
Stromal Factors as a Target for Immunotherapy in Melanoma and Non-Melanoma Skin Cancers


Abstract
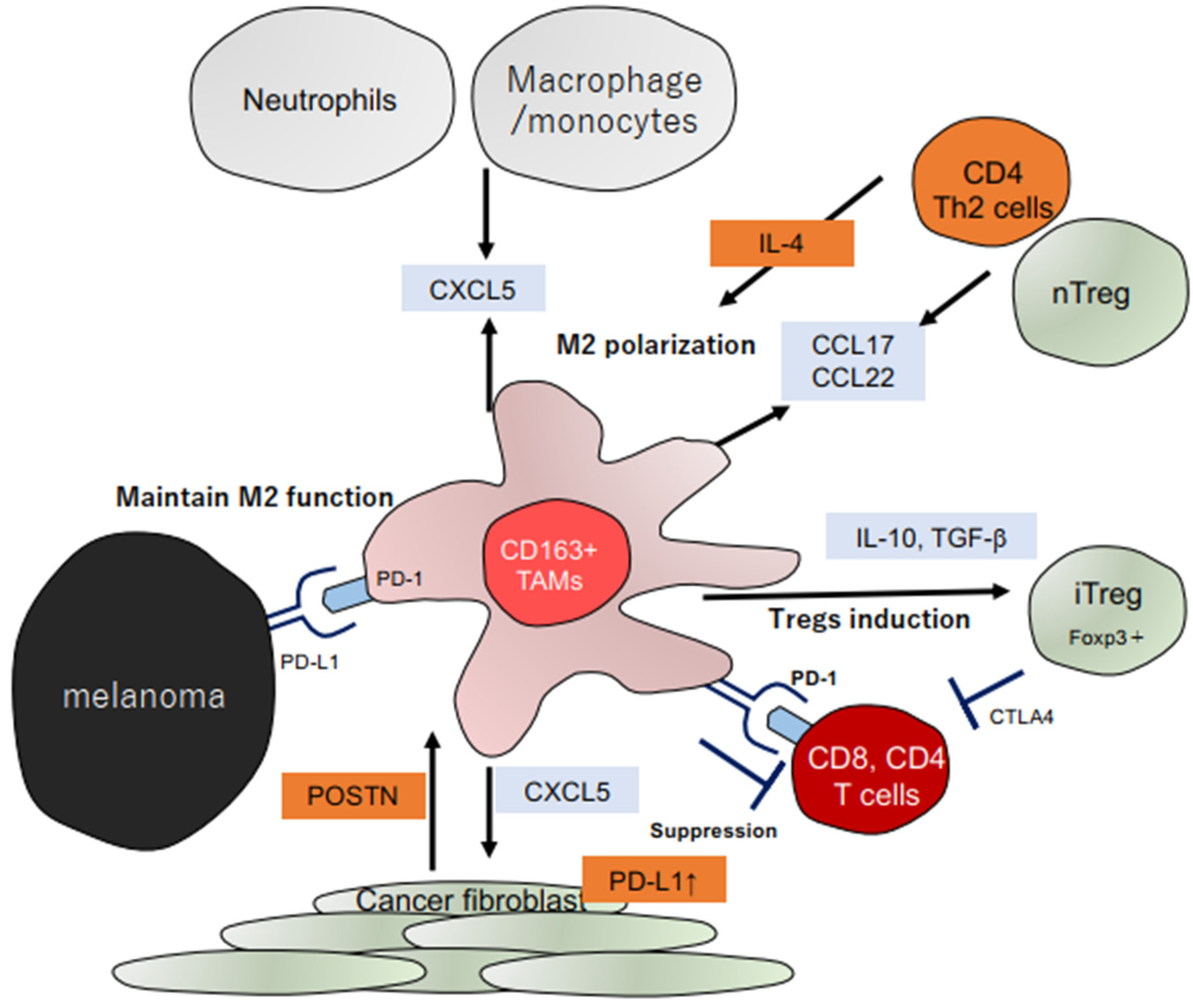
:1. Introduction
2. Profiles of Tumor-Infiltrating Leukocytes Determine the Characteristic Immunological Background of Skin Cancer
2.1. Importance of Stromal Factors in Developing Skin Cancers
2.1.1. Tumor-Promoting Roles of POSTN in Skin Cancers
2.1.2. PAI-1 in Skin Cancers
2.1.3. Other Stromal Factors in Skin Cancer
RANKL as an Immunostimulatory Molecule for TAMs
PD-L1 Expression in Skin Cancer
IL-4 as a Stromal Factor of Skin Cancer
2.2. Role of TAMs in Skin Cancers
2.2.1. Chemokine Production of TAMs
2.2.2. Angiogenetic Factors of TAMs
2.3. Other Stromal Cells in Skin Cancer
2.3.1. Tumor-Associated Neutrophils (TANs)
2.3.2. Cancer-Associated Fibroblasts (CAFs)
3. TILs in the Tumor Microenvironment as a Target to Improve Immunotherapy
4. Conclusions
Funding
Conflicts of Interest
References
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.J.; Rutkowski, P.; Lao, C.D.; Cowey, C.L.; Schadendorf, D.; Wagstaff, J.; Dummer, R.; et al. Five-Year Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2019, 381, 1535–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Kambayashi, Y.; Ohuchi, K.; Muto, Y.; Aiba, S. Treatment of Advanced Melanoma: Past, Present and Future. Life 2020, 16, 208. [Google Scholar] [CrossRef] [PubMed]
- Ishii, M.; Hirai, I.; Tanese, K.; Fusumae, T.; Nakamura, Y.; Fukuda, K.; Uchi, H.; Kabashima, K.; Otsuka, A.; Yokota, K.; et al. Anti-PD-1 antibody therapy for epithelial skin malignancies: An investigator-initiated, open-label, single-arm, multicenter, phase II clinical trial (NMSC-PD1 Study). Medicine 2020, 99, e22913. [Google Scholar] [CrossRef] [PubMed]
- Migden, M.R.; Khushalani, N.I.; Chang, A.L.S.; Lewis, K.D.; Schmults, C.D.; Hernandez-Aya, L.; Meier, F.; Schadendorf, D.; Guminski, A.; Hauschild, A.; et al. Cemiplimab in locally advanced cutaneous squamous cell carcinoma: Results from an open-label, phase 2, single-arm trial. Lancet Oncol. 2020, 21, 294–305. [Google Scholar] [CrossRef]
- Stratigos, A.J.; Sekulic, A.; Peris, K.; Bechter, O.; Prey, S.; Kaatz, M.; Lewis, K.D.; Basset-Seguin, N.; Chang, A.L.S.; Dalle, S.; et al. Cemiplimab in locally advanced basal cell carcinoma after hedgehog inhibitor therapy: An open-label, multi-centre, single-arm, phase 2 trial. Lancet Oncol. 2021, 22, 848–857. [Google Scholar] [CrossRef]
- Khodadoust, M.S.; Rook, A.H.; Porcu, P.; Foss, F.; Moskowitz, A.J.; Shustov, A.; Shanbhag, S.; Sokol, L.; Fling, S.P.; Ramchurren, N.; et al. Pembrolizumab in Relapsed and Refractory Mycosis Fungoides and Sézary Syndrome: A Multicenter Phase II Study. J. Clin. Oncol. 2020, 38, 20–28. [Google Scholar] [CrossRef]
- Fujimura, T.; Amagai, R.; Kambayashi, Y.; Aiba, S. Topical and Systemic Formulation Options for Cutaneous T Cell Lymphomas. Pharmaceutics 2021, 13, 200. [Google Scholar] [CrossRef]
- Wagner, M.J.; Othus, M.; Patel, S.P.; Ryan, C.; Sangal, A.; Powers, B.; Budd, G.T.; Victor, A.I.; Hsueh, C.T.; Chugh, R.; et al. Multicenter phase II trial (SWOG S1609, cohort 51) of ipilimumab and nivolumab in metastatic or unresectable angiosarcoma: A substudy of dual anti-CTLA-4 and anti-PD-1 blockade in rare tumors (DART). J. Immunother. Cancer 2021, 9, e002990. [Google Scholar] [CrossRef]
- Nakamura, Y.; Namikawa, K.; Yoshino, K.; Yoshikawa, S.; Uchi, H.; Goto, K.; Nakamura, Y.; Fukushima, S.; Kiniwa, Y.; Takenouchi, T.; et al. Anti-PD1 checkpoint inhibitor therapy in acral melanoma: A multicenter study of 193 Japanese patients. Ann. Oncol. 2020, 31, 1198–1206. [Google Scholar] [CrossRef]
- Hayward, N.K.; Wilmott, J.S.; Waddell, N.; Johansson, P.A.; Field, M.A.; Nones, K.; Patch, A.M.; Kakavand, H.; Alexandrov, L.B.; Burke, H.; et al. Whole-genome landscapes of major melanoma subtypes. Nature 2017, 545, 175–180. [Google Scholar] [CrossRef]
- Fujimura, T.; Aiba, S. Significance of Immunosuppressive Cells as a Target for Immunotherapies in Melanoma and Non-Melanoma Skin Cancers. Biomolecules 2020, 10, 1087. [Google Scholar] [CrossRef] [PubMed]
