
METCAM/MUC18 Plays a Tumor Suppressor Role in the Development of Nasopharyngeal Carcinoma Type I



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
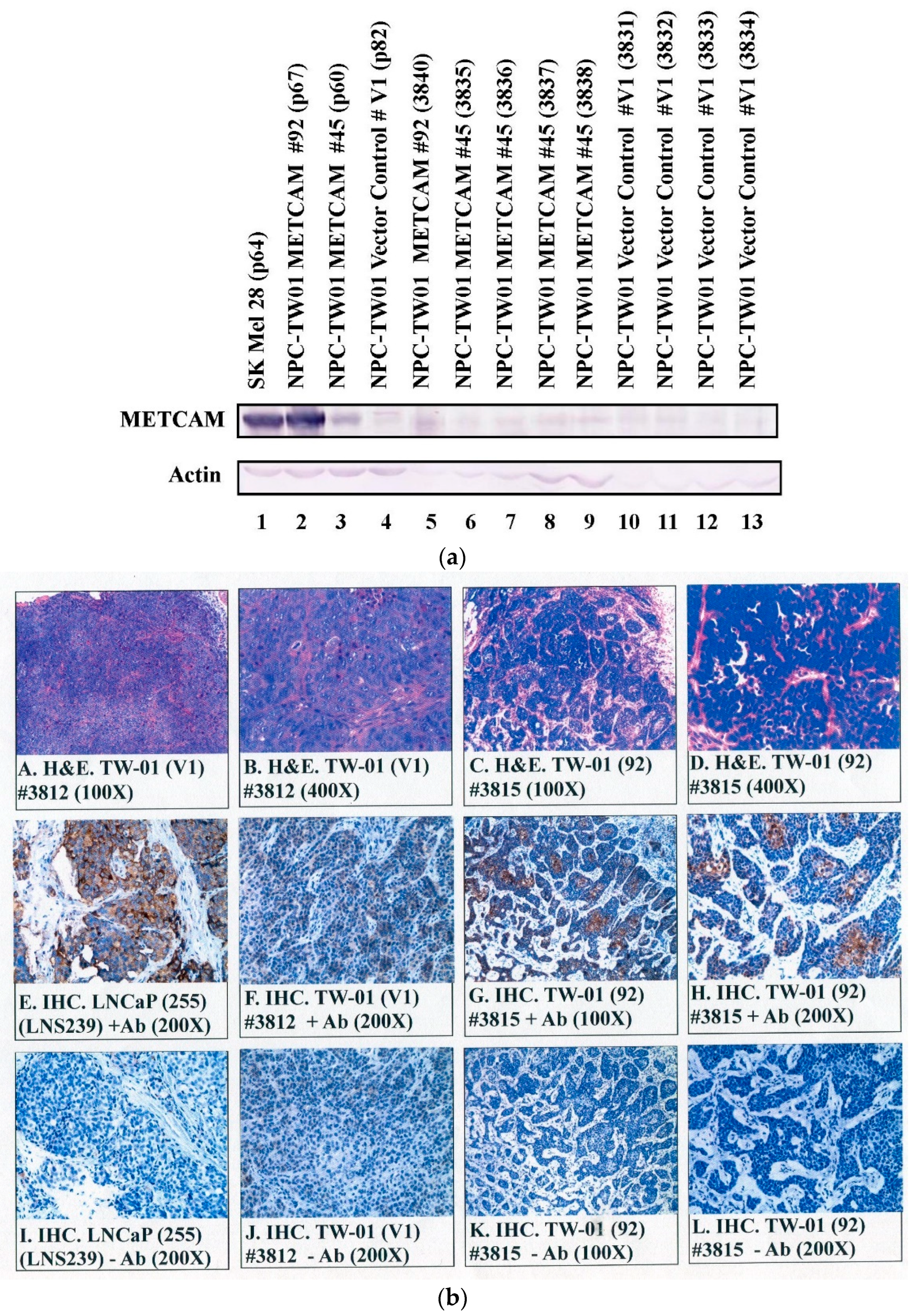
2.1. Expression of huMETCAM/MUC18 Protein in the Clones of the NPC-TW01 Cell Line
2.2. Over-Expression of huMETCAM/MUC18 Did Not Affect the In Vitro Growth Rate of the NPC-TW01 Cells
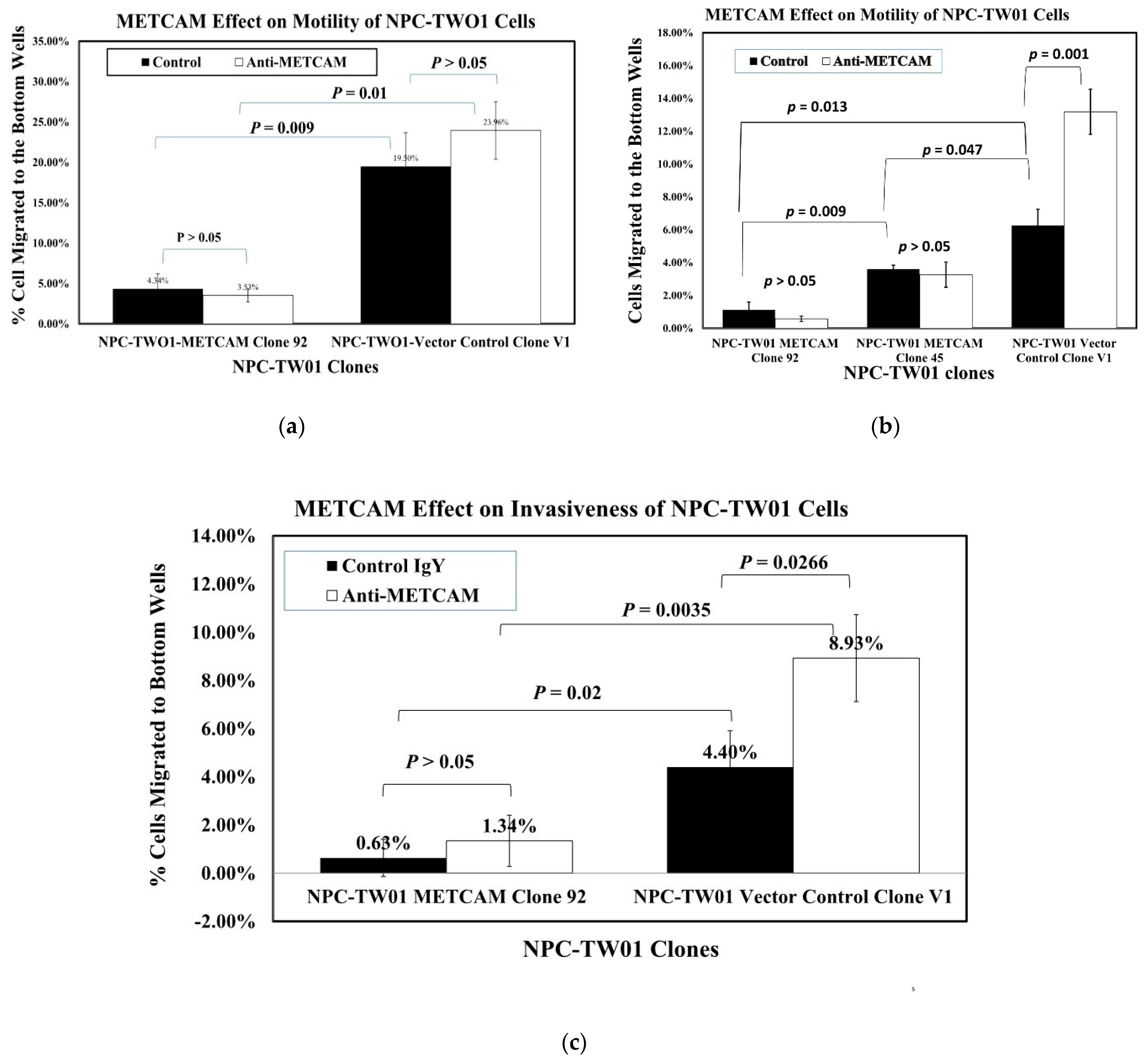
2.3. Overexpression of huMETCAM/MUC18 Decreases the Migration and Invasiveness of the NPC-TW01 Cells
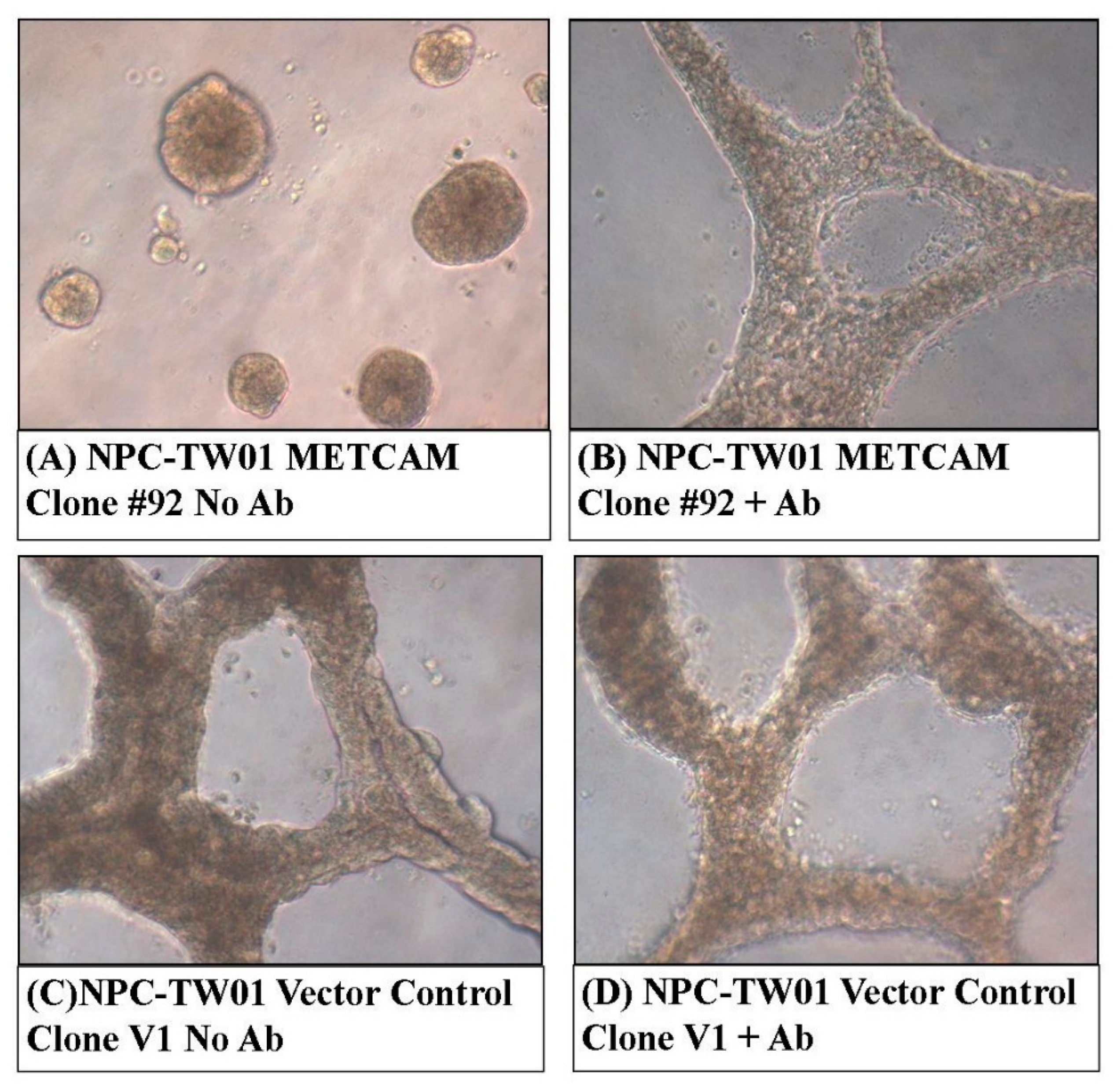
2.4. Over-Expression of huMETCAM/MUC18 Decreased Invasive Growth in the 3D Basement Membrane Assay
2.5. Overexpression of huMETCAM/MUC18 Decreases Tumorigenesis in Athymic Nude Mice
2.6. Expression of huMETCAM/MUC18 in Tumor Tissues
2.7. Preliminary Mechanisms of huMETCAM/MUC18-Induced Tumor Suppression in the NPC-TW01 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Antibodies
4.2. Growth of NPC-TW01 and Other Cell Lines
4.3. Lipofection of NPC-TW01 Cells and Selection for huMETCAM/MUC18-Expressing G418R-Clones
4.4. Determination of In Vitro (Intrinsic) Growth Rate of NPC-TW01 Clones
4.5. Cell Motility Assay
4.6. Cell Invasiveness Assay
4.7. Three-Dimensional (3D) Basement Membrane (Matrigel) Culture Assay
4.8. Determination of the Tumorigenicity of the NPC-TW01 Clones/Cells in Athymic Nude Mice
4.9. Western Blot (Immunoblot) Analysis
4.10. Histology and Immunohistochemistry
4.11. Determination of Vascular Density in Tumor Sections
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ho, J.H.C. Current knowledge of the epidemiology of nasopharyngeal carcinoma (NPC). In Oncogenesis and Herpesviruses; Biggs, P., de The’, G., Payne, L., Eds.; IARC: Lyon, France, 1972; pp. 357–366. [Google Scholar]
- Shanmugaratnam, K.; Sobin, L.H. The World Health Organization histological classification of tumors of the upper respiratory tract and ear. A commentary on the second edition. Cancer 1993, 71, 2689–2697. [Google Scholar] [CrossRef]
