
A dePEGylated Lipopeptide-Based Pan-Coronavirus Fusion Inhibitor Exhibits Potent and Broad-Spectrum Anti-HIV-1 Activity without Eliciting Anti-PEG Antibodies
 , , and
, , and
Abstract
:1. Introduction
2. Results
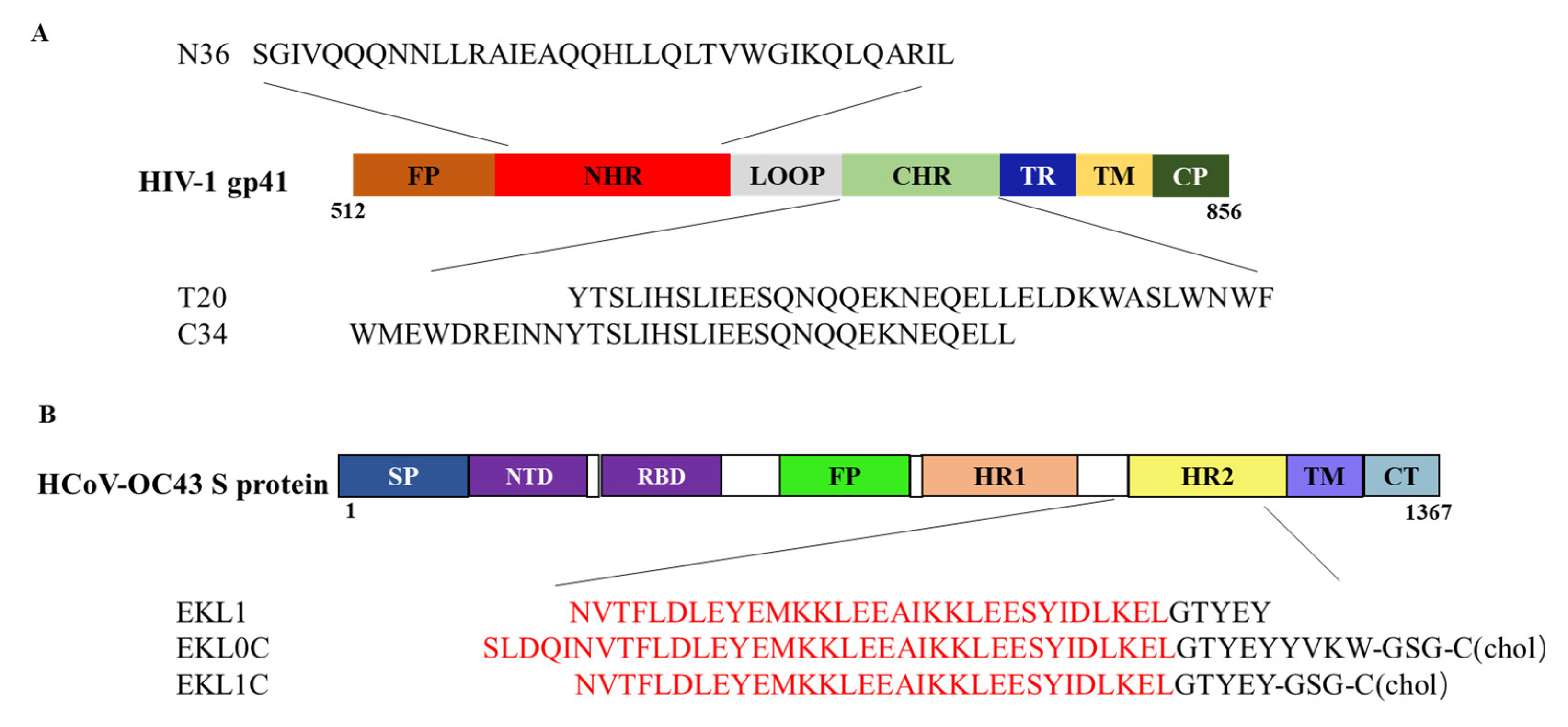
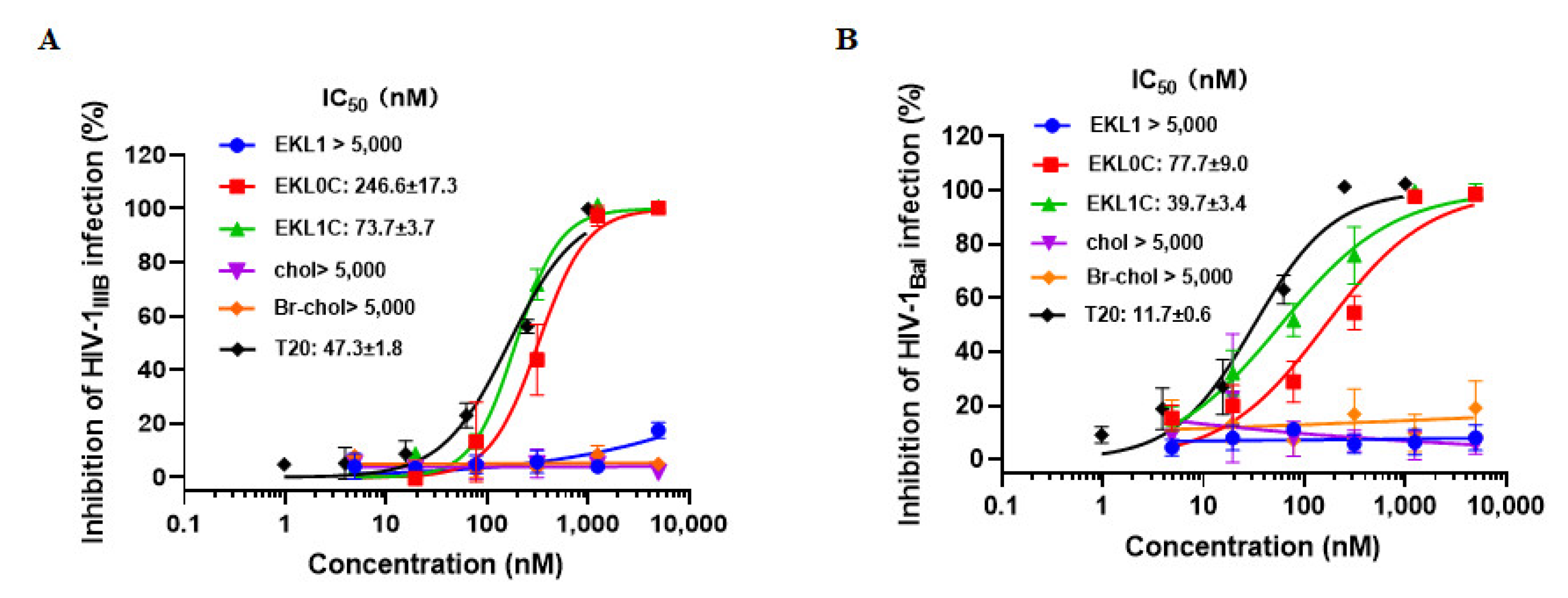
2.1. The dePEGylated Lipopeptide-Based Pan-CoV Fusion Inhibitor EKL1C Exhibited Potent Inhibitory Activity against a Broad Spectrum of Pseudotyped HIV-1 Strains
2.2. EKL1C Exhibited Potency against a Broad Spectrum of HIV-1 Laboratory-Adapted Strains
2.3. EKL1C Exhibited Potency against a Broad Spectrum of HIV-1 Primary Isolates
2.4. EKL1C Exhibited Potency against a Broad Spectrum of T20-Resistant HIV-1 Strains
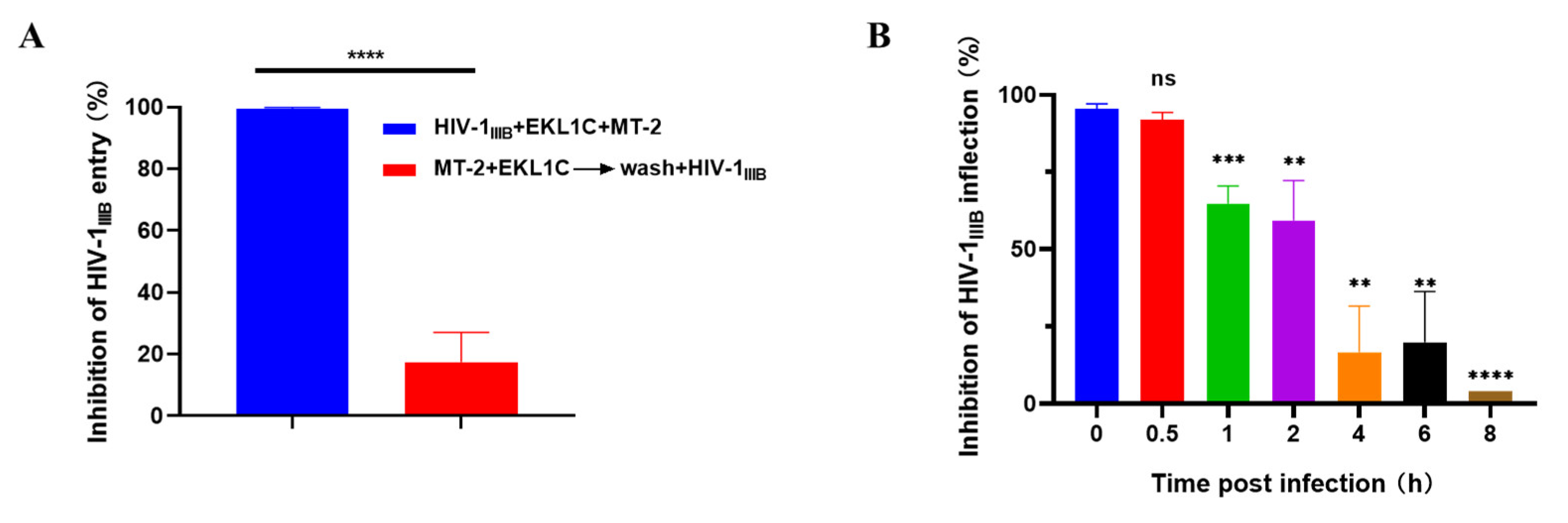
2.5. EKL1C Inhibited HIV-1 Infection at the Early Stage of Viral Replication
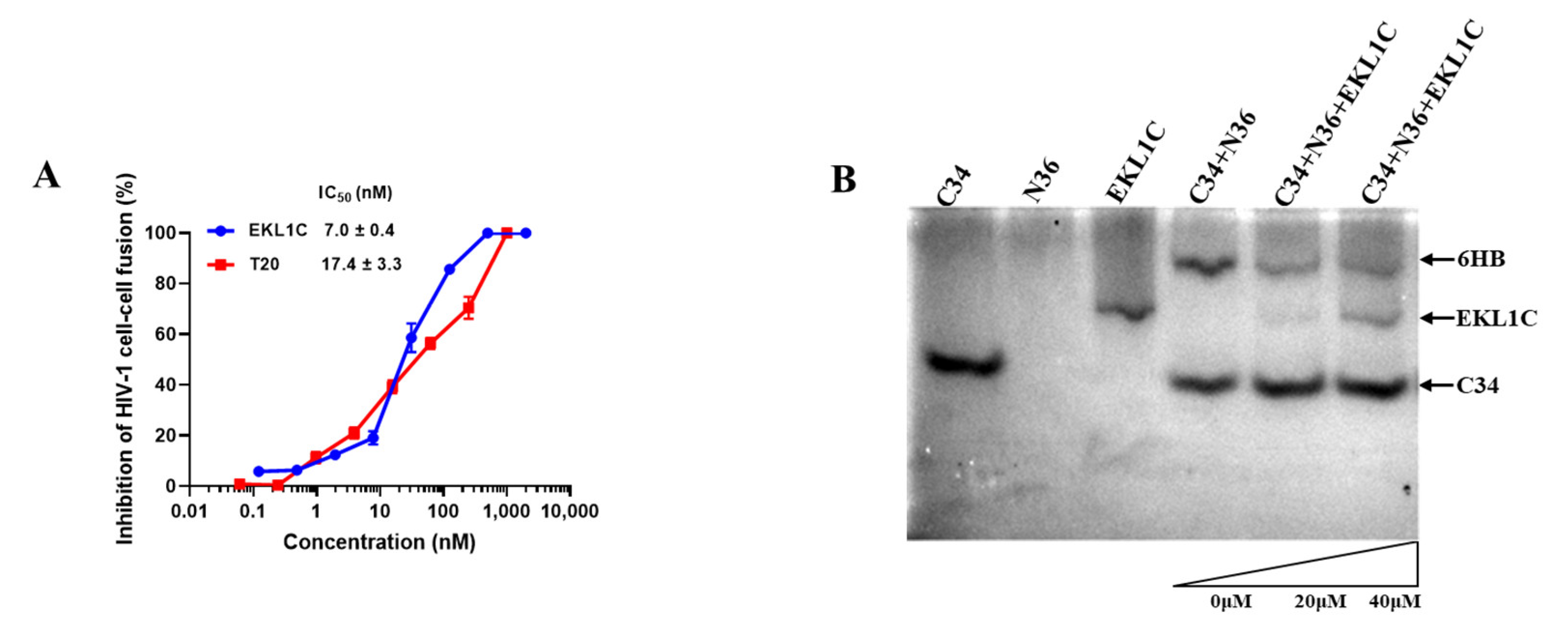
2.6. EKL1C Inhibited HIV-1-Env-Mediated Cell–Cell Fusion
2.7. EKL1C Bound to the HR1 of gp41 and Blocked the Formation of 6-HB
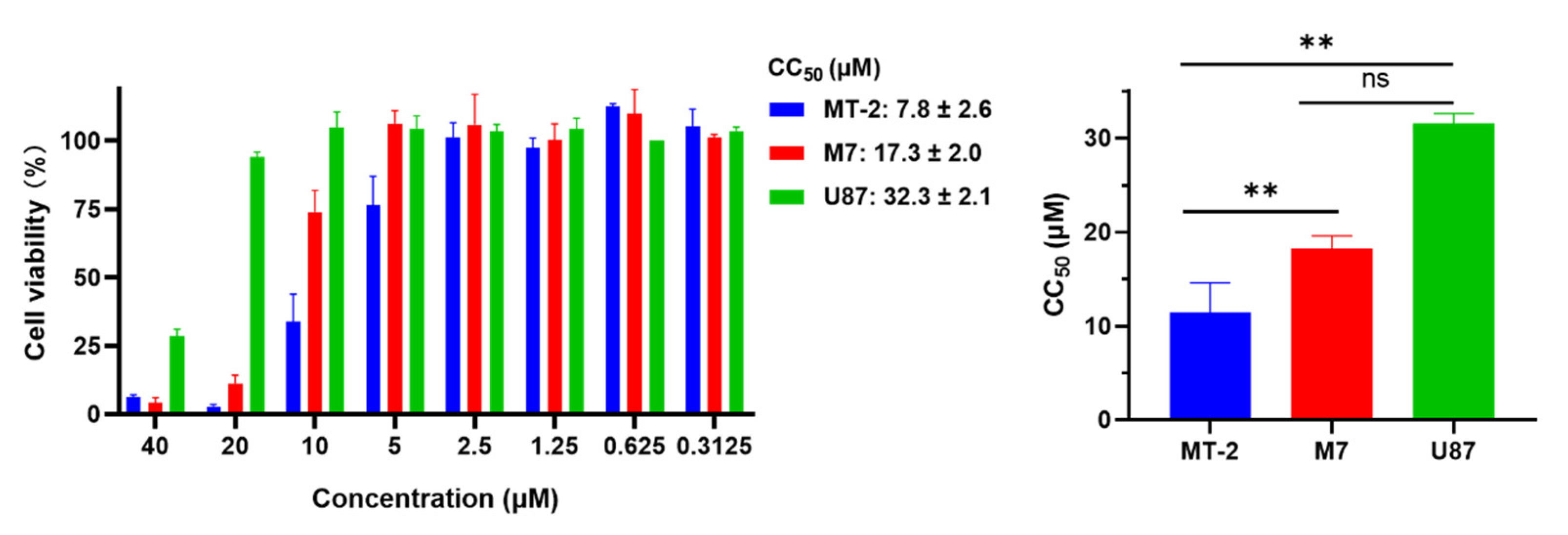
2.8. EKL1C at 5 μM Exhibited no Obvious In Vitro Cytotoxicity to MT-2 Cells, CEMx174 517 5.25 M7 Cells, and U87 CD4+ CCR5+ Cells
3. Discussion
4. Materials and Methods
4.1. Cells, Viruses, and Peptides
4.2. Plasmids
4.3. Inhibition of HIV-1 Pseudovirus Infection
4.4. Inhibition of HIV-1 Infection
4.5. Cell-Washout Assay
4.6. Time-of-Addition Assay
4.7. HIV-1-Env-Mediated Cell–Cell Fusion Assay
4.8. N-PAGE
4.9. Cytotoxicity Assay
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peeters, M.; Jung, M.; Ayouba, A. The origin and molecular epidemiology of HIV. Expert Rev. Anti-Infect. Ther. 2013, 11, 885–896. [Google Scholar] [CrossRef]
