
Diagnostic and Prognostic Role of Extracellular Vesicles in Pancreatic Cancer: Current Evidence and Future Perspectives
 , , ,
, , ,
Abstract
:1. Introduction
2. Extracellular Vesicles in Pancreatic Cancer: Role in Carcinogenesis, Cancer Progression, and Chemoresistance
3. Proteomics of Extracellular Vesicles in Pancreatic Cancer
4. EV-Derived miRNAs as Potential Biomarkers for Pancreatic Cancer
5. EVs and EV-Derived miRNAs in Pancreatic Precancerous Lesions
6. Future Perspectives
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic Adenocarcinoma. N. Engl. J. Med. 2014, 371, 2140–2141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2020. CA A Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting Cancer Incidence and Deaths to 2030: The Unexpected Burden of Thyroid, Liver, and Pancreas Cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, H.C.; Wallace, M.B.; Storz, P. Early Detection and Imaging Strategies to Reveal and Target Developing Pancreatic Cancer. Expert Rev. Anticancer Ther. 2020, 20, 81–83. [Google Scholar] [CrossRef] [Green Version]
- McGuigan, A.; Kelly, P.; Turkington, R.C.; Jones, C.; Coleman, H.G.; McCain, R.S. Pancreatic Cancer: A Review of Clinical Diagnosis, Epidemiology, Treatment and Outcomes. World J. Gastroenterol. 2018, 24, 4846–4861. [Google Scholar] [CrossRef] [PubMed]
- Goonetilleke, K.S.; Siriwardena, A.K. Systematic Review of Carbohydrate Antigen (CA 19-9) as a Biochemical Marker in the Diagnosis of Pancreatic Cancer. Eur. J. Surg. Oncol. 2007, 33, 266–270. [Google Scholar] [CrossRef]
- Gulla, A.; Hashimoto, D.; Wagner, D.; Damaseviciute, R.; Strupas, K.; Satoi, S. Interdisciplinary Approach of Establishing PDAC Resectability: Biochemical, Radiological and NAT Regimen Prognostic Factors—Literature Review. Medicina 2022, 58, 756. [Google Scholar] [CrossRef]
- Yang, J.; Xu, R.; Wang, C.; Qiu, J.; Ren, B.; You, L. Early Screening and Diagnosis Strategies of Pancreatic Cancer: A Comprehensive Review. Cancer Commun. 2021, 41, 1257–1274. [Google Scholar] [CrossRef]
- Kourembanas, S. Exosomes: Vehicles of Intercellular Signaling, Biomarkers, and Vectors of Cell Therapy. Annu. Rev. Physiol. 2015, 77, 13–27. [Google Scholar] [CrossRef]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-R.; Man, Q.-W.; Gao, X.; Lin, H.; Wang, J.; Su, F.-C.; Wang, H.-Q.; Bu, L.-L.; Liu, B.; Chen, G. Tissue-Derived Extracellular Vesicles in Cancers and Non-Cancer Diseases: Present and Future. J. Extracell Vesicles 2021, 10, e12175. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, K.; Liu, Y.; Xu, Y.; Zhang, F.; Yang, H.; Liu, J.; Pan, T.; Chen, J.; Wu, M.; et al. Exosomes Mediate the Cell-to-Cell Transmission of IFN-α-Induced Antiviral Activity. Nat. Immunol. 2013, 14, 793–803. [Google Scholar] [CrossRef] [PubMed]
- Waqas, M.Y.; Javid, M.A.; Nazir, M.M.; Niaz, N.; Nisar, M.F.; Manzoor, Z.; Bhatti, S.A.; Hameed, S.; Khaliq, M.H. Extracellular Vesicles and Exosome: Insight from Physiological Regulatory Perspectives. J. Physiol. Biochem. 2022, 78, 573–580. [Google Scholar] [CrossRef]
- Lee, Y.; El Andaloussi, S.; Wood, M.J.A. Exosomes and Microvesicles: Extracellular Vesicles for Genetic Information Transfer and Gene Therapy. Hum. Mol. Genet. 2012, 21, R125–R134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.A.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of Exosomes by Differential Centrifugation: Theoretical Analysis of a Commonly Used Protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef] [Green Version]