- Wei, W.; Sham, J.S. Nasopharyngeal carcinoma. Lancet 2005, 365, 2041–2054. [Google Scholar] [CrossRef]
- Gkretsi, V.; Stylianopoulos, T. Cell Adhesion and Matrix Stiffness: Coordinating Cancer Cell Invasion and Metastasis. Front. Oncol. 2018, 8, 145. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Dong, W.; Zhou, X.; Li, S. The significance of soluble intercellular adhesion molecule 1 (ICAM-1) and transformation growth factor alpha in patients with nasopharyngeal carcinomas. Acta Otolaryngol. Head Neck Surg. 2004, 130, 1205–1208. [Google Scholar]
- Huang, G.W.; Mo, W.N.; Kuang, G.Q.; Nong, H.T.; Wei, M.Y.; Sunagawa, M.; Kosugi, T. Expression of p16, nm-23-H1, E-cadherin, and CD44 gene products and their significance in nasopharyngeal carcinomas. Laryngoscope 2001, 111, 1465–1471. [Google Scholar] [CrossRef]
- Li, Z.; Ren, Y.; Lin, S.-X.; Liang, Y.-J.; Liang, H.-Z. Association of E-cadherin and beta-catenin with metastasis in nasopharyngeal carcinoma. Chin. Med. J. 2004, 117, 1232–1239. [Google Scholar]
- Yi, Z.-C.; Wang, H.; Zhang, G.-Y.; Xia, B. Downregulation of connexin 43 in nasopharyngeal carcinoma cells is related to promoter methylation. Oral Oncol. 2006, 43, 898–904. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.M.; Riethmüller, G.; Johnson, J.P. MUC18, a marker of tumor progression in human melanoma, shows sequence similarity to the neural cell adhesion molecules of the immunoglobulin superfamily. Proc. Natl. Acad. Sci. USA 1989, 86, 9891–9895. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.J. METCAM/MUC18 Expression and Cancer Metastasis. Curr. Genom. 2005, 6, 333–349. [Google Scholar] [CrossRef]
- Lin, J.C.; Chiang, C.F.; Wang, S.W.; Wang, W.Y.; Kwuan, P.C.; Wu, G.J. Significance and expression of human METCAM/MUC18 in nasopharyngeal carcinoma (NPC) and metastatic lesions. Asian Pac. J. Cancer Prev. 2014, 15, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Shih, I.-M. The role of CD146 (Mel-CAM) in biology and pathology. J. Pathol. 1999, 189, 4–11. [Google Scholar] [CrossRef]
