
Repetitive Cerulein-Induced Chronic Pancreatitis in Growing Pigs—A Pilot Study
 , , , , , , , ,
, , , , , , , ,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
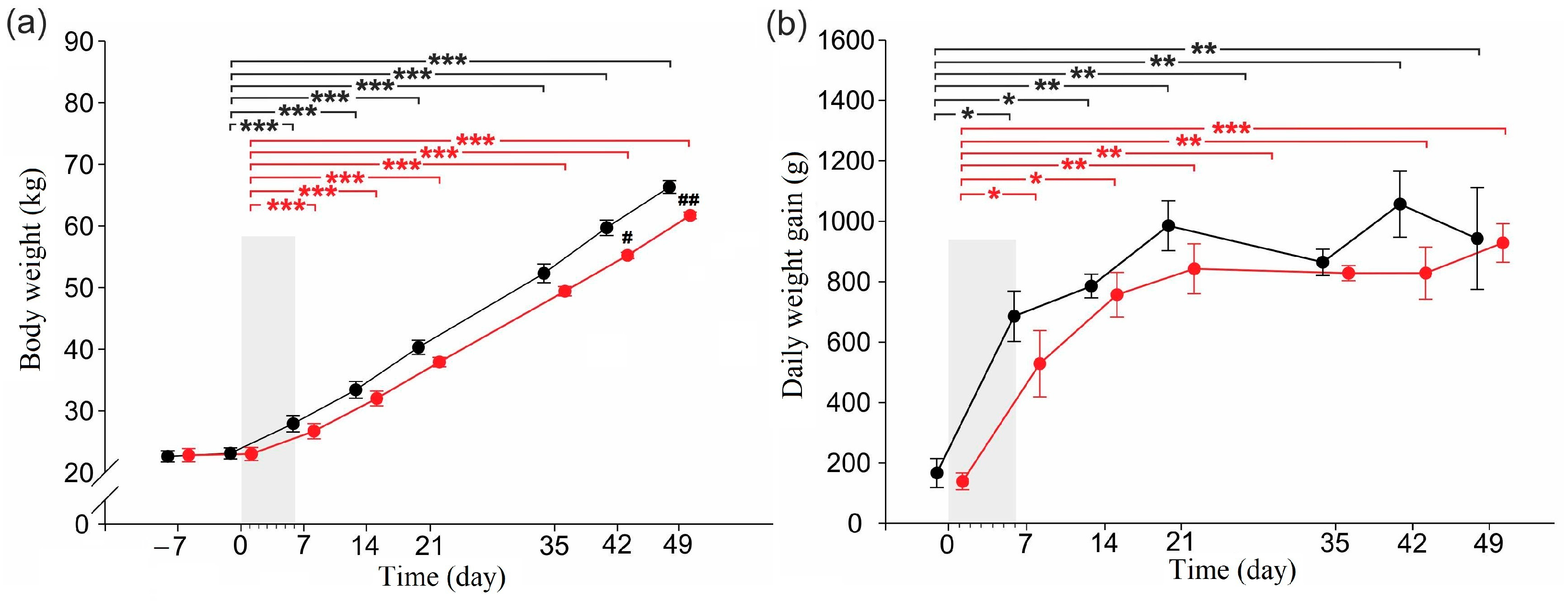
2.1. Body Weight and Daily Weight Gain
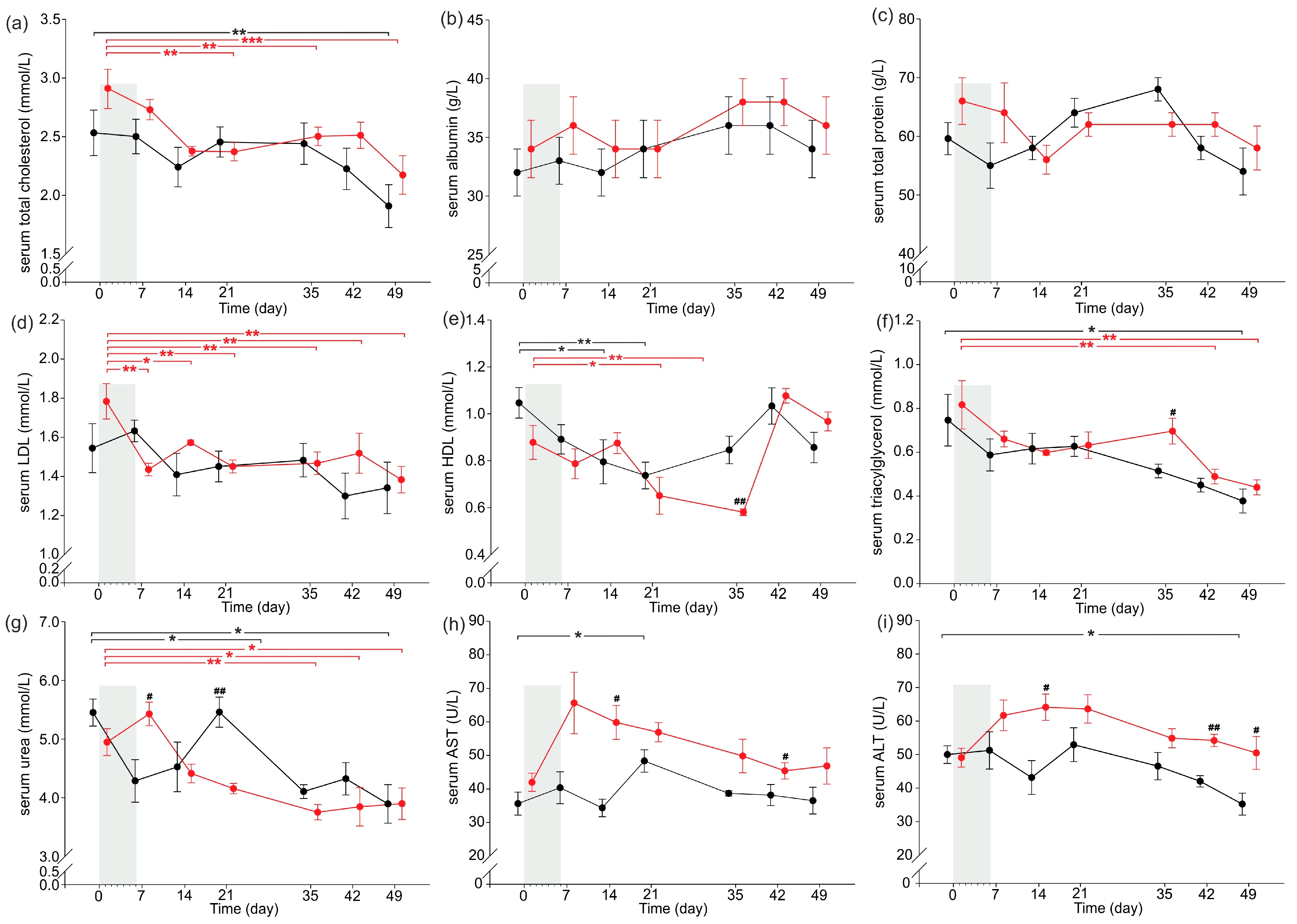
2.2. Serum Nutritional and Other Basal Biochemical Parameters
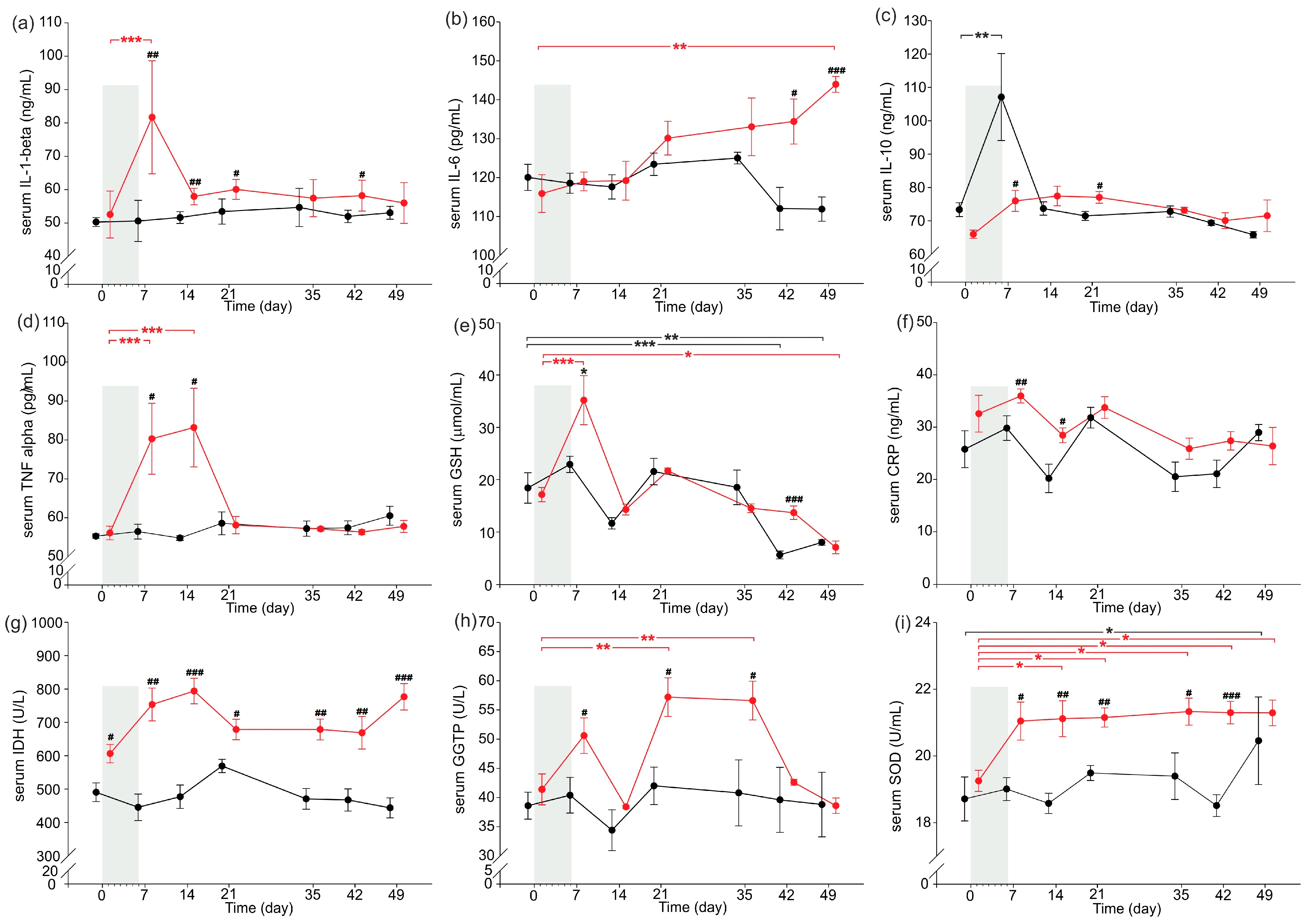
2.3. Inflammatory Parameters
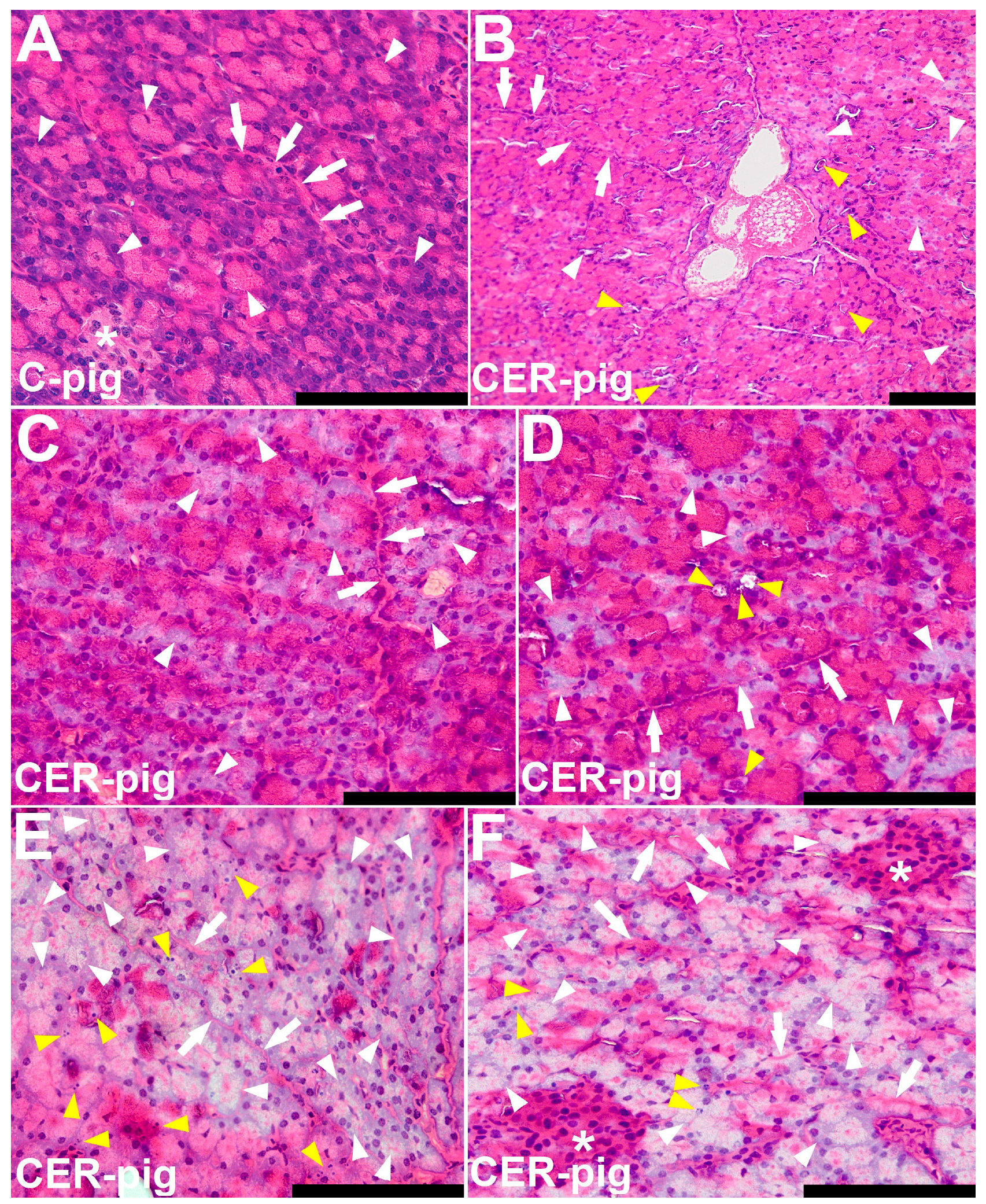
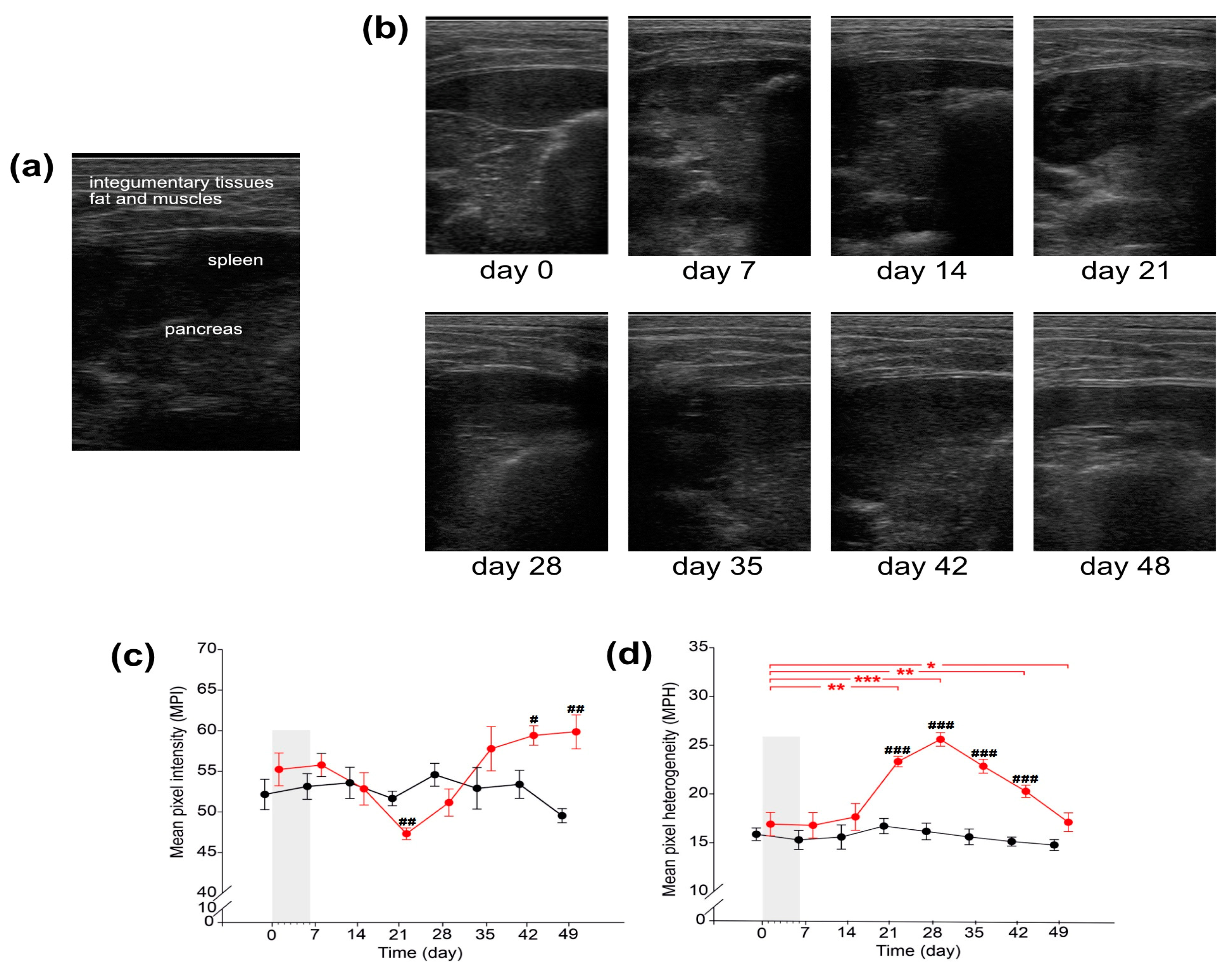
2.4. Pathomorphological Description of Pancreatic Samples
2.5. Pathomorphological Description of Pancreas Based on USG Examination
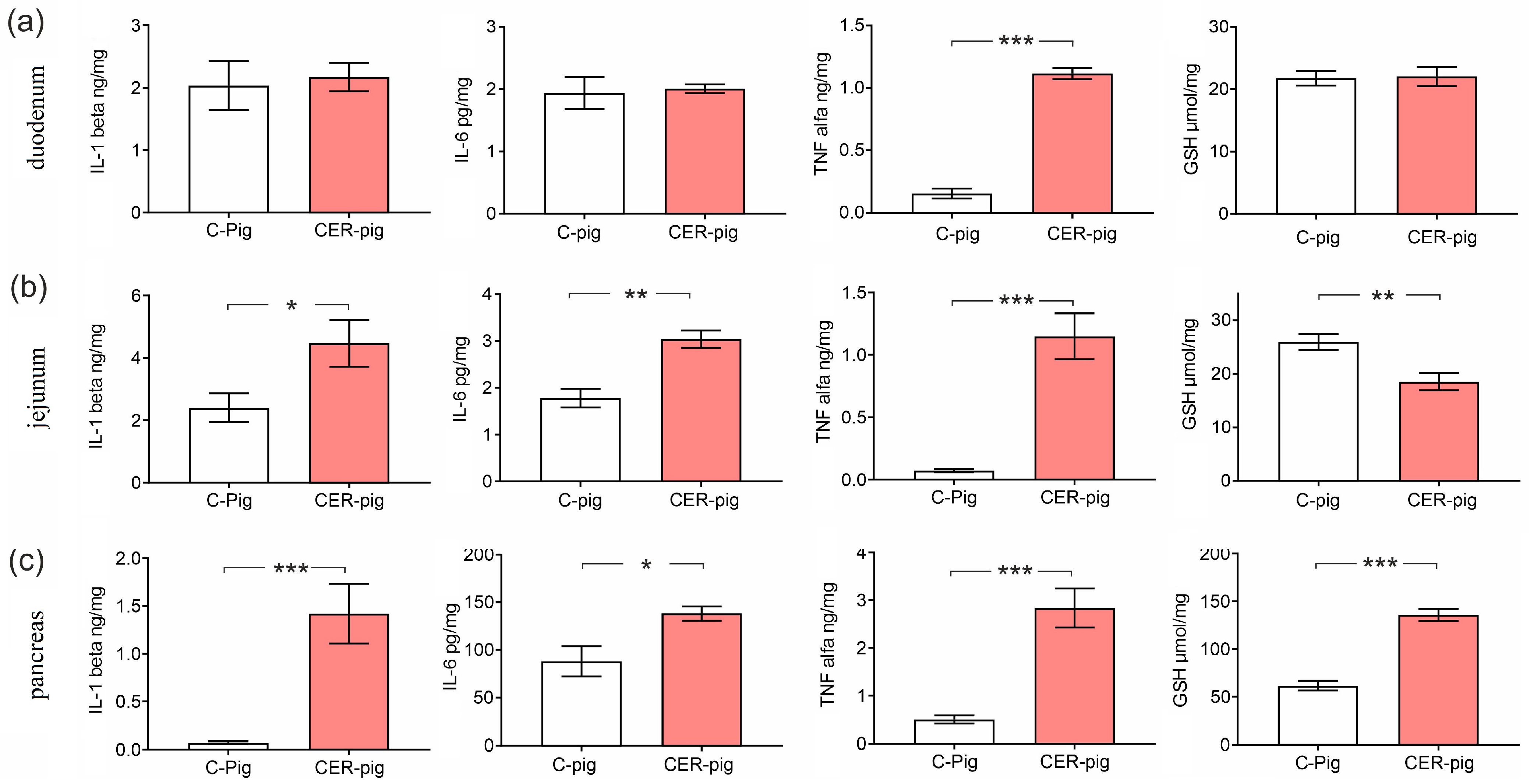
2.6. Interleukins and Reduced GSH in Tissues
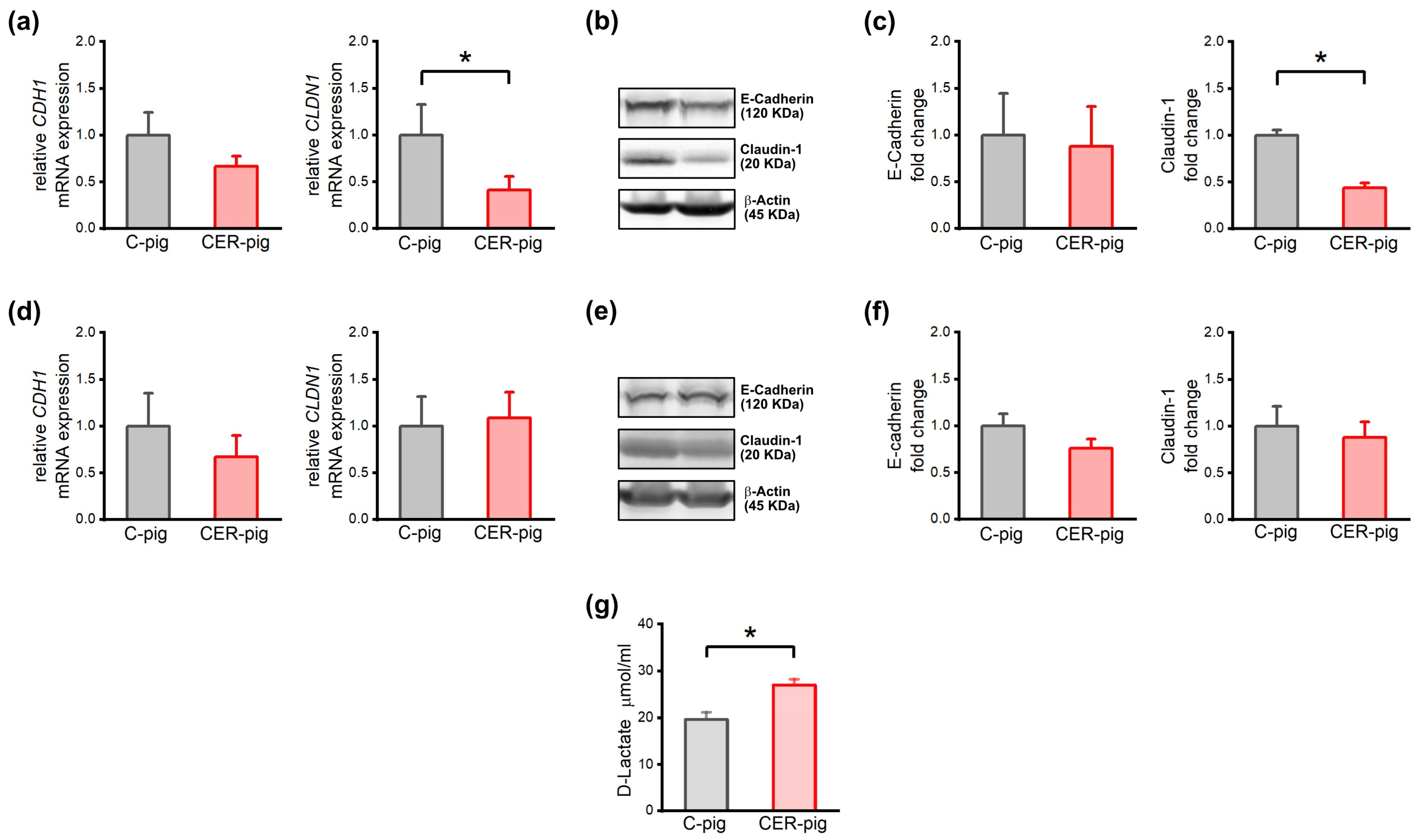
2.7. Expression of Intestinal Barrier Proteins and Their Genes and Intestinal Barrier Integrity
3. Discussion
4. Materials and Methods
4.1. Ethics
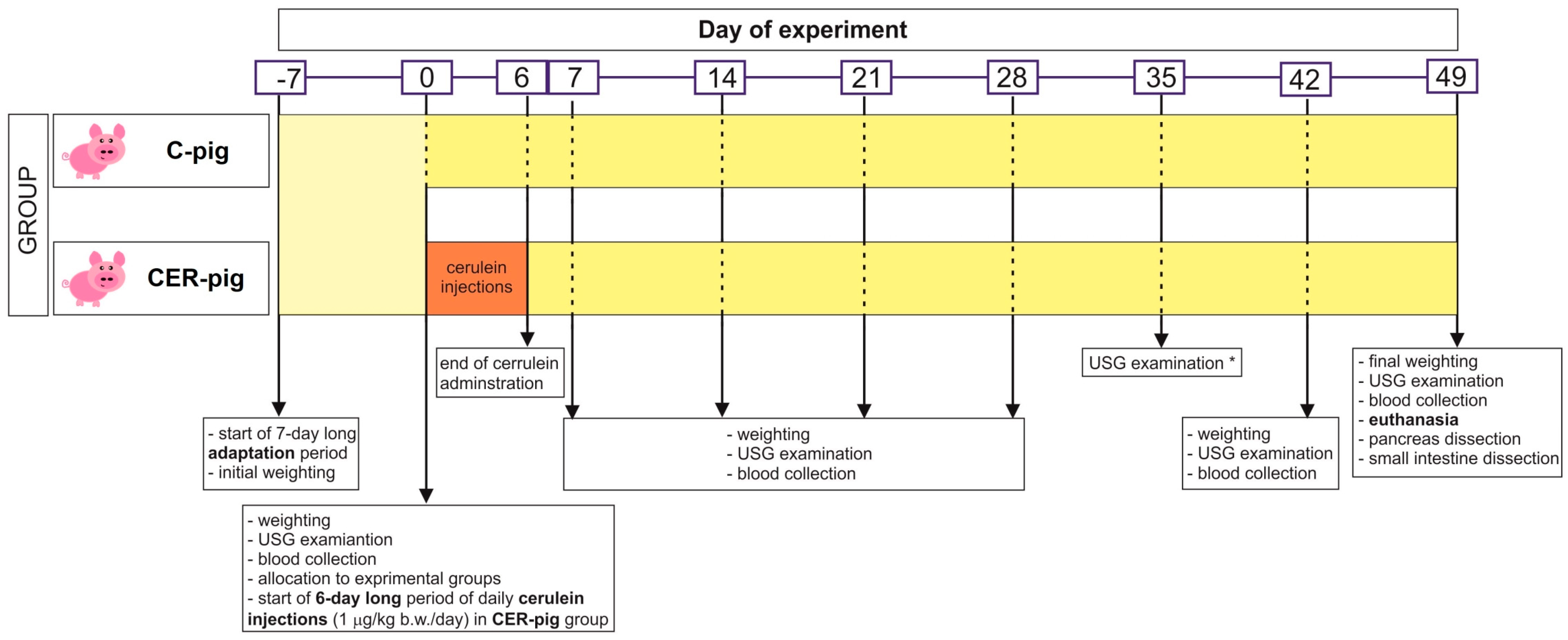
4.2. Animals and Experimental Design
4.3. USG Pancreas Examination
4.4. Inflammatory Parameters, Serum Nutritional and Other Basal Biochemical Parameters
4.5. Pancreas Histopathology
4.6. RT-qPCR Analysis of mRNA Expression of Genes Encoding Intestinal Barrier Proteins
4.7. Western Blot Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Pham, A.; Forsmark, C. Chronic Pancreatitis: Review and Update of Etiology, Risk Factors, and Management. F1000Reserch 2018, 7, 607. [Google Scholar] [CrossRef] [PubMed]
- Forsmark, C.E. Diagnosis and Management of Exocrine Pancreatic Insufficiency. Curr. Treat. Options Gastroenterol. 2018, 16, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Stram, M.; Liu, S.; Singhi, A.D. Chronic Pancreatitis. Surg. Pathol. Clin. 2016, 9, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Balázs, A.; Balla, Z.; Kui, B.; Maléth, J.; Rakonczay, Z.; Duerr, J.; Zhou-Suckow, Z.; Schatterny, J.; Sendler, M.; Mayerle, J.; et al. Ductal Mucus Obstruction and Reduced Fluid Secretion Are Early Defects in Chronic Pancreatitis. Front. Physiol. 2018, 9, 632. [Google Scholar] [CrossRef]
- Barry, K. Chronic Pancreatitis: Diagnosis and Treatment. Am. Fam. Physician 2018, 97, 385–393. [Google Scholar]
- Yadav, D.; Park, W.G.; Fogel, E.L.; Li, L.; Chari, S.T.; Feng, Z.; Fisher, W.E.; Forsmark, C.E.; Jeon, C.Y.; Habtezion, A.; et al. PROspective Evaluation of Chronic Pancreatitis for EpidEmiologic and Translational StuDies: Rationale and Study Design for PROCEED From the Consortium for the Study of Chronic Pancreatitis, Diabetes, and Pancreatic Cancer. Pancreas 2018, 47, 1229–1238. [Google Scholar] [CrossRef]
- Lerch, M.M.; Gorelick, F.S. Models of Acute and Chronic Pancreatitis. Gastroenterology 2013, 144, 1180–1193. [Google Scholar] [CrossRef]
- Kichler, A.; Jang, S. Chronic Pancreatitis: Epidemiology, Diagnosis, and Management Updates. Drugs 2020, 80, 1155–1168. [Google Scholar] [CrossRef]
