
External Hemin as an Inhibitor of Mitochondrial Large-Conductance Calcium-Activated Potassium Channel Activity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
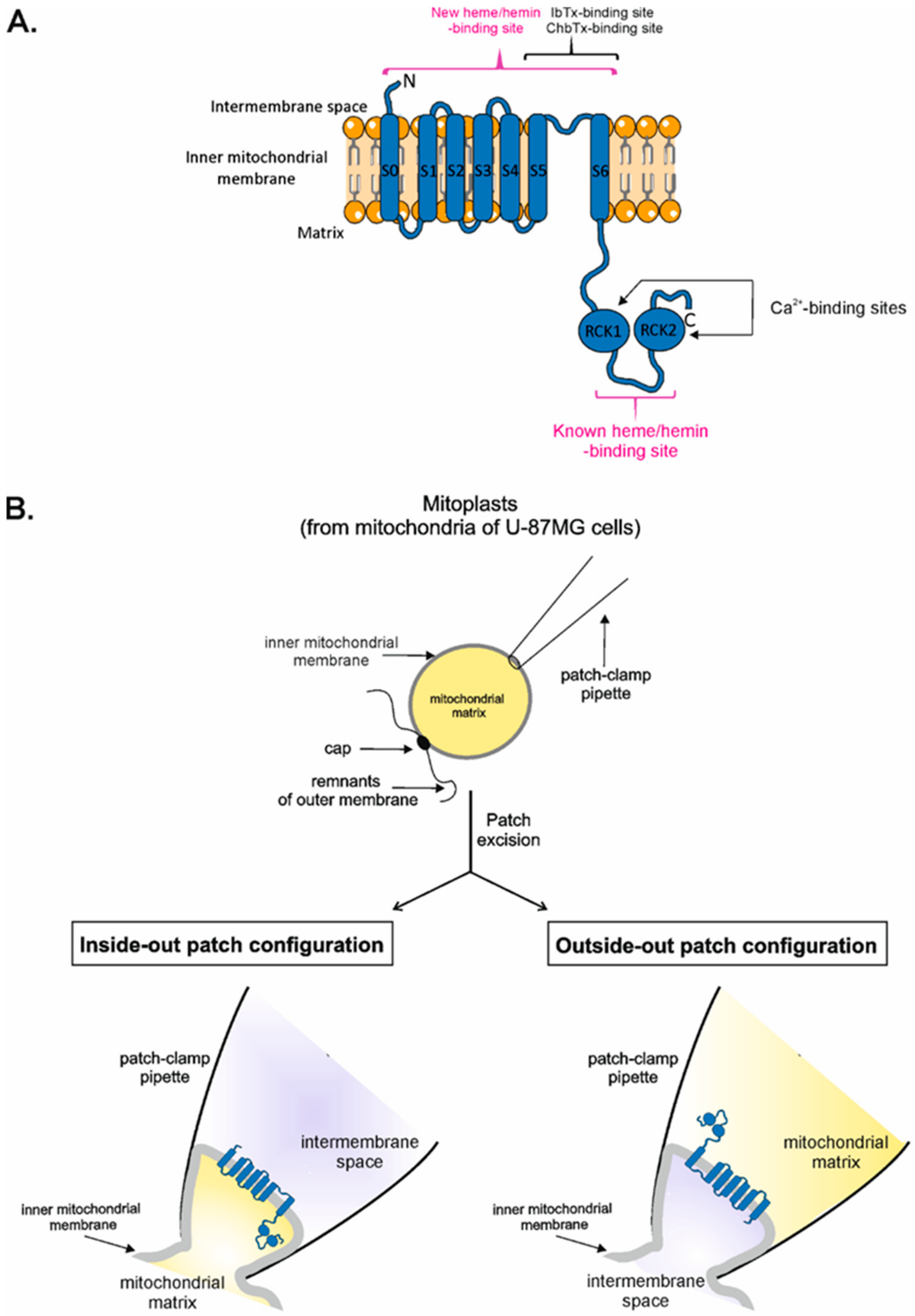
2.1. Definitions
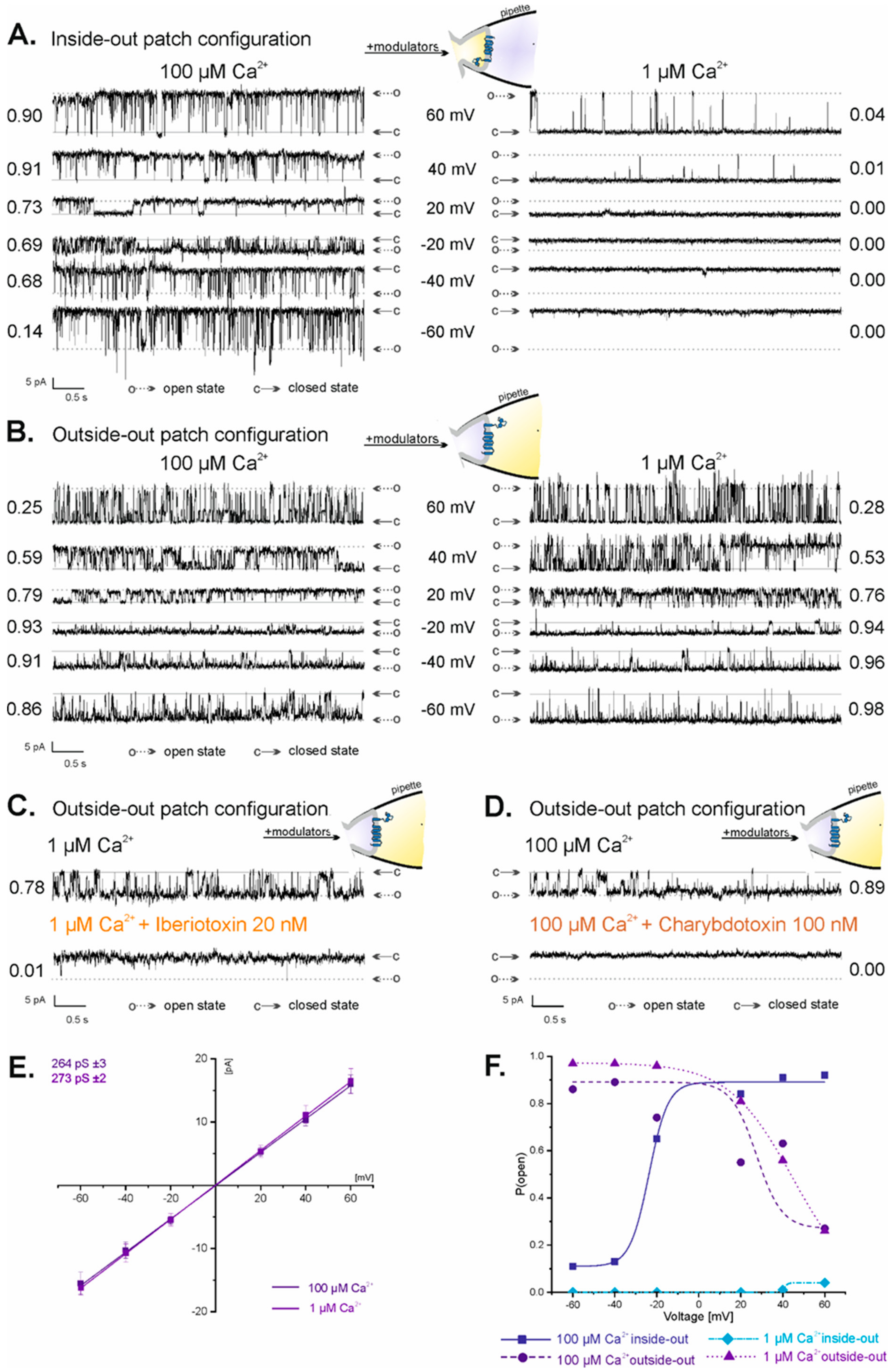
2.2. Vectorial Properties of mitoBKCa Channels
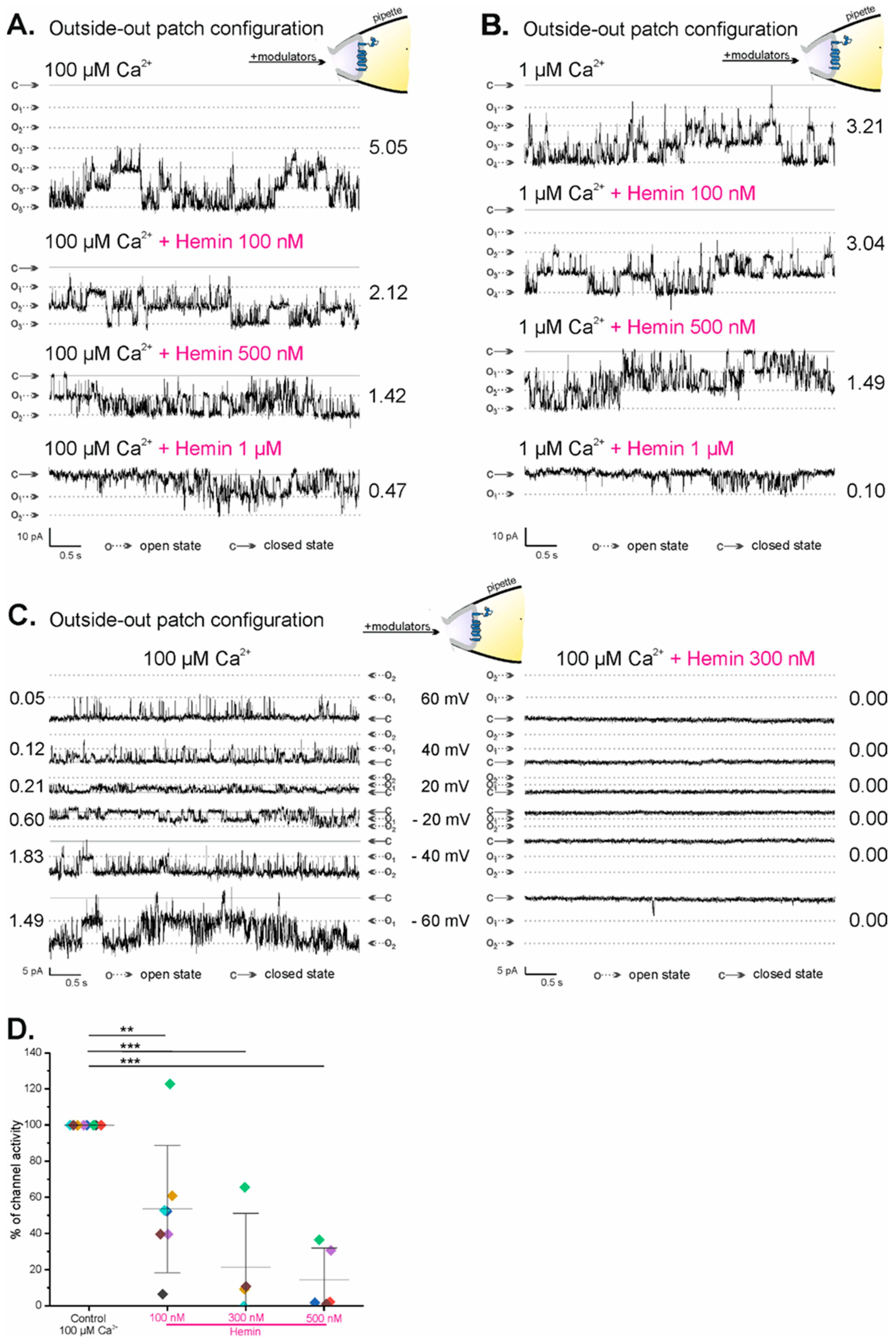
2.3. Regulation of mitoBKCa Channels by External Hemin
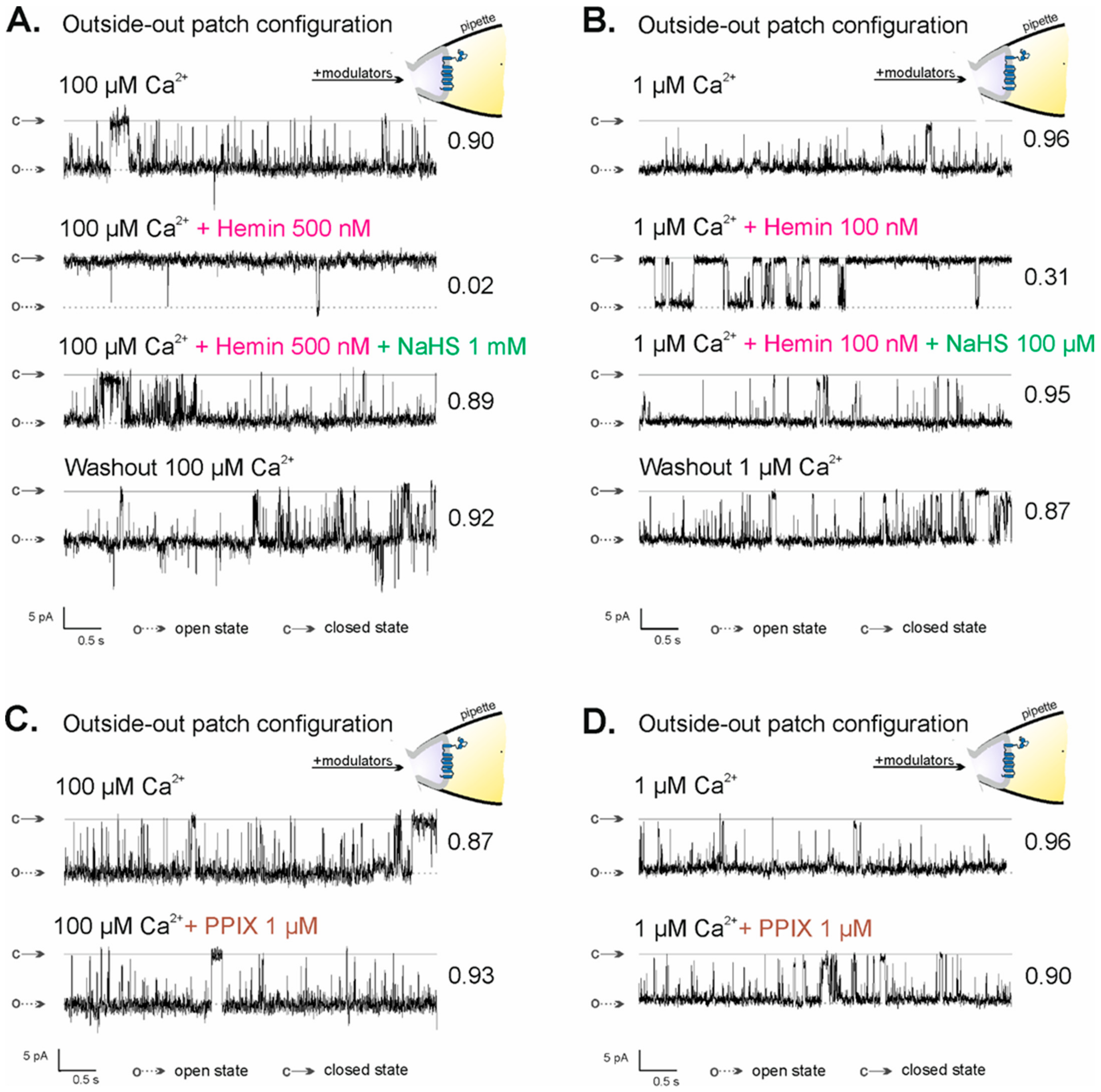
2.4. Regulation of mitoBKCa Channel Activity in the Outside-Out Configuration by NaHS and PPIX
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Mitochondria Isolation
4.3. Mitoplast Preparation
4.4. Patch-Clamp Experiments
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dreyer, I.; Uozumi, N. Potassium Channels in Plant Cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Fischer, W.B.; Sansom, M.S.P. Viral Ion Channels: Structure and Function. Biochim. Biophys. Acta Biomembr. 2002, 1561, 27–45. [Google Scholar] [CrossRef] [Green Version]
- Koprowski, P.; Kubalski, A. Bacterial Ion Channels and Their Eukaryotic Homologues. BioEssays 2001, 23, 1148–1158. [Google Scholar] [CrossRef]
- Koszela-Piotrowska, I.; Matkovic, K.; Szewczyk, A.; Jarmuszkiewicz, W. A Large-Conductance Calcium-Activated Potassium Channel in Potato (Solanum Tuberosum) Tuber Mitochondria. Biochem. J. 2009, 424, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Miller, C. An Overview of the Potassium Channel Family. Genome Biol. 2000, 1, reviews0004.1–reviews0004.5. [Google Scholar] [CrossRef]
- Checchetto, V.; Teardo, E.; Carraretto, L.; Leanza, L.; Szabo, I. Physiology of Intracellular Potassium Channels: A Unifying Role as Mediators of Counterion Fluxes? Biochim. Biophys. Acta Bioenerg. 2016, 1857, 1258–1266. [Google Scholar] [CrossRef]
- Contet, C.; Goulding, S.P.; Kuljis, D.A.; Barth, A.L. BK Channels in the Central Nervous System. Int. Rev. Neurobiol. 2016, 128, 281–342. [Google Scholar] [CrossRef] [Green Version]
- Pluznick, J.L.; Sansom, S.C. BK Channels in the Kidney: Role in K+ Secretion and Localization of Molecular Components. Am. J. Physiol. Renal Physiol. 2006, 291, F517–F529. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Han, B.; Zhang, M.; Wang, Y.H.; Tao, K.; Zhu, M.X.; He, K.; Zhang, Z.G.; Hou, S. Activation of BK Channels Prevents Hepatic Stellate Cell Activation and Liver Fibrosis Through the Suppression of TGFβ1/SMAD3 and JAK/STAT3 Profibrotic Signaling Pathways. Front. Pharmacol. 2020, 11, 165. [Google Scholar] [CrossRef]
- Yun, J.; Park, H.; Ko, J.H.; Lee, W.; Kim, K.; Kim, T.; Shin, J.; Kim, K.; Kim, K.; Song, J.H.; et al. Expression of Ca2+-Activated K+ Channels in Human Dermal Fibroblasts and Their Roles in Apoptosis. Skin Pharmacol. Physiol. 2010, 23, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Brenner, R.; Peréz, G.J.; Bonev, A.D.; Eckman, D.M.; Kosek, J.C.; Wiler, S.W.; Patterson, A.J.; Nelson, M.T.; Aldrich, R.W. Vasoregulation by the Β1 Subunit of the Calcium-Activated Potassium Channel. Nature 2000, 407, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Meredith, A.L.; Thorneloe, K.S.; Werner, M.E.; Nelson, M.T.; Aldrich, R.W. Overactive Bladder and Incontinence in the Absence of the BK Large Conductance Ca2+-Activated K+ Channel. J. Biol. Chem. 2004, 279, 36746–36752. [Google Scholar] [CrossRef] [PubMed]