- Zeng, G.-F.; Cai, S.-X.; Wu, G.-J. Up-regulation of METCAM/MUC18 promotes motility, invasion, and tumorigenesis of human breast cancer cells. BMC Cancer 2011, 11, 113. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-J.; Dickerson, E.B. Frequent and increased expression of human METCAM/MUC18 in cancer tissues and metastatic lesions is associated with the clinical progression of human ovarian carcinoma. Taiwan. J. Obstet. Gynecol. 2014, 53, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.-J. Dual Roles of METCAM in the Progression of Different Cancers. J. Oncol. 2012, 2012, 853797. [Google Scholar] [CrossRef] [Green Version]
- Zeng, G.; Cai, S.; Liu, Y.; Wu, G.-J. METCAM/MUC18 augments migration, invasion, and tumorigenicity of human breast cancer SK-BR-3 cells. Gene 2012, 492, 229–238. [Google Scholar] [CrossRef]
- Zeng, Q.; Li, W.; Lu, D.; Wu, Z.; Duan, H.; Luo, Y.; Feng, J.; Yang, D.; Fu, L.; Yan, X. CD146, an epithelial-mesenchymal transition inducer, is associated with triple-negative breast cancer. Proc. Natl. Acad. Sci. USA 2011, 109, 1127–1132. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-Y.; Wu, G.-J. Mechanism of METCAM/MUC18-promoted progression of human breast cancer cells: METCAM/MUC18 promoted tumorigenesis of human breast cancer SK-BR-3 cells in a dosage-specific manner. Taiwan. J. Obstet. Gynecol. 2016, 55, 202–212. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.-J.; Varma, V.A.; Wu, M.-W.H.; Wang, S.-W.; Qu, P.; Yang, H.; Petros, J.A.; Lim, S.D.; Amin, M.B. Expression of a human cell adhesion molecule, MUC18, in prostate cancer cell lines and tissues. Prostate 2001, 48, 305–315. [Google Scholar] [CrossRef]
- Wu, G.-J.; Wu, M.-W.H.; Wang, S.-W.; Liu, Z.; Qu, P.; Peng, Q.; Yang, H.; Varma, V.A.; Sun, Q.C.; Petros, J.A.; et al. Isolation and characterization of the major form of human MUC18 cDNA gene and correlation of MUC18 over-expression in prostate cancer cell lines and tissues with malignant progression. Gene 2001, 279, 17–31. [Google Scholar] [CrossRef]
