
IL-20 Activates ERK1/2 and Suppresses Splicing of X-Box Protein-1 in Intestinal Epithelial Cells but Does Not Improve Pathology in Acute or Chronic Models of Colitis
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
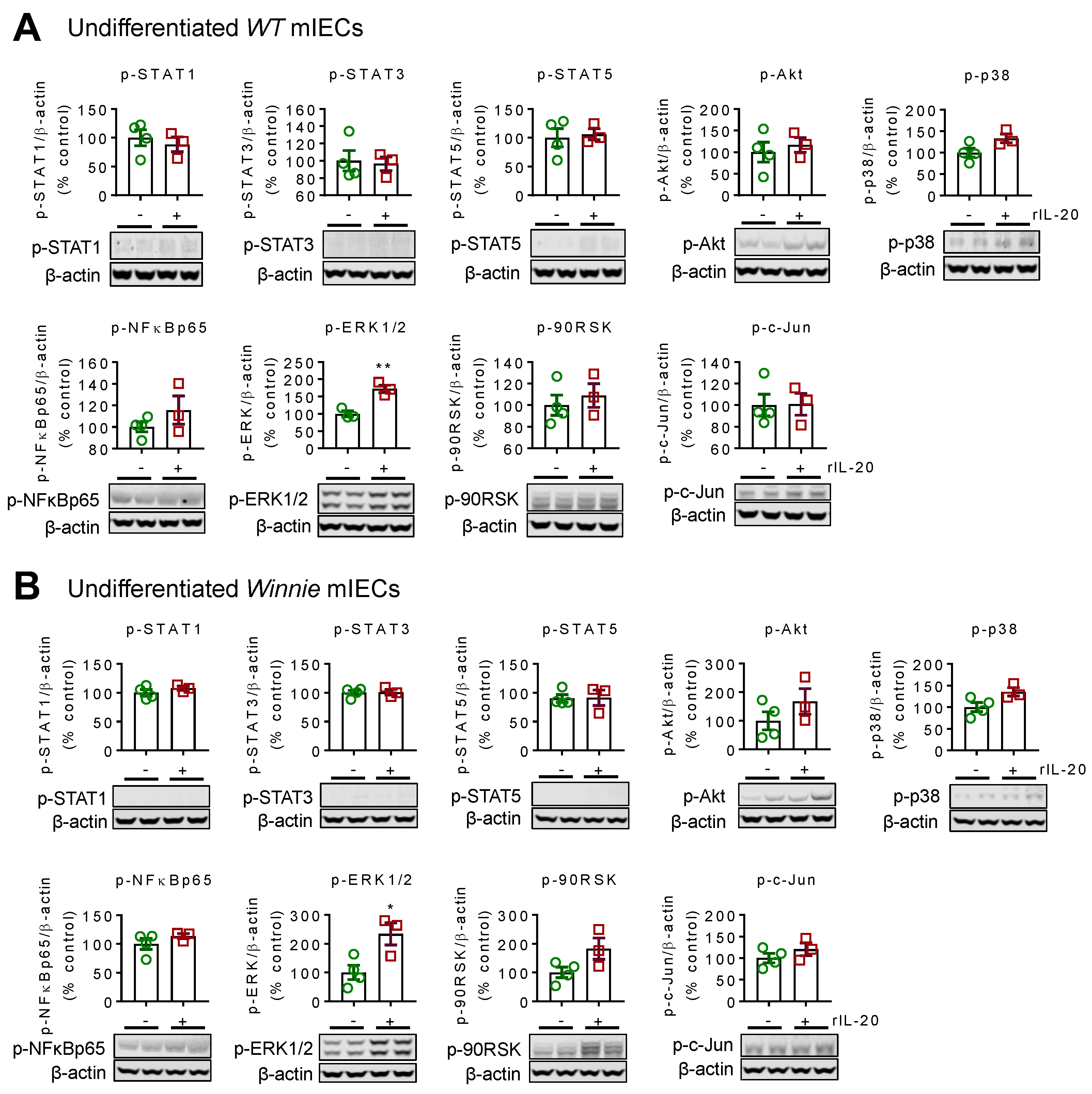
2.1. Recombinant IL-20 Activates ERK1/2 in Intestinal Epithelial Cells but Does Not Modulate Signaling in Goblet Cells
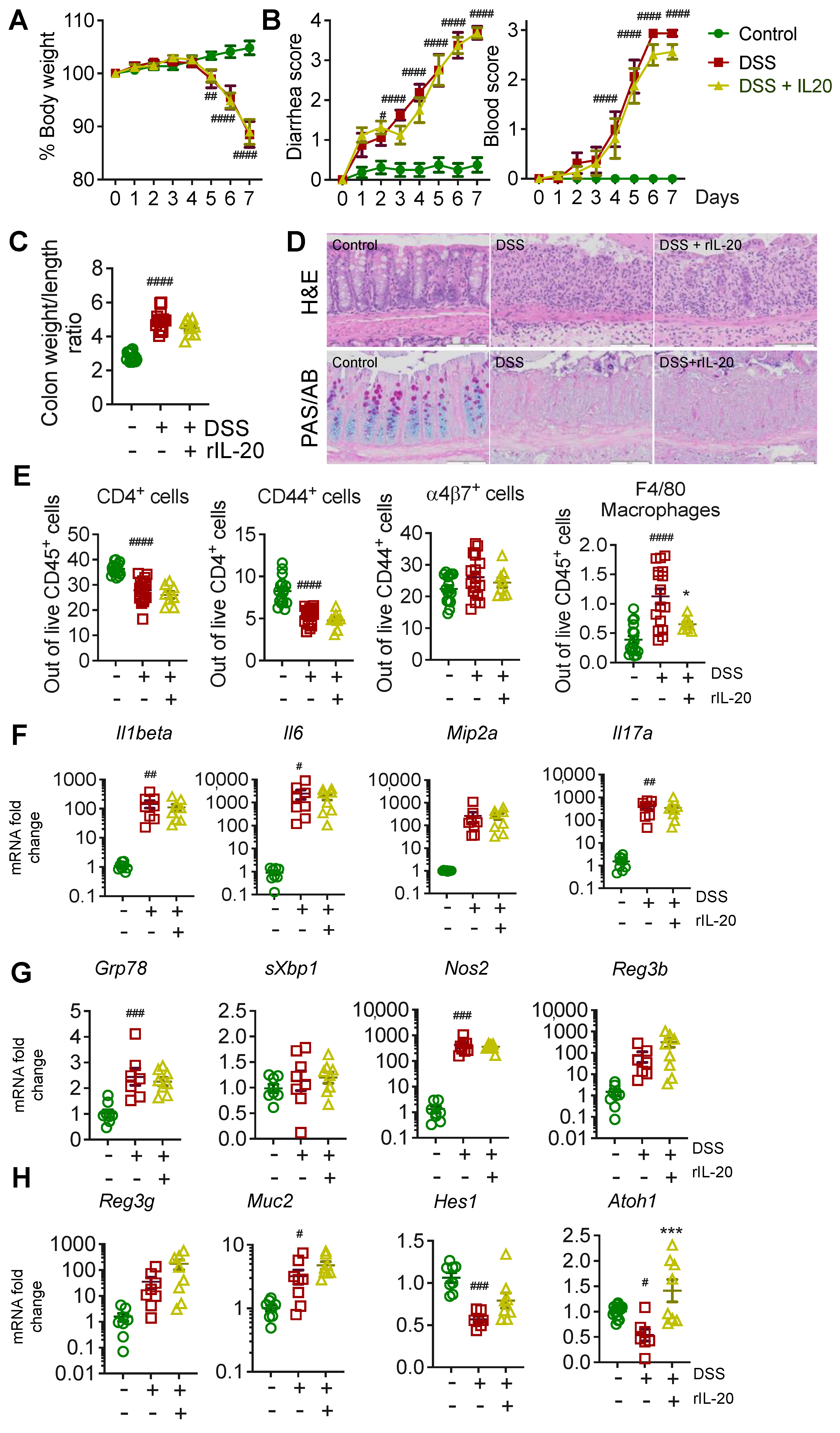
2.2. Recombinant IL-20 Treatment Did Not Modulate Pathology in Acute Chemical-Induced Colitis
2.3. IL-20 Is Capable of Reducing Endoplasmic Reticulum Stress; However, Has No Major Effects on Pathology in the Acute DSS-Induced Colitis Model
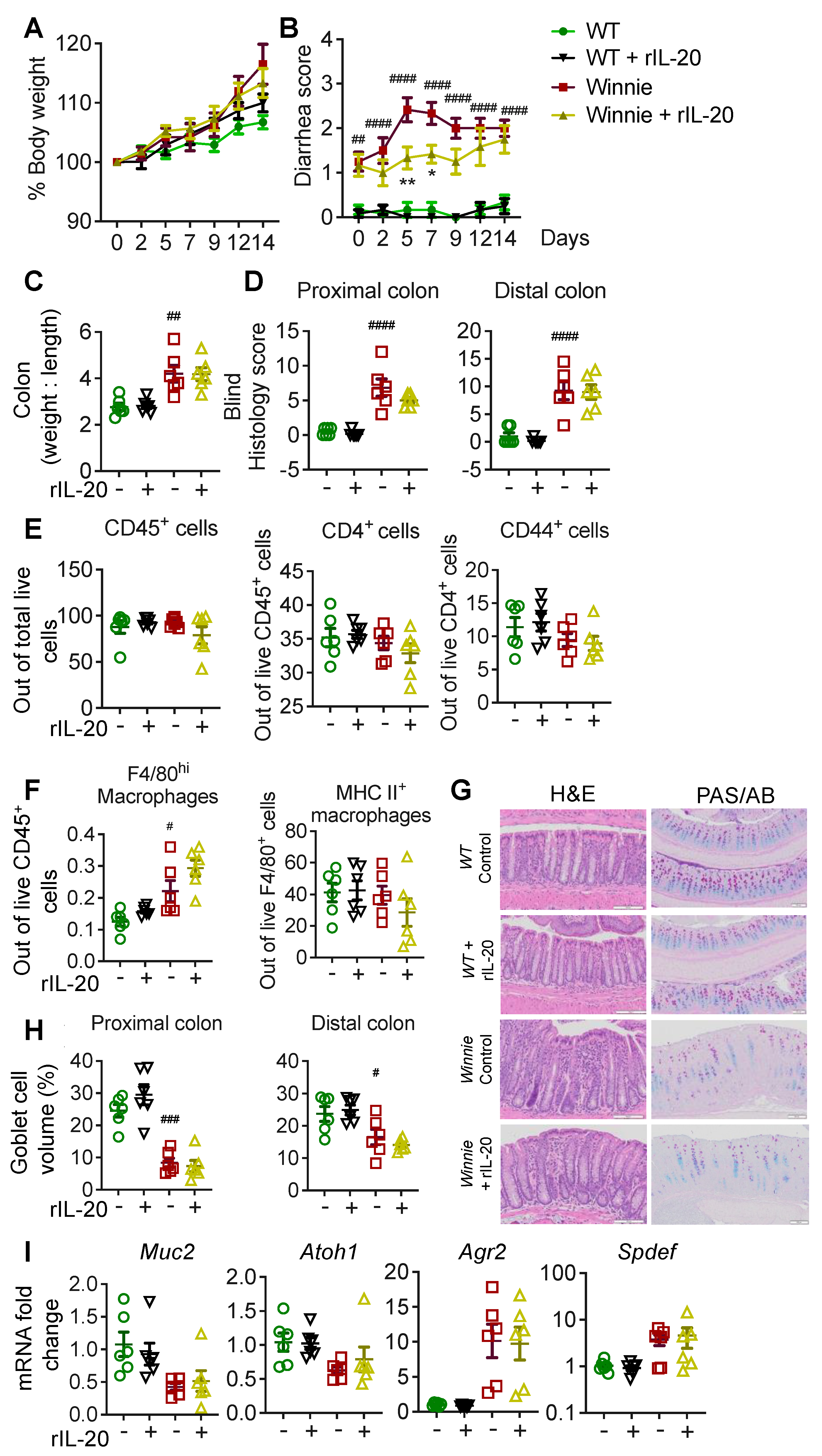
2.4. IL-20 Treatment Transiently Improves Diarrhea in Winnie Mice without Long-Term Improvements in Pathology
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals
4.3. Murine Primary Intestinal Epithelial Cell Culture (mIECs)
4.4. Western Blotting
4.5. Quantitative Reverse Transcriptase Polymerase Chain Reaction (qRT PCR)
4.6. Effects of rIL-20 in Acute DSS-Induced Colitis
4.7. Effects of rIL-20 in Winnie Spontaneous Colitis
4.8. Flow Cytometry Analyses of the Immune Cells
4.9. Histological Analysis of Intestinal Tissues
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Yamamoto-Furusho, J.K. Inflammatory bowel disease therapy: Blockade of cytokines and cytokine signaling pathways. Curr. Opin. Gastroenterol. 2018, 34, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tan, Z.; Zhang, R.; Kotenko, S.V.; Liang, P. Interleukin 24 (MDA-7/MOB-5) signals through two heterodimeric receptors, IL-22R1/IL-20R2 and IL-20R1/IL-20R2. J. Biol. Chem. 2002, 277, 7341–7347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parrish-Novak, J.; Xu, W.; Brender, T.; Yao, L.; Jones, C.; West, J.; Brandt, C.; Jelinek, L.; Madden, K.; McKernan, P.A.; et