- Muralidar, S.; Gopal, G.; Visaga Ambi, S. Targeting the viral-entry facilitators of SARS-CoV-2 as a therapeutic strategy in COVID-19. J. Med. Virol. 2021, 93, 5260–5276. [Google Scholar] [CrossRef]
- Harrison, S.C. Viral membrane fusion. Virology 2015, 479–480, 498–507. [Google Scholar] [CrossRef] [Green Version]
- Chen, B. Molecular Mechanism of HIV-1 Entry. Trends Microbiol. 2019, 27, 878–891. [Google Scholar] [CrossRef]
- Melikyan, G.B. HIV entry: A game of hide-and-fuse? Curr. Opin. Virol. 2014, 4, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, M.; Wang, W.; De Feo, C.J.; Vassell, R.; Weiss, C.D. Trimeric, coiled-coil extension on peptide fusion inhibitor of HIV-1 influences selection of resistance pathways. J. Biol. Chem. 2012, 287, 8297–8309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattnaik, G.P.; Chakraborty, H. Entry Inhibitors: Efficient Means to Block Viral Infection. J. Membr. Biol. 2020, 253, 425–444. [Google Scholar] [CrossRef] [PubMed]
- Huhmann, S.; Nyakatura, E.K.; Rohrhofer, A.; Moschner, J.; Schmidt, B.; Eichler, J.; Roth, C.; Koksch, B. Systematic Evaluation of Fluorination as Modification for Peptide-Based Fusion Inhibitors against HIV-1 Infection. ChemBioChem 2021, 22, 3443–3451. [Google Scholar] [CrossRef]
- Lan, Q.; Pu, J.; Cai, Y.; Zhou, J.; Wang, L.; Jiao, F.; Xu, W.; Wang, Q.; Xia, S.; Lu, L.; et al. Lipopeptide-based pan-CoV fusion inhibitors potently inhibit HIV-1 infection. Microbes Infect. 2021, 23, 10484. [Google Scholar] [CrossRef] [PubMed]