- Samengo, I.; Currò, D.; Barrese, V.; Taglialatela, M.; Martire, M. Large Conductance Calcium-Activated Potassium Channels: Their Expression and Modulation of Glutamate Release from Nerve Terminals Isolated from Rat Trigeminal Caudal Nucleus and Cerebral Cortex. Neurochem. Res. 2014, 39, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Griguoli, M.; Sgritta, M.; Cherubini, E. Presynaptic BK Channels Control Transmitter Release: Physiological Relevance and Potential Therapeutic Implications. J. Physiol. 2016, 594, 3489–3500. [Google Scholar] [CrossRef] [Green Version]
- Farajnia, S.; Meijer, J.H.; Michel, S. Age-Related Changes in Large-Conductance Calcium-Activated Potassium Channels in Mammalian Circadian Clock Neurons. Neurobiol. Aging 2015, 36, 2176–2183. [Google Scholar] [CrossRef] [Green Version]
- Kshatri, A.S.; Gonzalez-Hernandez, A.; Giraldez, T. Physiological Roles and Therapeutic Potential of Ca2+ Activated Potassium Channels in the Nervous System. Front. Mol. Neurosci. 2018, 11, 258. [Google Scholar] [CrossRef] [Green Version]
- Lee, U.S.; Cui, J. BK Channel Activation: Structural and Functional Insights. Trends Neurosci. 2010, 33, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhang, G.; Cui, J. BK Channels: Multiple Sensors, One Activation Gate. Front. Physiol. 2015, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Horrigan, F.T.; Aldrich, R.W. Coupling between Voltage Sensor Activation, Ca2+ Binding and Channel Opening in Large Conductance (BK) Potassium Channels. J. Gen. Physiol. 2002, 120, 267–305. [Google Scholar] [CrossRef] [Green Version]
- Wawrzkiewicz-Jałowiecka, A.; Trybek, P.; Machura, Ł.; Dworakowska, B.; Grzywna, Z.J. Mechanosensitivity of the BK Channels in Human Glioblastoma Cells: Kinetics and Dynamical Complexity. J. Membr. Biol. 2018, 251, 667–679. [Google Scholar] [CrossRef]
- Mallouk, N.; Allard, B. Stretch-Induced Activation of Ca2+-Activated K+ Channels in Mouse Skeletal Muscle Fibers. Am. J. Physiol. Cell Physiol. 2000, 278, C473–C479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirber, M.T.; Ordway, R.W.; Clapp, L.H.; Walsh, J.V.; Singer, J.J. Both Membrane Stretch and Fatty Acids Directly Activate Large Conductance Ca2+-Activated K+ Channels in Vascular Smooth Muscle Cells. FEBS Lett. 1992, 297, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Pallotta, B.S.; Magleby, K.L.; Barrett, J.N. Single Channel Recordings of Ca2+-Activated K+ Currents in Rat Muscle Cell Culture. Nature 1981, 293, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.R.; Constanti, A.; Brown, D.A.; Clark, R.B. Intracellular Ca2+ Activates a Fast Voltage-Sensitive K+ Current in Vertebrate Sympathetic Neurones. Nature 1982, 296, 746–749. [Google Scholar] [CrossRef]