al. Interleukins 19, 20, and 24 signal through two distinct receptor complexes. Differences in receptor-ligand interactions mediate unique biological functions. J. Biol. Chem. 2002, 277, 47517–47523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Llorente, L.; Yamamoto-Furusho, J.K. IL-10-- and IL-20--expressing epithelial and inflammatory cells are increased in patients with ulcerative colitis. J. Clin. Immunol. 2013, 33, 640–648. [Google Scholar] [CrossRef]
- Niess, J.H.; Hruz, P.; Kaymak, T. The Interleukin-20 Cytokines in Intestinal Diseases. Front. Immunol. 2018, 9, 1373. [Google Scholar] [CrossRef]
- Dumoutier, L.; Leemans, C.; Lejeune, D.; Kotenko, S.V.; Renauld, J.C. Cutting edge: STAT activation by IL-19, IL-20 and mda-7 through IL-20 receptor complexes of two types. J. Immunol. 2001, 167, 3545–3549. [Google Scholar] [CrossRef] [Green Version]
- Wegenka, U.M. IL-20: Biological functions mediated through two types of receptor complexes. Cytokine Growth Factor Rev. 2010, 21, 353–363. [Google Scholar] [CrossRef]
- Bosanquet, D.C.; Harding, K.G.; Ruge, F.; Sanders, A.J.; Jiang, W.G. Expression of IL-24 and IL-24 receptors in human wound tissues and the biological implications of IL-24 on keratinocytes. Wound Repair Regen. 2012, 20, 896–903. [Google Scholar] [CrossRef]
- Zhang, W.; Magadi, S.; Li, Z.; Smith, C.W.; Burns, A.R. IL-20 promotes epithelial healing of the injured mouse cornea. Exp. Eye Res. 2016, 154, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Kolumam, G.; Wu, X.; Lee, W.P.; Hackney, J.A.; Zavala-Solorio, J.; Gandham, V.; Danilenko, D.M.; Arora, P.; Wang, X.; Ouyang, W. IL-22R Ligands IL-20, IL-22, and IL-24 Promote Wound Healing in Diabetic db/db Mice. PLoS ONE 2017, 12, e0170639. