

Increased Arterial Responsiveness to Angiotensin II in Mice Conceived by Assisted Reproductive Technologies
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
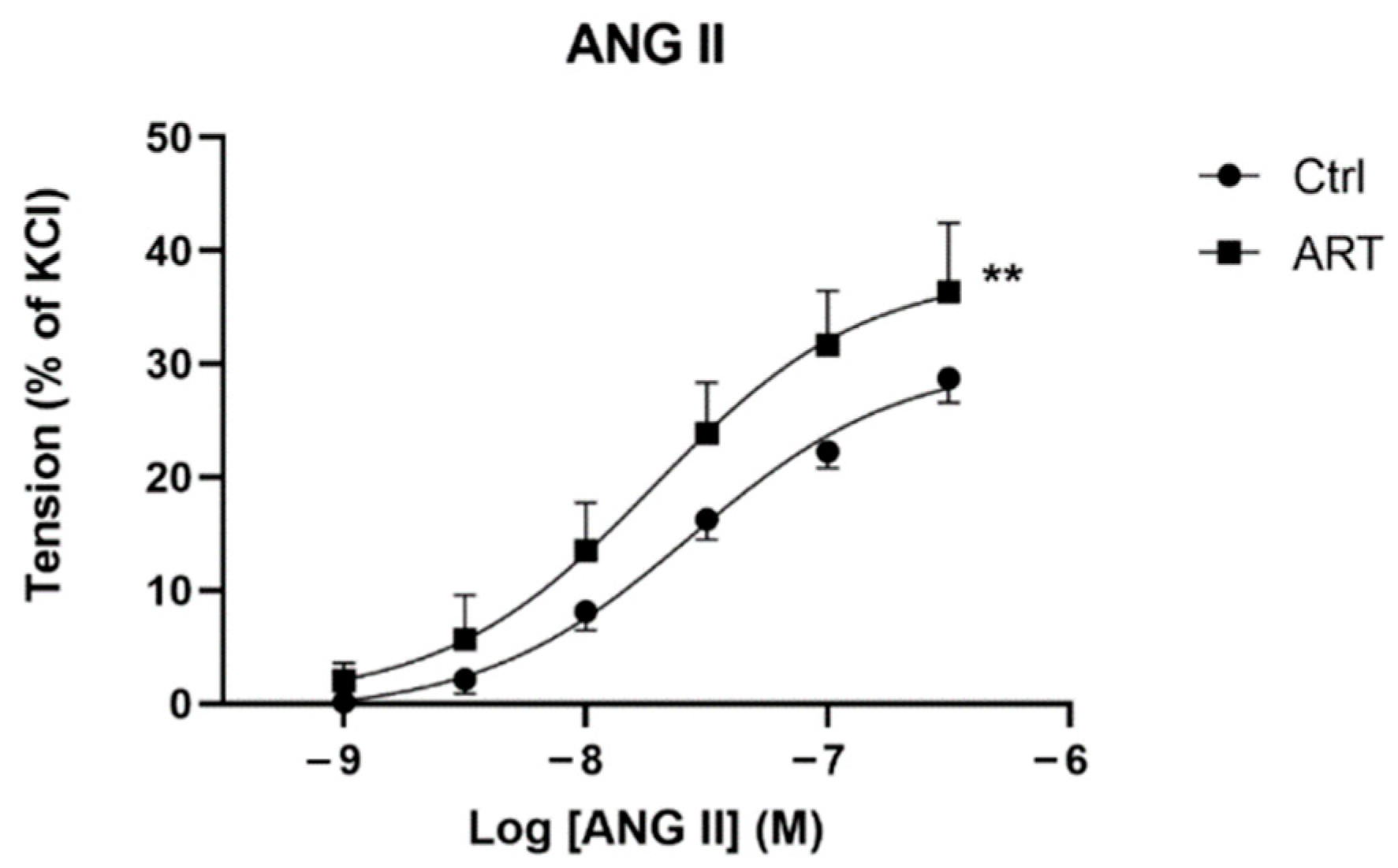
2.1. Increased ANG II Sensitivity Leads to Exaggerated Aortic Vasoconstriction in ART Mice
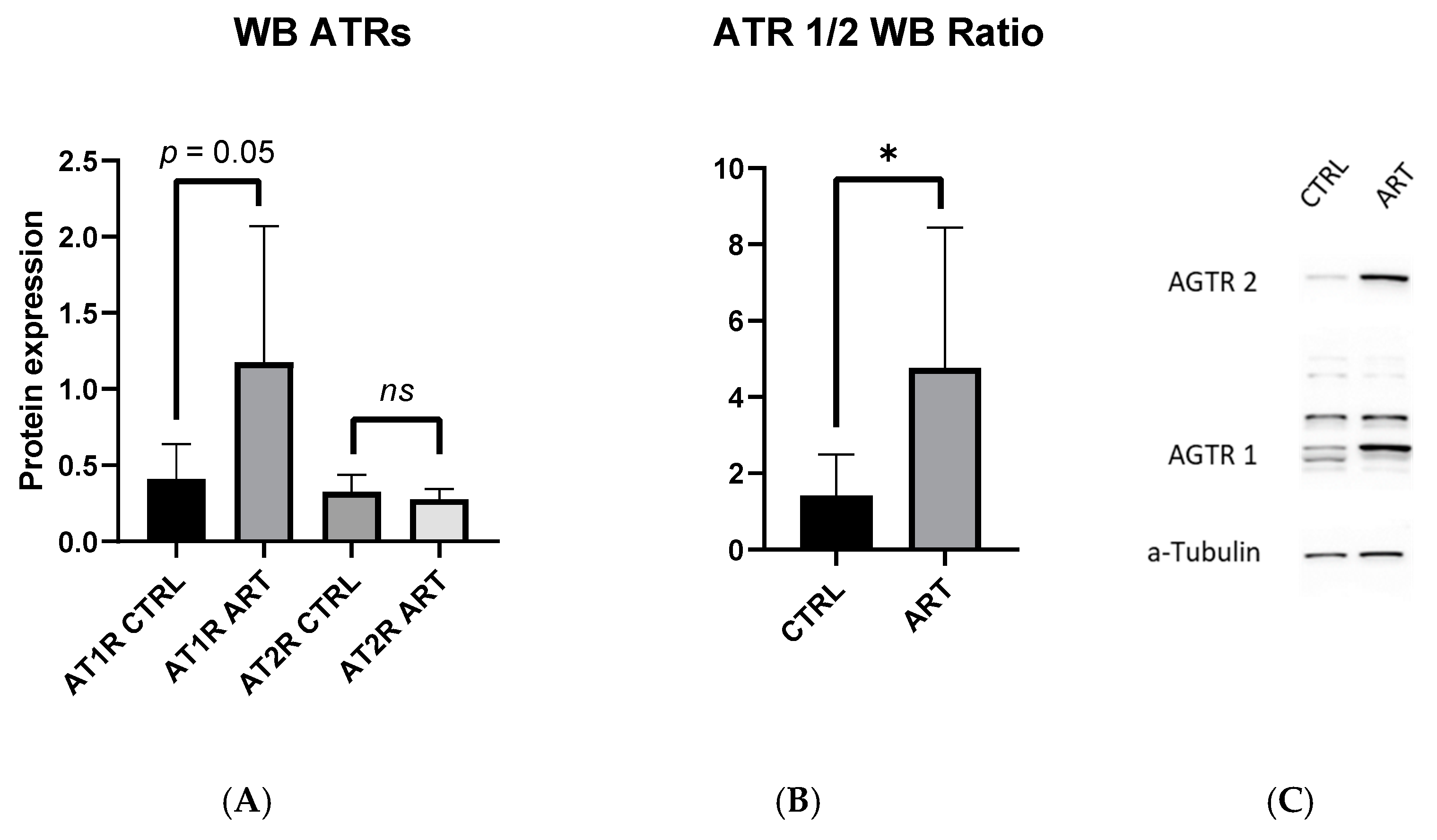
2.2. AT1R to AT2R Ratio in the Aorta Is Increased in ART Mice
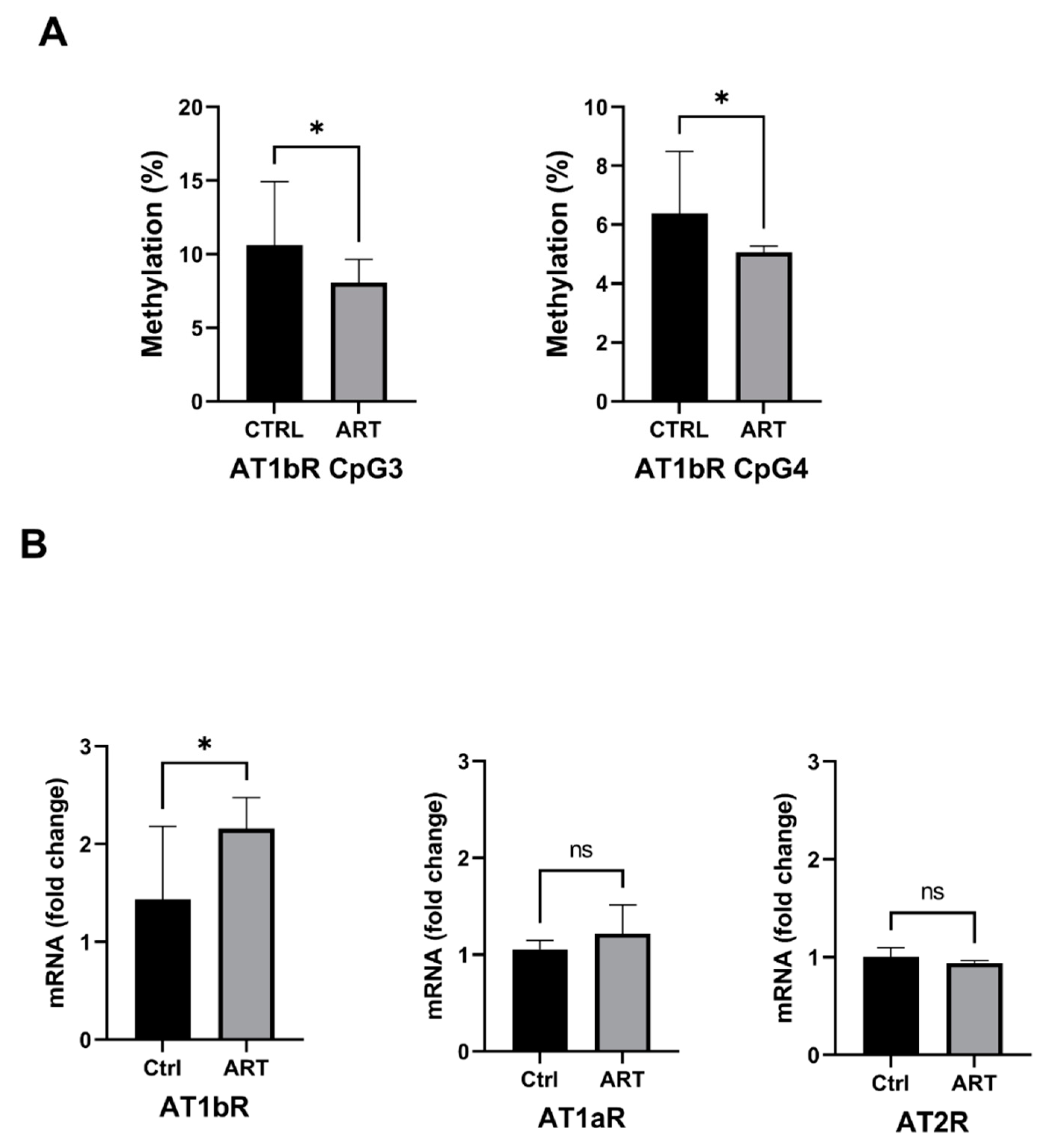
2.3. Increased Expression of the AT1bR Gene Is Correlated with Hypomethylation of Its Promoter in ART Mice
3. Discussion
4. Materials and Methods
4.1. Study Approval
4.2. Animals
4.3. In Vitro Fertilization and Embryo Culture
4.4. Embryo Transfer
4.5. Arterial Blood Pressure
4.6. Isolated Abdominal Aortic Ring Preparation and Vascular Contraction Experiments
4.7. Western Immunoblotting Analysis of Angiotensin II Receptor Proteins
4.8. Angiotensin II Receptors mRNA Preparation and Quantification by Real-Time qPCR
4.9. DNA Methylation Quantitative Measurement by Pyrosequencing of the Angiotensin II Receptor Genes Promoters
4.10. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ART | Assisted Reproductive Technologies |
| ANG II | Angiotensin II |
| AT1R | Angiotensin Receptor 1 |
| AT2R | Angiotensin Receptor 2 |
| eNOS | Endothelial Nitric Oxide Synthase |
| L-NNA | Nw-nitro-L-arginine |
| NO | Nitric Oxide |
| CpG | Cytosine-phosphate-guanine motifs |
References
