

Endometrial Cells Acutely Exposed to Phthalates In Vitro Do Not Phenocopy Endometriosis
 , ,
, ,
Abstract
:1. Introduction
2. Results
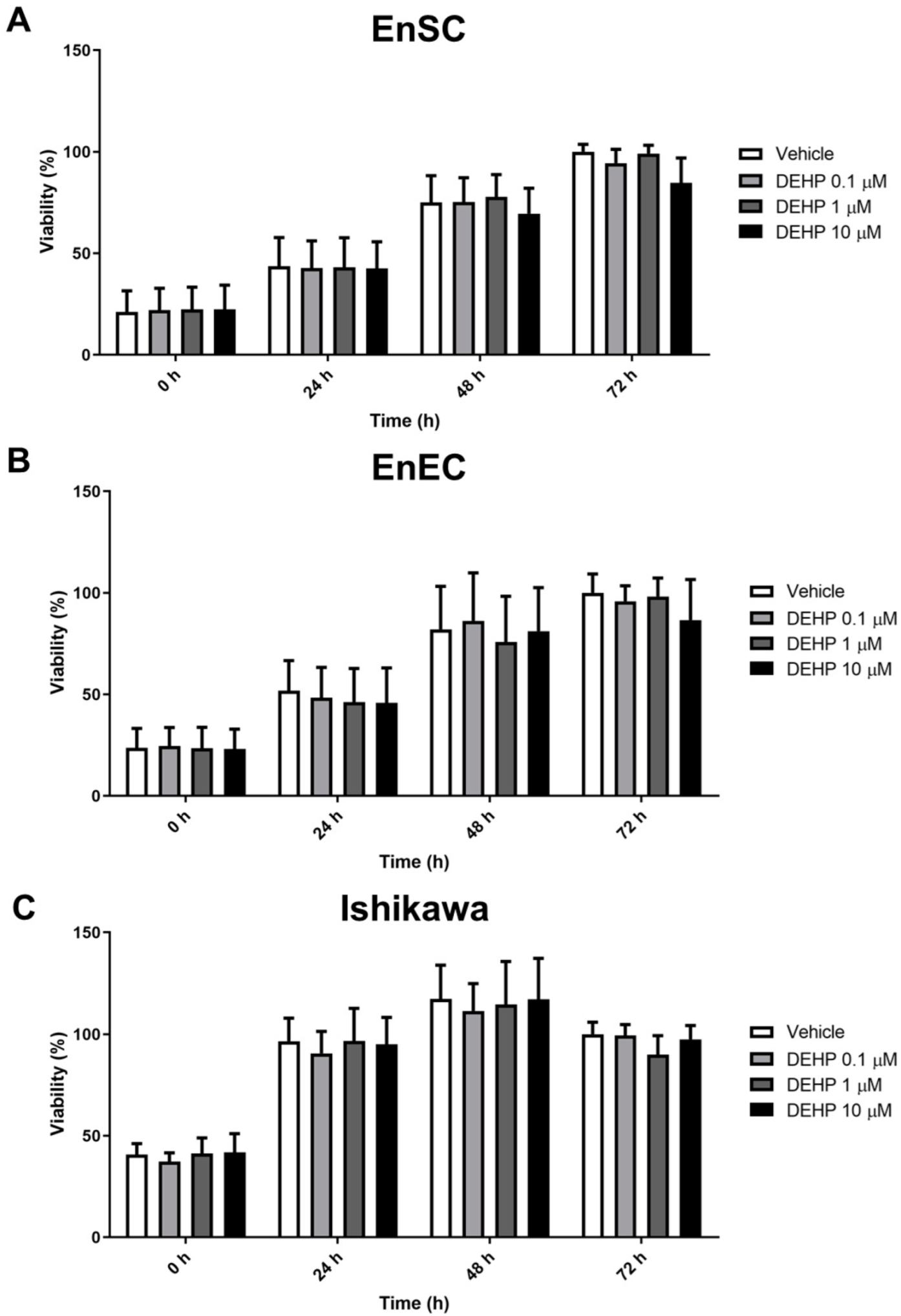
2.1. Acute DEHP Exposure Did Not Affect the Viability of Endometrial Cells
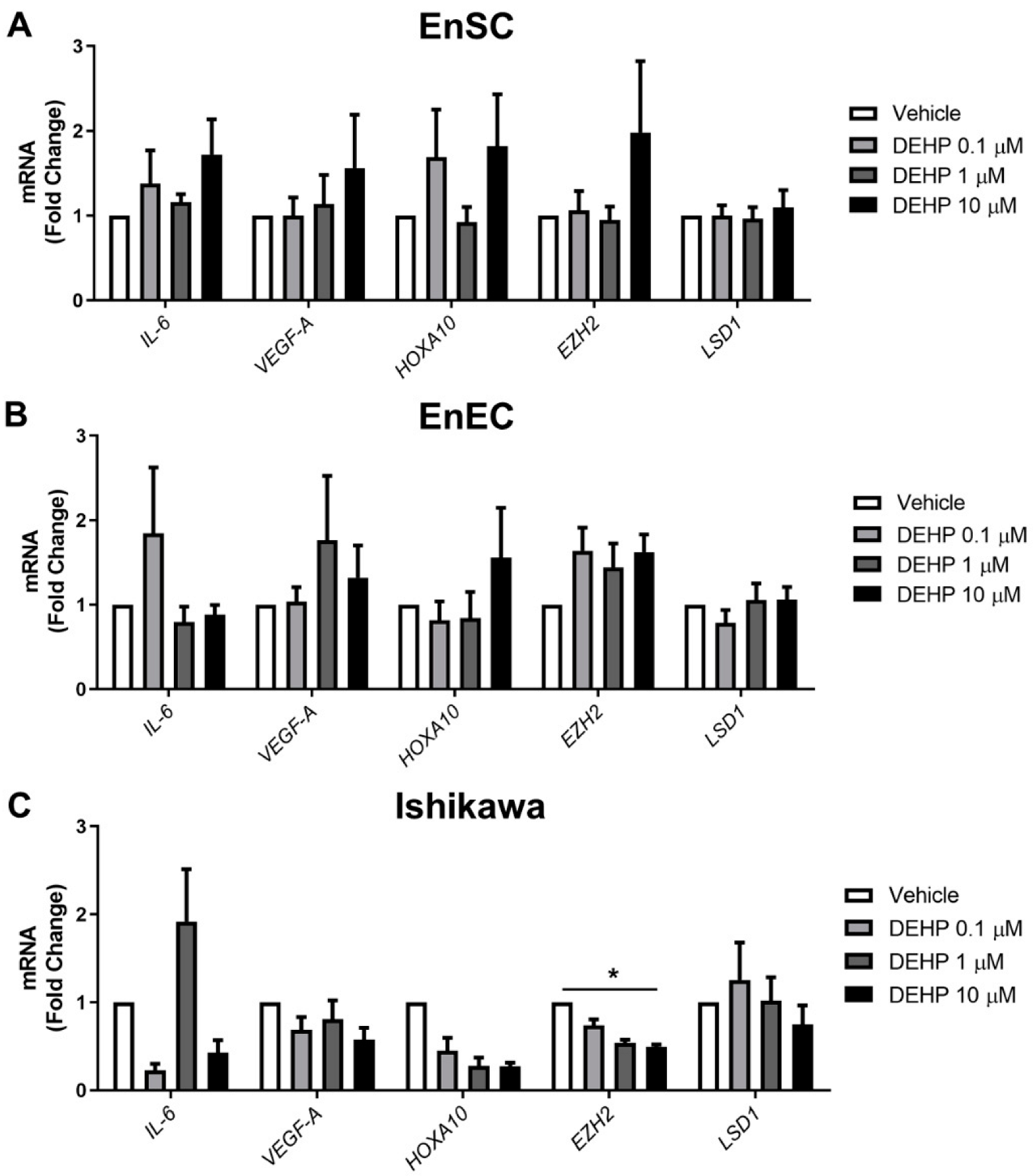
2.2. Endometriosis Marker Expression Was Not Altered after Acute DEHP Exposure
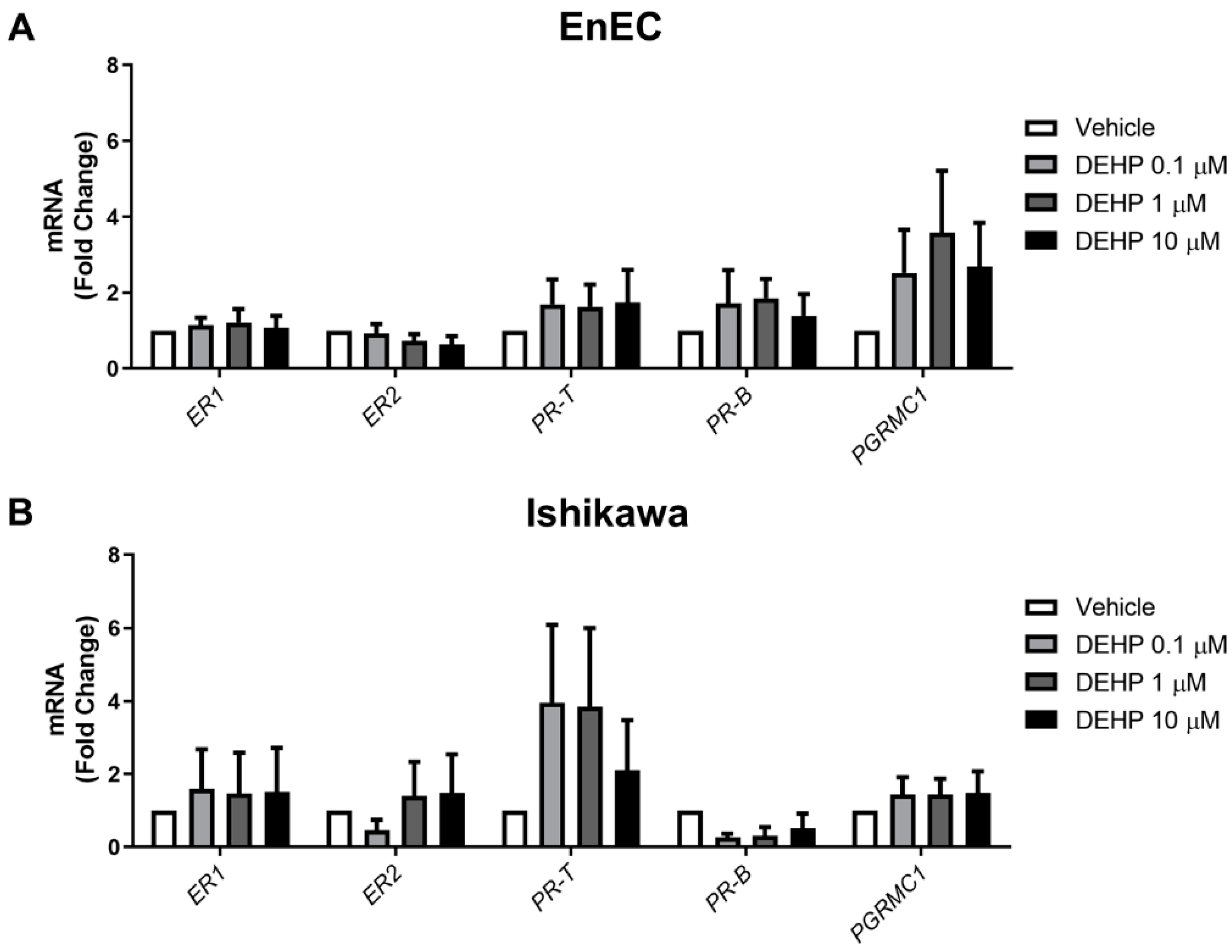
2.3. Acute DEHP Exposure Does Not Alter Steroid Receptor Expression in EnEC or Ishikawa Cells
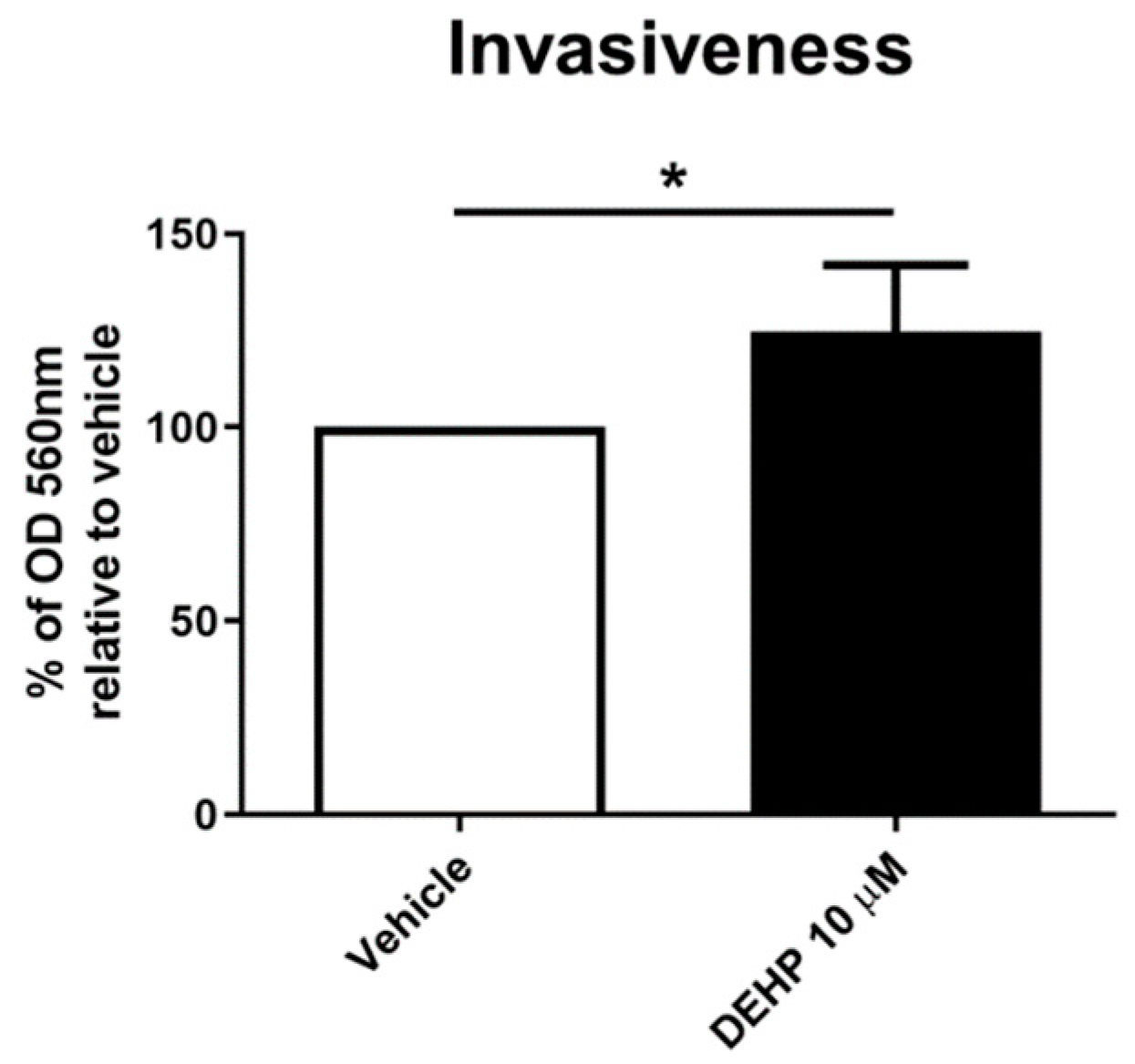
2.4. Acute DEHP Exposure Increases EnSC Invasiveness
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Isolation, Culture, and DEHP Exposure
4.3. Cell Viability Assay (MTS)
4.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and Quantitative PCR (q-PCR)
4.5. Invasion Assay
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bulun, S.E. Chapter 25—Endometriosis. In Yen and Jaffe’s Reproductive Endocrinology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 11, pp. 609–642.e7. [Google Scholar] [CrossRef]
- Klemmt, P.A.; Starzinski-Powitz, A. Molecular and Cellular Pathogenesis of Endometriosis. Curr. Women’s Health Rev. 2017, 13, 106–116. [Google Scholar] [CrossRef]
- Tapia-Orozco, N.; Santiago-Toledo, G.; Barrón, V.; Espinosa-García, A.M.; García-García, J.A.; García-Arrazola, R. Environmental epigenomics: Current approaches to assess epigenetic effects of endocrine disrupting compounds (EDC’s) on human health. Environ. Toxicol. Pharmacol. 2017, 51, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, S.; Masai, E.; Kamimura, N.; Takahashi, K.; Anderson, R.C.; Faisal, P.A. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J. Hazard. Mater. 2017, 340, 360–383. [Google Scholar] [CrossRef] [PubMed]
