
Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations

 , ,
, ,
Abstract
:1. Introduction
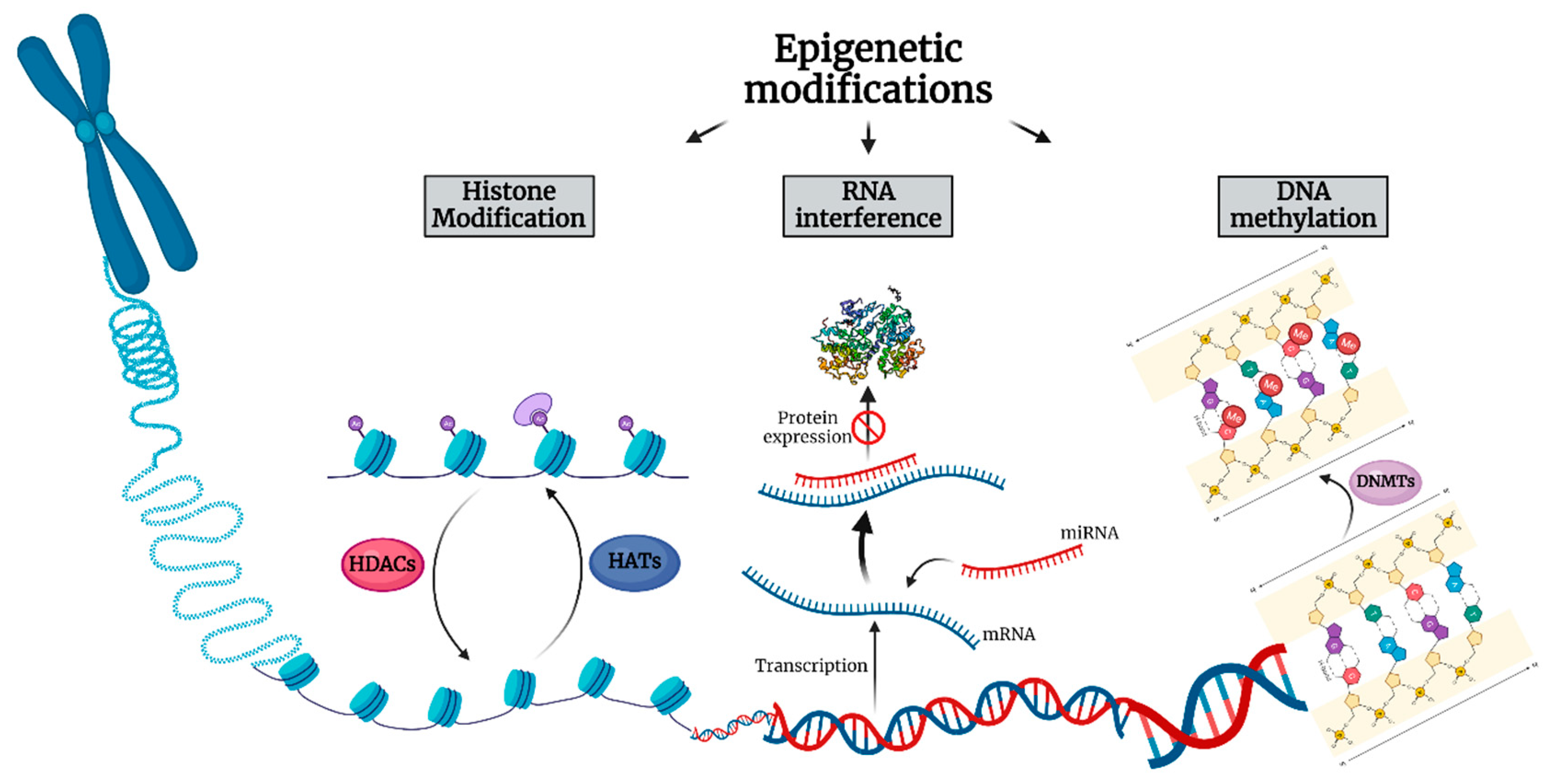
2. Alterations of Non-Coding RNAs in Thymic Epithelial Neoplasms
3. Alterations of DNA Methylation in TENs
4. Alterations of Histone Modifications in Thymic Epithelial Neoplasms
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valavanis, C.; Stanc, G.M.; Baltayiannis, N. Classification, histopathology and molecular pathology of thymic epithelial tumors: A review. J. BUON 2021, 26, 1198–1207. [Google Scholar] [PubMed]
- Marx, A.; Chan, J.K.C.; Chalabreysse, L.; Dacic, S.; Detterbeck, F.; French, C.A.; Hornick, J.L.; Inagaki, H.; Jain, D.; Lazar, A.J.; et al. The 2021 WHO Classification of Tumors of the Thymus and Mediastinum: What Is New in Thymic Epithelial, Germ Cell, and Mesenchymal Tumors? J. Thorac. Oncol. 2022, 17, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Marx, A.; Chan, J.K.; Coindre, J.-M.; Detterbeck, F.; Girard, N.; Harris, N.L.; Jaffe, E.S.; Kurrer, M.O.; Marom, E.M.; Moreira, A.L.; et al. The 2015 World Health Organization Classification of Tumors of the Thymus: Continuity and Changes. J. Thorac. Oncol. 2015, 10, 1383–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, A.; Ströbel, P.; Badve, S.; Chalabreysse, L.; Chan, J.K.; Chen, G.; de Leval, L.; Detterbeck, F.; Girard, N.; Huang, J.; et al. ITMIG consensus statement on the use of the WHO histological classification of thymoma and thymic carcinoma: Refined definitions, histological criteria, and reporting. J. Thorac. Oncol. 2014, 9, 596–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society. Key Statistics About Thymus Cancers; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- American Cancer Society. Survival Rates for Thymus Cancer; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- American Cancer Society. Signs and Symptoms of Thymus Cancers; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- American Cancer Society. Tests for Thymus Cancer; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- Lamas, A.; Lopez, E.; Carrio, R.; Lopez, D.M. Adipocyte and leptin accumulation in tumor-induced thymic involution. Int. J. Mol. Med. 2016, 37, 133–138. [Google Scholar] [CrossRef] [Green Version]
- American Cancer Society. Treatment of Thymus Cancers by Extent and Type of Tumor; American Cancer Society: Atlanta, GA, USA, 2021. [Google Scholar]
- Centers for Disease Control and Prevention. What is Epigenetics? Centers for Disease Control and Prevention: Atlanta, GA, USA, 2020. [Google Scholar]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Jin, B.; Li, Y.; Robertson, K.D. DNA methylation: Superior or subordinate in the epigenetic hierarchy? Genes Cancer 2011, 2, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Costello, J. DNA methylation: An epigenetic mark of cellular memory. Exp. Mol. Med. 2017, 49, e322. [Google Scholar] [CrossRef] [Green Version]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- McKenna, N.J.; O’Malley, B.W. Combinatorial control of gene expression by nuclear receptors and coregulators. Cell 2002, 108, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Grunstein, M. Histone acetylation in chromatin structure and transcription. Nature 1997, 389, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Archer, S.Y.; Hodin, R.A. Histone acetylation and cancer. Curr. Opin. Genet. Dev. 1999, 9, 171–174. [Google Scholar] [CrossRef]