- Leal, A.S.; Liby, K.T. Murine Models of Pancreatitis Leading to the Development of Pancreatic Cancer. Curr. Protoc. Pharmacol. 2018, 83, e48. [Google Scholar] [CrossRef]
- Gorelick, F.S.; Lerch, M.M. Do Animal Models of Acute Pancreatitis Reproduce Human Disease? Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 251–262. [Google Scholar] [CrossRef]
- Grupp, K.; Erbes, J.; Poppe, A.; Wodack, K.; Gocht, A.; Trepte, C.; Havel, J.; Mann, O.; Izbicki, J.R.; Bachmann, K. Melatonin Treatment of Pigs with Acute Pancreatitis Reduces Inflammatory Reaction of Pancreatic Tissue and Enhances Fitness Score of Pigs: Experimental Research. World J. Emerg. Surg. 2019, 14, 18. [Google Scholar] [CrossRef]
- Klauss, S.; Schorn, S.; Teller, S.; Steenfadt, H.; Friess, H.; Ceyhan, G.O.; Demir, I.E. Genetically Induced vs. Classical Animal Models of Chronic Pancreatitis: A Critical Comparison. FASEB J. 2018, 32, 5778–5792. [Google Scholar] [CrossRef]
- Quesada, R.; Andaluz, A.; Cáceres, M.; Moll, X.; Iglesias, M.; Dorcaratto, D.; Poves, I.; Berjano, E.; Grande, L.; Burdío, F. Long-Term Evolution of Acinar-to-Ductal Metaplasia and β-Cell Mass after Radiofrequency-Assisted Transection of the Pancreas in a Controlled Large Animal Model. Pancreatology 2016, 16, 38–43. [Google Scholar] [CrossRef]
- Roura, E.; Koopmans, S.-J.; Lallès, J.-P.; Le Huerou-Luron, I.; de Jager, N.; Schuurman, T.; Val-Laillet, D. Critical Review Evaluating the Pig as a Model for Human Nutritional Physiology. Nutr. Res. Rev. 2016, 29, 60–90. [Google Scholar] [CrossRef]
- Swindle, M.; Smith, A.C. Comparative Anatomy and Physiology of the Pig. Scand. J. Lab. Anim. Sci. 1998, 25, 11–21. [Google Scholar]
- Fasullo, M.; Omer, E.; Kaspar, M. Sarcopenia in Chronic Pancreatitis—Prevalence, Diagnosis, Mechanisms and Potential Therapies. Curr. Gastroenterol. Rep. 2022, 24, 53–63. [Google Scholar] [CrossRef]
- Kuan, L.L.; Dennison, A.R.; Garcea, G. Prevalence and Impact of Sarcopenia in Chronic Pancreatitis: A Review of the Literature. World J. Surg. 2021, 45, 590–597. [Google Scholar] [CrossRef]
- Machicado, J.D.; Yadav, D. Epidemiology of Recurrent Acute and Chronic Pancreatitis: Similarities and Differences. Dig. Dis. Sci. 2017, 62, 1683–1691. [Google Scholar] [CrossRef]
- Majumder, S.; Chari, S.T. Chronic Pancreatitis. Lancet 2016, 387, 1957–1966. [Google Scholar] [CrossRef]
- Erspamer, V. Progress Report: Caerulein. Gut 1970, 11, 79–87. [Google Scholar] [CrossRef]
- Bertaccini, G.; De Caro, G.; Endean, R.; Erspamer, V.; Impicciatore, M. The Actions of Caerulein on the Smooth Muscle of the Gastrointestinal Tract and the Gall Bladder. Br. J. Pharmacol. 1968, 34, 291–310. [Google Scholar] [CrossRef]
- Vincent, M.E.; Wetzner, S.M.; Robbins, A.H. Pharmacology, Clinical Uses, and Adverse Effects of Ceruletide, A Cholecystokinetic Agent. Pharmacother. J. Hum. Pharmacol. Drug Ther. 1982, 2, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Ganzina, F.; Santamaria, A. Caerulein (Ceruletide). A Review. Acta Gastroenterol. Belg. 1976, 39, 169–185. [Google Scholar] [PubMed]