- Conforti, A.; Carbone, L.; Simeon, V.; Chiodini, P.; Marrone, V.; Bagnulo, F.; Cariati, F.; Strina, I.; Alviggi, C. Unravelling the link between phthalate exposure and endometriosis in humans: A systematic review and meta-analysis of the literature. J. Assist. Reprod. Genet. 2021, 38, 2543–2557. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.A.; Hannon, P.R.; Johnson-Walker, Y.J.; Myint, M.S.; Flaws, J.A.; Nowak, R.A. Di (2-ethylhexyl) phthalate (DEHP) alters proliferation and uterine gland numbers in the uteri of adult exposed mice. Reprod. Toxicol. 2018, 77, 70–79. [Google Scholar] [CrossRef]
- Lovekamp-Swan, T.; Davis, B.J. Mechanisms of phthalate ester toxicity in the female reproductive system. Environ. Health Perspect. 2003, 111, 139–145. [Google Scholar] [CrossRef]
- Rowdhwal, S.S.S.; Chen, J. Toxic Effects of Di-2-ethylhexyl Phthalate: An Overview. Biomed Res. Int. 2018, 2018, 1750368. [Google Scholar] [CrossRef]
- Li, R.; Yu, C.; Gao, R.; Liu, X.; Lu, J.; Zhao, L.; Chen, X.; Ding, Y.; Wang, Y.; He, J. Effects of DEHP on endometrial receptivity and embryo implantation in pregnant mice. J. Hazard. Mater. 2012, 241–242, 231–240. [Google Scholar] [CrossRef]
- Wen, X.; Xiong, Y.; Qu, X.; Jin, L.; Zhou, C.; Zhang, M.; Zhang, Y. The risk of endometriosis after exposure to endocrine-disrupting chemicals: A meta-analysis of 30 epidemiology studies. Gynecol. Endocrinol. 2019, 35, 645–650. [Google Scholar] [CrossRef]
- Nazir, S.; Usman, Z.; Imran, M.; Lone, K.; Ahmad, G. Women diagnosed with endometriosis show high serum levels of diethyl hexyl phthalate. J. Hum. Reprod. Sci. 2018, 11, 131–136. [Google Scholar] [CrossRef]