- Povsic, T.J.; Lawrence, M.G.; Lincoff, A.M.; Mehran, R.; Rusconi, C.P.; Zelenkofske, S.L.; Huang, Z.; Sailstad, J.; Armstrong, P.W.; Steg, P.G.; et al. Pre-existing anti-PEG antibodies are associated with severe immediate allergic reactions to pegnivacogin, a PEGylated aptamer. J. Allergy. Clin. Immunol. 2016, 138, 1712–1715. [Google Scholar] [CrossRef] [Green Version]
- Garay, R.P.; El-Gewely, R.; Armstrong, J.K.; Garratty, G.; Richette, P. Antibodies against polyethylene glycol in healthy subjects and in patients treated with PEG-conjugated agents. Expert Opin. Drug Deliv. 2012, 9, 1319–1323. [Google Scholar] [CrossRef]
- Zhou, J.; Xu, W.; Liu, Z.; Wang, C.; Xia, S.; Lan, Q.; Cai, Y.; Su, S.; Pu, J.; Xing, L.; et al. A highly potent and stable pan-coronavirus fusion inhibitor as a candidate prophylactic and therapeutic for COVID-19 and other coronavirus diseases. Acta Pharm. Sin. B. 2022, 12, 1652–1661. [Google Scholar] [CrossRef] [PubMed]
- Reiser, K.; Mathys, L.; Curbo, S.; Pannecouque, C.; Noppen, S.; Liekens, S.; Engman, L.; Lundberg, M.; Balzarini, J.; Karlsson, A. The Cellular Thioredoxin-1/Thioredoxin Reductase-1 Driven Oxidoreduction Represents a Chemotherapeutic Target for HIV-1 Entry Inhibition. PloS ONE 2016, 11, e0147773. [Google Scholar] [CrossRef] [Green Version]
- Este, J.A. Virus entry as a target for anti-HIV intervention. Curr. Med. Chem. 2003, 10, 1617–1632. [Google Scholar] [CrossRef] [PubMed]
