.; Elder, C.T.; Su, G.; Leroy, V.; Ladd, Z.; Lu, G.; Mehaffey, J.H.; Salmon, M.D.; Hawkins, R.B.; et al. Endothelial pannexin-1 channels modulate macrophage and smooth muscle cell activation in abdominal aortic aneurysm formation. Nat. Commun. 2022, 13, 1521. [Google Scholar] [CrossRef]
- Shao, F.; Miao, Y.; Zhang, Y.; Han, L.; Ma, X.; Deng, J.; Jiang, C.; Kong, W.; Xu, Q.; Feng, J.; et al. B cell-derived anti-beta 2 glycoprotein I antibody contributes to hyperhomocysteinaemia-aggravated abdominal aortic aneurysm. Cardiovasc. Res. 2020, 116, 1897–1909. [Google Scholar] [CrossRef]
- Kurz, J.; Weiss, A.-C.; Lüdtke, T.H.-W.; Deuper, L.; Trowe, M.-O.; Thiesler, H.; Hildebrandt, H.; Heineke, J.; Duncan, S.A.; Kispert, A. GATA6 is a crucial factor for Myocd expression in the visceral smooth muscle cell differentiation program of the murine ureter. Development 2022, 149, dev200522. [Google Scholar] [CrossRef]
- Zhuang, T.; Liu, J.; Chen, X.; Pi, J.; Kuang, Y.; Wang, Y.; Tomlinson, B.; Chan, P.; Zhang, Q.; Li, Y.; et al. Cell-Specific Effects of GATA (GATA Zinc Finger Transcription Factor Family)-6 in Vascular Smooth Muscle and Endothelial Cells on Vascular Injury Neointimal Formation. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 888–901. [Google Scholar] [CrossRef] [Green Version]
- Alajbegovic, A.; Daoud, F.; Ali, N.; Kawka, K.; Holmberg, J.; Albinsson, S. Transcription factor GATA6 promotes migration of human coronary artery smooth muscle cells in vitro. Front. Physiol. 2022, 13, 1054819. [Google Scholar] [CrossRef]
- Jiao, H.; Walczak, B.E.; Lee, M.-S.; Lemieux, M.E.; Li, W.-J. GATA6 regulates aging of human mesenchymal stem/stromal cells. Stem Cells 2021, 39, 62–77. [Google Scholar] [CrossRef]
- Pabis, K.; Chiari, Y.; Sala, C.; Straka, E.; Giacconi, R.; Provinciali, M.; Li, X.; Brown-Borg, H.; Nowikovsky, K.; Valencak, T.G.; et al. Elevated metallothionein expression in long-lived species mediates the influence of cadmium accumulation on aging. Geroscience 2021, 43, 1975–1993. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wen, H.; Lv, J.; Luan, B.; Meng, J.; Gong, S.; Wen, J.; Xin, S. Therapeutic efficacy of mesenchymal stem cells for abdominal aortic aneurysm: A meta-analysis of preclinical studies. Stem Cell Res. Ther. 2022, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Bravo-San Pedro, J.M.; Kepp, O.; Kroemer, G. Regulated cell death and adaptive stress responses. Cell. Mol. Life Sci. 2016, 73, 2405–2410. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, A.; Kwartler, C.S.; Kaw, K.; Li, Y.; Kaw, A.; Chen, J.; LeMaire, S.A.; Shen, Y.H.; Milewicz, D.M. Cholesterol-Induced Phenotypic Modulation of Smooth Muscle Cells to Macrophage/Fibroblast–like Cells Is Driven by an Unfolded Protein Response. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 302–316. [Google Scholar] [CrossRef] [PubMed]
- Hamczyk, M.R.; Villa-Bellosta, R.; Quesada, V.; Gonzalo, P.; Vidak, S.; Nevado, R.M.; Andrés-Manzano, M.J.; Misteli, T.; López-Otín, C.; Andrés, V. Progerin accelerates atherosclerosis by inducing endoplasmic reticulum stress in vascular smooth muscle cells. EMBO Mol. Med. 2019, 11, e9736. [Google Scholar] [CrossRef]