- Wu, G.-J.; Peng, Q.; Fu, P.; Wang, S.-W.; Chiang, C.-F.; Dillehay, D.L.; Wu, M.-W.H. Ectopical expression of human MUC18 increases metastasis of human prostate cancer cells. Gene 2004, 327, 201–213. [Google Scholar] [CrossRef]
- Wu, G.-J.; Fu, P.; Chiang, C.-F.; Huss, W.J.; Greenberg, N.M.; Wu, M.-W.H. Increased expression of MUC18 correlates with the metastatic progression of mouse prostate adenocarcinoma in the TRAMP model. J. Urol. 2005, 173, 1778–1783. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-J.; Wu, M.-W.H.; Wang, C.; Liu, Y. Enforced Expression of METCAM/MUC18 Increases Tumorigenesis of Human Prostate Cancer LNCaP Cells in Nude Mice. J. Urol. 2011, 185, 1504–1512. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.J. Human METCAM/MUC18 as a Novel Biomarker to Drive and its Specific SiRNAs to Block the Malignant Progression of Prostate Cancer. J. Cell Sci. Ther. 2015, 6, 1000227. [Google Scholar] [CrossRef]
- Wu, G.J. Human METCAM/MUC18 is a New Diagnostic Marker of and a Driver for Promoting and its Specific SiRNAs, Derived Oligopeptides and Antibodies be used for Decreasing the Malignant Progression of Prostate Cancer. J. Stem Cell Res. Ther. 2016, 1, 35. [Google Scholar] [CrossRef]
- Xie, S.; Luca, M.; Huang, S.; Gutman, M.; Reich, R.; Johnson, J.P.; Bar-Eli, M. Expression of MCAM/MCU18 by human melanoma cells leads to increased tumor growth and metastasis. Cancer Res. 1997, 57, 2295–2303. [Google Scholar]
- Schlagbauer-Wadl, H.; Jansen, B.; Polterauer, P.; Wolff, K.; Eichler, H.-G.; Pehamberger, H.; Konakand, E.; Johnson, J.P. Influence of MUC18/MCAM/CD146 expression on human melanoma growth and metastasis in SCID mice. Int. J. Cancer 1999, 81, 951–955. [Google Scholar] [CrossRef]