- Taube, J.M.; Klein, A.; Brahmer, J.R.; Xu, H.; Pan, X.; Kim, J.H.; Chen, L.; Pardoll, D.M.; Topalian, S.L.; Anders, R.A. Association of PD-1, PD-1 ligands, and other features of the tumor immune microenvironment with response to anti-PD-1 therapy. Clin. Cancer Res. 2014, 20, 5064–5074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Green, M.D.; Li, S.; Sun, Y.; Journey, S.N.; Choi, J.E.; Rizvi, S.M.; Qin, A.; Waninger, J.J.; Lang, X.; et al. Liver metastasis restrains immunotherapy efficacy via macrophage-mediated T cell elimination. Nat. Med. 2021, 27, 152–164. [Google Scholar] [CrossRef] [PubMed]
- Blank, C.U.; Rozeman, E.A.; Fanchi, L.F.; Sikorska, K.; van de Wiel, B.; Kvistborg, P.; Krijgsman, O.; van den Braber, M.; Philips, D.; Broeks, A.; et al. Neoadjuvant versus adjuvant ipilimumab plus nivolumab in macroscopic stage III melanoma. Nat. Med. 2018, 24, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Okuyama, R.; Ohtani, T.; Ito, Y.; Haga, T.; Hashimoto, A.; Aiba, S. Perilesional treatment of metastatic melanoma with interferon-beta. Clin. Exp. Dermatol. 2009, 34, 793–799. [Google Scholar] [CrossRef]
- Homet Moreno, B.; Mok, S.; Comin-Anduix, B.; Hu-Lieskovan, S.; Ribas, A. Combined treatment with dabrafenib and trametinib with immune-stimulating antibodies for BRAF mutant melanoma. Oncoimmunology 2015, 5, e1052212. [Google Scholar] [CrossRef] [Green Version]
- Ohuchi, K.; Kambayashi, Y.; Hidaka, T.; Fujimura, T. Plasminogen activating inhibitor-1 might be a predictive marker for the efficacy of anti-PD1 antibody in advanced melanoma patients. Front. Oncol. 2021, 11, 798385. [Google Scholar] [CrossRef]
- Fujimura, T.; Kambayashi, Y.; Fujisawa, Y.; Hidaka, T.; Aiba, S. Tumor-associated macrophages: Therapeutic targets for skin cancer. Front Oncol 2018, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Tjiu, J.W.; Chen, J.S.; Shun, C.T.; Lin, S.J.; Liao, Y.H.; Chu, C.Y.; Tsai, T.F.; Chiu, H.C.; Dai, Y.S.; Inoue, H.; et al. Tumor-associated macrophage-induced invasion and angiogenesis of human basal cell carcinoma cells by cyclooxygenase-2 induction. J. Investig. Dermatol. 2009, 129, 1016–1025. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Kambayashi, Y.; Hidaka, T.; Hashimoto, A.; Haga, T.; Aiba, S. Comparison of Foxp3+ regulatory T cells and CD163+ macrophages in invasive and non-invasive extramammary Paget’s disease. Acta Derm. Venereol. 2012, 92, 625–628. [Google Scholar] [CrossRef] [Green Version]
- Kervarrec, T.; Gaboriaud, P.; Berthon, P.; Zaragoza, J.; Schrama, D.; Houben, R.; Le Corre, Y.; Hainaut-Wierzbicka, E.; Aubin, F.; Bens, G.; et al. Merkel cell carcinomas infiltrated with CD33+ myeloid cells and CD8+ T cells are associated with improved outcome. J. Am. Acad. Dermatol. 2018, 78, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Ring, S.; Umansky, V.; Mahnke, K.; Enk, A.H. Regulatory T cells (Treg) stimulate B7-H1 expression in myeloid derived suppressor cells (MDSC) in ret melanomas. J. Investig. Dermatol. 2012, 132, 1239–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kambayashi, Y.; Fujimura, T.; Hidaka, T.; Aiba, S. Biomarkers for Predicting Efficacies of Anti-PD1 Antibodies. Front. Med. 2019, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Sato, Y.; Tanita, K.; Kambayashi, Y.; Otsuka, A.; Fujisawa, Y.; Yoshino, K.; Matsushita, S.; Funakoshi, T.; Hata, H.; et al. Serum soluble CD163 and CXCL5 could be predictive markers for immune related adverse event in patients with advanced melanoma treated with nivolumab. Oncotarget 2018, 9, 15542–15551. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Sato, Y.; Tanita, K.; Kambayash, Y.; Otsuka, A.; Fujisawa, Y.; Yoshino, K.; Matsushita, S.; Funakoshi, T.; Hata, H.; et al. Serum level of soluble CD163 may be a predictive marker of the effectiveness of nivolumab in patients with advanced cutaneous melanoma. Front. Oncol. 2018, 8, 530. [Google Scholar] [CrossRef]