- Oller, J.; Gabandé-Rodríguez, E.; Ruiz-Rodríguez, M.J.; Desdín-Micó, G.; Aranda, J.F.; Rodrigues-Diez, R.; Ballesteros-Martínez, C.; Blanco, E.M.; Roldan-Montero, R.; Acuña, P.; et al. Extracellular Tuning of Mitochondrial Respiration Leads to Aortic Aneurysm. Circulation 2021, 143, 2091–2109. [Google Scholar] [CrossRef]
- Asano, K.; Cantalupo, A.; Sedes, L.; Ramirez, F. Pathophysiology and Therapeutics of Thoracic Aortic Aneurysm in Marfan Syndrome. Biomolecules 2022, 12, 128. [Google Scholar] [CrossRef]
- Sawada, H.; Katsumata, Y.; Higashi, H.; Zhang, C.; Li, Y.; Morgan, S.; Lee, L.H.; Singh, S.A.; Chen, J.Z.; Franklin, M.K.; et al. Second Heart Field–Derived Cells Contribute to Angiotensin II–Mediated Ascending Aortopathies. Circulation 2022, 145, 987–1001. [Google Scholar] [CrossRef]
- D’amico, F.; Doldo, E.; Pisano, C.; Scioli, M.G.; Centofanti, F.; Proietti, G.; Falconi, M.; Sangiuolo, F.; Ferlosio, A.; Ruvolo, G.; et al. Specific miRNA and Gene Deregulation Characterize the Increased Angiogenic Remodeling of Thoracic Aneurysmatic Aortopathy in Marfan Syndrome. Int. J. Mol. Sci. 2020, 21, 6886. [Google Scholar] [CrossRef]
- Si, X.; Chen, Q.; Zhang, J.; Zhou, W.; Chen, L.; Chen, J.; Deng, N.; Li, W.; Liu, D.; Wang, L.; et al. MicroRNA-23b prevents aortic aneurysm formation by inhibiting smooth muscle cell phenotypic switching via FoxO4 suppression. Life Sci. 2022, 288, 119092. [Google Scholar] [CrossRef]
- Yang, K.; Ren, J.; Li, X.; Wang, Z.; Xue, L.; Cui, S.; Sang, W.; Xu, T.; Zhang, J.; Yu, J.; et al. Prevention of aortic dissection and aneurysm via an ALDH2-mediated switch in vascular smooth muscle cell phenotype. Eur. Heart J. 2020, 41, 2442–2453. [Google Scholar] [CrossRef]
- Lino Cardenas, C.L.; Kessinger, C.W.; Cheng, Y.; Macdonald, C.; MacGillivray, T.; Ghoshhajra, B.; Huleihel, L.; Nuri, S.; Yeri, A.S.; Jaffer, F.A.; et al. An HDAC9-MALAT1-BRG1 complex mediates smooth muscle dysfunction in thoracic aortic aneurysm. Nat. Commun. 2018, 9, 1009. [Google Scholar] [CrossRef] [Green Version]
- Zalewski, D.P.; Ruszel, K.P.; Stępniewski, A.; Gałkowski, D.; Feldo, M.; Kocki, J.; Bogucka-Kocka, A. miRNA Regulatory Networks Associated with Peripheral Vascular Diseases. J. Clin. Med. 2022, 11, 3470. [Google Scholar] [CrossRef]
- Krishna, S.M.; Morton, S.K.; Li, J.; Golledge, J. Risk Factors and Mouse Models of Abdominal Aortic Aneurysm Rupture. Int. J. Mol. Sci. 2020, 21, 7250. [Google Scholar] [CrossRef]
- Lu, G.; Su, G.; Davis, J.P.; Schaheen, B.; Downs, E.; Roy, R.J.; Ailawadi, G.; Upchurch, G.R. A novel chronic advanced stage abdominal aortic aneurysm murine model. J. Vasc. Surg. 2017, 66, 232–242.e4. [Google Scholar] [CrossRef] [Green Version]
- Golledge, J.; Krishna, S.M.; Wang, Y. Mouse models for abdominal aortic aneurysm. Br. J. Pharmacol. 2020, 179, 792–810. [Google Scholar] [CrossRef]
- Sulé-Suso, J.; Forster, A.; Zholobenko, V.; Stone, N.; El Haj, A. Effects of CaCl2 and MgCl2 on Fourier Transform Infrared Spectra of Lung Cancer Cells. Appl. Spectrosc. 2004, 58, 61–67. [Google Scholar] [CrossRef]
- Sundararaman, S.S.; van der Vorst, E.P.C. Calcium-Sensing Receptor (CaSR), Its Impact on Inflammation and the Consequences on Cardiovascular Health. Int. J. Mol. Sci. 2021, 22, 2478. [Google Scholar] [CrossRef]
