
Extracellular Heat Shock Protein 70 Increases the Glucocorticoid Receptor and Dual-Specificity Phosphatase 1 via Toll-like Receptor 4 and Attenuates Inflammation in Airway Epithelial Cells

Abstract
:1. Introduction
2. Results
2.1. Effects of HSP70 on GR Expression
2.2. Upregulated Expression of DUSP-1
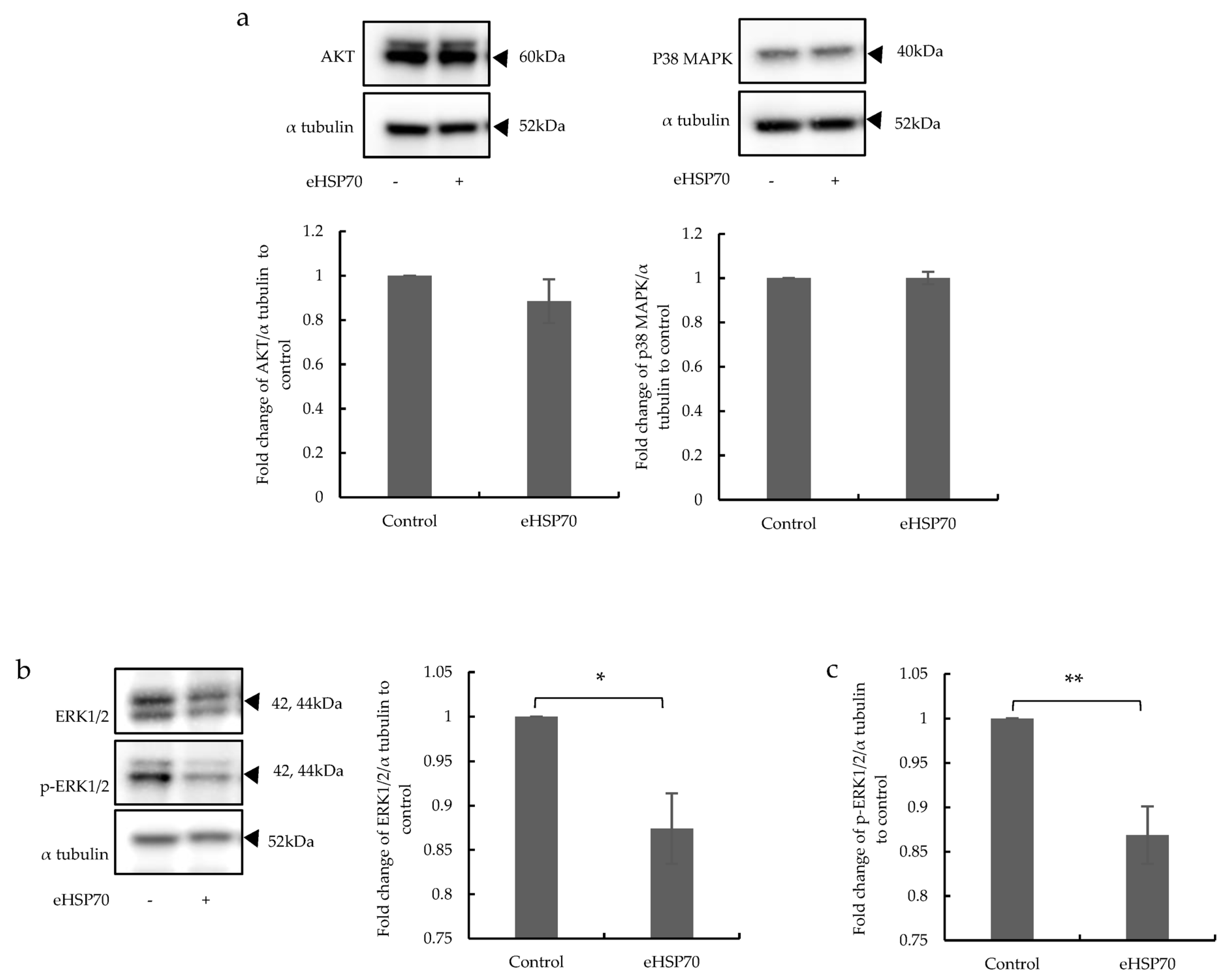
2.3. The Effects of eHSP70 Involve ERK1/2
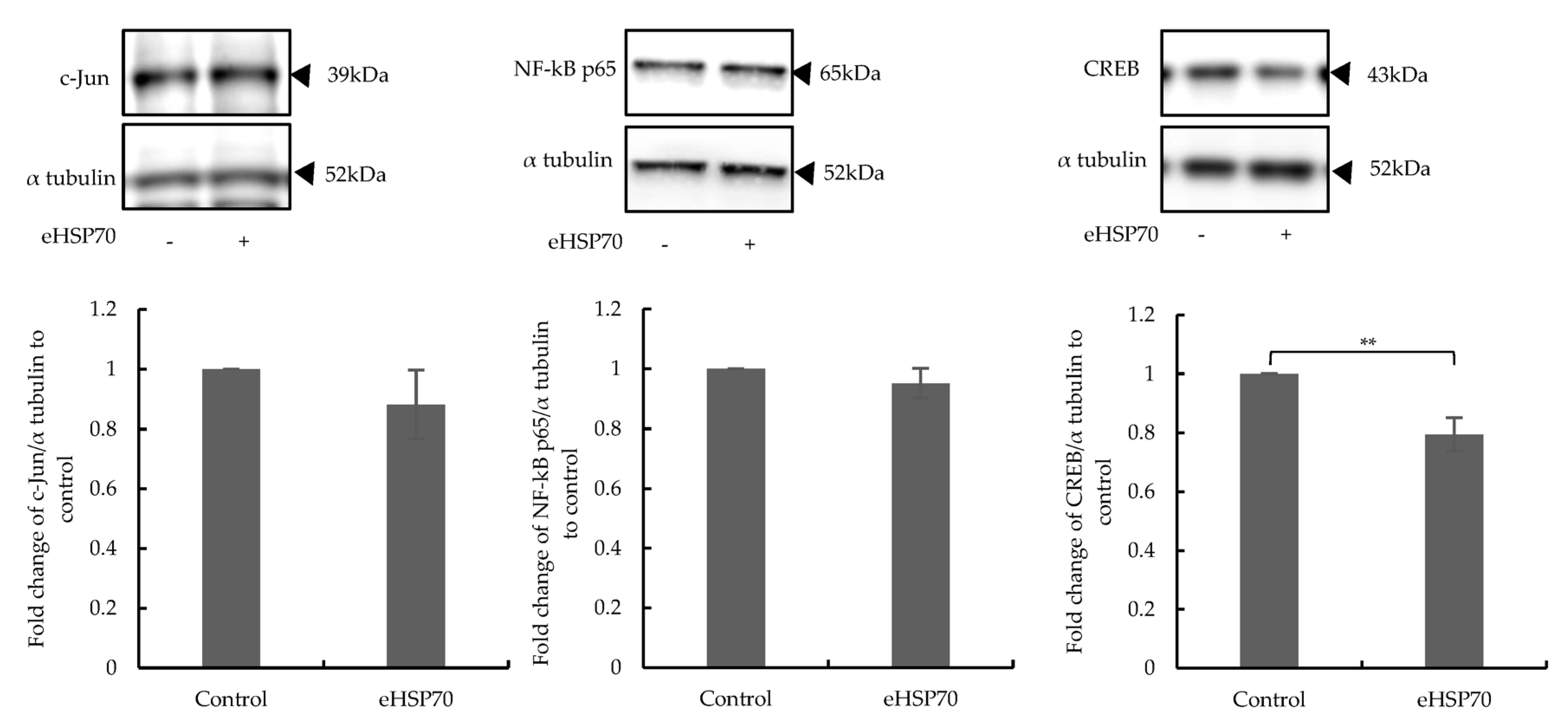
2.4. Downregulated Expression of CREB
2.5. Inhibitors of GR and TLRs Prevented the Influence of eHSP70
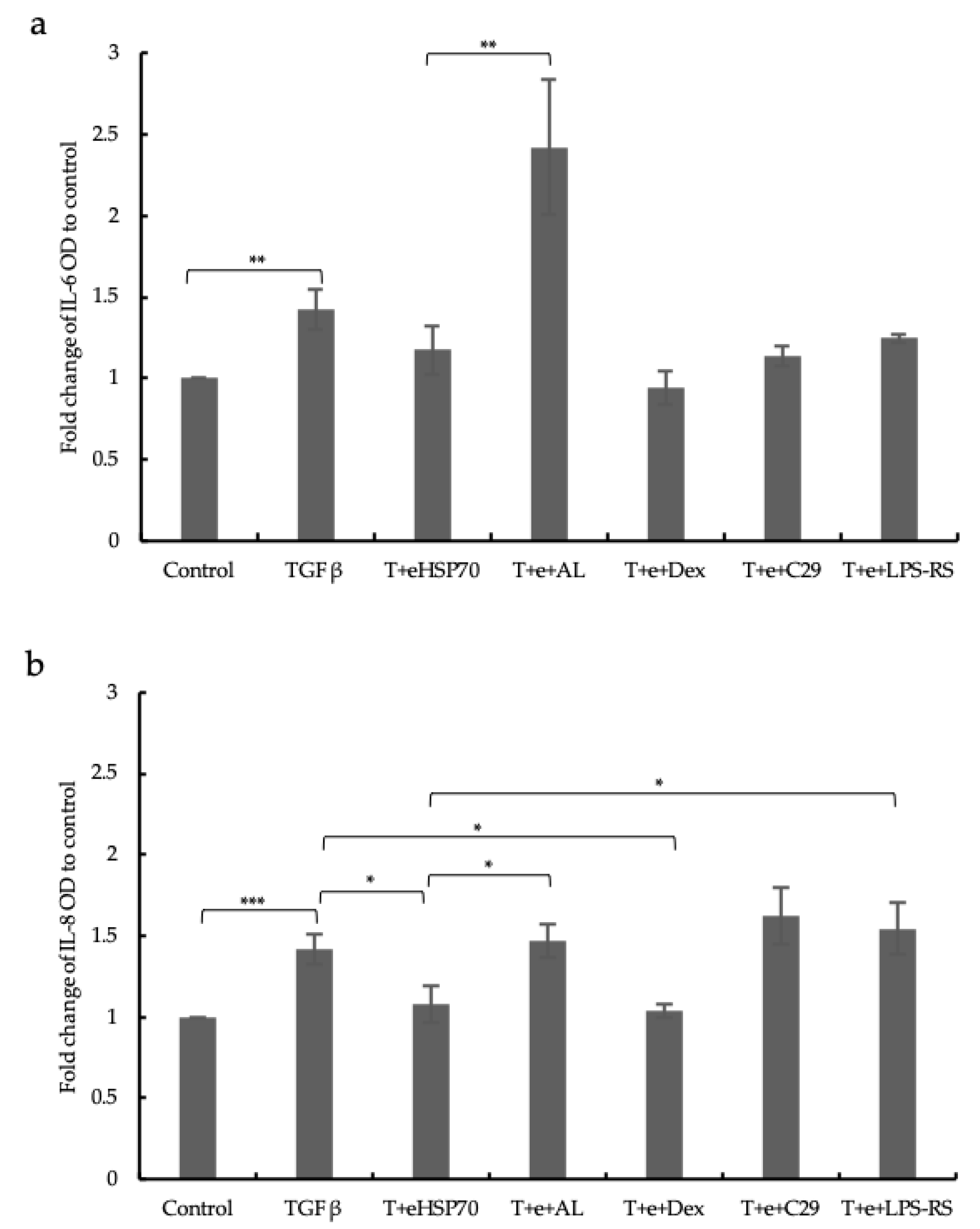
2.6. Cytokines Release
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Isolation and Treatment
4.3. Plasmid Transfection
4.4. RNA Extraction and Real-Time Reverse Transcription Polymerase Chain Reaction
4.5. Immunofluorescence Staining and Western Blot
4.6. Inhibitor Treatment
4.7. Enzyme-Linked Immunosorbent Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rosenzweig, R.; Nillegoda, N.B.; Mayer, M.P.; Bukau, B. The Hsp70 chaperone network. Nat. Rev. Mol. Cell Biol. 2019, 20, 665–680. [Google Scholar] [CrossRef] [PubMed]
- Frydman, J. Folding of newly translated proteins in vivo: The role of molecular chaperones. Annu. Rev. Biochem. 2001, 70, 603–647. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Sharma, R.; Shrestha, O.K.; Bingman, C.A.; Craig, E.A. Dual interaction of the Hsp70 J-protein cochaperone Zuotin with the 40S and 60S ribosomal subunits. Nat. Struct. Mol. Biol. 2016, 23, 1003–1010. [Google Scholar] [CrossRef] [Green Version]
- Craig, E.A. Hsp70 at the membrane: Driving protein translocation. BMC Biol. 2018, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Goloubinoff, P.; De Los Rios, P. The mechanism of Hsp70 chaperones: (entropic) pulling the models together. Trends Biochem. Sci. 2007, 32, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Jiang, Q.; Bai, X.; Yang, Y.F.; Ruan, M.Y.; Cai, S.Q. Tetrameric Assembly of K+ Channels Requires ER-Located Chaperone Proteins. Mol. Cell 2017, 65, 52–65. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.Y.R.; Noddings, C.M.; Kirschke, E.; Myasnikov, A.G.; Johnson, J.L.; Agard, D.A. Structure of Hsp90-Hsp70-Hop-GR reveals the Hsp90 client-loading mechanism. Nature 2022, 601, 460–464. [Google Scholar] [CrossRef]