- Reddy, B.S.; Rozati, R.; Reddy, B.V.R.; Raman, N.V.V.S.S. Association of phthalate esters with endometriosis in Indian women. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Cobellis, L.; Latini, G.; DeFelice, C.; Razzi, S.; Paris, I.; Ruggieri, F.; Mazzeo, P.; Petraglia, F. High plasma concentrations of di-(2-ethylhexyl)-phthalate in women with endometriosis. Hum. Reprod. 2003, 18, 1512–1515. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Chun, S.; Jang, J.Y.; Chae, H.D.; Kim, C.H.; Kang, B.M. Increased plasma levels of phthalate esters in women with advanced-stage endometriosis: A prospective case-control study. Fertil. Steril. 2011, 95, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Upson, K.; Sathyanarayana, S.; De Roos, A.J.; Thompson, M.L.; Scholes, D.; Dills, R.; Holt, V.L. Phthalates and risk of endometriosis. Environ. Res. 2013, 126, 91–97. [Google Scholar] [CrossRef]
- Kim, S.H.; Cho, S.H.; Ihm, H.J.; Oh, Y.S.; Heo, S.H.; Chun, S.; Im, H.; Chae, H.D.; Kim, C.H.; Kang, B.M. Possible role of phthalate in the pathogenesis of endometriosis: In vitro, animal, and human data. J. Clin. Endocrinol. Metab. 2015, 100, E1502–E1511. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, S.H.; Lee, H.W.; Chae, H.D.; Kim, C.H.; Kang, B.M. Increased viability of endometrial cells by in vitro treatment with di-(2-ethylhexyl) phthalate. Fertil. Steril. 2010, 94, 2413–2416. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, H.; Chen, Y.J.; Chi, Y.L.; Dong, S. The inflammation response to DEHP through PPARγ in endometrial cells. Int. J. Environ. Res. Public Health 2016, 13, 318. [Google Scholar] [CrossRef]
- Cho, Y.J.; Park, S.B.; Han, M. Di-(2-ethylhexyl)-phthalate induces oxidative stress in human endometrial stromal cells in vitro. Mol. Cell. Endocrinol. 2015, 407, 9–17. [Google Scholar] [CrossRef]