- Wagenseil, J.E.; Mecham, R.P.; Tellides, G.; Staiculescu, M.C.; Cocciolone, A.J.; Procknow, J.D.; Kim, J.; Hawes, J.Z.; Johnson, E.O.; Murshed, M.; et al. Vascular Extracellular Matrix and Arterial Mechanics. Physiol. Rev. 2009, 89, 957–989. [Google Scholar] [CrossRef] [Green Version]
- Nolasco, P.; Fernandes, C.G.; Ribeiro-Silva, J.C.; Oliveira, P.V.; Sacrini, M.; de Brito, I.V.; DE Bessa, T.; Pereira, L.V.; Tanaka, L.Y.; Alencar, A.; et al. Impaired vascular smooth muscle cell force-generating capacity and phenotypic deregulation in Marfan Syndrome mice. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2019, 1866, 165587. [Google Scholar] [CrossRef]
- Wong, L.; Kumar, A.; Gabela-Zuniga, B.; Chua, J.; Singh, G.; Happe, C.L.; Engler, A.J.; Fan, Y.; McCloskey, K.E. Substrate stiffness directs diverging vascular fates. Acta Biomater. 2019, 96, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Schnellmann, R.; Ntekoumes, D.; Choudhury, M.I.; Sun, S.; Wei, Z.; Gerecht, S. Stiffening Matrix Induces Age-Mediated Microvascular Phenotype Through Increased Cell Contractility and Destabilization of Adherens Junctions. Adv. Sci. 2022, 9, e2201483. [Google Scholar] [CrossRef] [PubMed]
- Talwar, S.; Kant, A.; Xu, T.; Shenoy, V.B.; Assoian, R.K. Mechanosensitive smooth muscle cell phenotypic plasticity emerging from a null state and the balance between Rac and Rho. Cell Rep. 2021, 35, 109019. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, S.-A.; Li, N.; Zhang, T.; Yao, W.; Zhao, H.; Pang, W.; Han, L.; Liu, J.; Zhou, J. Matrix stiffness exacerbates the proinflammatory responses of vascular smooth muscle cell through the DDR1-DNMT1 mechanotransduction axis. Bioact. Mater. 2022, 17, 406–424. [Google Scholar] [CrossRef]
- Qian, W.; Hadi, T.; Silvestro, M.; Ma, X.; Rivera, C.F.; Bajpai, A.; Li, R.; Zhang, Z.; Qu, H.; Tellaoui, R.S.; et al. Microskeletal stiffness promotes aortic aneurysm by sustaining pathological vascular smooth muscle cell mechanosensation via Piezo1. Nat. Commun. 2022, 13, 512. [Google Scholar] [CrossRef]
- Pasta, S.; Agnese, V.; Gallo, A.; Cosentino, F.; Di Giuseppe, M.; Gentile, G.; Raffa, G.M.; Maalouf, J.F.; Michelena, H.I.; Bellavia, D.; et al. Shear Stress and Aortic Strain Associations with Biomarkers of Ascending Thoracic Aortic Aneurysm. Ann. Thorac. Surg. 2020, 110, 1595–1604. [Google Scholar] [CrossRef]
- Gomez, D.; Swiatlowska, P.; Owens, G.K. Epigenetic Control of Smooth Muscle Cell Identity and Lineage Memory. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 2508–2516. [Google Scholar] [CrossRef] [Green Version]
- Gurung, R.; Choong, A.M.; Woo, C.C.; Foo, R.; Sorokin, V. Genetic and Epigenetic Mechanisms Underlying Vascular Smooth Muscle Cell Phenotypic Modulation in Abdominal Aortic Aneurysm. Int. J. Mol. Sci. 2020, 21, 6334. [Google Scholar] [CrossRef]