- Mogk, A.; Bukau, B. Role of sHsps in organizing cytosolic protein aggregation and disaggregation. Cell Stress Chaperones 2017, 22, 493–502. [Google Scholar] [CrossRef] [Green Version]
- Meriin, A.B.; Narayanan, A.; Meng, L.; Alexandrov, I.; Varelas, X.; Cisse, I.I.; Sherman, M.Y. Hsp70-Bag3 complex is a hub for proteotoxicity-induced signaling that controls protein aggregation. Proc. Natl. Acad. Sci. USA 2018, 115, E7043–E7052. [Google Scholar] [CrossRef] [Green Version]
- Nillegoda, N.B.; Kirstein, J.; Szlachcic, A.; Berynskyy, M.; Stank, A.; Stengel, F.; Arnsburg, K.; Gao, X.C.; Scior, A.; Aebersold, R.; et al. Crucial HSP70 co-chaperone complex unlocks metazoan protein disaggregation. Nature 2015, 524, 247–251. [Google Scholar] [CrossRef] [Green Version]
- Shorter, J. The Mammalian Disaggregase Machinery: Hsp110 Synergizes with Hsp70 and Hsp40 to Catalyze Protein Disaggregation and Reactivation in a Cell-Free System. PLoS ONE 2011, 6, e26319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwirowski, S.; Klosowska, A.; Obuchowski, I.; Nillegoda, N.B.; Pirog, A.; Zietkiewicz, S.; Bukau, B.; Mogk, A.; Liberek, K. Hsp70 displaces small heat shock proteins from aggregates to initiate protein refolding. EMBO J. 2017, 36, 783–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, A.M.; Miyata, Y.; Klinedinst, S.; Peng, H.M.; Chua, J.P.; Komiyama, T.; Li, X.K.; Morishima, Y.; Merry, D.E.; Pratt, W.B.; et al. Activation of Hsp70 reduces neurotoxicity by promoting polyglutamine protein degradation. Nat. Chem. Biol. 2013, 9, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, G.Y.; Hu, C.; Yun, Y.; Yang, W.S.; Dubiel, W.; Cheng, Y.; Wolf, D.A. Dual roles of HSP70 chaperone HSPA1 in quality control of nascent and newly synthesized proteins. EMBO J. 2021, 40, e106183. [Google Scholar] [CrossRef]
- Hunter-Lavin, C.; Davies, E.L.; Bacelar, M.M.; Marshall, M.J.; Andrew, S.M.; Williams, J.H. Hsp70 release from peripheral blood mononuclear cells. Biochem. Biophys. Res. Commun. 2004, 324, 511–517. [Google Scholar] [CrossRef]
- Fang, L.; Li, J.; Papakonstantinou, E.; Karakioulaki, M.; Sun, Q.; Schumann, D.; Tamm, M.; Stolz, D.; Roth, M. Secreted heat shock proteins control airway remodeling: Evidence from bronchial thermoplasty. J. Allergy Clin. Immunol. 2021, 148, 1249–1261.e8. [Google Scholar] [CrossRef]