- Barrett, J.N.; Magleby, K.L.; Pallotta, B.S. Properties of Single Calcium-activated Potassium Channels in Cultured Rat Muscle. J. Physiol. 1982, 331, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Latorre, R.; Castillo, K.; Carrasquel-Ursulaez, W.; Sepulveda, R.V.; Gonzalez-Nilo, F.; Gonzalez, C.; Alvarez, O. Molecular Determinants of BK Channel Functional Diversity and Functioning. Physiol. Rev. 2017, 97, 39–87. [Google Scholar] [CrossRef]
- Yusifov, T.; Savalli, N.; Gandhi, C.S.; Ottolia, M.; Olcese, R. The RCK2 Domain of the Human BKCa Channel Is a Calcium Sensor. Proc. Natl. Acad. Sci. USA 2008, 105, 376–381. [Google Scholar] [CrossRef] [Green Version]
- Xia, X.M.; Zeng, X.; Lingle, C.J. Multiple Regulatory Sites in Large-Conductance Calcium-Activated Potassium Channels. Nature 2002, 418, 880–884. [Google Scholar] [CrossRef]
- Schreiber, M.; Salkoff, L. A Novel Calcium-Sensing Domain in the BK Channel. Biophys. J. 1997, 73, 1355–1363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Huang, S.Y.; Yang, J.; Shi, J.; Yang, X.; Moller, A.; Zou, X.; Cui, J. Ion Sensing in the RCK1 Domain of BK Channels. Proc. Natl. Acad. Sci. USA 2010, 107, 18700–18705. [Google Scholar] [CrossRef]
- Tang, Q.Y.; Zhang, Z.; Meng, X.Y.; Cui, M.; Logothetis, D.E. Structural Determinants of Phosphatidylinositol 4,5-Bisphosphate (PIP2) Regulation of BK Channel Activity through the RCK1 Ca2+ Coordination Site. J. Biol. Chem. 2014, 289, 18860–18872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Aursnes, M.; Hansen, T.V.; Tungen, J.E.; Galpin, J.D.; Leisle, L.; Ahern, C.A.; Xu, R.; Heinemann, S.H.; Hoshi, T. Atomic Determinants of BK Channel Activation by Polyunsaturated Fatty Acids. Proc. Natl. Acad. Sci. USA 2016, 113, 13905–13910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.T.; Lovell, P.V.; Rishniw, M.; Kotlikoff, M.I.; Zeeman, M.L.; McCobb, D.P. Β2 and Β4 Subunits of BK Channels Confer Differential Sensitivity to Acute Modulation by Steroid Hormones. J. Neurophysiol. 2006, 95, 2878–2888. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, H.; Ohi, Y.; Muraki, K.; Watanabe, M.; Imaizumi, Y. BK Channel Activation by NS-1619 Is Partially Mediated by Intracellular Ca2+ Release in Smooth Muscle Cells of Porcine Coronary Artery. Br. J. Pharmacol. 2001, 132, 828–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Layne, J.J.; Nausch, B.; Olesen, S.P.; Nelson, M.T. BK Channel Activation by NS11021 Decreases Excitability and Contractility of Urinary Bladder Smooth Muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R378–R384. [Google Scholar] [CrossRef] [Green Version]
- Augustynek, B.; Koprowski, P.; Rotko, D.; Kunz, W.S.; Szewczyk, A.; Kulawiak, B. Mitochondrial BK Channel Openers CGS7181 and CGS7184 Exhibit Cytotoxic Properties. Int. J. Mol. Sci. 2018, 19, 353. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lingle, C.J. Paxilline Inhibits BK Channels by an Almost Exclusively Closed-Channel Block Mechanism. J. Gen. Physiol. 2014, 144, 415–440. [Google Scholar] [CrossRef] [Green Version]