- Floren, M.; Bonani, W.; Dharmarajan, A.; Motta, A.; Migliaresi, C.; Tan, W. Human mesenchymal stem cells cultured on silk hydrogels with variable stiffness and growth factor differentiate into mature smooth muscle cell phenotype. Acta Biomater. 2016, 31, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Walters, B.; Turner, P.A.; Rolauffs, B.; Hart, M.L.; Stegemann, J.P. Controlled Growth Factor Delivery and Cyclic Stretch Induces a Smooth Muscle Cell-like Phenotype in Adipose-Derived Stem Cells. Cells 2021, 10, 3123. [Google Scholar] [CrossRef]
- Tingting, T.; Wenjing, F.; Qian, Z.; Hengquan, W.; Simin, Z.; Zhisheng, J.; Shunlin, Q. The TGF-β pathway plays a key role in aortic aneurysms. Clin. Chim. Acta 2020, 501, 222–228. [Google Scholar] [CrossRef]
- Li, G.; Wang, M.; Caulk, A.W.; Cilfone, N.A.; Gujja, S.; Qin, L.; Chen, P.-Y.; Chen, Z.; Yousef, S.; Jiao, Y.; et al. Chronic mTOR activation induces a degradative smooth muscle cell phenotype. J. Clin. Investig. 2020, 130, 1233–1251. [Google Scholar] [CrossRef]
- Zhou, Z.; Liu, Y.; Gao, S.; Zhou, M.; Qi, F.; Ding, N.; Zhang, J.; Li, R.; Wang, J.; Shi, J.; et al. Excessive DNA damage mediates ECM degradation via the RBBP8/NOTCH1 pathway in sporadic aortic dissection. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2022, 1868, 166303. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| VSMC Phenotypes | Technique | Tissue | Annotation | Marker Genes | Characteristics |
|---|---|---|---|---|---|
| Secreting VSMC [111] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Fibromyocyte | ACTA2; MYL9; COL1A2; COL8A1 | Share properties of fibroblasts and muscle cells; Upregulate collagen, proteoglycan genes. |
| Secreting VSMC [112] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Secreting VSMCs | ACTA2; MYH11; COL1A1; COL1A2; FABP4 | Significantly increase their percentage in AA lesions. |
| Secreting VSMC [113] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Myofibroblast | ACTA2; TAGLN; DCN; LUM; VCAN; CLU; FN1; LTBP2; CTGF; AEBP1 | Share characteristics of VSMCs and fibroblasts; Characterized by ECM organization, proliferation and migration. |
| Secreting VSMC [114] | scRNA-seq | WT and Yap1f/f; Myh11-CreERT2 mice; Challenged by HFD and AngII | ECM-producing VSMCs | Not discussed | Not discussed |
| Proliferating VSMC [111] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Proliferating VSMC1 | TPM2; MAP1B; MYH11; CCND1; CALD1; MYH10; TPM4; FGF1; SPARC; FTH1 | Upregulate cyclin gene expression; Downregulate cell–cell junction; Express high level of synthetic VSMC marker genes; Express low levels of collagen and proteoglycan genes. |
| Proliferating VSMC2 | GAS6; IGFBP2; MGP; FTH1; SPARC; FGF1; TPM4; MYH10; CCND1; TPM2 | Same as above | |||
| Proliferating VSMC [115] | Lineage tracing | Apoe−/− (Apoe−/−; Myh11CreERT2T2;mT/mGf/f) mice; TGFβR2iSMC (Myh11CreERT2;mT/mGf/f;Tgfbr2f/f) mice; Challenged by HCHFD; | MSC-like VSMCs | CD105; CD73; CD90; CD44; Sca-1 | Monoclonal expansion of lineage-traced VSMCs was found in 9/10 samples. |
| Proliferating VSMC [116] | Lineage tracing | Myh11-CreERt2/Rosa26-Confetti mice; Challenged by AngII and anti-TGF; | / | / | Monochromatic patches of VSMCs were found in the medial layer of 5 out of the 6 animals analyzed. |
| Mesenchymal VSMC [112] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Msenchymal-like VSMCs | ACTA2; MYH11; CD34; PDGFRA | Play a poor role in the communication network with immune cells. |