- Watanabe, O.; Baccino, F.M.; Steer, M.L.; Meldolesi, J. Supramaximal Caerulein Stimulation and Ultrastructure of Rat Pancreatic Acinar Cell: Early Morphological Changes during Development of Experimental Pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 1984, 246, G457–G467. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, Y. Effect of Tetragastrin, Caerulein and Pancreozymin on the Enzyme Secretion of the Rabbit Pancreas in Vitro and in Vivo. Jpn. J. Pharmacol. 1972, 22, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Dockray, G.J. The Action of Secretin, Cholecystokinin-Pancreozymin and Caerulein on Pancreatic Secretion in the Rat. J. Physiol. 1972, 225, 679–692. [Google Scholar] [CrossRef]
- Coruzzi, G.; Bertaccini, G.; de Carstiglione, R.; Perseo, G. Stimulatory Action of (β-Asp9) Ceruletide on Dog Pancreatic Exocrine Secretion. Peptides 1985, 6, 29–31. [Google Scholar] [CrossRef]
- Lampel, M.; Kern, H.F. Acute Interstitial Pancreatitis in the Rat Induced by Excessive Doses of a Pancreatic Secretagogue. Virchows Arch. A Path. Anat. Histol. 1977, 373, 97–117. [Google Scholar] [CrossRef]
- Willemer, S.; Elsässer, H.-P.; Adler, G. Hormone-Induced Pancreatitis. Eur. Surg. Res. 1992, 24, 29–39. [Google Scholar] [CrossRef]
- Yu, J.H.; Lim, J.W.; Kim, H. Altered Gene Expression in Cerulein-Stimulated Pancreatic Acinar Cells: Pathologic Mechanism of Acute Pancreatitis. Korean J. Physiol. Pharm. 2009, 13, 409. [Google Scholar] [CrossRef]
- Yang, X.; Yao, L.; Fu, X.; Mukherjee, R.; Xia, Q.; Jakubowska, M.A.; Ferdek, P.E.; Huang, W. Experimental Acute Pancreatitis Models: History, Current Status, and Role in Translational Research. Front. Physiol. 2020, 11, 614591. [Google Scholar] [CrossRef]
- Bieger, W.; Seybold, J.; Kern, H.F. Studies on Intracellular Transport of Secretory Proteins in the Rat Exocrine Pancreas: V. Kinetic Studies on Accelerated Transport Following Caerulein Infusion in Vivo. Cell Tissue Res. 1976, 170, 203–219. [Google Scholar] [CrossRef]
- Bansod, S.; Aslam Saifi, M.; Khurana, A.; Godugu, C. Nimbolide Abrogates Cerulein-Induced Chronic Pancreatitis by Modulating β-Catenin/Smad in a Sirtuin-Dependent Way. Pharmacol. Res. 2020, 156, 104756. [Google Scholar] [CrossRef]
- Saluja, A.K.; Dudeja, V. Relevance of Animal Models of Pancreatic Cancer and Pancreatitis to Human Disease. Gastroenterology 2013, 144, 1194–1198. [Google Scholar] [CrossRef]
- Zhan, X.; Wang, F.; Bi, Y.; Ji, B. Animal Models of Gastrointestinal and Liver Diseases. Animal Models of Acute and Chronic Pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G343–G355. [Google Scholar] [CrossRef]