- Fujimura, T.; Sato, Y.; Tanita, K.; Lyu, C.; Kambayash, Y.; Amagai, R.; Otsuka, A.; Fujisawa, Y.; Yoshino, K.; Matsushita, S.; et al. Association of baseline serum levels of CXCL5 with the efficacy of nivolumab in advanced melanoma. Front. Med. 2019, 6, 86. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Tanita, K.; Sato, Y.; Lyu, C.; Kambayashi, Y.; Fujisawa, Y.; Uchi, H.; Yamamoto, Y.; Otsuka, A.; Yoshino, K.; et al. Immune checkpoint inhibitor-induced vitiligo in advanced melanoma could be related to increased levels of CCL19. Br. J. Dermatol. 2020, 182, 1297–1300. [Google Scholar] [CrossRef]
- Fujimura, T.; Tanita, K.; Ohuchi, K.; Sato, Y.; Lyu, C.; Kambayashi, Y.; Fujisawa, Y.; Tanaka, R.; Hashimoto, A.; Aiba, S. Increased serum CCL26 level is a potential biomarker for the effectiveness of anti-PD1 antibodies in patients with advanced melanoma. Melanoma Res. 2020, 30, 613–618. [Google Scholar] [CrossRef]
- Fukuda, K.; Sugihara, E.; Ohta, S.; Izuhara, K.; Funakoshi, T.; Amagai, M.; Saya, H. Periostin Is a Key Niche Component for Wound Metastasis of Melanoma. PLoS ONE 2015, 10, e0129704. [Google Scholar] [CrossRef]
- Kambayashi, Y.; Fujimura, T.; Furudate, S.; Asano, M.; Kakizaki, A.; Aiba, S. The possible interaction between receptor activator of nuclear factor kappa-B ligand (RANKL) expressed by extramammary Paget cells and its ligand on dermal macrophages. J. Investig. Dermatol. 2015, 135, 2547–2550. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.T.; McLeod, K.; Kim, S.; Conway, S.J.; Hamilton, D.W. Periostin as a multifunctional modulator of the wound healing response. Cell Tissue Res. 2016, 365, 453–465. [Google Scholar] [CrossRef] [Green Version]
- Orecchia, P.; Conte, R.; Balza, E.; Castellani, P.; Borsi, L.; Zardi, L.; Mingari, M.C.; Carnemolla, B. Identification of a novel cell binding site of periostin involved in tumour growth. Eur. J. Cancer 2011, 47, 2221–2229. [Google Scholar] [CrossRef]
- Yoshida, H.; Koodie, L.; Jacobsen, K.; Hanzawa, K.; Miyamoto, Y.; Yamamoto, M. B4GALNT1 induces angiogenesis, anchorage independence growth and motility, and promotes tumorigenesis in melanoma by induction of ganglioside GM2/GD2. Sci. Rep. 2020, 10, 1199. [Google Scholar] [CrossRef] [Green Version]
- Ohno, F.; Nakahara, T.; Kido-Nakahara, M.; Ito, T.; Nunomura, S.; Izuhara, K.; Furue, M. Periostin Links Skin Inflammation to Melanoma Progression in Humans and Mice. Int. J. Mol. Sci. 2019, 20, 169. [Google Scholar] [CrossRef] [Green Version]
- Lincoln, V.; Chao, L.; Woodley, D.T.; Murrell, D.; Kim, M.; O’Toole, E.A.; Ly, A.; Cogan, J.; Mosallaei, D.; Wysong, A.; et al. Over-expression of stromal periostin correlates with poor prognosis of cutaneous squamous cell carcinomas. Exp. Dermatol. 2021, 30, 698–704. [Google Scholar] [CrossRef]
- Fujimura, T.; Kambayashi, Y.; Furudate, S.; Kakizaki, A.; Hidaka, T.; Haga, T.; Hashimoto, A.; Morimoto, R.; Aiba, S. Isolated adrenocorticotropic hormone deficiency possibly caused by nivolumab in a metastatic melanoma patient. J. Dermatol. 2017, 44, e13–e14. [Google Scholar] [CrossRef] [Green Version]
- Villani, R.; Murigneux, V.; Alexis, J.; Sim, S.L.; Wagels, M.; Saunders, N.; Soyer, H.P.; Parmentier, L.; Nikolaev, S.; Fink, J.L.; et al. Subtype-Specific Analyses Reveal Infiltrative Basal Cell Carcinomas Are Highly Interactive with their Environment. J. Investig. Dermatol. 2021, 141, 2380–2390. [Google Scholar] [CrossRef]
- Fujimura, T.; Kakizaki, A.; Sato, Y.; Tanita, K.; Furudate, S.; Aiba, S. The Immunological Roles of Periostin/Tumor-Associated Macrophage Axis in Development of Dermatofibrosarcoma Protuberans. Anticancer Res 2017, 37, 2867–2873. [Google Scholar]
- Fujimura, T.; Furudate, S.; Kambayahsi, Y.; Kakizaki, A.; Yamamoto, Y.; Okuhira, H.; Fujimoto, N.; Aiba, S. Phospho-STAT5B Expression Is a Prognostic Marker for Merkel Cell Carcinoma. Anticancer Res. 2017, 37, 2335–2341. [Google Scholar] [CrossRef] [Green Version]
- Furudate, S.; Fujimura, T.; Kakizaki, A.; Kambayashi, Y.; Asano, M.; Watabe, A.; Aiba, S. The possible interaction between periostin expressed by cancer stroma and tumor-associated macrophages in developing mycosis fungoides. Exp. Dermatol. 2016, 25, 107–112. [Google Scholar] [CrossRef]