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Wang, R.; Jeet, V.; McGuckin, M.A.; Hasnain, S.Z. Interleukin (IL)-22 from IL-20 Subfamily of Cytokines Induces Colonic Epithelial Cell Proliferation Predominantly through ERK1/2 Pathway. Int. J. Mol. Sci. 2019, 20, 3468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto-Furusho, J.K.; De-Leon-Rendon, J.L.; de la Torre, M.G.; Alvarez-Leon, E.; Vargas-Alarcon, G. Genetic polymorphisms of interleukin 20 (IL-20) in patients with ulcerative colitis. Immunol. Lett. 2013, 149, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Gulhane, M.; Murray, L.; Lourie, R.; Tong, H.; Sheng, Y.H.; Wang, R.; Kang, A.; Schreiber, V.; Wong, K.Y.; Magor, G.; et al. High Fat Diets Induce Colonic Epithelial Cell Stress and Inflammation that is Reversed by IL-22. Sci. Rep. 2016, 6, 28990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grey, M.J.; De Luca, H.; Ward, D.V.; Kreulen, I.A.; Bugda Gwilt, K.; Foley, S.E.; Thiagarajah, J.R.; McCormick, B.A.; Turner, J.R.; Lencer, W.I. The epithelial-specific ER stress sensor ERN2/IRE1beta enables host-microbiota crosstalk to affect colon goblet cell development. J. Clin. Invest. 2022, 132, e153519. [Google Scholar] [CrossRef]
- Andoh, A.; Shioya, M.; Nishida, A.; Bamba, S.; Tsujikawa, T.; Kim-Mitsuyama, S.; Fujiyama, Y. Expression of IL-24, an activator of the JAK1/STAT3/SOCS3 cascade, is enhanced in inflammatory bowel disease. J. Immunol. 2009, 183, 687–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.H.; Chung, E.; Li, Z.; Wan, Y.W.; Mahe, M.M.; Chen, M.S.; Noah, T.K.; Bell, K.N.; Yalamanchili, H.K.; Klisch, T.J.; et al. Transcriptional Regulation by ATOH1 and its Target SPDEF in the Intestine. Cell Mol. Gastroenterol. Hepatol. 2017, 3, 51–71. [Google Scholar] [CrossRef] [Green Version]