- Faddy, M.J.; Gosden, M.D.; Gosden, R.G. A demographic projection of the contribution of assisted reproductive technologies to world population growth. Reprod. Biomed Online 2018, 36, 455–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceelen, M.; van Weissenbruch, M.M.; Vermeiden, J.P.; van Leeuwen, F.E.; Delemarre-van de Waal, H.A. Cardiometabolic differences in children born after in vitro fertilization: Follow-up study. J. Clin. Endocrinol. Metab. 2008, 93, 1682–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherrer, U.; Rimoldi, S.F.; Rexhaj, E.; Stuber, T.; Duplain, H.; Garcin, S.; de Marchi, S.F.; Nicod, P.; Germond, M.; Allemann, Y.; et al. Systemic and pulmonary vascular dysfunction in children conceived by assisted reproductive technologies. Circulation 2012, 125, 1890–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenzuela-Alcaraz, B.; Crispi, F.; Bijnens, B.; Cruz-Lemini, M.; Creus, M.; Sitges, M.; Bartrons, J.; Civico, S.; Balasch, J.; Gratacós, E. Assisted Reproductive Technologies Are Associated With Cardiovascular Remodeling In Utero That Persists Postnatally. Circulation 2013, 128, 1442–1450. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.Y.; Liu, X.M.; Jin, L.; Wang, T.T.; Ullah, K.; Sheng, J.Z.; Huang, H.F. Cardiovascular and metabolic profiles of offspring conceived by assisted reproductive technologies: A systematic review and meta-analysis. Fertil. Steril. 2017, 107, 622–631.e625. [Google Scholar] [CrossRef] [Green Version]