- Kim, H.G.; Lim, Y.S.; Hwang, S.; Kim, H.Y.; Moon, Y.; Song, Y.J.; Na, Y.J.; Yoon, S. Di-(2-ethylhexyl) Phthalate Triggers Proliferation, Migration, Stemness, and Epithelial–Mesenchymal Transition in Human Endometrial and Endometriotic Epithelial Cells via the Transforming Growth Factor-β/Smad Signaling Pathway. Int. J. Mol. Sci. 2022, 23, 3938. [Google Scholar] [CrossRef]
- Kim, L.Y.; Kim, M.R.; Kim, J.H.; Cho, H.H. Aldo-keto reductase activity after diethylhexyl phthalate exposure in eutopic and ectopic endometrial cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 215, 215–219. [Google Scholar] [CrossRef]
- Reddy, B.S.; Rozati, R.; Reddy, S.; Kodampur, S.; Reddy, P.; Reddy, R. High plasma concentrations of polychlorinated biphenyls and phthalate esters in women with endometriosis: A prospective case control study. Fertil. Steril. 2006, 85, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Huang, Z.; Xiao, Z.; Li, S.; Ma, Q. The cytochrome P4501A1 gene polymorphisms and endometriosis: A meta-analysis. J. Assist. Reprod. Genet. 2016, 33, 1373–1383. [Google Scholar] [CrossRef] [PubMed]
- Ramon, L.A.; Braza-Bols, A.; Gilabert-Estelles, J.; Gilabert, J.; España, F.; Chirivella, M.; Estelles, A. MicroRNAs expression in endometriosis and their relation to angiogenic factors. Hum. Reprod. 2011, 26, 1082–1090. [Google Scholar] [CrossRef]
- Jana, S.K.; Banerjee, P.; Mukherjee, R.; Chakravarty, B.; Chaudhury, K. HOXA-11 mediated dysregulation of matrix remodeling during implantation window in women with endometriosis. J. Assist. Reprod. Genet. 2013, 30, 1505–1512. [Google Scholar] [CrossRef]
- Sun, Q.; Ding, D.; Liu, X.; Guo, S.W. Tranylcypromine, a lysine-specific demethylase 1 (LSD1) inhibitor, suppresses lesion growth and improves generalized hyperalgesia in mouse with induced endometriosis. Reprod. Biol. Endocrinol. 2016, 14, 17. [Google Scholar] [CrossRef]