- Pockley, A.G.; Muthanal, M.; Calderwood, S.K. The dual immunoregulatory roles of stress proteins. Trends Biochem. Sci. 2008, 33, 71–79. [Google Scholar] [CrossRef]
- Borges, T.J.; Wieten, L.; van Herwijnen, M.J.C.; Broere, F.; van der Zee, R.; Bonorino, C.; van Eden, W. The anti-inflammatory mechanisms of Hsp70. Front. Immunol. 2012, 3, 95. [Google Scholar] [CrossRef] [Green Version]
- Wachstein, J.; Tischer, S.; Figueiredo, C.; Limbourg, A.; Falk, C.; Immenschuh, S.; Blasczyk, R.; Eiz-Vesper, B. HSP70 enhances immunosuppressive function of CD4+CD25+FoxP3+ T regulatory cells and cytotoxicity in CD4+CD25− T cells. PLoS ONE 2012, 7, e51747. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Bao, H.Y.; Jin, C.C.; Zhou, J.C.; Hua, F.; Li, K.; Lv, X.X.; Cui, B.; Hu, Z.W.; Zhang, X.W. Targeting Extracellular Heat Shock Protein 70 Ameliorates Doxorubicin-Induced Heart Failure Through Resolution of Toll-Like Receptor 2-Mediated Myocardial Inflammation. J. Am. Heart Assoc. 2019, 8, e012338. [Google Scholar] [CrossRef]
- Rodrigues-Krause, J.; Krause, M.; O’Hagan, C.; De Vito, G.; Boreham, C.; Murphy, C.; Newsholme, P.; Colleran, G. Divergence of intracellular and extracellular HSP72 in type 2 diabetes: Does fat matter? Cell Stress Chaperones 2012, 17, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidari, F.; Rabizadeh, S.; Sadat Salehi, S.; Akhavan, S.; Khaloo, P.; Alemi, H.; Mirmiranpour, H.; Esteghamati, A.; Nakhjavani, M. Serum HSP70 level in patients with endometrial cancer with and without diabetes. Gynecol. Endocrinol. 2020, 36, 351–355. [Google Scholar] [CrossRef]
- Kim, J.Y.; Barua, S.; Huang, M.Y.; Park, J.; Yenari, M.A.; Lee, J.E. Heat Shock Protein 70 (HSP70) Induction: Chaperonotherapy for Neuroprotection after Brain Injury. Cells 2020, 9, 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Kovalchin, J.T.; Muhlenkamp, P.; Chandawarkar, R.Y. Exogenous heat shock protein 70 binds macrophage lipid raft microdomain and stimulates phagocytosis, processing, and MHC-II presentation of antigens. Blood 2006, 107, 1636–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulina, A.; Grdic Rajkovic, M.; Jaksic Despot, D.; Jelic, D.; Dojder, A.; Cepelak, I.; Rumora, L. Extracellular Hsp70 induces inflammation