- Meister, T.A.; Rimoldi, S.F.; Soria, R.; von Arx, R.; Messerli, F.H.; Sartori, C.; Scherrer, U.; Rexhaj, E. Association of Assisted Reproductive Technologies With Arterial Hypertension During Adolescence. J. Am. Coll. Cardiol. 2018, 72, 1267–1274. [Google Scholar] [CrossRef]
- Watkins, A.J.; Platt, D.; Papenbrock, T.; Wilkins, A.; Eckert, J.J.; Kwong, W.Y.; Osmond, C.; Hanson, M.; Fleming, T.P. Mouse embryo culture induces changes in postnatal phenotype including raised systolic blood pressure. Proc. Natl. Acad. Sci. USA 2007, 104, 5449–5454. [Google Scholar] [CrossRef] [Green Version]
- Rexhaj, E.; Paoloni-Giacobino, A.; Rimoldi, S.F.; Fuster, D.G.; Anderegg, M.; Somm, E.; Bouillet, E.; Allemann, Y.; Sartori, C.; Scherrer, U. Mice generated by in vitro fertilization exhibit vascular dysfunction and shortened life span. J. Clin. Invest. 2013, 123, 5052–5060. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Y.; Le, F.; Wang, N.; Zhang, F.; Luo, Y.; Lou, Y.; Hu, M.; Wang, L.; Thurston, L.M.; et al. Alteration in the expression of the renin-angiotensin system in the myocardium of mice conceived by in vitro fertilization. Biol. Reprod. 2018, 99, 1276–1288. [Google Scholar] [CrossRef]
- Zhang, M.; Lu, L.; Zhang, Y.; Li, X.; Fan, X.; Chen, X.; Tang, J.; Han, B.; Li, M.; Tao, J.; et al. Methylation-reprogrammed AGTR1 results in increased vasoconstriction by angiotensin II in human umbilical cord vessel following in vitro fertilization-embryo transfer. Life Sci. 2019, 234, 116792. [Google Scholar] [CrossRef]