- Louis, J.M.; Baber, J.L.; Clore, G.M. The C34 Peptide Fusion Inhibitor Binds to the Six-Helix Bundle Core Domain of HIV-1 gp41 by Displacement of the C-Terminal Helical Repeat Region. Biochemistry 2015, 54, 6796–6805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, S.; Yan, L.; Xu, W.; Agrawal, A.S.; Algaissi, A.; Tseng, C.K.; Wang, Q.; Du, L.; Tan, W.; Wilson, I.A.; et al. A pan-coronavirus fusion inhibitor targeting the HR1 domain of human coronavirus spike. Sci. Adv. 2019, 5, eaav4580. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Liu, M.; Wang, C.; Xu, W.; Lan, Q.; Feng, S.; Qi, F.; Bao, L.; Du, L.; Liu, S.; et al. Inhibition of SARS-CoV-2 (previously 2019-nCoV) infection by a highly potent pan-coronavirus fusion inhibitor targeting its spike protein that harbors a high capacity to mediate membrane fusion. Cell Res. 2020, 30, 343–355. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhao, Q.; Jiang, S. Determination of the HIV-1 gp41 fusogenic core conformation modeled by synthetic peptides: Applicable for identification of HIV-1 fusion inhibitors. Peptides. 2003, 24, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, X.; Wang, H.; Pu, J.; Li, Q.; Li, J.; Liu, Y.; Lu, L.; Jiang, S. A “Two-Birds-One-Stone” Approach toward the Design of Bifunctional Human Immunodeficiency Virus Type 1 Entry Inhibitors Targeting the CCR5 Coreceptor and gp41 N-Terminal Heptad Repeat Region. J. Med. Chem. 2021, 64, 11460–11471. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.T.; Shugars, D.C.; Greenwell, T.K.; McDanal, C.B.; Matthews, T.J. Peptides corresponding to a predictive alpha-helical domain of human immunodeficiency virus type 1 gp41 are potent inhibitors of virus infection. Proc. Natl. Acad. Sci. USA 1994, 91, 9770–9774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, S.; Lin, K.; Strick, N.; Neurath, A.R. HIV-1 inhibition by a peptide. Nature. 