and modulates LPS/LTA-stimulated inflammatory response in THP-1 cells. Cell Stress Chaperones 2018, 23, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Borges, T.J.; Lopes, R.L.; Pinho, N.G.; Machado, F.D.; Souza, A.P.; Bonorino, C. Extracellular Hsp70 inhibits pro-inflammatory cytokine production by IL-10 driven down-regulation of C/EBPbeta and C/EBPdelta. Int. J. Hyperthermia 2013, 29, 455–463. [Google Scholar] [CrossRef]
- Fong, J.J.; Sreedhara, K.; Deng, L.; Varki, N.M.; Angata, T.; Liu, Q.; Nizet, V.; Varki, A. Immunomodulatory activity of extracellular Hsp70 mediated via paired receptors Siglec-5 and Siglec-14. EMBO J. 2015, 34, 2775–2788. [Google Scholar] [CrossRef] [Green Version]
- Stocki, P.; Dickinson, A.M. The immunosuppressive activity of heat shock protein 70. Autoimmune Dis. 2012, 2012, 617213. [Google Scholar] [CrossRef] [Green Version]
- Vardas, K.; Ilia, S.; Sertedaki, A.; Charmandari, E.; Briassouli, E.; Goukos, D.; Apostolou, K.; Psarra, K.; Botoula, E.; Tsagarakis, S.; et al. Increased glucocorticoid receptor expression in sepsis is related to heat shock proteins, cytokines, and cortisol and is associated with increased mortality. Intensive Care Med. Exp. 2017, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Vandevyver, S.; Dejager, L.; Van Bogaert, T.; Kleyman, A.; Liu, Y.; Tuckermann, J.; Libert, C. Glucocorticoid receptor dimerization induces MKP1 to protect against TNF-induced inflammation. J. Clin. Investig. 2012, 122, 2130–2140. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; King, E.M.; Chandrasekhar, A.; Newton, R. Roles for the mitogen-activated protein kinase (MAPK) phosphatase, DUSP1, in feedback control of inflammatory gene expression and repression by dexamethasone. J. Biol. Chem. 2014, 289, 13667–13679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Shepherd, E.G.; Manson, M.E.; Nelin, L.D.; Sorokin, A.; Liu, Y. The role of mitogen-activated protein kinase phosphatase-1 in the response of alveolar macrophages to lipopolysaccharide: Attenuation of proinflammatory cytokine biosynthesis via feedback control of p38. J. Biol. Chem. 2005, 280, 8101–8108. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Wang, X.; Nelin, L.D.; Yao, Y.; Matta, R.; Manson, M.E.; Baliga, R.S.; Meng, X.; Smith, C.V.; Bauer, J.A.; et al. MAP kinase phosphatase 1 controls innate immune responses and suppresses endotoxic shock. J. Exp. Med. 2006, 203, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Barry, S.P.; Roth, R.J.; Wu, J.J.; Jones, E.A.; Bennett, A.M.; Flavell, R.A. Dynamic regulation of pro- and anti-inflammatory cytokines by MAPK phosphatase 1 (MKP-1) in innate immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 2274–2279. [Google Scholar] [CrossRef] [PubMed]
- Bonni, A.; Brunet, A.; West, A.E.; Datta, S.R.; Takasu, M.A.; Greenberg, M.E. Cell survival promoted by the Ras-MAPK signaling pathway by transcription-dependent and -independent mechanisms. Science 1999, 286, 1358–1362. [Google Scholar] [CrossRef] [Green Version]
- Mossman, B.T.; Lounsbury, K.M.; Reddy, S.P. Oxidants and signaling by mitogen-activated protein kinases in lung epithelium. Am. J. Respir. Cell Mol. Biol. 2006, 34, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Hippenstiel, S.; Opitz, B.; Schmeck, B.; Suttorp, N. Lung epithelium as a sentinel and effector system in pneumonia molecular mechanisms of pathogen recognition and signal transduction. Respir. Res. 2006, 7, 97. [Google Scholar] [CrossRef] [Green Version]
- Profita, M.; Bonanno, A.; Siena, L.; Ferraro, M.; Montalbano, A.M.; Pompeo, F.; Riccobono, L.; Pieper, M.P.; Gjomarkaj, M. Acetylcholine mediates the release of IL-8 in human bronchial epithelial cells by a NFkB/ERK-dependent mechanism. Eur. J. Pharmacol. 2008, 582, 145–153. [Google Scholar] [CrossRef]
- Berube, J.; Roussel, L.; Nattagh, L.; Rousseau, S. Loss of cystic fibrosis transmembrane conductance regulator function enhances activation of p38 and ERK MAPKs, increasing interleukin-6 synthesis in airway epithelial cells exposed to Pseudomonas aeruginosa. J. Biol. Chem. 2010, 285, 22299–22307. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, D. Transforming growth factor beta: A central modulator of pulmonary and airway inflammation and fibrosis. Proc. Am. Thorac. Soc. 2006, 3, 413–417. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Fenoglio, S.; Gao, D.C.; Camiolo, M.; Stiles, B.; Lindsted, T.; Schlederer, M.; Johns, C.; Altorki, N.; Mittal, V.; et al. TGF-beta IL-6 axis mediates selective and adaptive mechanisms of resistance to molecular targeted therapy in lung cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 15535–15540. [Google Scholar] [CrossRef] [PubMed]