- Zhang, Q.; Dong, P.; Liu, X.; Sakuragi, N.; Guo, S.W. Enhancer of Zeste homolog 2 (EZH2) induces epithelial-mesenchymal transition in endometriosis. Sci. Rep. 2017, 7, 6804. [Google Scholar] [CrossRef] [PubMed]
- Borghese, B.; Zondervan, K.T.; Abrao, M.S.; Chapron, C.; Vaiman, D. Recent insights on the genetics and epigenetics of endometriosis. Clin. Genet. 2017, 91, 254–264. [Google Scholar] [CrossRef]
- Grimstad, F.W.; Decherney, A. A Review of the Epigenetic Contributions to Endometriosis. Clin. Obstet. Gynecol. 2017, 60, 467–476. [Google Scholar] [CrossRef]
- Alavian-Ghavanini, A.; Rüegg, J. Understanding Epigenetic Effects of Endocrine Disrupting Chemicals: From Mechanisms to Novel Test Methods. Basic Clin. Pharmacol. Toxicol. 2018, 122, 38–45. [Google Scholar] [CrossRef]
- Wu, Y.; Halverson, G.; Basir, Z.; Strawn, E.; Yan, P.; Guo, S.W. Aberrant methylation at HOXA10 may be responsible for its aberrant expression in the endometrium of patients with endometriosis. Am. J. Obstet. Gynecol. 2005, 193, 371–380. [Google Scholar] [CrossRef]
- Bunch, K.; Tinnemore, D.; Huff, S.; Hoffer, Z.S.; Burney, R.O.; Stallings, J.D. Expression patterns of progesterone receptor membrane components 1 and 2 in endometria from women with and without endometriosis. Reprod. Sci. 2014, 21, 190–197. [Google Scholar] [CrossRef]
- Béliard, A.; Noël, A.; Foidart, J.M. Reduction of apoptosis and proliferation in endometriosis. Fertil. Steril. 2004, 82, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Bruner-Tran, K.L.; Eisenberg, E.; Yeaman, G.R.; Anderson, T.A.; McBean, J.; Osteen, K.G. Steroid and cytokine regulation of matrix metalloproteinase expression in endometriosis and the establishment of experimental endometriosis in nude mice. J. Clin. Endocrinol. Metab. 2002, 87, 4782–4791. [Google Scholar] [CrossRef]