- Fujimura, T.; Okabe, T.; Tanita, K.; Sato, Y.; Lyu, C.; Kambayashi, Y.; Maruyama, S.; Aiba, S. A novel technique to diagnose non-melanoma skin cancer by thermal conductivity measurements: Correlations with cancer stromal factors. Exp. Dermatol. 2019, 28, 1029–1035. [Google Scholar] [CrossRef]
- Rubina, K.A.; Sysoeva, V.Y.; Zagorujko, E.I.; Tsokolaeva, Z.I.; Kurdina, M.I.; Parfyonova, Y.V.; Tkachuk, V.A. Increased expression of uPA, uPAR, and PAI-1 in psoriatic skin and in basal cell carcinomas. Arch. Dermatol. Res. 2017, 309, 433–442. [Google Scholar] [CrossRef]
- Klein, R.M.; Bernstein, D.; Higgins, S.P.; Higgins, C.E.; Higgins, P.J. SERPINE1 expression discriminates site-specific metastasis in human melanoma. Exp. Dermatol. 2012, 21, 551–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.J.; Lee, C.H.; Chen, W.Y.; Yang, J.L.; Tzeng, H.T. Inhibition of PAI-1 Blocks PD-L1 Endocytosis and Improves the Response of Melanoma Cells to Immune Checkpoint Blockade. J. Investg. Dermatol. 2021, 141, 2690–2698. [Google Scholar] [CrossRef]
- Del Rosso, M.; Fibbi, G.; Laurenzana, A.; Margheri, F.; Chillà, A. A Possible Role for PAI-1 Blockade in Melanoma Immunotherapy. J. Investig. Dermatol. 2021, 141, 2566–2568. [Google Scholar] [CrossRef]
- Tzeng, H.T.; Yang, J.L.; Tseng, Y.J.; Lee, C.H.; Chen, W.J.; Chyuan, I.T. Plasminogen Activator Inhibitor-1 Secretion by Autophagy Contributes to Melanoma Resistance to Chemotherapy through Tumor Microenvironment Modulation. Cancers 2021, 13, 1253. [Google Scholar] [CrossRef]
- Fujimura, T.; Yoshino, K.; Kato, H.; Fukushima, S.; Otsuka, A.; Matsushita, S.; Miyata, T.; Amagai, R.; Ohuchi, K.; Furudate, S.; et al. Anti-PD-1 plus TM5614 immunotherapy for unresectable malignant melanoma: An investigator-initiated, open-label, single-arm, multicentre, phase 2 clinical trial. Med. Protoc. Case Rep. 2021, 2, e0197. [Google Scholar]
- Thapa, B.; Koo, B.H.; Kim, Y.H.; Kwon, H.J.; Kim, D.S. Plasminogen activator inhibitor-1 regulates infiltration of macrophages into melanoma via phosphorylation of FAK-Tyr925. Biochem. Biophys. Res. Commun. 2014, 450, 1696–1701. [Google Scholar] [CrossRef]
- Kadeh, H.; Saravani, S.; Heydari, F.; Shahraki, S. Differential immunohistochemical expression of matrix metalloproteinase-10 (MMP-10) in non-melanoma skin cancers of the head and neck. Pathol. Res. Pract. 2016, 212, 867–871. [Google Scholar] [CrossRef]
- Freytag, J.; Wilkins-Port, C.E.; Higgins, C.E.; Carlson, J.A.; Noel, A.; Foidart, J.M.; Higgins, S.P.; Samarakoon, R.; Higgins, P.J. PAI-1 Regulates the Invasive Phenotype in Human Cutaneous Squamous Cell Carcinoma. J. Oncol. 2009, 2009, 963209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins-Port, C.E.; Higgins, C.E.; Freytag, J.; Higgins, S.P.; Carlson, J.A.; Higgins, P.J. PAI-1 is a Critical Upstream Regulator of the TGF-beta1/EGF-Induced Invasive Phenotype in Mutant p53 Human Cutaneous Squamous Cell Carcinoma. J. Biomed. Biotechnol. 2007, 2007, 85208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguire, T.; Chin, D.; Soutar, D.; Duffy, M.J. Low levels of urokinase plasminogen activator components in basal cell carcinoma of the skin. Int. J. Cancer 2000, 85, 457–459. [Google Scholar] [CrossRef]
- Ilich, A.; Kumar, V.; Henderson, M.; Mallick, R.; Wells, P.; Carrier, M.; Key, N.S. Biomarkers in cancer patients at risk for venous thromboembolism: Data from the AVERT study. Thromb. Res. 2020, 191 (Suppl. 1), S31–S36. [Google Scholar] [CrossRef]
- Boutsikas, G.; Terpos, E.; Markopoulos, A.; Papatheodorou, A.; Stefanou, A.; Georgiou, G.; Galani, Z.; Komninaka, V.; Telonis, V.; Anagnostopoulos, I.; et al. Protein Z (PZ) and plasminogen activator inhibitor-1 (PAI-1) plasma levels in patients with previously untreated Hodgkin lymphoma (HL). J. BU ON Off. J. Balk. Union Oncol. 2017, 22, 1022–1031. [Google Scholar]