- Greening, D.W.; Xu, R.; Ji, H.; Tauro, B.J.; Simpson, R.J. A Protocol for Exosome Isolation and Characterization: Evaluation of Ultracentrifugation, Density-Gradient Separation, and Immunoaffinity Capture Methods. Methods Mol. Biol. 2015, 1295, 179–209. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Monguió-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.l.; Beyer, K.; Borràs, F.E. Size-Exclusion Chromatography-Based Isolation Minimally Alters Extracellular Vesicles’ Characteristics Compared to Precipitating Agents. Sci. Rep. 2016, 6, 33641. [Google Scholar] [CrossRef] [Green Version]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.F.; Wiegmans, A.P.; Leimgruber, A.; Möller, A. Optimized Exosome Isolation Protocol for Cell Culture Supernatant and Human Plasma. J. Extracell Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef]
- Salih, M.; Zietse, R.; Hoorn, E.J. Urinary Extracellular Vesicles and the Kidney: Biomarkers and Beyond. Am. J. Physiol. Renal Physiol. 2014, 306, F1251–F1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tataruch-Weinert, D.; Musante, L.; Kretz, O.; Holthofer, H. Urinary Extracellular Vesicles for RNA Extraction: Optimization of a Protocol Devoid of Prokaryote Contamination. J. Extracell Vesicles 2016, 5, 30281. [Google Scholar] [CrossRef] [PubMed]
- Deregibus, M.C.; Figliolini, F.; D’Antico, S.; Manzini, P.M.; Pasquino, C.; De Lena, M.; Tetta, C.; Brizzi, M.F.; Camussi, G. Charge-Based Precipitation of Extracellular Vesicles. Int. J. Mol. Med. 2016, 38, 1359–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreu, Z.; Rivas, E.; Sanguino-Pascual, A.; Lamana, A.; Marazuela, M.; González-Alvaro, I.; Sánchez-Madrid, F.; de la Fuente, H.; Yáñez-Mó, M. Comparative Analysis of EV Isolation Procedures for MiRNAs Detection in Serum Samples. J. Extracell Vesicles 2016, 5, 31655. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, Z.; Lynn, K.D.; Thorpe, P.E.; Schroit, A.J. A Novel “Salting-out” Procedure for the Isolation of Tumor-Derived Exosomes. J. Immunol. Methods 2014, 407, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Gallart-Palau, X.; Serra, A.; Sze, S.K. Enrichment of Extracellular Vesicles from Tissues of the Central Nervous System by PROSPR. Mol. Neurodegener. 2016, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Han, C.; Labuz, J.M.; Kim, J.; Kim, J.; Cho, S.; Gho, Y.S.; Takayama, S.; Park, J. High-Yield Isolation of Extracellular Vesicles Using Aqueous Two-Phase System. Sci. Rep. 2015, 5, 13103. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.; Carpenter, E.; Issadore, D. Detection and Isolation of Circulating Exosomes and Microvesicles for Cancer Monitoring and Diagnostics Using Micro-/Nano-Based Devices. Analyst 2016, 141, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Nakai, W.; Yoshida, T.; Diez, D.; Miyatake, Y.; Nishibu, T.; Imawaka, N.; Naruse, K.; Sadamura, Y.; Hanayama, R. A Novel Affinity-Based Method for the Isolation of Highly Purified Extracellular Vesicles. Sci. Rep. 2016, 6, 33935. [Google Scholar] [CrossRef] [Green Version]
- Balaj, L.; Atai, N.A.; Chen, W.; Mu, D.; Tannous, B.A.; Breakefield, X.O.; Skog, J.; Maguire, C.A. Heparin Affinity Purification of Extracellular Vesicles. Sci. Rep. 2015, 5, 10266. [Google Scholar] [CrossRef]