- Wu, G.-J.; Fu, P.; Wang, S.-W.; Wu, M.-W.H. Enforced Expression of MCAM/MUC18 Increases In vitro Motility and Invasiveness and In vivo Metastasis of Two Mouse Melanoma K1735 Sublines in a Syngeneic Mouse Model. Mol. Cancer Res. 2008, 6, 1666–1677. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.-J.; Zeng, G.-F. METCAM/MUC18 is a novel tumor and metastasis suppressor for the human ovarian cancer SKOV3 cells. BMC Cancer 2016, 16, 136. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.-J. METCAM/MUC18 plays a Novel Tumor and Metastasis Suppressor Role in the Progression of Human Ovarian Cancer Cells. Obstet. Gynecol. Int. J. 2017, 6, 210. [Google Scholar] [CrossRef]
- Wu, G.-J. METCAM/MUC18 Decreases the Malignant Propensity of Human Ovarian Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 2976. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.-J. Ectopic expression of MCAM/MUC18 increases in vitro motility and invasiveness, but decreases in vivo tumorigenesis and metastasis of a mouse melanoma K1735-9 subline in a syngeneic mouse model. Clin. Exp. Metastasis 2016, 33, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C. The putative role of huMETCAM in modulating the development and progression of nasopharyngeal carcinoma. Master’s Thesis, Chung Yuan Christian University, Taoyuan, Taiwan, 2014. Available online: http://.www.lib.cycu.edu.tw/thesis (accessed on 1 January 2019).
- Liu, Y.C.; Ko, C.C.; Chen, Y.J.; Wu, G.J. METCAM/MUC18 plays a tumor promoter role in the development of nasopharyngeal carcinoma type III. 2022; in preparation. [Google Scholar]
- Leone, A.; Flatow, U.; King, C.R.; Sandeen, M.A.; Margulies, I.M.; Liotta, L.A.; Steeg, P.S. Reduced tumor incidence, metastatic potential, and cytokine responsiveness of nm-23 transfected melanoma cells. Cell 1991, 65, 25. [Google Scholar] [CrossRef]
- Lee, G.Y.; Kenny, P.A.; Lee, E.H.; Bissell, M.J. Three-dimensional culture models of normal and malignant breast epithelial cells. Nat. Methods 2007, 4, 359–365. [Google Scholar] [CrossRef]