- Ropero, S.; Esteller, M. The role of histone deacetylases (HDACs) in human cancer. Mol. Oncol. 2007, 1, 19–25. [Google Scholar] [CrossRef]
- Goutas, D.; Theocharis, S.; Tsourouflis, G. Unraveling the Epigenetic Role and Clinical Impact of Histone Deacetylases in Neoplasia. Diagnostics 2021, 11, 1346. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Lee, E.S. Non-coding RNA: What is functional and what is junk? Front. Genet. 2015, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, M.; Zhuo, Y.; Shan, B. MicroRNAs, Long Noncoding RNAs, and Their Functions in Human Disease. Methods Mol. Biol. 2017, 1617, 1–25. [Google Scholar] [PubMed]
- Grillone, K.; Riillo, C.; Scionti, F.; Rocca, R.; Tradigo, G.; Guzzi, P.H.; Alcaro, S.; Di Martino, M.T.; Tagliaferri, P.; Tassone, P. Non-coding RNAs in cancer: Platforms and strategies for investigating the genomic “dark matter”. J. Exp. Clin. Cancer Res. 2020, 39, 117. [Google Scholar] [CrossRef] [PubMed]
- Ganci, F.; Vico, C.; Korita, E.; Sacconi, A.; Gallo, E.; Mori, F.; Cambria, A.; Russo, E.; Anile, M.; Vitolo, D.; et al. MicroRNA expression profiling of thymic epithelial tumors. Lung Cancer 2014, 85, 197–204. [Google Scholar] [CrossRef]
- Wei, J.; Liu, Z.; Wu, K.; Yang, N.; He, Y.; Chen, G.G.; Zhang, J.; Lin, J. Identification of prognostic and subtype-specific potential miRNAs in thymoma. Epigenomics 2017, 9, 647–657. [Google Scholar] [CrossRef]
- Bellissimo, T.; Russo, E.; Ganci, F.; Vico, C.; Sacconi, A.; Longo, F.; Vitolo, D.; Anile, M.; Disio, D.; Marino, M.; et al. Circulating miR-21-5p and miR-148a-3p as emerging non-invasive biomarkers in thymic epithelial tumors. Cancer Biol. Ther. 2016, 17, 79–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radovich, M.; Solzak, J.P.; Hancock, B.A.; Conces, M.L.; Atale, R.; Porter, R.F.; Zhu, J.; Glasscock, J.; Kesler, K.A.; Badve, S.S.; et al. A large microRNA cluster on chromosome 19 is a transcriptional hallmark of WHO type A and AB thymomas. Br. J. Cancer 2016, 114, 477–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enkner, F.; Pichlhöfer, B.; Zaharie, A.T.; Krunic, M.; Holper, T.M.; Janik, S.; Moser, B.; Schlangen, K.; Neudert, B.; Walter, K.; et al. Molecular Profiling of Thymoma and Thymic Carcinoma: Genetic Differences and Potential Novel Therapeutic Targets. Pathol. Oncol. Res. 2017, 23, 551–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, G.; Ren, R.; Fang, X. Identification and Characterization of Non-Coding RNAs in Thymoma. Med. Sci. Monit. 2021, 27, e929727. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Chen, Y.; Tian, Z.; Lu, C.; Chen, L.; Ma, X. lncRNAs classifier to accurately predict the recurrence of thymic epithelial tumors. Thorac. Cancer 2020, 11, 1773–1783. [Google Scholar] [CrossRef] [PubMed]