- Yahata, T.; Ibrahim, A.A.; Hirano, K.I.; Muguruma, Y.; Naka, K.; Hozumi, K.; Vaughan, D.E.; Miyata, T.; Ando, K. Targeting of plasminogen activator inhibitor-1 activity promotes elimination of chronic myeloid leukemia stem cells. Haematologica 2021, 106, 483–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noy, R.; Pollard, J.W. Tumor-associated macrophages: From mechanisms to therapy. Immunity 2014, 41, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, T.; Kakizaki, A.; Furudate, S.; Kambayashi, Y.; Aiba, S. Tumor-associated macrophages in skin: How to treat their heterogeneity and plasticity. J. Dermatol. Sci. 2016, 83, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Aleza, C.; Nguyen, B.; Yoldi, G.; Ciscar, M.; Barranco, A.; Hernández-Jiménez, E.; Maetens, M.; Salgado, R.; Zafeiroglou, M.; Pellegrini, P.; et al. Inhibition of RANK signaling in breast cancer induces an anti-tumor immune response orchestrated by CD8+ T cells. Nat. Commun. 2020, 11, 6335. [Google Scholar] [CrossRef] [PubMed]
- Angela, Y.; Haferkamp, S.; Weishaupt, C.; Ugurel, S.; Becker, J.C.; Oberndörfer, F.; Alar, V.; Satzger, I.; Gutzmer, R. Combination of denosumab and immune checkpoint inhibition: Experience in 29 patients with metastatic melanoma and bone metastases. Cancer Immunol. Immunother. 2019, 68, 1187–1194. [Google Scholar] [CrossRef]
- Liede, A.; Hernandez, R.K.; Wade, S.W.; Bo, R.; Nussbaum, N.C.; Ahern, E.; Dougall, W.C.; Smyth, M.J. An observational study of concomitant immunotherapies and denosumab in patients with advanced melanoma or lung cancer. Oncoimmunology 2018, 5, e1480301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amôr, N.G.; Santos, P.S.D.S.; Campanelli, A.P. The Tumor Microenvironment in SCC: Mechanisms and Therapeutic Opportunities. Front. Cell Dev. Biol. 2021, 9, 636544. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Kambayashi, Y.; Tono, H.; Lyu, C.; Ohuchi, K.; Hashimoto, A.; Aiba, S. Successful treatment of unresectable recurrent cutaneous squamous cell carcinoma of the scalp with meningeal invasion with nivolumab monotherapy. Dermatol. Ther. 2020, 33, e13672. [Google Scholar] [CrossRef] [PubMed]
- Lipson, E.J.; Vincent, J.G.; Loyo, M.; Kagohara, L.T.; Luber, B.S.; Wang, H.; Xu, H.; Nayar, S.K.; Wang, T.S.; Sidransky, D.; et al. PD-L1 expression in the Merkel cell carcinoma microenvironment: Association with inflammation, Merkel cell polyomavirus and overall survival. Cancer Immunol. Res. 2013, 1, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iga, N.; Otsuka, A.; Yamamoto, Y.; Nakashima, C.; Honda, T.; Kitoh, A.; Nakajima, S.; Egawa, G.; Nomura, T.; Dainichi, T.; et al. Accumulation of exhausted CD8+ T cells in extramammary Paget’s disease. PLoS ONE 2019, 14, e0211135. [Google Scholar] [CrossRef]
- Fujimura, T.; Kambayashi, Y.; Kakizaki, A.; Furudate, S.; Aiba, S. RANKL expression is a useful marker for differentiation of pagetoid squamous cell carcinoma in situ from extramammary Paget disease. J. Cutan. Pathol. 2016, 43, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Honda, Y.; Otsuka, A.; Ono, S.; Yamamoto, Y.; Seidel, J.A.; Morita, S.; Hirata, M.; Kataoka, T.R.; Takenouchi, T.; Fujii, K.; et al. Infiltration of PD-1-positive cells in combination with tumor site PD-L1 expression is a positive prognostic factor in cutaneous angiosarcoma. Oncoimmunology 2016, 6, e1253657. [Google Scholar] [CrossRef] [Green Version]
- Hino, R.; Kabashima, K.; Kato, Y.; Yagi, H.; Nakamura, M.; Honjo, T.; Okazaki, T.; Tokura, Y. Tumor cell expression of programmed cell death-1 ligand 1 is a prognostic factor for malignant melanoma. Cancer 2010, 116, 1757–1766. [Google Scholar] [CrossRef] [PubMed]
- Carlino, M.S.; Long, G.V.; Schadendorf, D.; Robert, C.; Ribas, A.; Richtig, E.; Nyakas, M.; Caglevic, C.; Tarhini, A.; Blank, C.; et al. Outcomes by line of therapy and programmed death ligand 1 expression in patients with advanced melanoma treated with pembrolizumab or ipilimumab in KEYNOTE-006: A randomised clinical trial. Eur. J. Cancer 2018, 101, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D.; et al. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef] [PubMed]