- Grad, I.; Picard, D. The glucocorticoid responses are shaped by molecular chaperones. Mol. Cell Endocrinol. 2007, 275, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Town, T.; Gokarn, V.; Flavell, R.A.; Chandawarkar, R.Y. HSP70 enhances macrophage phagocytosis by interaction with lipid raft-associated TLR-7 and upregulating p38 MAPK and PI3K pathways. J. Surg. Res. 2006, 136, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Koletsa, T.; Zhou, L.; Fang, L.; Roth, M.; Karakioulaki, M.; Savic, S.; Grize, L.; Tamm, M.; Stolz, D. Bronchial thermoplasty in asthma: An exploratory histopathological evaluation in distinct asthma endotypes/phenotypes. Respir. Res. 2021, 22, 186. [Google Scholar] [CrossRef]
- Torrego, A.; Herth, F.J.; Munoz-Fernandez, A.M.; Puente, L.; Facciolongo, N.; Bicknell, S.; Novali, M.; Gasparini, S.; Bonifazi, M.; Dheda, K.; et al. Bronchial Thermoplasty Global Registry (BTGR): 2-year results. BMJ Open 2021, 11, e053854. [Google Scholar] [CrossRef]
- Denner, D.R.; Doeing, D.C.; Hogarth, D.K.; Dugan, K.; Naureckas, E.T.; White, S.R. Airway Inflammation after Bronchial Thermoplasty for Severe Asthma. Ann. Am. Thorac. Soc. 2015, 12, 1302–1309. [Google Scholar] [CrossRef] [Green Version]
- Postigo, M.; Hall, C.S.; Castro, M. Predicting the Response to Bronchial Thermoplasty: The Needier, the Better. J. Allergy Clin. Immunol. Pract. 2020, 8, 1261–1262. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, C.T.; Kim, Y.W.; Han, S.K.; Shim, Y.S.; Yoo, C.G. Preheating accelerates mitogen-activated protein (MAP) kinase inactivation post-heat shock via a heat shock protein 70-mediated increase in phosphorylated MAP kinase phosphatase-1. J. Biol. Chem. 2005, 280, 13179–13186. [Google Scholar] [CrossRef] [Green Version]
- Junchao, Y.; Zhen, W.; Yuan, W.; Liying, X.; Libin, J.; Yuanhong, Z.; Wei, Z.; Ruilin, C.; Lu, Z. Anti- trachea inflammatory effects of diosgenin from Dioscorea nipponica through interactions with glucocorticoid receptor alpha. J. Int. Med. Res. 2017, 45, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Barlow, C.A.; Shukla, A.; Mossman, B.T.; Lounsbury, K.M. Oxidant-mediated cAMP response element binding