- Tito, C.; Ganci, F.; Sacconi, A.; Masciarelli, S.; Fontemaggi, G.; Pulito, C.; Gallo, E.; Laquintana, V.; Iaiza, A.; De Angelis, L.; et al. LINC00174 is a novel prognostic factor in thymic epithelial tumors involved in cell migration and lipid metabolism. Cell Death Dis. 2020, 11, 959. [Google Scholar] [CrossRef]
- Wang, J.; Huang, H.; Zhang, X.; Ma, H. LOXL1AS1 promotes thymoma and thymic carcinoma progression by regulating miR5255pHSPA9. Oncol. Rep. 2021, 45, 117. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Xu, S.; Yang, Y.; Wei, D.; Wang, W.; Huang, X. Aberrant decrease of microRNA19b regulates TSLP expression and contributes to Th17 cells development in myasthenia gravis related thymomas. J. Neuroimmunol. 2015, 288, 34–39. [Google Scholar] [CrossRef]
- Xin, Y.; Cai, H.; Lu, T.; Zhang, Y.; Yang, Y.; Cui, Y. miR-20b Inhibits T Cell Proliferation and Activation via NFAT Signaling Pathway in Thymoma-Associated Myasthenia Gravis. Biomed. Res. Int. 2016, 2016, 9595718. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Zhang, Y.; Song, H. A disparate role of RP11-424C20.2/UHRF1 axis through control of tumor immune escape in liver hepatocellular carcinoma and thymoma. Aging 2019, 11, 6422–6439. [Google Scholar] [CrossRef]
- Zhuang, J.; Guan, M.; Liu, M.; Liu, Y.; Yang, S.; Hu, Z.; Lai, F.; He, F. Immune-Related Molecular Profiling of Thymoma With Myasthenia Gravis. Front. Genet. 2021, 12, 756493. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Bai, L.; Ji, L.; Wu, L.; Li, G. Bioinformatics analysis of the key potential ceRNA biomarkers in human thymic epithelial tumors. Medicine 2021, 100, e26271. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yuan, Y.; Xiao, H.; Dai, J.; Ye, Y.; Zhang, Q.; Zhang, Z.; Jiang, Y.; Luo, J.; Hu, J.; et al. Discovery and validation of DNA methylation markers for overall survival prognosis in patients with thymic epithelial tumors. Clin. Epigenet. 2019, 11, 38. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Meng, Y.; Niu, Y.; Li, S.; Liu, H.; He, J.; Zhang, Y.; Liang, N.; Liu, L.; Mao, X.; et al. Genomewide DNA methylation profile of thymomas and potential epigenetic regulation of thymoma subtypes. Oncol. Rep. 2019, 41, 2762–2774. [Google Scholar] [PubMed]
- Coppedè, F.; Ricciardi, R.; Denaro, M.; De Rosa, A.; Provenzano, C.; Bartoccioni, E.; Baggiani, A.; Lucchi, M.; Mussi, A.; Migliore, L. Association of the DNMT3B -579G>T polymorphism with risk of thymomas in patients with myasthenia gravis. PLoS ONE 2013, 8, e80846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coppedè, F.; Ricciardi, R.; Lopomo, A.; Stoccoro, A.; De Rosa, A.; Guida, M.; Petrucci, L.; Maestri, M.; Lucchi, M.; Migliore, L. Investigation of MLH1, MGMT, CDKN2A, and RASSF1A Gene Methylation in Thymomas From Patients With Myasthenia Gravis. Front. Mol. Neurosci. 2020, 13, 567676. [Google Scholar] [CrossRef]
- Mokhtar, M.; Kondo, K.; Namura, T.; Ali, A.H.; Fujita, Y.; Takai, C.; Takizawa, H.; Nakagawa, Y.; Toba, H.; Kajiura, K.; et al. Methylation and expression profiles of MGMT gene in thymic epithelial tumors. Lung Cancer 2014, 83, 279–287. [Google Scholar] [CrossRef]