- Aghdassi, A.A.; Mayerle, J.; Christochowitz, S.; Weiss, F.U.; Sendler, M.; Lerch, M.M. Animal Models for Investigating Chronic Pancreatitis. Fibrogenesis Tissue Repair. 2011, 4, 26. [Google Scholar] [CrossRef]
- Demir, I.E.; Friess, H.; Ceyhan, G.O. Neural Plasticity in Pancreatitis and Pancreatic Cancer. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 649–659. [Google Scholar] [CrossRef]
- Schneider, A.; Whitcomb, D.C. Hereditary Pancreatitis: A Model for Inflammatory Diseases of the Pancreas. Best Pract. Res. Clin. Gastroenterol. 2002, 16, 347–363. [Google Scholar] [CrossRef]
- Foitzik, T.; Hotz, H.G.; Eibl, G.; Buhr, H.J. Experimental Models of Acute Pancreatitis: Are They Suitable for Evaluating Therapy? Int. J. Color. Dis. 2000, 15, 127–135. [Google Scholar] [CrossRef]
- Völzke, H.; Baumeister, S.E.; Alte, D.; Hoffmann, W.; Schwahn, C.; Simon, P.; John, U.; Lerch, M.M. Independent Risk Factors for Gallstone Formation in a Region with High Cholelithiasis Prevalence. Digestion 2005, 71, 97–105. [Google Scholar] [CrossRef]
- Whitcomb, D.C.; Frulloni, L.; Garg, P.; Greer, J.B.; Schneider, A.; Yadav, D.; Shimosegawa, T. Chronic Pancreatitis: An International Draft Consensus Proposal for a New Mechanistic Definition. Pancreatology 2016, 16, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Sah, R.P.; Dawra, R.K.; Saluja, A.K. New Insights into the Pathogenesis of Pancreatitis. Curr. Opin. Gastroenterol. 2013, 29, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K. Mechanisms of Pancreatic Fibrosis and Applications to the Treatment of Chronic Pancreatitis. J. Gastroenterol. 2008, 43, 823–832. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Noguera, A.; D’Onofrio, M. Ultrasonography of the Pancreas. 1. Conventional Imaging. Abdom. Imaging 2007, 32, 136–149. [Google Scholar] [CrossRef]
- Newcom, D.W.; Baas, T.J.; Lampe, J.F. Prediction of Intramuscular Fat Percentage in Live Swine Using Real-Time Ultrasound1. J. Anim. Sci. 2002, 80, 3046–3052. [Google Scholar] [CrossRef]
- Ayuso, D.; González, A.; Hernández, F.; Corral, J.M.; Izquierdo, M. Prediction of Carcass Composition, Ham and Foreleg Weights, and Lean Meat Yields of Iberian Pigs Using Ultrasound Measurements in Live Animals1. J. Anim. Sci. 2013, 91, 1884–1892. [Google Scholar] [CrossRef]
- Scuro, L.A.; Cavallini, G.; Benini, L.; Brocco, G.; Bovo, P.; Riela, A.; Togni, M.; Cataudella, G.; Bassi, C.; Pederzoli, P. Pancreatic Calcifications in Patients with Chronic Pancreatitis. A Sign of Long-Lasting or Severe Disease? Int. J. Pancreatol. 1990, 6, 139–150. [Google Scholar] [CrossRef]
- Sarner, M.; Cotton, P.B. Classification of Pancreatitis. Gut 1984, 25, 756–759. [Google Scholar] [CrossRef]
- Braganza, J.M.; Lee, S.H.; McCloy, R.F.; McMahon, M.J. Chronic Pancreatitis. Lancet 2011, 377, 1184–1197. [Google Scholar] [CrossRef]
- Steer, M.L.; Waxman, I.; Freedman, S. Chronic Pancreatitis. N. Engl. J. Med. 1995, 332, 1482–1490. [Google Scholar] [CrossRef]