- Knol, J.C.; de Reus, I.; Schelfhorst, T.; Beekhof, R.; de Wit, M.; Piersma, S.R.; Pham, T.V.; Smit, E.F.; Verheul, H.M.W.; Jiménez, C.R. Peptide-Mediated “miniprep” Isolation of Extracellular Vesicles Is Suitable for High-Throughput Proteomics. EuPA Open Proteom. 2016, 11, 11–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Royo, F.; Zuñiga-Garcia, P.; Sanchez-Mosquera, P.; Egia, A.; Perez, A.; Loizaga, A.; Arceo, R.; Lacasa, I.; Rabade, A.; Arrieta, E.; et al. Different EV Enrichment Methods Suitable for Clinical Settings Yield Different Subpopulations of Urinary Extracellular Vesicles from Human Samples. J. Extracell Vesicles 2016, 5, 29497. [Google Scholar] [CrossRef]
- Caponnetto, F.; Manini, I.; Skrap, M.; Palmai-Pallag, T.; Di Loreto, C.; Beltrami, A.P.; Cesselli, D.; Ferrari, E. Size-Dependent Cellular Uptake of Exosomes. Nanomedicine 2017, 13, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Costa-Silva, B.; Shen, T.-L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.; Di Giannatale, A.; Ceder, S.; et al. Tumour Exosome Integrins Determine Organotropic Metastasis. Nature 2015, 527, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R.F. Routes and Mechanisms of Extracellular Vesicle Uptake. J. Extracell Vesicles 2014, 3, 24641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [Green Version]
- Peinado, H.; Alečković, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.; Hergueta-Redondo, M.; Williams, C.; García-Santos, G.; Ghajar, C.; et al. Melanoma Exosomes Educate Bone Marrow Progenitor Cells toward a Pro-Metastatic Phenotype through MET. Nat. Med. 2012, 18, 883–891. [Google Scholar] [CrossRef] [Green Version]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic Cancer Exosomes Initiate Pre-Metastatic Niche Formation in the Liver. Nat. Cell Biol. 2015, 17, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Moeng, S.; Son, S.W.; Lee, J.S.; Lee, H.Y.; Kim, T.H.; Choi, S.Y.; Kuh, H.J.; Park, J.K. Extracellular Vesicles (EVs) and Pancreatic Cancer: From the Role of EVs to the Interference with EV-Mediated Reciprocal Communication. Biomedicines 2020, 8, 267. [Google Scholar] [CrossRef]
- Chang, C.-H.; Pauklin, S. Extracellular Vesicles in Pancreatic Cancer Progression and Therapies. Cell Death Dis. 2021, 12, 1–12. [Google Scholar] [CrossRef]
- Ruivo, C.F.; Bastos, N.; Adem, B.; Batista, I.; Duraes, C.; Melo, C.A.; Castaldo, S.A.; Campos-Laborie, F.; Moutinho-Ribeiro, P.; Morão, B.; et al. Extracellular Vesicles from Pancreatic Cancer Stem Cells Lead an Intratumor Communication Network (EVNet) to Fuel Tumour Progression. Gut 2022, 71, 2043–2068. [Google Scholar] [CrossRef] [PubMed]
- Leca, J.; Martinez, S.; Lac, S.; Nigri, J.; Secq, V.; Rubis, M.; Bressy, C.; Sergé, A.; Lavaut, M.-N.; Dusetti, N.; et al. Cancer-Associated Fibroblast-Derived Annexin A6+ Extracellular Vesicles Support Pancreatic Cancer Aggressiveness. J. Clin. Investig. 2016, 126, 4140–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, S.; Lai, L.A.; Simeone, D.M.; Dawson, D.W.; Yan, Y.; Crnogorac-Jurcevic, T.; Chen, R.; Brentnall, T.A. Proteomes of Extracellular Vesicles From Pancreatic Cancer Cells and Cancer-Associated Fibroblasts. Pancreas 2022, 51, 790–799. [Google Scholar] [CrossRef]
- Takahashi, K.; Taniue, K.; Ono, Y.; Fujiya, M.; Mizukami, Y.; Okumura, T. Long Non-Coding RNAs in Epithelial-Mesenchymal Transition of Pancreatic Cancer. Front. Mol. Biosci. 2021, 8, 717890. [Google Scholar] [CrossRef] [PubMed]
- Novizio, N.; Belvedere, R.; Pessolano, E.; Morello, S.; Tosco, A.; Campiglia, P.; Filippelli, A.; Petrella, A. ANXA1 Contained in EVs Regulates Macrophage Polarization in Tumor Microenvironment and Promotes Pancreatic Cancer Progression and Metastasis. Int. J. Mol. Sci. 2021, 22, 11018. [Google Scholar] [CrossRef]