- Suzuki, M.; Chen, H.; Shigematsu, H.; Ando, S.; Iida, T.; Nakajima, T.; Fujisawa, T.; Kimura, H. Aberrant methylation: Common in thymic carcinomas, rare in thymomas. Oncol. Rep. 2005, 14, 1621–1624. [Google Scholar] [CrossRef]
- Chen, C.; Yin, B.; Wei, Q.; Li, D.; Hu, J.; Yu, F.; Lu, Q. Aberrant DNA methylation in thymic epithelial tumors. Cancer Investig. 2009, 27, 582–591. [Google Scholar] [CrossRef]
- Gu, Z.; Mitsui, H.; Inomata, K.; Honda, M.; Endo, C.; Sakurada, A.; Sato, M.; Okada, Y.; Kondo, T.; Horii, A. The methylation status of FBXW7 beta-form correlates with histological subtype in human thymoma. Biochem. Biophys. Res. Commun. 2008, 377, 685–688. [Google Scholar] [CrossRef]
- Hirabayashi, H.; Fujii, Y.; Sakaguchi, M.; Tanaka, H.; Yoon, H.E.; Komoto, Y.; Inoue, M.; Miyoshi, S.; Matsuda, H. p16INK4, pRB, p53 and cyclin D1 expression and hypermethylation of CDKN2 gene in thymoma and thymic carcinoma. Int. J. Cancer 1997, 73, 639–644. [Google Scholar] [CrossRef]
- Hirose, Y.; Kondo, K.; Takizawa, H.; Nagao, T.; Nakagawa, Y.; Fujino, H.; Toba, H.; Kenzaki, K.; Sakiyama, S.; Tangoku, A. Aberrant methylation of tumour-related genes in thymic epithelial tumours. Lung Cancer 2009, 64, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Iaiza, A.; Tito, C.; Ianniello, Z.; Ganci, F.; Laquintana, V.; Gallo, E.; Sacconi, A.; Masciarelli, S.; De Angelis, L.; Aversa, S.; et al. METTL3-dependent MALAT1 delocalization drives c-Myc induction in thymic epithelial tumors. Clin. Epigenet. 2021, 13, 173. [Google Scholar] [CrossRef]
- Kishibuchi, R.; Kondo, K.; Soejima, S.; Tsuboi, M.; Kajiura, K.; Kawakami, Y.; Kawakita, N.; Sawada, T.; Toba, H.; Yoshida, M.; et al. DNA methylation of GHSR, GNG4, HOXD9 and SALL3 is a common epigenetic alteration in thymic carcinoma. Int. J. Oncol. 2020, 56, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Tegshee, B.; Kondo, K.; Soejima, S.; Muguruma, K.; Tsuboi, M.; Kajiura, K.; Kawakami, Y.; Kawakita, N.; Toba, H.; Yoshida, M.; et al. GHSR methylation-dependent expression of a variant ligand and receptor of the ghrelin system induces thymoma tumorigenesis. Oncol. Lett. 2021, 22, 793. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Stoccoro, A.; Nicolì, V.; Gallo, R.; De Rosa, A.; Guida, M.; Maestri, M.; Lucchi, M.; Ricciardi, R.; Migliore, L. Investigation of GHSR methylation levels in thymomas from patients with Myasthenia Gravis. Gene 2020, 752, 144774. [Google Scholar] [CrossRef]
- Kont, V.; Murumägi, A.; Tykocinski, L.-O.; Kinkel, S.A.; Webster, K.E.; Kisand, K.; Tserel, L.; Pihlap, M.; Ströbel, P.; Scott, H.; et al. DNA methylation signatures of the AIRE promoter in thymic epithelial cells, thymomas and normal tissues. Mol. Immunol. 2011, 49, 518–526. [Google Scholar] [CrossRef]