- Rischin, D.; Migden, M.R.; Lim, A.M.; Schmults, C.D.; Khushalani, N.I.; Hughes, B.G.M.; Schadendorf, D.; Dunn, L.A.; Hernandez-Aya, L.; Chang, A.L.S.; et al. Phase 2 study of cemiplimab in patients with metastatic cutaneous squamous cell carcinoma: Primary analysis of fixed-dosing, long-term outcome of weight-based dosing. J. Immunother. Cancer 2020, 8, e000775. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, S.P.; Bhatia, S.; Brohl, A.S.; Hamid, O.; Mehnert, J.M.; Terheyden, P.; Shih, K.C.; Brownell, I.; Lebbé, C.; Lewis, K.D.; et al. Avelumab in patients with previously treated metastatic Merkel cell carcinoma (JAVELIN Merkel 200): Updated overall survival data after >5 years of follow-up. ESMO Open 2021, 6, 100290. [Google Scholar] [CrossRef] [PubMed]
- Tanita, K.; Fujimura, T.; Sato, Y.; Lyu, C.; Kambayashi, Y.; Ogata, D.; Fukushima, S.; Miyashita, A.; Nakajima, H.; Nakamura, M.; et al. Bexarotene Reduces Production of CCL22 From Tumor-Associated Macrophages in Cutaneous T-Cell Lymphoma. Front. Oncol. 2019, 9, 907. [Google Scholar] [CrossRef] [PubMed]
- Van Gorp, H.; Delputte, P.L.; Nauwynck, H.J. Scavenger receptor CD163, a Jack-of-all-trades and potential target for cell-directed therapy. Mol. Immunol. 2010, 47, 1650–1660. [Google Scholar] [CrossRef] [PubMed]
- Sugaya, M.; Miyagaki, T.; Ohmatsu, H.; Suga, H.; Kai, H.; Kamata, M.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; et al. Association of the numbers of CD163+ cells in lesional skin and serum levels of soluble CD163 with disease progression of cutaneous T cell lymphoma. J. Dermatol. Sci. 2012, 68, 45–51. [Google Scholar] [CrossRef]
- Furudate, S.; Fujimura, T.; Kakizaki, A.; Hidaka, T.; Asano, M.; Aiba, S. Tumor-associated M2 macrophages in mycosis fungoides acquire immunomodulatory function by interferon alpha and interferon gamma. J. Dermatol. Sci. 2016, 83, 182–189. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, J.; Zhang, J.; Li, S.; Wang, H.; Du, J. Cancer-associated fibroblasts promote PD-L1 expression in mice cancer cells via secreting CXCL5. Int. J. Cancer 2019, 145, 1946–1957. [Google Scholar] [CrossRef] [Green Version]
- Serafini, P.; Mgebroff, S.; Noonan, K.; Borrello, I. Myeloid-derived suppressor cells promote cross-tolerance in B-cell lymphoma by expanding regulatory T cells. Cancer Res. 2008, 68, 5439–5449. [Google Scholar] [CrossRef] [Green Version]
- Fujimura, T.; Kambayashi, Y.; Furudate, S.; Asano, M.; Kakizaki, A.; Aiba, S. Receptor Activator of NF-κB Ligand Promotes the Production of CCL17 from RANK+ M2 Macrophages. J. Investig. Dermatol. 2015, 135, 2884–2887. [Google Scholar] [CrossRef] [Green Version]
- Lyu, C.; Fujimura, T.; Amagai, R.; Ohuchi, K.; Sato, Y.; Tanita, K.; Matsushita, S.; Fujisawa, Y.; Otsuka, A.; Yamamoto, Y.; et al. Increased expression of dermal LL37 may trigger migration of CCR7+ regulatory T cells in extramammary Paget’s disease. J. Dermatol. Sci. 2020, 99, 65–68. [Google Scholar] [CrossRef]
- Sato, Y.; Fujimura, T.; Tanita, K.; Lyu, C.; Matsushita, S.; Fujisawa, Y.; Otsuka, A.; Yamamoto, Y.; Hidaka, T.; Aiba, S. Malassezia-derived aryl hydrocarbon receptor (AhR) ligand augments CCL20/Th17/soluble CD163 pathogenic axis in extramammary Paget’s disease. Exp. Dermatol. 2019, 28, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, J.S.; Fuentes-Duculan, J.; Suárez-Fariñas, M.; Pierson, K.C.; Pitts-Kiefer, A.; Fan, L.; Belkin, D.A.; Wang, C.Q.; Bhuvanendran, S.; Johnson-Huang, L.M.; et al. Tumor-associated macrophages in the cutaneous SCC microenvironment are heterogeneously activated. J. Investig. Dermatol. 2011, 131, 1322–1330. [Google Scholar] [CrossRef] [Green Version]
- Amôr, N.G.; de Oliveira, C.E.; Gasparoto, T.H.; Boas, V.G.V.; Perri, G.; Kaneno, R.; Lara, V.S.; Garlet, G.P.; da Silva, J.S.; Martins, G.A.; et al. ST2/IL-33 signaling promotes malignant development of experimental squamous cell carcinoma by decreasing NK cells cytotoxicity and modulating the intratumoral cell infiltrate. Oncotarget 2018, 9, 25768. [Google Scholar] [CrossRef] [Green Version]