- Lin, C.T.; Wong, C.I.; Chan, W.Y.; Tzung, K.W.; Ho, J.K.; Hsu, M.M.; Chuang, S.M. Establishment and characterization of two nasopharyngeal carcinoma cell lines. Lab. Investig. 1990, 62, 713–724. [Google Scholar]
- Lin, C.T.; Chan, W.Y.; Chen, W.; Huang, H.M.; Wu, H.C.; Hsu, M.M.; Chuang, S.M.; Wang, C.C. Characterization of seven newly established nasopharyngeal carcinoma cell lines. Lab. Investig. 1993, 68, 716–727. [Google Scholar]
- Cheung, A.K.L.; Ip, J.C.Y.; Chu, A.C.H.; Cheng, Y.; Leong, M.M.L.; Ko, J.M.Y.; Shuen, W.H.; Lung, H.L.; Lung, M.L. PTPRG suppresses tumor growth and invasion via inhibition of Akt signaling in nasopharyngeal carcinoma. Oncotarget 2015, 6, 13434–13447. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Du, L.; Chen, D.; Ye, Z.; Duan, H.; Tu, T.; Feng, J.; Yang, Y.; Chen, Q.; Yan, X. Reduced CD146 expression promotes tumorigenesis and cancer stemness in colorectal cancer through activating Wnt/β-catenin signaling. Oncotarget 2016, 7, 40704–40718. [Google Scholar] [CrossRef]
- Wu, G.J. Dual role of METCAM/MUC18 in the progression of human cancer cells. In Gene Expression and Regulation in Mammalian Cells; Uchiumi, F., Ed.; Chapter 13; InTech-Open Access Publisher: Croatia, Balkans, 2018; pp. 257–288. [Google Scholar]
- Aguirre-Chiso, J.A. Models, mechanisms and clinical evidence for cancer dormancy. Nat. Rev. Cancer 2007, 7, 834–846. [Google Scholar] [CrossRef] [Green Version]
- Pickl, W.F.; Majdic, O.; Fischer, G.F.; Petzelbauer, P.; Faé, I.; Waclavicek, M.; Stöckl, J.; Scheinecker, C.; Vidicki, T.; Aschauer, H.; et al. MUC18/MCAM (CD146), an activation antigen of human T lymphocytes. J. Immunol. 1997, 158, 2107–2115. [Google Scholar]
- Elshal, M.F.; Khan, S.S.; Takahashi, Y.; Solomon, M.A.; McCoy, J.P. CD146 (Mel-CAM), an adhesion marker of endothelial cells, is a novel marker of lymphocyte subset activation in normal peripheral blood. Blood 2005, 106, 2923–2924. [Google Scholar] [CrossRef] [PubMed]