- Galvez, A.; Gimenez-Gallego, G.; Reuben, J.P.; Roy-Contancin, L.; Feigenbaum, P.; Kaczorowski, G.J.; Garcia, M.L. Purification and Characterization of a Unique, Potent, Peptidyl Probe for the High Conductance Calcium-Activated Potassium Channel from Venom of the Scorpion Buthus Tamulus. J. Biol. Chem. 1990, 265, 11083–11090. [Google Scholar] [CrossRef]
- Miller, C.; Moczydlowski, E.; Latorre, R.; Phillips, M. Charybdotoxin, a Protein Inhibitor of Single Ca2+-Activated K+ Channels from Mammalian Skeletal Muscle. Nature 1985, 313, 316–318. [Google Scholar] [CrossRef]
- Giangiacomo, K.M.; Garcia, M.L.; McManus, O.B. Mechanism of Iberiotoxin Block of the Large-Conductance Calcium-Activated Potassium Channel from Bovine Aortic Smooth Muscle. Biochemistry 1992, 31, 6719–6727. [Google Scholar] [CrossRef]
- Banerjee, A.; Lee, A.; Campbell, E.; MacKinnon, R. Structure of a Pore-Blocking Toxin in Complex with a Eukaryotic Voltage-Dependent K+ Channel. eLife 2013, 2, e00594. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.D.; Xu, R.; Reynolds, M.F.; Garcia, M.L.; Heinemann, S.H.; Hoshi, T. Haem Can Bind to and Inhibit Mammalian Calcium-Dependent Slo1 BK Channels. Nature 2003, 425, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Jaggar, J.H.; Li, A.; Parfenova, H.; Liu, J.; Umstot, E.S.; Dopico, A.M.; Leffler, C.W. Heme Is a Carbon Monoxide Receptor for Large-Conductance Ca2+-Activated K+ Channels. Circ. Res. 2005, 97, 805–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, J.W.A.; Leach, N.; Ferguson, S.J. The Histidine of the C-Type Cytochrome CXXCH Haem-Binding Motif Is Essential for Haem Attachment by the Escherichia Coli Cytochrome c Maturation (Ccm) Apparatus. Biochem. J. 2005, 389 Pt 2, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Khalimonchuk, O.; Smith, P.M.; Winge, D.R. Structure, Function, and Assembly of Heme Centers in Mitochondrial Respiratory Complexes. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1604–1616. [Google Scholar] [CrossRef] [Green Version]
- Kranz, R.G.; Richard-Fogal, C.; Taylor, J.-S.; Frawley, E.R. Cytochrome c Biogenesis: Mechanisms for Covalent Modifications and Trafficking of Heme and for Heme-Iron Redox Control. Microbiol. Mol. Biol. Rev. 2009, 73, 510–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, J.M.; Daltrop, O.; Allen, J.W.A.; Ferguson, S.J. C-Type Cytochrome Formation: Chemical and Biological Enigmas. Acc. Chem. Res. 2004, 37, 999–1007. [Google Scholar] [CrossRef]
- Yi, L.; Morgan, J.T.; Ragsdale, S.W. Identification of a Thiol/Disulfide Redox Switch in the Human BK Channel That Controls Its Affinity for Heme and CO. J. Biol. Chem. 2010, 285, 20117–20127. [Google Scholar] [CrossRef] [Green Version]