- Gaiser, M.R.; Weis, C.A.; Gaiser, T.; Jiang, H.; Buder-Bakhaya, K.; Herpel, E.; Warth, A.; Xiao, Y.; Miao, L.; Brownell, I. Merkel cell carcinoma expresses the immunoregulatory ligand CD200 and induces immunosuppressive macrophages and regulatory T cells. Oncoimmunology 2018, 7, e1426517. [Google Scholar] [CrossRef]
- Werchau, S.; Toberer, F.; Enk, A.; Dammann, R.; Helmbold, P. Merkel cell carcinoma induces lymphatic microvessel formation. J. Am. Acad. Dermatol. 2012, 67, 215–225. [Google Scholar] [CrossRef]
- Atzori, M.G.; Ceci, C.; Ruffini, F.; Trapani, M.; Barbaccia, M.L.; Tentori, L.; D’Atri, S.; Lacal, P.M.; Graziani, G. Role of VEGFR-1 in melanoma acquired resistance to the BRAF inhibitor vemurafenib. J. Cell Mol. Med. 2020, 24, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Linde, N.; Lederle, W.; Depner, S.; van Rooijen, N.; Gutschalk, C.M.; Mueller, M.M. Vascular endothelial growth factor-induced skin carcinogenesis depends on recruitment and alternative activation of macrophages. J. Pathol. 2012, 227, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimian, T.G.; Squiban, C.; Roque, T.; Lugo-Martinez, H.; Hneino, M.; Buard, V.; Gourmelon, P.; Benderitter, M.; Milliat, F.; Tamarat, R. Plasminogen activator inhibitor-1 controls bone marrow-derived cells therapeutic effect through MMP9 signaling: Role in physiological and pathological wound healing. Stem Cells 2012, 30, 1436–1446. [Google Scholar] [CrossRef]
- Kale, S.; Raja, R.; Thorat, D.; Soundararajan, G.; Patil, T.V.; Kundu, G.C. Osteopontin signaling upregulates cyclooxygenase-2 expression in tumor-associated macrophages leading to enhanced angiogenesis and melanoma growth via α9β1 integrin. Oncogene 2014, 33, 2295–2306. [Google Scholar] [CrossRef] [PubMed]
- Kambayashi, Y.; Fujimura, T.; Furudate, S.; Lyu, C.; Hidaka, T.; Kakizaki, A.; Sato, Y.; Tanita, K.; Aiba, S. The Expression of Matrix Metalloproteinases in Receptor Activator of Nuclear Factor Kappa-B Ligand (RANKL)-expressing Cancer of Apocrine Origin. Anticancer Res. 2018, 38, 113–120. [Google Scholar] [PubMed] [Green Version]
- Litvinov, I.V.; Netchiporouk, E.; Cordeiro, B.; Doré, M.A.; Moreau, L.; Pehr, K.; Gilbert, M.; Zhou, Y.; Sasseville, D.; Kupper, T.S. The Use of Transcriptional Profiling to Improve Personalized Diagnosis and Management of Cutaneous T-cell Lymphoma (CTCL). Clin. Cancer Res. 2015, 21, 2820–2829. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Guo, Y.; Zhu, P.; Zhang, D.; Liu, S.; Tang, M.; Wang, Y.; Jin, Z.; Li, D.; Yan, D.; et al. TRIM59 loss in M2 macrophages promotes melanoma migration and invasion by upregulating MMP-9 and Madcam1. Aging 2019, 11, 8623–8641. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.R.; Huttenlocher, A. Neutrophils in the tumor microenvironment. Trends Immunol. 2016, 37, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffelt, S.B.; Kersten, K.; Doornebal, C.W.; Weiden, J.; Vrijland, K.; Hau, C.S.; Verstegen, N.J.M.; Ciampricotti, M.; Hawinkels, L.J.A.C.; Jonkers, J.; et al. IL-17-producing γδ T cells and neutrophils conspire to promote breast cancer metastasis. Nature 2015, 522, 345–348. [Google Scholar] [CrossRef]
- Uribe-Querol, E.; Rosales, C. Neutrophils in cancer: Two sides of the same coin. J. Immunol. Res. 2015, 2015, 983698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Chen, X.; Zhao, J.; Martin, B.; Zepp, J.A.; Ko, J.S.; Gu, C.; Cai, G.; Ouyang, W.; Sen, G.; et al. A novel IL-17 signaling pathway controlling keratinocyte proliferation and tumorigenesis via the TRAF4-ERK5 axis. J. Exp. Med. 2015, 212, 1571–1587. [Google Scholar] [CrossRef]
- Gasparoto, T.H.; de Oliveira, C.E.; de Freitas, L.T.; Pinheiro, C.R.; Ramos, R.N.; da Silva, A.L.; Cavassani, K.A.; Campanelli, A.P. Inflammatory events during murine squamous cell carcinoma development. J. Inflamm. 2012, 9, 46. [Google Scholar] [CrossRef] [Green Version]
- Sato, Y.; Fujimura, T.; Hidaka, T.; Lyu, C.; Tanita, K.; Matsushita, S.; Yamamoto, M.; Aiba, S. Possible Roles of Proinflammatory Signaling in Keratinocytes Through Aryl Hydrocarbon Receptor Ligands for the Development of Squamous Cell Carcinoma. Front. Immunol. 2020, 11, 534323. [Google Scholar] [CrossRef]