1993, 365, 11. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Blacklow, S.C.; Kim, P.S. A trimeric structural domain of the HIV-1 transmembrane glycoprotein. Nat. Struct. Biol. 1995, 2, 1075–1082. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C.; Fass, D.; Berger, J.M.; Kim, P.S. Core structure of gp41 from the HIV envelope glycoprotein. Cell. 1997, 89, 263–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio, A.A.; Filsinger Interrante, M.V.; Bell, B.N.; Brown, C.L.; Bruun, T.U.J.; LaBranche, C.C.; Montefiori, D.C.; Kim, P.S. A Derivative of the D5 Monoclonal Antibody That Targets the gp41 N-Heptad Repeat of HIV-1 with Broad Tier-2-Neutralizing Activity. J. Virol. 2021, 95, e0235020. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cheng, J.; Lu, H.; Li, J.; Hu, J.; Qi, Z.; Liu, Z.; Jiang, S.; Dai, Q. Potent HIV fusion ienhibitors against Enfuvirtide-resistant HIV-1 strains. Proc. Natl. Acad. Sci. USA 2008, 105, 16332–16337. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Curreli, F.; Zhang, X.; Bhattacharya, S.; Waheed, A.A.; Cooper, A.; Cowburn, D.; Freed, E.O.; Debnath, A.K. Antiviral activity of α-helical stapled peptides designed from the HIV-1 capsid dimerization domain. Retrovirology. 2011, 8, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, B.; Sanna, G.; Madeddu, S.; Hollmann, A.; Santos, N.C. Combining 25-Hydroxycholesterol with an HIV Fusion Inhibitor Peptide: Interaction with Biomembrane Model Systems and Human Blood Cells. ACS. Infect. Dis. 2019, 5, 582–591. [Google Scholar] [CrossRef]