- Ye, L.; Li, X.; Kong, X.; Wang, W.; Bi, Y.; Hu, L.; Cui, B.; Li, X.; Ning, G. Hypomethylation in the promoter region of POMC gene correlates with ectopic overexpression in thymic carcinoids. J. Endocrinol. 2005, 185, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Lopomo, A.; Ricciardi, R.; Maestri, M.; De Rosa, A.; Melfi, F.; Lucchi, M.; Mussi, A.; Coppedè, F.; Migliore, L. Gene-Specific Methylation Analysis in Thymomas of Patients with Myasthenia Gravis. Int. J. Mol. Sci. 2016, 17, 2121. [Google Scholar] [CrossRef] [Green Version]
- Masunaga, A.; Omatsu, M.; Kunimura, T.; Uematsu, S.; Kamio, Y.; Kitami, A.; Miyagi, Y.; Hiroshima, K.; Suzuki, T. Expression of PTEN and its pseudogene PTENP1, and promoter methylation of PTEN in non-tumourous thymus and thymic tumours. J. Clin. Pathol. 2017, 70, 690–696. [Google Scholar] [CrossRef]
- Sabbir, M.G.; Prieditis, H.; Ravinsky, E.; Mowat, M.R. The role of Dlc1 isoform 2 in K-Ras2(G12D) induced thymic cancer. PLoS ONE 2012, 7, e40302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, M.; Fujiwara, Y.; Asao, T.; Honda, T.; Shimada, Y.; Kanai, Y.; Tsuta, K.; Kono, K.; Watanabe, S.; Ohe, Y.; et al. The genomic and epigenomic landscape in thymic carcinoma. Carcinogenesis 2017, 38, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Soejima, S.; Kondo, K.; Tsuboi, M.; Muguruma, K.; Tegshee, B.; Kawakami, Y.; Kajiura, K.; Kawakita, N.; Toba, H.; Yoshida, M.; et al. GAD1 expression and its methylation as indicators of malignant behavior in thymic epithelial tumors. Oncol. Lett. 2021, 21, 483. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, S.; Wang, Y.; Chen, Y.; Zhang, P.; Liu, Y.; Zhang, H.; Tao, Z.; Xiong, K. High expression of KITLG is a new hallmark activating the MAPK pathway in type A and AB thymoma. Thorac. Cancer 2020, 11, 1944–1954. [Google Scholar] [CrossRef]
- Arjonen, A.; Mäkelä, R.; Härmä, V.; Rintanen, N.; Kuopio, T.; Kononen, J.; Rantala, J.K. Image-based ex vivo drug screen to assess targeted therapies in recurrent thymoma. Lung Cancer 2020, 145, 27–32. [Google Scholar] [CrossRef]
- Giaccone, G.; Rajan, A.; Berman, A.; Kelly, R.J.; Szabo, E.; Lopez-Chavez, A.; Trepel, J.; Lee, M.-J.; Cao, L.; Espinoza-Delgado, I.; et al. Phase II study of belinostat in patients with recurrent or refractory advanced thymic epithelial tumors. J. Clin. Oncol. 2011, 29, 2052–2059. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Rajan, A.; Szabo, E.; Tomita, Y.; Carter, C.A.; Scepura, B.; Lopez-Chavez, A.; Lee, M.-J.; Redon, C.E.; Frosch, A.; et al. A phase I/II trial of belinostat in combination with cisplatin, doxorubicin, and cyclophosphamide in thymic epithelial tumors: A clinical and translational study. Clin. Cancer Res. 2014, 20, 5392–5402. [Google Scholar] [CrossRef] [Green Version]
- Bellissimo, T.; Ganci, F.; Gallo, E.; Sacconi, A.; Tito, C.; De Angelis, L.; Pulito, C.; Masciarelli, S.; Diso, D.; Anile, M.; et al. Thymic Epithelial Tumors phenotype relies on miR-145-5p epigenetic regulation. Mol. Cancer 2017, 16, 88. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Non-Coding RNA | Cell Lines/Tissue Samples | Methods | Main Results | References |
|---|---|---|---|---|