- Kyama, C.M.; Overbergh, L.; Mihalyi, A.; Meuleman, C.; Mwenda, J.M.; Mathieu, C.; D’Hooghe, T.M. Endometrial and peritoneal expression of aromatase, cytokines, and adhesion factors in women with endometriosis. Fertil. Steril. 2008, 89, 301–310. [Google Scholar] [CrossRef]
- Igarashi, T.M.; Bruner-Tran, K.L.; Yeaman, G.R.; Lessey, B.A.; Edwards, D.P.; Eisenberg, E.; Osteen, K.G. Reduced expression of progesterone receptor-B in the endometrium of women with endometriosis and in cocultures of endometrial cells exposed to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Fertil. Steril. 2005, 84, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Salsano, S.; Pérez-Debén, S.; Quiñonero, A.; González-Martín, R.; Domínguez, F. Phytoestrogen exposure alters endometrial stromal cells and interferes with decidualization signaling. Fertil. Steril. 2019, 112, 947–958.e3. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Debén, S.; Gonzalez-Martin, R.; Palomar, A.; Quiñonero, A.; Salsano, S.; Dominguez, F. Copper and lead exposures disturb reproductive features of primary endometrial stromal and epithelial cells. Reprod. Toxicol. 2020, 93, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, F.; Avila, S.; Cervero, A.; Martín, J.; Pellicer, A.; Castrillo, J.L.; Simón, C. A Combined Approach for Gene Discovery Identifies Insulin-Like Growth Factor-Binding Protein-Related Protein 1 as a New Gene Implicated in Human Endometrial Receptivity. J. Clin. Endocrinol. Metab. 2003, 88, 1849–1857. [Google Scholar] [CrossRef]