- Li, H.; Yu, F.; Xia, S.; Yu, Y.; Wang, Q.; Lv, M.; Wang, Y.; Jiang, S.; Lu, L. Chemically Modified Human Serum Albumin Potently Blocks Entry of Ebola Pseudoviruses and Viruslike Particles. Antimicrob. Agents Chemother. 2017, 61, e02168-16. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Li, W.; Li, Z.; Li, Y.; Luo, J.; Yu, L.; Yang, J.; Qiu, M.; Cheng, H.; Xu, W.; et al. A Novel CXCR4 Targeting Protein SDF-1/54 as an HIV-1 Entry Inhibitor. Viruses 2019, 11, 874. [Google Scholar] [CrossRef] [Green Version]
- Gallo, S.A.; Clore, G.M.; Louis, J.M.; Bewley, C.A.; Blumenthal, R. Temperature-dependent intermediates in HIV-1 envelope glycoprotein-mediated fusion revealed by inhibitors that target N- and C-terminal helical regions of HIV-1 gp41. Biochemistry 2004, 43, 8230–8233. [Google Scholar] [CrossRef]
- Lu, H.; Zhao, Q.; Xu, Z.; Jiang, S. Automatic quantitation of HIV-1 mediated cell-to-cell fusion with a digital image analysis system (DIAS): Application for rapid screening of HIV-1 fusion inhibitors. J. Virol. Methods 2003, 107, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Klug, Y.A.; Ashkenazi, A.; Viard, M.; Porat, Z.; Blumenthal, R.; Shai, Y. Early and late HIV-1 membrane fusion events are impaired by sphinganine lipidated peptides that target the fusion site. Biochem. J. 2014, 461, 213–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.; Rasquinha, G.; Du, L.; Wang, Q.; Xu, W.; Li, W.; Lu, L.; Jiang, S. A Peptide-Based