- Vooijs, M.; Liu, Z.; Kopan, R. Notch: Architect, landscaper, and guardian of the intestine. Gastroenterology 2011, 141, 448–459. [Google Scholar] [CrossRef] [Green Version]
- Bech, R.; Jalilian, B.; Agger, R.; Iversen, L.; Erlandsen, M.; Otkjaer, K.; Johansen, C.; Paludan, S.R.; Rosenberg, C.A.; Kragballe, K.; et al. Interleukin 20 regulates dendritic cell migration and expression of co-stimulatory molecules. Mol. Cell. Ther. 2016, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Myles, I.A.; Fontecilla, N.M.; Valdez, P.A.; Vithayathil, P.J.; Naik, S.; Belkaid, Y.; Ouyang, W.; Datta, S.K. Signaling via the IL-20 receptor inhibits cutaneous production of IL-1beta and IL-17A to promote infection with methicillin-resistant Staphylococcus aureus. Nat. Immunol. 2013, 14, 804–811. [Google Scholar] [CrossRef]
- Hasnain, S.Z.; Tauro, S.; Das, I.; Tong, H.; Chen, A.C.; Jeffery, P.L.; McDonald, V.; Florin, T.H.; McGuckin, M.A. IL-10 promotes production of intestinal mucus by suppressing protein misfolding and endoplasmic reticulum stress in goblet cells. Gastroenterology 2013, 144, 357–368.e9. [Google Scholar] [CrossRef]
- Wang, R.; Moniruzzaman, M.; Wong, K.Y.; Wiid, P.; Harding, A.; Giri, R.; Tong, W.H.; Creagh, J.; Begun, J.; McGuckin, M.A.; et al. Gut microbiota shape the inflammatory response in mice with an epithelial defect. Gut Microbes 2021, 13, 1–18. [Google Scholar] [CrossRef]
- Wang, R.; Hasnain, S.Z.; Tong, H.; Das, I.; Che-Hao Chen, A.; Oancea, I.; Proctor, M.; Florin, T.H.; Eri, R.D.; McGuckin, M.A. Neutralizing IL-23 is superior to blocking IL-17 in suppressing intestinal inflammation in a spontaneous murine colitis model. Inflamm. Bowel. Dis. 2015, 21, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Das, I.; Png, C.W.; Oancea, I.; Hasnain, S.Z.; Lourie, R.; Proctor, M.; Eri, R.D.; Sheng, Y.; Crane, D.I.; Florin, T.H.; et