- Bednarczyk, P.; Skalska, J.; Glab, M.; Kulawiak, B.; Dolowy, K.; Szewczyk, A. Large Conductance Potassium Ion Channel from Rat Brain Mitochondria. Biochim. Biophys. Acta-Bioenerg. 2006, 461, 1–12. [Google Scholar]
- Sek, A.; Kampa, R.P.; Kulawiak, B.; Szewczyk, A.; Bednarczyk, P. Identification of the Large-conductance Ca2+-regulated Potassium Channel in Mitochondria of Human Bronchial Epithelial Cells. Molecules 2021, 26, 3233. [Google Scholar] [CrossRef]
- Siemen, D.; Loupatatzis, C.; Borecky, J.; Gulbins, E.; Lang, F. Ca2+-Activated K Channel of the BK-Type in the Inner Mitochondrial Membrane of a Human Glioma Cell Line. Biochem. Biophys. Res. Commun. 1999, 257, 549–554. [Google Scholar] [CrossRef]
- Lorigo, M.; Oliveira, N.; Cairrao, E. Clinical Importance of the Human Umbilical Artery Potassium Channels. Cells 2020, 9, 1956. [Google Scholar] [CrossRef]
- Hofmann, F. A Concise Discussion of the Regulatory Role of cGMP Kinase I in Cardiac Physiology and Pathology. Basic Res. Cardiol. 2018, 113, 31. [Google Scholar] [CrossRef]
- Balderas, E.; Zhang, J.; Stefani, E.; Toro, L. Mitochondrial BKCa Channel. Front. Physiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, A.; Bednarczyk, P.; Jędraszko, J.; Kampa, R.P.; Koprowski, P.; Krajewska, M.; Kucman, S.; Kulawiak, B.; Laskowski, M.; Rotko, D.; et al. Mitochondrial Potassium Channels-an Overview. Postepy Biochem. 2018, 64, 196–212. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Lu, R.; Bopassa, J.C.; Meredith, A.L.; Stefani, E.; Toro, L. MitoBKCa Is Encoded by the Kcnma1 Gene, and a Splicing Sequence Defines Its Mitochondrial Location. Proc. Natl. Acad. Sci. USA 2013, 110, 10836–10841. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Liu, Y.; Wang, S.; McDonald, T.; Van Eyk, J.E.; Sidor, A.; O’Rourke, B. Cytoprotective Role of Ca2+-Activated K+ Channels in the Cardiac Inner Mitochondrial Membrane. Science 2002, 298, 1029–1033. [Google Scholar] [CrossRef]
- Borchert, G.H.; Yang, C.; Kolář, F. Mitochondrial BKCa Channels Contribute to Protection of Cardiomyocytes Isolated from Chronically Hypoxic Rats. Am. J. Physiol. Hear. Circ. Physiol. 2011, 300, H507–H513. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Saito, T.; Saegusa, N.; Nakaya, H. Mitochondrial Ca2+-Activated K+ Channels in Cardiac Myocytes: A Mechanism of the Cardioprotective Effect and Modulation by Protein Kinase A. Circulation 2005, 111, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Bentzen, B.H.; Osadchii, O.; Jespersen, T.; Hansen, R.S.; Olesen, S.P.; Grunnet, M. Activation of Big Conductance Ca2+-Activated K+ Channels (BK) Protects the Heart against Ischemia-Reperfusion Injury. Pflugers Arch. Eur. J. Physiol. 2009, 457, 979–988. [Google Scholar] [CrossRef]
- Gałecka, S.; Kulawiak, B.; Bednarczyk, P.; Singh, H.; Szewczyk, A. Single Channel Properties of Mitochondrial Large Conductance Potassium Channel Formed by BK-VEDEC Splice Variant. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Kicinska, A.; Kampa, R.P.; Daniluk, J.; Sek, A.; Jarmuszkiewicz, W.; Szewczyk, A.; Bednarczyk, P. Regulation of the Mitochondrial BKCa Channel by the Citrus Flavonoid Naringenin as a Potential Means of Preventing Cell Damage. Molecules 2020, 25, 3010. [Google Scholar] [CrossRef]