HIV-1 Fusion Inhibitor with Two Tail-Anchors and Palmitic Acid Exhibits Substantially Improved In Vitro and Ex Vivo Anti-HIV-1 Activity and Prolonged In Vivo Half-Life. Molecules 2019, 24, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Titer | IC50 (nM) | |
|---|---|---|---|
| EKL1C | T20 | ||
| Q769env.h5 (A, R5) | 2 | 22.1 ± 14.4 | 179.3 ± 76.9 |
| Q842env.d16 (A, R5) | 2 | 51.2 ± 12.7 | 23.8 ± 30.3 |
| Q259env.w6 (A, R5) | 2 | 39.0 ± 33.9 | 50.8 ± 22.9 |
| Bal (B, R5) | 3 | 37.6 ± 26.2 | 23.2 ± 15.8 |
| REJO4541 (B, R5) | - | 17.6 ± 15.8 | 11.6 ± 13.7 |
| SF162 (B, R5) | 1A | 186.5 ± 85.3 | 302.7 ± 164.1 |
| JRFL (B, R5) | 2 | 158.4 ± 57.9 | 222.4 ± 170.3 |
| pRHPA4259 clone 7 (B, R5) | 2 | 147.4 ± 64.2 | 14.5 ± 5.9 |
| ZM53M.PB12 (C, R5) | 2 | 6.6 ± 6.8 | 51.4 ± 41.0 |
| HIV-25710-2, clone 43 (C, R5) | 1B | 23.8 ± 20.6 | 53.0 ± 27.1 |
| QA013.70I.ENV.M12 (D, R5) | 2 | 31.5 ± 13.8 | 6.5 ± 3.5 |
| QD435.100M.ENV.E1 (D, R5) | 2 | 48.1 ± 33.1 | 72.0 ± 24.8 |
| Subtype G clone 252 (G, R5) | 2 | 98.5 ± 59.4 | 27.5 ± 9.5 |
| CRF01_AE clone 269 (A/E, R5) | - | 5.1 ± 1.234 | 14.2 ± 17.5 |
| GX11.13 (A/E, R5) | - | 10.3 ± 7.5 | 74.1 ± 60.2 |
| CRF02_AG clone 266 (A/G, R5) | 2 | 37.5 ± 3.3 | 37.1 ± 7.2 |
| CRF02_AG clone 271 (A/G, R5) | 1B | 19.1 ± 8.2 | 11.5 ± 6.4 |
| CRF02_AG clone 278 (A/G, R5) | 3 | 19.4 ± 14.2 | 28.4 ± 28.6 |
| BC02 (B/C, R5) | - | 20.5 ± 11.0 | 31.7 ± 34.0 |
| CH119 (B/C, R5) | 2 | 74.2 ± 45.6 | 394.2 ± 71.6 |
| Virus | Subtype, Tropism | IC50 (nM) | |
|---|---|---|---|
| EKL1C | T20 | ||
| MN/H9 (84US_MNp) | (A, X4) | 98.1 ± 29.2 | 52.6 ± 7.4 |