- Despoix, N.; Walzer, T.; Jouve, N.; Blot-Chabaud, M.; Bardin, N.; Paul, P.; Lyonnet, L.; Vivier, E.; Dignat-George, F.; Vély, F. Mouse CD146/MCAM is a marker of natural killer cell maturation. Eur. J Immunol. 2008, 38, 2855–2864. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Johansson, M.; Coussens, L.M. Immune cells as mediators of solid tumor metastasis. Cancer Metastasis Rev. 2008, 27, 11–18. [Google Scholar] [CrossRef]
- Yang, H.; Wu, M.W.H.; Chern, S.W.W.; Liu, Z.; Wu, G.J. Isolation and characterization of murine MUC18 cDNA gene, and correlation of MUC18 expression in murine melanoma cell lines with metastatic ability. Gene 2001, 265, 133–145. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology; Green Publishing Associates and Wiley-Interscience Press: New York, NY, USA, 1987; Section 4.9. [Google Scholar]
- Passaniti, A.; Isaacs, J.T.; Haney, J.A.; Adler, S.W.; Cujdik, T.J.; Long, P.V.; Kleinman, H.K. Stimulation of human prostatic carcinoma tumor growth in athymic mice and control of migration in culture by extracellular matrix. Int. J. Cancer 1992, 51, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Noël, A.; De Pauw-Gillet, M.-C.; Purnell, G.; Nusgens, B.; Lapiere, C.-M.; Foidart, J.-M. Enhancement of tumorigenicity of human breast adenocarcinoma cells in nude mice by matrigel and fibroblasts. Br. J. Cancer 1993, 68, 909–915. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-C.; Chen, Y.-J.; Wu, G.-J. METCAM/MUC18 Plays a Tumor Suppressor Role in the Development of Nasopharyngeal Carcinoma Type I. Int. J. Mol. Sci. 2022, 23, 13389. https://doi.org/10.3390/ijms232113389
Liu Y-C, Chen Y-J, Wu G-J. METCAM/MUC18 Plays a Tumor Suppressor Role in the Development of Nasopharyngeal Carcinoma Type I. International Journal of Molecular Sciences. 2022; 23(21):13389. https://doi.org/10.3390/ijms232113389
Chicago/Turabian StyleLiu, Yen-Chun, Yu-Jen Chen, and Guang-Jer Wu. 2022. "METCAM/MUC18 Plays a Tumor Suppressor Role in the Development of Nasopharyngeal Carcinoma Type I" International Journal of Molecular Sciences 23, no. 21: 13389. https://doi.org/10.3390/ijms232113389