- Pirlog, R.; Calin, G.A. KRAS Mutations as Essential Promoters of Lymphangiogenesis via Extracellular Vesicles in Pancreatic Cancer. J. Clin. Investig. 2022, 132, e161454. [Google Scholar] [CrossRef] [PubMed]
- AmraneDjedidi, R.; Rousseau, A.; Larsen, A.K.; Elalamy, I.; Van Dreden, P.; Gerotziafas, G.T. Extracellular Vesicles Derived from Pancreatic Cancer Cells BXPC3 or Breast Cancer Cells MCF7 Induce a Permanent Procoagulant Shift to Endothelial Cells. Thromb. Res. 2020, 187, 170–179. [Google Scholar] [CrossRef]
- Zhang, Y.-F.; Zhou, Y.-Z.; Zhang, B.; Huang, S.-F.; Li, P.-P.; He, X.-M.; Cao, G.-D.; Kang, M.-X.; Dong, X.; Wu, Y.-L. Pancreatic Cancer-Derived Exosomes Promoted Pancreatic Stellate Cells Recruitment by Pancreatic Cancer. J. Cancer 2019, 10, 4397–4407. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; He, D.; Li, L.; Zhang, S.; Wang, L.; Fan, Z.; Wang, Y. Extracellular Vesicles in Pancreatic Cancer Immune Escape: Emerging Roles and Mechanisms. Pharmacol. Res. 2022, 183, 106364. [Google Scholar] [CrossRef]
- Zhao, J.; Schlößer, H.A.; Wang, Z.; Qin, J.; Li, J.; Popp, F.; Popp, M.C.; Alakus, H.; Chon, S.-H.; Hansen, H.P.; et al. Tumor-Derived Extracellular Vesicles Inhibit Natural Killer Cell Function in Pancreatic Cancer. Cancers 2019, 11, 874. [Google Scholar] [CrossRef]
- Wang, S.; Gao, Y. Pancreatic Cancer Cell-Derived MicroRNA-155-5p-Containing Extracellular Vesicles Promote Immune Evasion by Triggering EHF-Dependent Activation of Akt/NF-ΚB Signaling Pathway. Int. Immunopharmacol. 2021, 100, 107990. [Google Scholar] [CrossRef]
- Gorzalczany, Y.; Merimsky, O.; Sagi-Eisenberg, R. Mast Cells Are Directly Activated by Cancer Cell–Derived Extracellular Vesicles by a CD73- and Adenosine-Dependent Mechanism. Transl. Oncol. 2019, 12, 1549–1556. [Google Scholar] [CrossRef] [PubMed]
- Namee, N.M.; O’Driscoll, L. Extracellular Vesicles and Anti-Cancer Drug Resistance. Biochim. Biophys. Acta Rev. Cancer 2018, 1870, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Kawamoto, K.; et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 42339. [Google Scholar] [CrossRef] [PubMed]
- Richards, K.E.; Zeleniak, A.E.; Fishel, M.L.; Wu, J.; Littlepage, L.E.; Hill, R. Cancer-Associated Fibroblast Exosomes Regulate Survival and Proliferation of Pancreatic Cancer Cells. Oncogene 2017, 36, 1770–1778. [Google Scholar] [CrossRef] [Green Version]
- Melo, S.A.; Luecke, L.B.; Kahlert, C.; Fernandez, A.F.; Gammon, S.T.; Kaye, J.; LeBleu, V.S.; Mittendorf, E.A.; Weitz, J.; Rahbari, N.; et al. Glypican-1 Identifies Cancer Exosomes and Detects Early Pancreatic Cancer. Nature 2015, 523, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Frampton, A.E.; Prado, M.M.; López-Jiménez, E.; Fajardo-Puerta, A.B.; Jawad, Z.A.R.; Lawton, P.; Giovannetti, E.; Habib, N.A.; Castellano, L.; Stebbing, J.; et al. Glypican-1 Is Enriched in Circulating-Exosomes in Pancreatic Cancer and Correlates with Tumor Burden. Oncotarget 2018, 9, 19006–19013. [Google Scholar] [CrossRef] [Green Version]
- Lucien, F.; Lac, V.; Billadeau, D.D.; Borgida, A.; Gallinger, S.; Leong, H.S. Glypican-1 and Glycoprotein 2 Bearing Extracellular Vesicles Do Not Discern Pancreatic Cancer from Benign Pancreatic Diseases. Oncotarget 2019, 10, 1045–1055. [Google Scholar] [CrossRef] [Green Version]