- Kampa, R.P.; Kicinska, A.; Jarmuszkiewicz, W.; Pasikowska-Piwko, M.; Dolegowska, B.; Debowska, R.; Szewczyk, A.; Bednarczyk, P. Naringenin as an Opener of Mitochondrial Potassium Channels in Dermal Fibroblasts. Exp. Dermatol. 2019, 28, 543–550. [Google Scholar] [CrossRef]
- Bednarczyk, P.; Koziel, A.; Jarmuszkiewicz, W.; Szewczyk, A. Large-Conductance Ca2+-Activated Potassium Channel in Mitochondria of Endothelial EA.Hy926 Cells. Am. J. Physiol. Hear. Circ. Physiol. 2013, 304, H1415–H1427. [Google Scholar] [CrossRef]
- Gu, X.Q.; Pamenter, M.E.; Siemen, D.; Sun, X.; Haddad, G.G. Mitochondrial but Not Plasmalemmal BK Channels Are Hypoxia-Sensitive in Human Glioma. Glia 2014, 62, 504–513. [Google Scholar] [CrossRef]
- Skalska, J.; Bednarczyk, P.; Piwońska, M.; Kulawiak, B.; Wilczynski, G.; Dołowy, K.; Kudin, A.P.; Kunz, W.S.; Szewczyk, A. Calcium Ions Regulate K+ Uptake into Brain Mitochondria: The Evidence for a Novel Potassium Channel. Int. J. Mol. Sci. 2009, 10, 1104–1120. [Google Scholar] [CrossRef] [Green Version]
- Augustynek, B.; Kudin, A.P.; Bednarczyk, P.; Szewczyk, A.; Kunz, W.S. Hemin Inhibits the Large Conductance Potassium Channel in Brain Mitochondria: A Putative Novel Mechanism of Neurodegeneration. Exp. Neurol. 2014, 257, 70–75. [Google Scholar] [CrossRef]
- Robinson, S.R.; Dang, T.N.; Dringen, R.; Bishop, G.M. Hemin Toxicity: A Preventable Source of Brain Damage Following Hemorrhagic Stroke. Redox Rep. 2009, 14, 228–235. [Google Scholar] [CrossRef]
- Walewska, A.; Szewczyk, A.; Krajewska, M.; Koprowski, P. Targeting Mitochondrial Large-Conductance Calcium-Activated Potassium Channel by Hydrogen Sulfide via Heme-Binding Site. J. Pharmacol. Exp. Ther. 2022, 381, 137–150. [Google Scholar] [CrossRef]
- Walewska, A.; Kulawiak, B.; Szewczyk, A.; Koprowski, P. Mechanosensitivity of Mitochondrial Large-Conductance Calcium-Activated Potassium Channels. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 797–805. [Google Scholar] [CrossRef]
- Lavi, A.; Weitman, H.; Holmes, R.T.; Smith, K.M.; Ehrenberg, B. The Depth of Porphyrin in a Membrane and the Membrane’s Physical Properties Affect the Photosensitizing Efficiency. Biophys J. 2002, 82, 2101–2110. [Google Scholar] [CrossRef]
- Schmitt, T.H.; Frezzatti, W.A.; Schreier, S. Hemin-induced lipid membrane disorder and increased permeability: A molecular model for the mechanism of cell lysis. Arch Biochem Biophys. 1993, 307, 96–103. [Google Scholar] [CrossRef]
- Burton, M.J.; Kapetanaki, S.M.; Chernova, T.; Jamieson, A.G.; Dorlet, P.; Santolini, J.; Moody, P.C.E.; Mitcheson, J.S.; Davies, N.W.; Schmid, R.; et al. A Heme-Binding Domain Controls Regulation of ATP-Dependent Potassium Channels. Proc. Natl. Acad. Sci. USA 2016, 113, 3785–3790. [Google Scholar] [CrossRef] [Green Version]