protein activation: Calcium regulation and role in apoptosis of lung epithelial cells. Am. J. Respir. Cell Mol. Biol. 2006, 34, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Asea, A.; Rehli, M.; Kabingu, E.; Boch, J.A.; Bare, O.; Auron, P.E.; Stevenson, M.A.; Calderwood, S.K. Novel signal transduction pathway utilized by extracellular HSP70: Role of toll-like receptor (TLR) 2 and TLR4. J. Biol. Chem. 2002, 277, 15028–15034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, K.N.; Kitajima, M.; Imamura, T.; Hiraki, K.; Fujishita, A.; Sekine, I.; Ishimaru, T.; Masuzaki, H. Toll-like receptor 4-mediated growth of endometriosis by human heat-shock protein 70. Hum. Reprod. 2008, 23, 2210–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Liu, Z.L.; Ding, H.; Zhou, Y.; Doan, H.A.; Sin, K.W.T.; Zhu, Z.J.R.; Flores, R.; Wen, Y.F.; Gong, X.; et al. Tumor induces muscle wasting in mice through releasing extracellular Hsp70 and Hsp90. Nat. Commun. 2017, 8, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosemans, G.; Merckx, C.; De Bleecker, J.L.; De Paepe, B. Inducible Heat Shock Protein 70 Levels in Patients and the mdx Mouse Affirm Regulation during Skeletal Muscle Regeneration in Muscular Dystrophy. Front. Biosci. (Schol. Ed.) 2022, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shan, P.; Srivastava, A.; Jiang, G.; Zhang, X.; Lee, P.J. An Endothelial Hsp70-TLR4 Axis Limits Nox3 Expression and Protects Against Oxidant Injury in Lungs. Antioxid. Redox Signal. 2016, 24, 991–1012. [Google Scholar] [CrossRef] [Green Version]
- Vicencio, J.M.; Yellon, D.M.; Sivaraman, V.; Das, D.; Boi-Doku, C.; Arjun, S.; Zheng, Y.; Riquelme, J.A.; Kearney, J.; Sharma, V.; et al. Plasma exosomes protect the myocardium from ischemia-reperfusion injury. J. Am. Coll. Cardiol. 2015, 65, 1525–1536. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, T.J.; Greenstein, J.I.; Loewenstern, J.; Degermentzidis, E.; Yao, L. Anti-inflammatory peptide regulates the supply of heat shock protein 70 monomers: Implications for aging and age-related disease. Rejuvenation Res. 2015, 18, 136–144. [Google Scholar] [CrossRef]