- Logsdon, C.D.; Ji, B. The Role of Protein Synthesis and Digestive Enzymes in Acinar Cell Injury. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 362–370. [Google Scholar] [CrossRef]
- Watanabe, T.; Kudo, M.; Strober, W. Immunopathogenesis of Pancreatitis. Mucosal. Immunol. 2017, 10, 283–298. [Google Scholar] [CrossRef]
- Gukovskaya, A.S.; Gukovsky, I.; Zaninovic, V.; Song, M.; Sandoval, D.; Gukovsky, S.; Pandol, S.J. Pancreatic Acinar Cells Produce, Release, and Respond to Tumor Necrosis Factor-Alpha. Role in Regulating Cell Death and Pancreatitis. J. Clin. Investig. 1997, 100, 1853–1862. [Google Scholar] [CrossRef]
- Sendler, M.; Dummer, A.; Weiss, F.U.; Krüger, B.; Wartmann, T.; Scharffetter-Kochanek, K.; van Rooijen, N.; Malla, S.R.; Aghdassi, A.; Halangk, W.; et al. Tumour Necrosis Factor α Secretion Induces Protease Activation and Acinar Cell Necrosis in Acute Experimental Pancreatitis in Mice. Gut 2013, 62, 430–439. [Google Scholar] [CrossRef]
- Malmstrøm, M.L.; Hansen, M.B.; Andersen, A.M.; Ersbøll, A.K.; Nielsen, O.H.; Jørgensen, L.N.; Novovic, S. Cytokines and Organ Failure in Acute Pancreatitis: Inflammatory Response in Acute Pancreatitis. Pancreas 2012, 41, 271–277. [Google Scholar] [CrossRef]
- Castell, J.V.; Gómez-Lechón, M.J.; David, M.; Andus, T.; Geiger, T.; Trullenque, R.; Fabra, R.; Heinrich, P.C. Interleukin-6 Is the Major Regulator of Acute Phase Protein Synthesis in Adult Human Hepatocytes. FEBS Lett. 1989, 242, 237–239. [Google Scholar] [CrossRef]
- Hoque, R.; Malik, A.F.; Gorelick, F.; Mehal, W.Z. Sterile Inflammatory Response in Acute Pancreatitis. Pancreas 2012, 41, 353–357. [Google Scholar] [CrossRef]
- Castellheim, A.; Brekke, O.-L.; Espevik, T.; Harboe, M.; Mollnes, T.E. Innate Immune Responses to Danger Signals in Systemic Inflammatory Response Syndrome and Sepsis. Scand. J. Immunol. 2009, 69, 479–491. [Google Scholar] [CrossRef]
- Tao, J.; Gong, D.; Ji, D.; Xu, B.; Liu, Z.; Li, L. Improvement of Monocyte Secretion Function in a Porcine Pancreatitis Model by Continuous Dose-Dependent Veno-Venous Hemofiltration. Int. J. Artif. Organs 2008, 31, 716–721. [Google Scholar] [CrossRef]
- Pezzilli, R.; Melzi D’eril, G.V.; Morselli-Labate, A.M.; Merlini, G.; Barakat, B.; Bosoni, T. Serum Amyloid A, Procalcitonin, and C-Reactive Protein in Early Assessment of Severity of Acute Pancreatitis. Dig. Dis. Sci. 2000, 45, 1072–1078. [Google Scholar] [CrossRef]
- Yin, X.; Xu, J.; Zhang, Q.; Yang, L.; Duan, Y. Quantification Analysis of Lactate Dehydrogenase and C-Reactive Protein in Evaluation of the Severity and Prognosis of the Acute Pancreatitis. Cell Mol. Biol. (Noisy-Le-Grand) 2020, 66, 122–125. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Xiong, J.; Zhang, Y.; Peng, T.; Huang, M.; Lin, Y.; Guo, Y.; Wu, H.; Wang, C. Serum Lactate Dehydrogenase Is Predictive of Persistent Organ Failure in Acute Pancreatitis. J. Crit. Care 2017, 41, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Tiribelli, C.; Ostrow, J.D. New Concepts in Bilirubin and Jaundice: Report of the Third International Bilirubin Workshop, April 6–8, 1995, Trieste, Italy. Hepatology 1996, 24, 1296–1311. [Google Scholar] [CrossRef] [PubMed]