- Simón, C.; Mercader, A.; Garcia-Velasco, J.; Nikas, G.; Moreno, C.; Remohí, J.; Pellicer, A. Coculture of human embryos with autologous human endometrial epithelial cells in patients with implantation failure. J. Clin. Endocrinol. Metab. 1999, 84, 2638–2646. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| IL-6 | GATGGATGCTTCCAATCTGG | TGGCATTTGTGGTTGGGTCA |
| VEGF-A | AGGGCAGAGAATCACGAAG | TGGTGATGTTGGACTGCTCA |
| HOXA10 | TGCTCCCTTCGCCAAATTA | GATGAGCGAGTCGACCAAA |
| EZH2 | TTCATGCAACACCCAACACT | CTCCCTCCAAATGCTGGTAA |
| LSD1 | CTAATGCCACACCTCTCTCAAC | CACACGAGTAGCCATTCCTTAC |
| ER1 | GCTTCGATGATGGGCTTAC | CTGATCATGGAGGGTCAAATC |
| ER2 | GATCGCTAGAACAACACACCTTAC | CGACCAGACTCCATAGTGATA |
| PR-T | GTGGGAGCTGTAAGGTCTTCTTTAA | AACGATGCAGTCATTTCTTCCA |
| PR-B | TCGGACACCTTGCCTGAAGT | CAGGGCCGAGGGAAGAGTAG |
| PGRMC1 | GGAAGAGATGCATCCAGGG | TGAGTACACAGTGGGCTCCT |
| GAPDH | AGATCAAGAAGGTGGTGAAG | TTGTCATACCAGGAAATGAGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Martin, R.; Palomar, A.; Medina-Laver, Y.; Quiñonero, A.; Domínguez, F. Endometrial Cells Acutely Exposed to Phthalates In Vitro Do Not Phenocopy Endometriosis. Int. J. Mol. Sci. 2022, 23, 11041. https://doi.org/10.3390/ijms231911041
Gonzalez-Martin R, Palomar A, Medina-Laver Y, Quiñonero A, Domínguez F. Endometrial Cells Acutely Exposed to Phthalates In Vitro Do Not Phenocopy Endometriosis. International Journal of Molecular Sciences. 2022; 23(19):11041. https://doi.org/10.3390/ijms231911041
Chicago/Turabian StyleGonzalez-Martin, Roberto, Andrea Palomar, Yassmin Medina-Laver, Alicia Quiñonero, and Francisco Domínguez. 2022. "Endometrial Cells Acutely Exposed to Phthalates In Vitro Do Not Phenocopy Endometriosis" International Journal of Molecular Sciences 23, no. 19: 11041. https://doi.org/10.3390/ijms231911041