al. Glucocorticoids alleviate intestinal ER stress by enhancing protein folding and degradation of misfolded proteins. J. Exp. Med. 2013, 210, 1201–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasnain, S.Z.; Borg, D.J.; Harcourt, B.E.; Tong, H.; Sheng, Y.H.; Ng, C.P.; Das, I.; Wang, R.; Chen, A.C.; Loudovaris, T.; et al. Glycemic control in diabetes is restored by therapeutic manipulation of cytokines that regulate beta cell stress. Nat. Med. 2014, 20, 1417–1426. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, H.; Stappenbeck, T.S. In vitro expansion and genetic modification of gastrointestinal stem cells in spheroid culture. Nat. Protoc. 2013, 8, 2471–2482. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moniruzzaman, M.; Wong, K.Y.; Wang, R.; Symon, H.; Mueller, A.; Rahman, M.A.; Hasnain, S.Z. IL-20 Activates ERK1/2 and Suppresses Splicing of X-Box Protein-1 in Intestinal Epithelial Cells but Does Not Improve Pathology in Acute or Chronic Models of Colitis. Int. J. Mol. Sci. 2023, 24, 174. https://doi.org/10.3390/ijms24010174
Moniruzzaman M, Wong KY, Wang R, Symon H, Mueller A, Rahman MA, Hasnain SZ. IL-20 Activates ERK1/2 and Suppresses Splicing of X-Box Protein-1 in Intestinal Epithelial Cells but Does Not Improve Pathology in Acute or Chronic Models of Colitis. International Journal of Molecular Sciences. 2023; 24(1):174. https://doi.org/10.3390/ijms24010174
Chicago/Turabian StyleMoniruzzaman, Md., Kuan Yau Wong, Ran Wang, Hamish Symon, Alexandra Mueller, M. Arifur Rahman, and Sumaira Z. Hasnain. 2023. "IL-20 Activates ERK1/2 and Suppresses Splicing of X-Box Protein-1 in Intestinal Epithelial Cells but Does Not Improve Pathology in Acute or Chronic Models of Colitis" International Journal of Molecular Sciences 24, no. 1: 174. https://doi.org/10.3390/ijms24010174