- Buscail, E.; Cauvin, T.; Fernandez, B.; Buscail, C.; Marty, M.; Lapuyade, B.; Subtil, C.; Adam, J.-P.; Vendrely, V.; Dabernat, S.; et al. Intraductal Papillary Mucinous Neoplasms of the Pancreas and European Guidelines: Importance of the Surgery Type in the Decision-Making Process. BMC Surg. 2019, 19, 115. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Liu, P.; Wu, Y.; Meng, X.; Wu, M.; Han, J.; Tan, X. Exosomal Zinc Transporter ZIP4 Promotes Cancer Growth and Is a Novel Diagnostic Biomarker for Pancreatic Cancer. Cancer Sci. 2018, 109, 2946–2956. [Google Scholar] [CrossRef]
- Castillo, J.; Bernard, V.; San Lucas, F.A.; Allenson, K.; Capello, M.; Kim, D.U.; Gascoyne, P.; Mulu, F.C.; Stephens, B.M.; Huang, J.; et al. Surfaceome Profiling Enables Isolation of Cancer-Specific Exosomal Cargo in Liquid Biopsies from Pancreatic Cancer Patients. Ann. Oncol. 2018, 29, 223–229. [Google Scholar] [CrossRef]
- Wang, H.; Rana, S.; Giese, N.; Büchler, M.W.; Zöller, M. Tspan8, CD44v6 and Alpha6beta4 Are Biomarkers of Migrating Pancreatic Cancer-Initiating Cells: Pancreatic Cancer-Initiating Cell Markers. Int. J. Cancer 2013, 133, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Yue, S.; Mu, W.; Erb, U.; Zöller, M. The Tetraspanins CD151 and Tspan8 Are Essential Exosome Components for the Crosstalk between Cancer Initiating Cells and Their Surrounding. Oncotarget 2015, 6, 2366–2384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funamizu, N.; Hu, C.; Lacy, C.; Schetter, A.; Zhang, G.; He, P.; Gaedcke, J.; Ghadimi, M.B.; Ried, T.; Yfantis, H.G.; et al. Macrophage Migration Inhibitory Factor Induces Epithelial to Mesenchymal Transition, Enhances Tumor Aggressiveness and Predicts Clinical Outcome in Resected Pancreatic Ductal Adenocarcinoma. Int. J. Cancer 2013, 132, 785–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, K.; Liu, F.; Fan, J.; Sun, D.; Liu, C.; Lyon, C.J.; Bernard, D.W.; Li, Y.; Yokoi, K.; Katz, M.H.; et al. Nanoplasmonic Quantification of Tumor-Derived Extracellular Vesicles in Plasma Microsamples for Diagnosis and Treatment Monitoring. Nat. Biomed. Eng. 2017, 1, 0021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakomori, S. Glycosylation Defining Cancer Malignancy: New Wine in an Old Bottle. Proc. Natl. Acad. Sci. USA 2002, 99, 10231–10233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, A.; Kuno, A.; Nakagawa, T.; Ikehara, Y.; Irimura, T.; Yamamoto, M.; Nakanuma, Y.; Miyoshi, E.; Nakamori, S.; Nakanishi, H.; et al. Lectin Microarray-Based Sero-Biomarker Verification Targeting Aberrant O-Linked Glycosylation on Mucin 1. Anal. Chem. 2015, 87, 7274–7281. [Google Scholar] [CrossRef] [PubMed]
- Yokose, T.; Kabe, Y.; Matsuda, A.; Kitago, M.; Matsuda, S.; Hirai, M.; Nakagawa, T.; Masugi, Y.; Hishiki, T.; Nakamura, Y.; et al. O-Glycan-Altered Extracellular Vesicles: A Specific Serum Marker Elevated in Pancreatic Cancer. Cancers 2020, 12, 2469. [Google Scholar] [CrossRef]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in Cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef]
- Cheng, L.; Sharples, R.A.; Scicluna, B.J.; Hill, A.F. Exosomes Provide a Protective and Enriched Source of MiRNA for Biomarker Profiling Compared to Intracellular and Cell-Free Blood. J. Extracell Vesicles 2014, 3, 23743. [Google Scholar] [CrossRef]
- Flammang, I.; Reese, M.; Yang, Z.; Eble, J.A.; Dhayat, S.A. Tumor-Suppressive MiR-192-5p Has Prognostic Value in Pancreatic Ductal Adenocarcinoma. Cancers 2020, 12, 1693. [Google Scholar] [CrossRef] [PubMed]