- Borchert, G.H.; Hlaváčková, M.; Kolář, F. Pharmacological Activation of Mitochondrial BKCa Channels Protects Isolated Cardiomyocytes against Simulated Reperfusion-Induced Injury. Exp. Biol. Med. 2013, 238, 233–241. [Google Scholar] [CrossRef]
- Soltysinska, E.; Bentzen, B.H.; Barthmes, M.; Hattel, H.; Thrush, A.B.; Harper, M.E.; Qvortrup, K.; Larsen, F.J.; Schiffer, T.A.; Losa-Reyna, J.; et al. KCNMA1 Encoded Cardiac BK Channels Afford Protection against Ischemia-Reperfusion Injury. PLoS ONE 2014, 9, e103402. [Google Scholar] [CrossRef] [Green Version]
- Frankenreiter, S.; Bednarczyk, P.; Kniess, A.; Bork, N.I.; Straubinger, J.; Koprowski, P.; Wrzosek, A.; Mohr, E.; Logan, A.; Murphy, M.P.; et al. CGMP-Elevating Compounds and Ischemic Conditioning Provide Cardioprotection Against Ischemia and Reperfusion Injury via Cardiomyocyte-Specific BK Channels. Circulation 2017, 136, 2337–2355. [Google Scholar] [CrossRef] [Green Version]
- Kampa, R.P.; Sęk, A.; Szewczyk, A.; Bednarczyk, P. Cytoprotective Effects of the Flavonoid Quercetin by Activating Mitochondrial BKCa Channels in Endothelial Cells. Biomed. Pharmacother. 2021, 142, 112039. [Google Scholar] [CrossRef]
- Debska-Vielhaber, G.; Godlewski, M.M.; Kicinska, A.; Skalska, J.; Kulawiak, B.; Piwonska, M.; Zablocki, K.; Kunz, W.S.; Szewczyk, A.; Motyl, T. Large-Conductance K+ Channel Openers Induce Death of Human Glioma Cells. J. Physiol. Pharmacol. 2009, 60, 27–36. [Google Scholar]
- Bury, M.; Girault, A.; Megalizzi, V.; Spiegl-Kreinecker, S.; Mathieu, V.; Berger, W.; Evidente, A.; Kornienko, A.; Gailly, P.; Vandier, C.; et al. Ophiobolin A Induces Paraptosis-like Cell Death in Human Glioblastoma Cells by Decreasing BKCa Channel Activity. Cell Death Dis. 2013, 4, e561. [Google Scholar] [CrossRef] [Green Version]
- Leanza, L.; Venturini, E.; Kadow, S.; Carpinteiro, A.; Gulbins, E.; Becker, K.A. Targeting a Mitochondrial Potassium Channel to Fight Cancer. Cell Calcium. 2015, 58, 131–138. [Google Scholar] [CrossRef]
- Nilsson, R.; Schultz, I.J.; Pierce, E.L.; Soltis, K.A.; Naranuntarat, A.; Ward, D.M.; Baughman, J.M.; Paradkar, P.N.; Kingsley, P.D.; Culotta, V.C.; et al. Discovery of Genes Essential for Heme Biosynthesis through Large-Scale Gene Expression Analysis. Cell Metab. 2009, 10, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawicki, K.T.; Chang, H.C.; Ardehali, H. Role of Heme in Cardiovascular Physiology and Disease. J. Am. Heart Assoc. 2015, 4, e001138. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Lengalova, A.; Martínek, V.; Martínková, M. Heme: Emergent Roles of Heme in Signal Transduction, Functional Regulation and as Catalytic Centres. Chem. Soc. Rev. 2019, 48, 5624–5657. [Google Scholar] [CrossRef]
- Tsiftsoglou, A.S.; Tsamadou, A.I.; Papadopoulou, L.C. Heme as Key Regulator of Major Mammalian Cellular Functions: Molecular, Cellular, and Pharmacological Aspects. Pharmacol. Ther. 2006, 111, 327–345. [Google Scholar] [CrossRef]