| 87 miRNAs | 54 thymic tumor samples, 12 normal counterparts | Reverse transcription quantitative real-time Polymerase Chain Reaction (RT-qPCR), Immunohistochemistry |
| [25] |
| 91 miRNAs | 17 type A, 38 type AB, 13 type B1, 31 type B2, 13 type B3, 11 type C thymomas | KEGG pathway analysis of miRNA target genes |
| [26] |
| miRNA-21-5p, miRNA-148a-3p, miRNA-141-3p, miRNA-34b-5p, miRNA-34c-5p, miRNA-455-5p | Peripheral blood samples from five patients with TEN | MicroRNA expression analysis |
| [27] |
| Large microRNA cluster on chr19q13.42 |
| RNA-sequencing, qPCR, protein analyses, drug sensitivity experiments |
| [28] |
| miRNA cluster C14MC on chromosome 14q32, miRNA-21, miRNA-9-3, miRNA-375, miRNA-34b, miRNA-34c, miRNA-130a, miRNA-195 | 18 type A, 19 type B3 thymomas, 35 TC | Cancer gene panel sequencing, miRNA sequencing, fluorescence in situ hybridization (FISH), immunohistochemistry |
| [29] |
| 65 differentially-expressed lncRNAs, 1695 miRNAs | 25 thymomas, 25 healthy thymus specimens | lncRNA-miRNA-mRNA functional enrichment analyses |
| [30] |
| lncRNAs ADAMTS9-AS1, HSD52, LINC00968, LINC01697 | TCGA | Statistics |
| [31] |
| lncRNA LINC00174 |
| Bioinformatics analysis of expression data from TCGA and Istituto Regina Elena thymoma cohorts, cDNA reverse transcriptase, RT-q PCR |
| [32] |
| miR-525-5p |
| RT-qPCR, dual luciferase reporter assay, cell counting kit 8 assay, flow cytometry, transwell assay, western blot analysis, tumor-burdened assay |
| [33] |
| miRNA-19b | nine type A, 11 type B1, 20 type B2, 12 type B3 thymomas, 11 normal thymi | Luciferase reporter assay, qRT-PCR, western blot |
| [34] |
| miRNA-20b |
| qRT-PCR, MTT assay, cell cycle analysis, immunostaining, flow cytometric analysis, western blot |
| [35] |
| RP11-424C20.2, miRNA-378a-3p, miRNA-422a | DreamBase, TCGA | RP11-424C20.2 cellular localization prediction, GO and KEGG enrichment analysis, correlation of UHRF1 expression with immune infiltration analysis |
| [36] |
| lncRNAs AC004943.1, WT1-AS, FOXG1-AS1 | 11 type A, 26 type AB, 10 type B1, 22 type B2, nine type B3 thymomas, 9 TC | Statistical analysis |
| [37] |
| 16 samples of type A, 35 samples of type AB, 57 samples of type B thymomas, 11 samples of TC, two samples of normal tissue. | GO and KEGG pathway annotation of DEmRNAs, overall survival analysis and establishment of the TEN-specific prognostic significance |
| [38] |
| DNA Methylation Sites | Cell Lines/Tissue Samples | Methods | Main Results | References |
|---|---|---|---|---|
| 5155 hyper-,6967 hypo-methylated CpG sites in type A–B3 TEN and TC | TCGA thymoma datasets with DNA methylation profiles of 124 tumor tissues and 2 matched adjacent normal tissues from 124 cases of thymoma, 54 thymomas, 46 TC | Statistical analysis, pyrosequencing |
| [39] |
| 10,000 CpG sites | 1 atypical type A, 1 type A, 1 type AB, 1 type B1, 1 type B2, 2 type B3 thymomas, 1 atypical TC | DNA isolation and bisulfite treatment, illumina 850K methylation microarray, statistical analysis |
| [40] |
| DNMT3B-579T allele | Peripheral blood of 324 AChR+ MG patients, 735 healthy matched unrelated controls, 94 patients with thymoma | Genotyping analysis performed by means of PCR-RFLP techniques |