- Kiriyama, S.; Gabata, T.; Takada, T.; Hirata, K.; Yoshida, M.; Mayumi, T.; Hirota, M.; Kadoya, M.; Yamanouchi, E.; Hattori, T.; et al. New Diagnostic Criteria of Acute Pancreatitis. J. Hepato Biliary Pancreat Sci. 2010, 17, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Witt, H.; Apte, M.V.; Keim, V.; Wilson, J.S. Chronic Pancreatitis: Challenges and Advances in Pathogenesis, Genetics, Diagnosis, and Therapy. Gastroenterology 2007, 132, 1557–1573. [Google Scholar] [CrossRef]
- Shintakuya, R.; Uemura, K.; Murakami, Y.; Kondo, N.; Nakagawa, N.; Urabe, K.; Okano, K.; Awai, K.; Higaki, T.; Sueda, T. Sarcopenia Is Closely Associated with Pancreatic Exocrine Insufficiency in Patients with Pancreatic Disease. Pancreatology 2017, 17, 70–75. [Google Scholar] [CrossRef]
- Su, S.-B.; Motoo, Y.; Xie, M.-J.; Mouri, H.; Asayama, K.; Sawabu, N. Superoxide Dismutase Is Induced During Rat Pancreatic Acinar Cell Injury. Pancreas 2002, 24, 146–152. [Google Scholar] [CrossRef]
- Guice, K.S.; Miller, D.E.; Oldham, K.T.; Townsend, C.M.; Thompson, J.C. Superoxide Dismutase and Catalase: A Possible Role in Established Pancreatitis. Am. J. Surg. 1986, 151, 163–169. [Google Scholar] [CrossRef]
- Zheng, X.; Li, L.; Zhu, Y.; Huang, X.; Zhang, Y.; Yu, B.; He, W.; Lv, N. Superoxide Dismutase Predicts Persistent Circulation Failure and Mortality in the Early Stage of Acute Pancreatitis. Dig. Dis. Sci. 2020, 65, 3551–3557. [Google Scholar] [CrossRef]
- Kodydkova, J.; Vavrova, L.; Stankova, B.; Macasek, J.; Krechler, T.; Zak, A. Antioxidant Status and Oxidative Stress Markers in Pancreatic Cancer and Chronic Pancreatitis. Pancreas 2013, 42, 614–621. [Google Scholar] [CrossRef]
- Khan, J.; Solakivi, T.; Seppänen, H.; Lappalainen-Lehto, R.; Järvinen, S.; Ronkainen, J.; Sand, J.; Nordback, I. Serum Lipid and Fatty Acid Profiles Are Highly Changed in Patients with Alcohol Induced Acute Pancreatitis. Pancreatology 2012, 12, 44–48. [Google Scholar] [CrossRef]
- Ignacio de Ulíbarri, J.; González-Madroño, A.; de Villar, N.G.P.; González, P.; González, B.; Mancha, A.; Rodríguez, F.; Fernández, G. CONUT: A Tool for Controlling Nutritional Status. First Validation in a Hospital Population. Nutr. Hosp. 2005, 20, 38–45. [Google Scholar]
- Wan, J.; He, W.; Zhu, Y.; Zhu, Y.; Zeng, H.; Liu, P.; Xia, L.; Lu, N. Stratified Analysis and Clinical Significance of Elevated Serum Triglyceride Levels in Early Acute Pancreatitis: A Retrospective Study. Lipids. Health Dis. 2017, 16, 124. [Google Scholar] [CrossRef]