- Zhou, Y.Z.; Wang, Y.; Xu, F.X.; Song, C.X.; Yang, X.; Zhang, Z.; Yi, M.F.; Ma, N.; Zhou, X.F.; He, J.N. Small HSPs play an important role in crosstalk between HSF-HSP and ROS pathways in heat stress response through transcriptomic analysis in lilies (Lilium longiflorum). BMC Plant Biol. 2022, 22, 202. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Dilution | Catalog Number | Company |
|---|---|---|---|
| p38 MAPK | 1:500 | 9212S | Cell Signaling (Danvers, MA, USA) |
| c-Jun | 1:1000 | ab40766 | Abcam (Cambridge, UK) |
| NF-kB p65 | 1:400 | D14E12 | Cell Signaling |
| HSP70 | 1:2000 | #4873 | Cell Signaling |
| α Tubulin | 1:5000 | MAB9344 | R&D systems |
| DUSP-1 | 1:1000 | 48625 | Cell Signaling |
| ERK 1/2 | 1:1000 | 9102l | Cell Signaling |
| p-ERK1/2 | 1:1000 | 9101s | Cell Signaling |
| AKT | 1:1000 | 4691s | Cell Signaling |
| CREB | 1:1000 | #4820 | Cell Signaling |
| GR | 1:1000 | ab183127 | Abcam |
| GAPDH | 1:1000 | ab181602 | Abcam |
| Primary Antibody | Dilution | Catalog Number | Company |
|---|---|---|---|
| DUSP-1 | 1:100 | 48625 | Cell Signaling |
| GR | 1:500 | ab183127 | Abcam |
| Gene | Forward Primer (5′-3′) | Reverse Primer (3′–5′) |
|---|---|---|
| α tubulin | AGGAGTCCAGATCGGCAATG | GTCCCCACCACCAATGGTTT |
| hDUSP1 | CTGCCTTGATCAACGTCTCA | CTGTGCCTTGTGGTTGTCCT |
| GR | ATAGCTCTGTTCCAGACTCAACT | TCCTGAAACCTGGTATTGCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, L.; Fang, L.; Tamm, M.; Stolz, D.; Roth, M. Extracellular Heat Shock Protein 70 Increases the Glucocorticoid Receptor and Dual-Specificity Phosphatase 1 via Toll-like Receptor 4 and Attenuates Inflammation in Airway Epithelial Cells. Int. J. Mol. Sci. 2023, 24, 11700. https://doi.org/10.3390/ijms241411700
Zhou L, Fang L, Tamm M, Stolz D, Roth M. Extracellular Heat Shock Protein 70 Increases the Glucocorticoid Receptor and Dual-Specificity Phosphatase 1 via Toll-like Receptor 4 and Attenuates Inflammation in Airway Epithelial Cells. International Journal of Molecular Sciences. 2023; 24(14):11700. https://doi.org/10.3390/ijms241411700
Chicago/Turabian StyleZhou, Liang, Lei Fang, Michael Tamm, Daiana Stolz, and Michael Roth. 2023. "Extracellular Heat Shock Protein 70 Increases the Glucocorticoid Receptor and Dual-Specificity Phosphatase 1 via Toll-like Receptor 4 and Attenuates Inflammation in Airway Epithelial Cells" International Journal of Molecular Sciences 24, no. 14: 11700. https://doi.org/10.3390/ijms241411700