- Reese, M.; Flammang, I.; Yang, Z.; Dhayat, S.A. Potential of Exosomal MicroRNA-200b as Liquid Biopsy Marker in Pancreatic Ductal Adenocarcinoma. Cancers 2020, 12, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Omura, N.; Hong, S.-M.; Vincent, A.; Walter, K.; Griffith, M.; Borges, M.; Goggins, M. Pancreatic Cancers Epigenetically Silence SIP1 and Hypomethylate and Overexpress MiR-200a/200b in Association with Elevated Circulating MiR-200a and MiR-200b Levels. Cancer Res. 2010, 70, 5226–5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pu, X.; Ding, G.; Wu, M.; Zhou, S.; Jia, S.; Cao, L. Elevated Expression of Exosomal MicroRNA-21 as a Potential Biomarker for the Early Diagnosis of Pancreatic Cancer Using a Tethered Cationic Lipoplex Nanoparticle Biochip. Oncol. Lett. 2020, 19, 2062–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhavan, B.; Yue, S.; Galli, U.; Rana, S.; Gross, W.; Müller, M.; Giese, N.A.; Kalthoff, H.; Becker, T.; Büchler, M.W.; et al. Combined Evaluation of a Panel of Protein and MiRNA Serum-Exosome Biomarkers for Pancreatic Cancer Diagnosis Increases Sensitivity and Specificity. Int. J. Cancer 2015, 136, 2616–2627. [Google Scholar] [CrossRef]
- Que, R.; Ding, G.; Chen, J.; Cao, L. Analysis of Serum Exosomal MicroRNAs and Clinicopathologic Features of Patients with Pancreatic Adenocarcinoma. World J. Surg. Oncol. 2013, 11, 219. [Google Scholar] [CrossRef] [Green Version]
- Singhi, A.D.; Koay, E.J.; Chari, S.T.; Maitra, A. Early Detection of Pancreatic Cancer: Opportunities and Challenges. Gastroenterology 2019, 156, 2024–2040. [Google Scholar] [CrossRef] [Green Version]
- van Huijgevoort, N.C.M.; Del Chiaro, M.; Wolfgang, C.L.; van Hooft, J.E.; Besselink, M.G. Diagnosis and Management of Pancreatic Cystic Neoplasms: Current Evidence and Guidelines. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 676–689. [Google Scholar] [CrossRef]
- European Study Group on Cystic Tumours of the Pancreas European Evidence-Based Guidelines on Pancreatic Cystic Neoplasms. Gut 2018, 67, 789–804. [CrossRef] [Green Version]
- Tanaka, M.; Ishikawa, S.; Ushiku, T.; Morikawa, T.; Isagawa, T.; Yamagishi, M.; Yamamoto, H.; Katoh, H.; Takeshita, K.; Arita, J.; et al. EVI1 Modulates Oncogenic Role of GPC1 in Pancreatic Carcinogenesis. Oncotarget 2017, 8, 99552–99566. [Google Scholar] [CrossRef]
- Yang, K.S.; Ciprani, D.; O’Shea, A.; Liss, A.S.; Yang, R.; Fletcher-Mercaldo, S.; Mino-Kenudson, M.; Fernández-del Castillo, C.; Weissleder, R. Extracellular Vesicle Analysis Allows for Identification of Invasive IPMN. Gastroenterology 2021, 160, 1345–1358.e11. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Jain, M.; Batra, S.K. Liquid Biopsy for Identification of High-Risk Cystic Lesions of Pancreas. Gastroenterology 2021, 160, 1016–1018. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Suzuki, R.; Takagi, T.; Sugimoto, M.; Ohira, H. Circulating Extracellular Vesicle-encapsulated MicroRNA as Screening Biomarkers for Intraductal Papillary Mucinous Neoplasm. Oncol. Lett. 2020, 20, 315. [Google Scholar] [CrossRef] [PubMed]