| Immune-related VSMC [112] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Macrophage-like VSMCs | ACTA2; MYH11; CD14; CD68 | Communicate with all immune cells and sent maximum number of the signaling pathways. |
| T-cell-like VSMCs | ACTA2; MYH11; CD3D; CD3G | Act as a signal sender in the communication network with immune cells. | |||
| Immune-related VSMC [111] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Inflammatory1 | CXCR4; CCL4; CXCL12; TNF; CCL20; IFNG; XCL1; XCL2; LTB; CCL5; TNFSF9; IL32 | Show a T lymphocyte–like gene expression profile. |
| Inflammatory2 | C1QA; C1QB; IL1RN; IL1B; IL18; CCL3; CCL4L2; CCL3L1; CXCL1; CXCL3; CXCL8; CXCL16 | Characterized by expression of macrophage markers. | |||
| Inflammatory3 | IFIT1; IFI6; CXCL9; CXCL10; CCL28; IL15 | Expressed interferon-induced gene. | |||
| Immune-related VSMC [117] | scRNA-seq | 4 healthy controls; 5 diseased aortas from ATAD patients; | Monocyte-like VSMCs | CD93; THBD | Characterized by expression of monocyte markers. |
| Immune-related VSMC [115] | Lineage tracing | Apoe−/− (Apoe−/−; Myh11CreERT2T2;mT/mGf/f) mice; TGFβR2iSMC (Myh11CreERT2;mT/mGf/f;Tgfbr2f/f) mice; Challenged by HCHFD; | Macrophage-like cells | Mac2 | Characterized by expression of macrophage markers. |
| Proinflammatory VSMC [118] | scRNA-seq | C57BL/6J mice; Challenged by elastase; | Inflammatory-like SMCs | Mac2; Sparcl1; Igfbp5; Sncg; Thbs1; Notch3; Cd68; Cxcl1; Cxcl2; Il1r1 | Displayed a pro-inflammatory phenotype; Increased at late stage of AAA development. |
| Senescent VSMC [117] | scRNA-seq | 4 healthy controls; 5 diseased aortas from ATAD patients; | VSMC8 | MT1G; MT1M | Express metallothionein superfamily genes; Have minimum differentiation potential. |
| Senescent VSMC [118] | scRNA-seq | C57BL/6J mice; Challenged by elastase; | Dedifferentiated SMCs | Ifrd1; Klf4; gata6; Mt1; Mt2; Hk2; | Show the lowest expression of contractile markers; Have altered metabolic profile to glycolysis. |
| Stressed VSMC [111] | scRNA-seq | 3 normal controls; 8 aneurysmal aorta samples from ATAA patients; | Stressed VSMCs | FOS; ATF3; JUN; HSPB8; ACTC1; BRD2; KLF10; DUSP1; | Share several features with contractile VSMCs, except the activation of stress response genes. |
| Stressed VSMC [118] | scRNA-seq | C57BL/6J mice; Challenged by elastase | Proliferative VSMCs | Fos; Jun; Klf2; Klf4; gata6; Atf3 | Upregulate expression of genes associated with stress; Gain an inflammation-activated profile. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Cai, Z.; He, B. Smooth Muscle Heterogeneity and Plasticity in Health and Aortic Aneurysmal Disease. Int. J. Mol. Sci. 2023, 24, 11701. https://doi.org/10.3390/ijms241411701
Hu Y, Cai Z, He B. Smooth Muscle Heterogeneity and Plasticity in Health and Aortic Aneurysmal Disease. International Journal of Molecular Sciences. 2023; 24(14):11701. https://doi.org/10.3390/ijms241411701
Chicago/Turabian StyleHu, Yunwen, Zhaohua Cai, and Ben He. 2023. "Smooth Muscle Heterogeneity and Plasticity in Health and Aortic Aneurysmal Disease" International Journal of Molecular Sciences 24, no. 14: 11701. https://doi.org/10.3390/ijms241411701