- Hoshi, T.; Zagotta, W.N.; Aldrich, R.W. Biophysical and Molecular Mechanisms of Shaker Potassium Channel Inactivation. Science 1990, 250, 533–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoshi, T.; Zagotta, W.N.; Aldrich, R.W. Two Types of Inactivation in Shaker K+ Channels: Effects of Alterations in the Carboxy-Terminal Region. Neuron 1991, 7, 547–556. [Google Scholar] [CrossRef]
- Sahoo, N.; Goradia, N.; Ohlenschläger, O.; Schönherr, R.; Friedrich, M.; Plass, W.; Kappl, R.; Hoshi, T.; Heinemann, S.H. Heme Impairs the Ball-and-Chain Inactivation of Potassium Channels. Proc. Natl. Acad. Sci. USA 2013, 110, E4036–E4044. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Bai, J.; Yang, Y.H.; Hoshi, N.; Chen, D.B. Hydrogen Sulfide Relaxes Human Uterine Artery via Activating Smooth Muscle BKCa Channels. Antioxidants 2020, 9, 1127. [Google Scholar] [CrossRef]
- Sitdikova, G.F.; Weiger, T.M.; Hermann, A. Hydrogen Sulfide Increases Calcium-Activated Potassium (BK) Channel Activity of Rat Pituitary Tumor Cells. Pflugers Arch. Eur. J. Physiol. 2010, 459, 389–397. [Google Scholar] [CrossRef]
- Quan, X.; Luo, H.; Liu, Y.; Xia, H.; Chen, W.; Tang, Q. Hydrogen Sulfide Regulates the Colonic Motility by Inhibiting Both L-Type Calcium Channels and BKCa Channels in Smooth Muscle Cells of Rat Colon. PLoS ONE 2015, 10, e0121331. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, H.; Kong, G.; Shim, W.; Zhang, G. Hydrogen Sulfide Augments the Proliferation and Survival of Human Induced Pluripotent Stem Cell-Derived Mesenchymal Stromal Cells through Inhibition of BKCa. Cytotherapy 2013, 15, 1395–1405. [Google Scholar] [CrossRef] [PubMed]
- Bednarczyk, P.; Wieckowski, M.R.; Broszkiewicz, M.; Skowronek, K.; Siemen, D.; Szewczyk, A. Putative Structural and Functional Coupling of the Mitochondrial BKCa Channel to the Respiratory Chain. PLoS ONE 2013, 8, e68125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednarczyk, P.; Kampa, R.P.; Gałecka, S.; Se, A.; Walewska, A.; Koprowski, P. Patch-Clamp Recording of the Activity of Ion Channels in the Inner Mitochondrial Membrane. Methods Mol. Biol. 2021, 2276, 235–248. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walewska, A.; Szewczyk, A.; Koprowski, P. External Hemin as an Inhibitor of Mitochondrial Large-Conductance Calcium-Activated Potassium Channel Activity. Int. J. Mol. Sci. 2022, 23, 13391. https://doi.org/10.3390/ijms232113391
Walewska A, Szewczyk A, Koprowski P. External Hemin as an Inhibitor of Mitochondrial Large-Conductance Calcium-Activated Potassium Channel Activity. International Journal of Molecular Sciences. 2022; 23(21):13391. https://doi.org/10.3390/ijms232113391
Chicago/Turabian StyleWalewska, Agnieszka, Adam Szewczyk, and Piotr Koprowski. 2022. "External Hemin as an Inhibitor of Mitochondrial Large-Conductance Calcium-Activated Potassium Channel Activity" International Journal of Molecular Sciences 23, no. 21: 13391. https://doi.org/10.3390/ijms232113391