| BZ167/GS 010 (89BZ_167) | (B, X4) | 118.4 ± 17.0 | 47.3 ± 5.9 |
| 92UG024 | (D, X4) | 80.2 ± 7.6 | 31.0 ± 5.6 |
| J32228M4 | (D, R5) | 70.1 ± 3.2 | 26.2 ± 3.1 |
| HIVBCF02 | (O, R5) | 192.9 ± 37.6 | 41.3 ± 1.4 |
| Virus | IC50 (nM) | |
|---|---|---|
| EKL1C | T20 | |
| V38A, N42D | 36.1 ± 11.2 | >1000 |
| HIV-1 NL4-3 D36G (WT) | 17.2 ± 10.2 | 10.2 ± 4.1 |
| V38A | 97.9 ± 28.3 | >1000 |
| N42T, N43K | 9.4 ± 1.2 | >1000 |
| V38E, N42S | 19.7 ± 10.3 | >1000 |
| V38A, N42T | 17.4 ± 1.5 | >1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Wang, C.; Xu, W.; Xing, L.; Zhou, J.; Pu, J.; Fu, M.; Lu, L.; Jiang, S.; Wang, Q. A dePEGylated Lipopeptide-Based Pan-Coronavirus Fusion Inhibitor Exhibits Potent and Broad-Spectrum Anti-HIV-1 Activity without Eliciting Anti-PEG Antibodies. Int. J. Mol. Sci. 2023, 24, 9779. https://doi.org/10.3390/ijms24119779
Xu L, Wang C, Xu W, Xing L, Zhou J, Pu J, Fu M, Lu L, Jiang S, Wang Q. A dePEGylated Lipopeptide-Based Pan-Coronavirus Fusion Inhibitor Exhibits Potent and Broad-Spectrum Anti-HIV-1 Activity without Eliciting Anti-PEG Antibodies. International Journal of Molecular Sciences. 2023; 24(11):9779. https://doi.org/10.3390/ijms24119779
Chicago/Turabian StyleXu, Ling, Chao Wang, Wei Xu, Lixiao Xing, Jie Zhou, Jing Pu, Mingming Fu, Lu Lu, Shibo Jiang, and Qian Wang. 2023. "A dePEGylated Lipopeptide-Based Pan-Coronavirus Fusion Inhibitor Exhibits Potent and Broad-Spectrum Anti-HIV-1 Activity without Eliciting Anti-PEG Antibodies" International Journal of Molecular Sciences 24, no. 11: 9779. https://doi.org/10.3390/ijms24119779