- Ozerlat, I. Blood Urea Nitrogen Levels Predict Mortality Risk in Acute Pancreatitis. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 359. [Google Scholar] [CrossRef]
- Wu, B.U.; Bakker, O.J.; Papachristou, G.I.; Besselink, M.G.; Repas, K.; van Santvoort, H.C.; Muddana, V.; Singh, V.K.; Whitcomb, D.C.; Gooszen, H.G.; et al. Blood Urea Nitrogen in the Early Assessment of Acute Pancreatitis: An International Validation Study. Arch Intern. Med. 2011, 171, 669–676. [Google Scholar] [CrossRef]
- Hirano, K.; Saito, T.; Mizuno, S.; Tada, M.; Sasahira, N.; Isayama, H.; Matsukawa, M.; Umefune, G.; Akiyama, D.; Saito, K.; et al. Total Cholesterol Level for Assessing Pancreatic Insufficiency Due to Chronic Pancreatitis. Gut Liver 2014, 8, 563–568. [Google Scholar] [CrossRef]
- Liu, H.; Li, W.; Wang, X.; Li, J.; Yu, W. Early Gut Mucosal Dysfunction in Patients With Acute Pancreatitis. Pancreas 2008, 36, 192–196. [Google Scholar] [CrossRef]
- Ammori, B. Early Increase in Intestinal Permeability in Patients with Severe Acute Pancreatitis: Correlation with Endotoxemia, Organ Failure, and Mortality. J. Gastrointest. Surg. 1999, 3, 252–262. [Google Scholar] [CrossRef]
- Hart, P.A.; Conwell, D.L. Chronic Pancreatitis: Managing a Difficult Disease. Am. J. Gastroenterol. 2020, 115, 49–55. [Google Scholar] [CrossRef]
- Günzel, D.; Fromm, M. Claudins and Other Tight Junction Proteins. In Comprehensive Physiology; Terjung, R., Ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 1819–1852. ISBN 978-0-470-65071-4. [Google Scholar]
- Poritz, L.S.; Harris, L.R.; Kelly, A.A.; Koltun, W.A. Increase in the Tight Junction Protein Claudin-1 in Intestinal Inflammation. Dig Dis. Sci. 2011, 56, 2802. [Google Scholar] [CrossRef]
- Kyuno, D. Targeting Tight Junctions during Epithelial to Mesenchymal Transition in Human Pancreatic Cancer. WJG 2014, 20, 10813. [Google Scholar] [CrossRef] [PubMed]
- van Roy, F.; Berx, G. The Cell-Cell Adhesion Molecule E-Cadherin. Cell. Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef] [PubMed]
- Kaneta, Y.; Sato, T.; Hikiba, Y.; Sugimori, M.; Sue, S.; Kaneko, H.; Irie, K.; Sasaki, T.; Kondo, M.; Chuma, M.; et al. Loss of Pancreatic E-Cadherin Causes Pancreatitis-Like Changes and Contributes to Carcinogenesis. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhu, S.; Wang, B.; Li, X.; Liu, Y.; Qin, Q.; Gong, Q.; Niu, Y.; Xiang, C.; Chen, J.; et al. Chronic Pancreatitis and Pancreatic Cancer Demonstrate Active Epithelial–Mesenchymal Transition Profile, Regulated by MiR-217-SIRT1 Pathway. Cancer Lett. 2014, 355, 184–191. [Google Scholar] [CrossRef]
- Avril, M.; DePaolo, R.W. “Driver-Passenger” Bacteria and Their Metabolites in the Pathogenesis of Colorectal Cancer. Gut Microbes 2021, 13, 1941710. [Google Scholar] [CrossRef]