- Romano, V.; Belviso, I.; Venuta, A.; Ruocco, M.R.; Masone, S.; Aliotta, F.; Fiume, G.; Montagnani, S.; Avagliano, A.; Arcucci, A. Influence of Tumor Microenvironment and Fibroblast Population Plasticity on Melanoma Growth, Therapy Resistance and Immunoescape. Int. J. Mol. Sci. 2021, 22, 5283. [Google Scholar] [CrossRef]
- Omland, S.H.; Wettergren, E.E.; Mollerup, S.; Asplund, M.; Mourier, T.; Hansen, A.J.; Gniadecki, R. Cancer associated fibroblasts (CAFs) are activated in cutaneous basal cell carcinoma and in the peritumoural skin. BMC Cancer 2017, 17, 675. [Google Scholar] [CrossRef]
- Wong, P.F.; Wei, W.; Gupta, S.; Smithy, J.W.; Zelterman, D.; Kluger, H.M.; Rimm, D.L. Multiplex quantitative analysis of cancer-associated fibroblasts and immunotherapy outcome in metastatic melanoma. J. Immunother. Cancer 2019, 7, 197. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhou, L.; Xiao, Y.; Andl, T.; Zhang, Y. BRAF Inhibitors Reprogram Cancer-Associated Fibroblasts to Drive Matrix Remodeling and Therapeutic Escape in Melanoma. Cancer Res. 2022, 82, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Berestjuk, I.; Lecacheur, M.; Carminati, A.; Diazzi, S.; Rovera, C.; Prod’homme, V.; Ohanna, M.; Popovic, A.; Mallavialle, A.; Larbret, F.; et al. Targeting Discoidin Domain Receptors DDR1 and DDR2 overcomes matrix-mediated tumor cell adaptation and tolerance to BRAF-targeted therapy in melanoma. EMBO Mol. Med. 2021, 14, e11814. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Spassova, I.; Gravemeyer, J.; Ritter, C.; Horny, K.; Lange, A.; Gambichler, T.; Ødum, N.; Schrama, D.; Schadendorf, D.; et al. Merkel cell carcinoma-derived exosome-shuttle miR-375 induces fibroblast polarization by inhibition of RBPJ and p53. Oncogene 2021, 40, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, S.J.; Moerman-Herzog, A.; Wong, H.K. Normal and cancer fibroblasts differentially regulate TWIST1, TOX and cytokine gene expression in cutaneous T-cell lymphoma. BMC Cancer 2021, 21, 492. [Google Scholar] [CrossRef]
- Yagi, T.; Baba, Y.; Okadome, K.; Kiyozumi, Y.; Hiyoshi, Y.; Ishimoto, T.; Iwatsuki, M.; Miyamoto, Y.; Yoshida, N.; Watanabe, M.; et al. Tumour-associated macrophages are associated with poor prognosis and programmed death ligand 1 expression in oesophageal cancer. Eur. J. Cancer 2019, 111, 38–49. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Producing Cells | Target Cells | Cancer Species | Function | |
|---|---|---|---|---|
| POSTN | CAFs | TAMs | Melanoma | Immunomodulation |
| T cells | Mycosis fungoides | Angiogenesis-promoting | ||
| CAFs | Merkel cell carcinoma | Inflammation | ||
| Dendritic cells | fibrosis | |||
| PAI-1 | Tumor cells | Tumor cells | Melanoma | Immunomodulation |
| Endothelial cells | Endothelial cells | BCC | Angiogenesis-promoting | |
| macrophages | TAMs | cSCC | Thrombus formation | |
| adipocytes | TILs | |||
| RANKL | Tumor cells | TAMs | Extramammary Paget’s disease | Immunomodulation |
| T cells | Langerhans cells | Melanoma | Osteoclast | |
| Epithelial cells | Dendritic cells | Apocrine carcinoma | Angiogenesis-promoting | |
| PD-L1 | Tumor cells | effector T cells | Melanoma | |
| T cells | TAMs | cSCC | ||
| macrophages | BCC | Immunosuppression | ||
| dendritic cells | Merkel cell carcinoma | Immunomodulation | ||
| Other non-melanoma skin cancer | ||||
| Cutaneous angiosarcoma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujimura, T. Stromal Factors as a Target for Immunotherapy in Melanoma and Non-Melanoma Skin Cancers. Int. J. Mol. Sci. 2022, 23, 4044. https://doi.org/10.3390/ijms23074044
Fujimura T. Stromal Factors as a Target for Immunotherapy in Melanoma and Non-Melanoma Skin Cancers. International Journal of Molecular Sciences. 2022; 23(7):4044. https://doi.org/10.3390/ijms23074044
Chicago/Turabian StyleFujimura, Taku. 2022. "Stromal Factors as a Target for Immunotherapy in Melanoma and Non-Melanoma Skin Cancers" International Journal of Molecular Sciences 23, no. 7: 4044. https://doi.org/10.3390/ijms23074044