| [41] |
| MLH1, MGMT, CDKN2A, RASSF1A | Blood samples and surgically resected thymomas from 69 patients with thymomatous MG and in the adjacent normal thymus available from 44 of them | Methylation sensitive-high resolution melting |
| [42] |
| MGMT | four type A, 12 type AB, 13 type B1, 7 type B2, eight type B3 thymomas, 23 TC | Nested methylation-specific PCR (MSP), immunohistochemical analysis |
| [43] |
| SPARC | six thymoma, five TC, 22 non-small cell lung carcinoma samples | MSP, mutation assay |
| [44] |
| hMLH1, MGMT, p-16INK4a, RASSF1A, FHIT, APC1A, RARB, DAPK, E-cadherin | 10 type A, 10 type AB, 12 type B1, nine type B2, seven type B3 thymomas, 17 TC | Real-time RT-PCR, nested MSP, immunohistologic analysis of global DNA methylation, measurement of global DNA methylation by an ELISA-like reaction |
| [45] |
| FBXW7 β-form promoter | one type A, three type AB, five type B1, three type B2, one type B3 thymomas | Genomic sodium bisulfite sequencing analysis |
| [46] |
| CDKN2 | 36 thymomas (non-invasive type, 16 cases; invasive type, 20 cases), three TC | Immunohistochemistry, PCR-SSCP, sequencing, PCR-based methylation assay, southern hybridization |
| [47] |
| DAPK, p-16, MGMT, HPP1 | one type A, six type AB, 10 type B1, four type B2, five type B3 thymomas, six TC | Sodium bisulfite modification |
| [48] |
| METTL3 |
| Immunohistochemistry, clonogenic assays, real-time qRT-PCR analysis, lysis and immunoblotting analysis, polysome profiling and treatment with puromycin, m6A immunoprecipitation, FISH |
| [49] |
| GNG4, GHSR, HOXD9, SALL3 | five type A, two type AB, four type B1, 10 type B2, and nine type B3 thymomas, 12 TC, four NECTT samples | Bisulfite conversion of genomic DNA, bisulfite pyrosequencing |
| [50] |
| GHSR | six type A, six type AB, eight type B1, 11 type B2, 10 type B3 thymomas, 13 TC, four NECTT samples | RT-qPCR, immunohistochemistry |
| [51] |
| GHSR | 12 type A, 12 type AB, five type B1, 22 type B2, eight type B3, five type B2-B3, one not specified thymoma samples | Bisulfite modification, methylation- sensitive high-resolution melting analysis |
| [52] |
| AIRE |
| Reverse transcriptase quantitative RT-PCR, bisulfite genomic DNA sequencing, quick and quantitative chromatin immunoprecipitation |
| [53] |
| POMC | three normal thymuses, one large cell lung cancer, five thymic carcinoid tumors | Immunohistochemistry, quantitation of POMC gene expression, bisulphite modification and sequencing |
| [54] |
| MTHFR, DNMT3A | 13 type A, 13 type AB, five type B1, 23 type B2, eight type B3, five type B2/3, two non-specified thymoma samples | Bisulfite modification, methylation- sensitive–high-resolution melting analysis |
| [55] |
| PTEN | three type A, eight type AB, 11 type B1, six type B2, five type B3 thymoma samples, four TC, two normal thymi | Immunohistochemistry, PCR direct sequencing, methylation-specific PCR, reverse transcription real-time PCR after target cell collection with laser microdissection |
| [56] |
| Dlc1 |