- Obafemi, T.F.; Yu, P.; Li, J.; Davis, J.M.; Liu, K.; Cheng, B.; Zhao, X.; Shen, Q.; Younes, M.; Ko, T.C.; et al. Comparable Responses in Male and Female Mice to Cerulein-Induced Chronic Pancreatic Injury and Recovery. J. Pancreas 2018, 19, 236–243. [Google Scholar]
- Freitag, M.; Standl, T.G.; Kleinhans, H.; Gottschalk, A.; Mann, O.; Rempf, C.; Bachmann, K.; Gocht, A.; Petri, S.; Izbicki, J.R.; et al. Improvement of Impaired Microcirculation and Tissue Oxygenation by Hemodilution with Hydroxyethyl Starch plus Cell-Free Hemoglobin in Acute Porcine Pancreatitis. Pancreatology 2006, 6, 232–239. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Swine: Eleventh Revised Edition; The National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-48903-4.
- Grela, E.; Skomiał, J.; Raj, S.; Skiba, G.; Święch, E.; Fandrejewski, H.; Czech, A.; Frankiewicz, A.; Świątkiewicz, M. Zalecenia Żywieniowe i Wartość Pokarmowa Pasz dla Świń: Monografia; Wydanie III Uzupełnione z Oprogramowaniem; Instytut Fizjologii i Żywienia Zwierząt im; Jana Kielanowskiego Polskiej Akademii Nauk: Jabłonna, Poland, 2020; ISBN 978-83-951612-7-8. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Yun, G.; Kim, Y.H.; Lee, Y.J.; Kim, B.; Hwang, J.-H.; Choi, D.J. Tumor Heterogeneity of Pancreas Head Cancer Assessed by CT Texture Analysis: Association with Survival Outcomes after Curative Resection. Sci. Rep. 2018, 8, 7226. [Google Scholar] [CrossRef]
- Piórkowska, K.; Oczkowicz, M.; Różycki, M.; Ropka-Molik, K.; Kajtoch, A.P.-. Novel Porcine Housekeeping Genes for Real-Time RT-PCR Experiments Normalization in Adipose Tissue: Assessment of Leptin MRNA Quantity in Different Pig Breeds. Meat Sci. 2011, 87, 191–195. [Google Scholar] [CrossRef]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. New Long-Non Coding RNAs Related to Fat Deposition Based on Pig Model. Ann. Anim. Sci. 2022, 22, 1211–1224. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tomaszewska, E.; Świątkiewicz, M.; Muszyński, S.; Donaldson, J.; Ropka-Molik, K.; Arciszewski, M.B.; Murawski, M.; Schwarz, T.; Dobrowolski, P.; Szymańczyk, S.; et al. Repetitive Cerulein-Induced Chronic Pancreatitis in Growing Pigs—A Pilot Study. Int. J. Mol. Sci. 2023, 24, 7715. https://doi.org/10.3390/ijms24097715
Tomaszewska E, Świątkiewicz M, Muszyński S, Donaldson J, Ropka-Molik K, Arciszewski MB, Murawski M, Schwarz T, Dobrowolski P, Szymańczyk S, et al. Repetitive Cerulein-Induced Chronic Pancreatitis in Growing Pigs—A Pilot Study. International Journal of Molecular Sciences. 2023; 24(9):7715. https://doi.org/10.3390/ijms24097715
Chicago/Turabian StyleTomaszewska, Ewa, Małgorzata Świątkiewicz, Siemowit Muszyński, Janine Donaldson, Katarzyna Ropka-Molik, Marcin B. Arciszewski, Maciej Murawski, Tomasz Schwarz, Piotr Dobrowolski, Sylwia Szymańczyk, and et al. 2023. "Repetitive Cerulein-Induced Chronic Pancreatitis in Growing Pigs—A Pilot Study" International Journal of Molecular Sciences 24, no. 9: 7715. https://doi.org/10.3390/ijms24097715