- Sonohara, F.; Yamada, S.; Takeda, S.; Hayashi, M.; Suenaga, M.; Sunagawa, Y.; Tashiro, M.; Takami, H.; Kanda, M.; Tanaka, C.; et al. Exploration of Exosomal Micro RNA Biomarkers Related to Epithelial-to-Mesenchymal Transition in Pancreatic Cancer. Anticancer Res. 2020, 40, 1843–1853. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Fujiya, M.; Konishi, H.; Sasajima, J.; Fujibayashi, S.; Hayashi, A.; Utsumi, T.; Sato, H.; Iwama, T.; Ijiri, M.; et al. An Elevated Expression of Serum Exosomal MicroRNA-191, − 21, −451a of Pancreatic Neoplasm Is Considered to Be Efficient Diagnostic Marker. BMC Cancer 2018, 18, 116. [Google Scholar] [CrossRef] [Green Version]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. Biomed. Res. Int. 2018, 2018, 8545347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Altelaar, A.F.M.; Munoz, J.; Heck, A.J.R. Next-Generation Proteomics: Towards an Integrative View of Proteome Dynamics. Nat. Rev. Genet. 2013, 14, 35–48. [Google Scholar] [CrossRef]
- Yang, K.S.; Im, H.; Hong, S.; Pergolini, I.; Del Castillo, A.F.; Wang, R.; Clardy, S.; Huang, C.-H.; Pille, C.; Ferrone, S.; et al. Multiparametric Plasma EV Profiling Facilitates Diagnosis of Pancreatic Malignancy. Sci. Transl. Med. 2017, 9, eaal3226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, L.; Li, D.; Campbell, D.H.; Walsh, B.J.; Packer, N.H.; Dong, Q.; Wang, E.; Wang, Y. Phenotypic Profiling of Pancreatic Ductal Adenocarcinoma Plasma-Derived Small Extracellular Vesicles for Cancer Diagnosis and Cancer Stage Prediction: A Proof-of-Concept Study. Anal. Methods 2022, 14, 2255–2265. [Google Scholar] [CrossRef]
- Cheng, L.; Hill, A.F. Therapeutically Harnessing Extracellular Vesicles. Nat. Rev. Drug Discov. 2022, 21, 379–399. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.; Zhou, Y.; Shi, J. Extrachromosomal Circular DNA: A New Potential Role in Cancer Progression. J. Transl. Med. 2021, 19, 257. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Y.; Li, J.; Zhou, X. Extrachromosomal Circular DNA (EccDNA): An Emerging Star in Cancer. Biomark. Res. 2022, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Yi, Y.; Wang, C.; Li, X.; Zhou, M.; Peng, Q.; Zhou, J.; Yang, Y.; He, Q. Extrachromosomal Circular DNA (EccDNA): From Chaos to Function. Front. Cell Dev. Biol. 2021, 9, 792555. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Jiang, W.; Ye, L.; Li, T.; Yu, X.; Liu, L. Classification of Extrachromosomal Circular DNA with a Focus on the Role of Extrachromosomal DNA (EcDNA) in Tumor Heterogeneity and Progression. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188392. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, S.S.; Dunlay, C.J.; Simeone, D.M.; Nagrath, S. Microfluidic Device (ExoChip) for on-Chip Isolation, Quantification and Characterization of Circulating Exosomes. Lab. Chip 2014, 14, 1891–1900. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Greening, D.W.; Zhu, H.-J.; Takahashi, N.; Simpson, R.J. Extracellular Vesicle Isolation and Characterization: Toward Clinical Application. J. Clin. Investig. 2016, 126, 1152–1162. [Google Scholar] [CrossRef] [Green Version]
- Hoshino, A.; Kim, H.S.; Bojmar, L.; Gyan, K.E.; Cioffi, M.; Hernandez, J.; Zambirinis, C.P.; Rodrigues, G.; Molina, H.; Heissel, S.; et al. Extracellular Vesicle and Particle Biomarkers Define Multiple Human Cancers. Cell 2020, 182, 1044–1061.e18. [Google Scholar] [CrossRef]



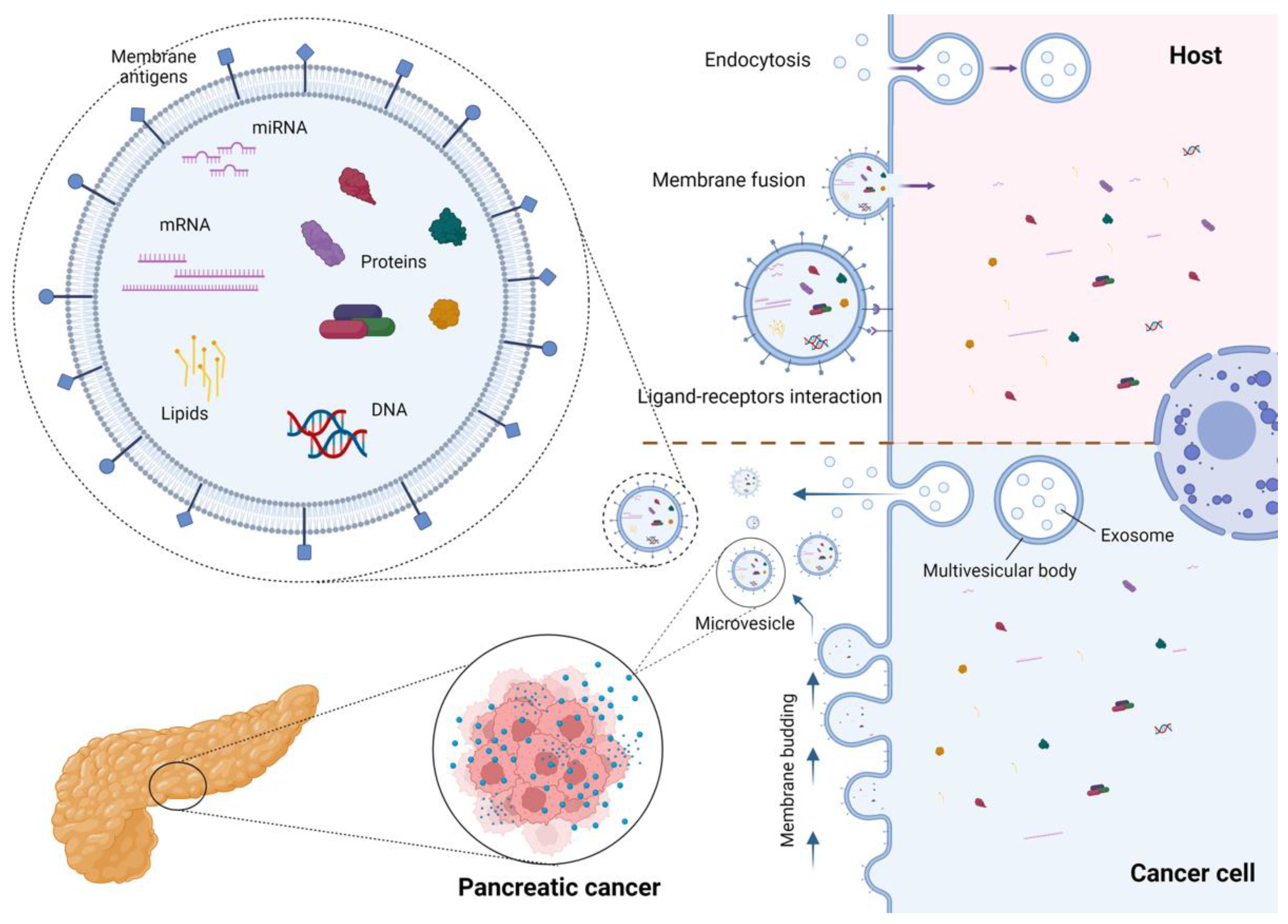{kind=link}
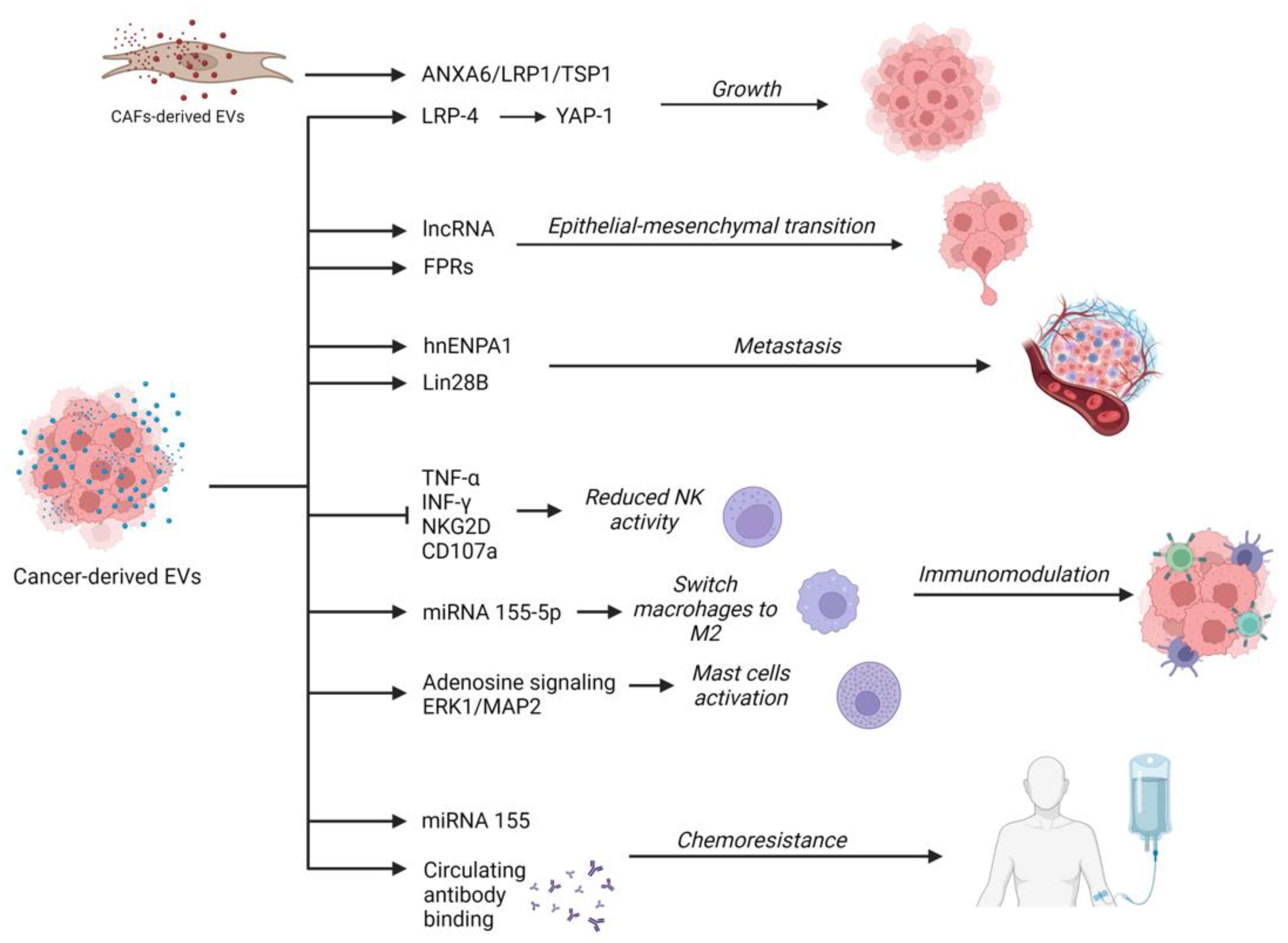{kind=link}
| Principle for EVs Isolation | Method | Type of Sample | EV Size | EV Yield | Purity | Time | Main Advantages | Main Disadvantages |
|---|---|---|---|---|---|---|---|---|
| Buoyant density or size-selection | Differential centrifugation [17] | Plasma Urine Cell medium | 20–100 nm | Medium | Low | 140–600 min | Cost | Low reproducibility Impurities Damage of exosomes Variable efficiency |