| Fluorescence microscopy, multiplex RT-PCR, western blotting, immunohistochemistry, microdissection, loss of heterozygosity analysis, DNA methylation study of Dlc1 promoter region, transendothelial migration and in vitro filipodia formation assay of the T lymphoma cells, treatment of cell culture with 5 azacytidine |
| [57] |
| TET2 | nine squamous cell carcinomas, one undifferentiated carcinoma | Exome sequencing, analysis of mutational signature, genome copy number analysis and tumor content estimation, whole transcriptome sequencing, DNA methylation analysis, gene ontology analysis |
| [58] |
| GAD1 | nine type A, 11 type AB, 19 type B1, 20 type B2, 14 type B3, 17 type C thymoma samples | Bisulfite conversion of genomic DNA, global methylation analysis, bisulfite pyrosequencing, RT-qPCR, immunohistochemistry |
| [59] |
| KITLG |
| RNA-seq data analysis, KITLG small-interfering RNA silencing, KITLG-overexpressing plasmid construction, real-time PCR, western blot analysis |
| [60] |
| HDACI | Cell Lines/Tissue Samples/Patient Collective | Methods | Main Results | References |
|---|---|---|---|---|
| Belinostat, Panobinostat | Bronchial metastatic lesion of a 54-year old female patient with a metastatic thymoma with pleural invasion | Mutation analysis, high content imaging drug screening |
| [61] |
| Belinostat | 25 patients with thymoma, 16 patients with TC | Pharmacodynamic analyses: Protein acetylation, peripheral blood mononuclear cell immune subsets, circulating angiogenic markers |
| [62] |
| Belinostat | two patients with type B1, seven with type B2, three with type B3 thymoma, 14 patients with TC | Monitoring for treatment-related adverse events, clinical laboratory tests, vital signs, physical examinations, 12-lead electrocardiograms, pharmacokinetic evaluations, multiparametric flow cytometry, CT scan |
| [63] |
| VPA | 10 peritumoral thymi, 14 thymomas, 5 TC | Microarray hybridization, RT-qPCR, immunohistochemistry, transfection and treatment, lysate preparation and immunoblotting analysis, transwell migration assay, clonogenic assays, immunocytochemistry and morphological analysis, cell cycle analysis, ATPlite luminescence assay system, formaldehyde cross-linking and chromatin immunoprecipitation, plasmid construction and dual-luciferase reporter assay |
| [64] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Psilopatis, I.; Pergaris, A.; Vrettou, K.; Theocharis, S.; Troungos, C. Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations. Int. J. Mol. Sci. 2022, 23, 4045. https://doi.org/10.3390/ijms23074045
Psilopatis I, Pergaris A, Vrettou K, Theocharis S, Troungos C. Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations. International Journal of Molecular Sciences. 2022; 23(7):4045. https://doi.org/10.3390/ijms23074045
Chicago/Turabian StylePsilopatis, Iason, Alexandros Pergaris, Kleio Vrettou, Stamatios Theocharis, and Constantinos Troungos. 2022. "Thymic Epithelial Neoplasms: Focusing on the Epigenetic Alterations" International Journal of Molecular Sciences 23, no. 7: 4045. https://doi.org/10.3390/ijms23074045