| Density gradient centrifugation [18] | Urine Cell medium | 20–100 nm | Medium | High | 250 min–2 days | Purity | Complexity Loss of sample | |
| Size-exclusive chromatography [19] | Plasma Platelet-free plasma Urine Cell medium | 50–500 nm | Medium | Medium | 1 mL/min + column washing | Purity Reproducibility EVs integrity | Cost Complexity | |
| Ultrafiltration [20,21] | Plasma Urine Cell medium | 50–250 nm | Low | High | 130 min | Simplicity Pure preparation | Loss of sample Deformation of EVs | |
| Hydrostatic dialysis [22] | Urine | 50–90 nm | - | 30 min–1 h per 75 mL | Cost | Additional purification from bacteria | ||
| Precipitation or phase separation | Precipitation with protamine [23] | Cell culture | - | - | Low | 55 min + incubation overnight | Cost Simplicity Preservation of EVs integrity | Need purification Long duration |
| Precipitation with polymers [24] | Plasma (PEG) Urine, plasma, or cell medium (ExoQuick) | 50–200 nm | High | Low | 45–65 min | Cost (PEG) Simplicity EVs integrity | Cost with commercial kits Contamination | |
| Precipitation with sodium acetate [25] | Cell culture | - | - | High | 130 min | Cost Simplicity EVs integrity Purity Efficiency | Contamination with non-EVs protein | |
| Precipitation of protein with organic solvent [26] | Plasma | 20–300 nm | - | - | 105 min | Cost Simplicity | Aggregation | |
| Two-phase isolation [27] | Plasma Mixture of exosomes and proteins | Exosomes | - | High | 75–195 min | Cost Simplicity Purity Efficiency | Contamination with polymer | |
| Affinity Interaction | Antibodies to EV receptors [28] | Cell medium Plasma | 40–150 nm | Low | High | 240 min | Purity High selectivity | Cost Damage of EVs |
| Phosphatidylserine-binding proteins [29] | Plasma Urine Cell medium | 106 nm (with Tim protein) | 12 h incubation | Reversible binding Simplicity | Cost | |||
| Heparin-modified sorbents [30] | Plasma Cell medium | - | 24 h incubation | Cost EVs integrity | Need initial purification and ultracentrifugation | |||
| Binding of heat shock proteins [31] | Plasma Urine Cell medium | 30–100 nm | <1 h | EVs integrity | Cost | |||
| Lectins [32] | Urine | - | 12 h incubation | Cost Simplicity Purity | Need initial purification and ultracentrifugation | |||
| Size selection, affinity interaction | Microfluidic technologies [33] | Blood Plasma Cell culture | Medium | Low | <2 h | Rapidness Efficiency | Cost Complexity |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicoletti, A.; Negri, M.; Paratore, M.; Vitale, F.; Ainora, M.E.; Nista, E.C.; Gasbarrini, A.; Zocco, M.A.; Zileri Dal Verme, L. Diagnostic and Prognostic Role of Extracellular Vesicles in Pancreatic Cancer: Current Evidence and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 885. https://doi.org/10.3390/ijms24010885
Nicoletti A, Negri M, Paratore M, Vitale F, Ainora ME, Nista EC, Gasbarrini A, Zocco MA, Zileri Dal Verme L. Diagnostic and Prognostic Role of Extracellular Vesicles in Pancreatic Cancer: Current Evidence and Future Perspectives. International Journal of Molecular Sciences. 2023; 24(1):885. https://doi.org/10.3390/ijms24010885
Chicago/Turabian StyleNicoletti, Alberto, Marcantonio Negri, Mattia Paratore, Federica Vitale, Maria Elena Ainora, Enrico Celestino Nista, Antonio Gasbarrini, Maria Assunta Zocco, and Lorenzo Zileri Dal Verme. 2023. "Diagnostic and Prognostic Role of Extracellular Vesicles in Pancreatic Cancer: Current Evidence and Future Perspectives" International Journal of Molecular Sciences 24, no. 1: 885. https://doi.org/10.3390/ijms24010885