- Valenzuela-Alcaraz, B.; Cruz-Lemini, M.; Rodríguez-López, M.; Goncé, A.; García-Otero, L.; Ayuso, H.; Sitges, M.; Bijnens, B.; Balasch, J.; Gratacós, E.; et al. Fetal cardiac remodeling in twin pregnancy conceived by assisted reproductive technology. Ultrasound Obs. Gynecol. 2018, 51, 94–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vehaskari, V.M.; Stewart, T.; Lafont, D.; Soyez, C.; Seth, D.; Manning, J. Kidney angiotensin and angiotensin receptor expression in prenatally programmed hypertension. Am. J. Physiol. Ren. Physiol. 2004, 287, F262–F267. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rodriguez, P., Jr.; Tong, W.; Xue, Q.; Li, Y.; Hu, S.; Zhang, L. Fetal hypoxia results in programming of aberrant angiotensin ii receptor expression patterns and kidney development. Int. J. Med. Sci. 2013, 10, 532–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.R.; Cullen-McEwen, L.A.; Kett, M.M.; Boon, W.M.; Dowling, J.; Bertram, J.F.; Moritz, K.M. Prenatal corticosterone exposure results in altered AT1/AT2, nephron deficit and hypertension in the rat offspring. J. Physiol. 2007, 579, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Alcaraz, B.; Serafini, A.; Sepulveda-Martínez, A.; Casals, G.; Rodríguez-López, M.; Garcia-Otero, L.; Cruz-Lemini, M.; Bijnens, B.; Sitges, M.; Balasch, J.; et al. Postnatal persistence of fetal cardiovascular remodelling associated with assisted reproductive technologies: A cohort study. BJOG 2019, 126, 291–298. [Google Scholar] [CrossRef] [Green Version]
- Rexhaj, E.; Pireva, A.; Paoloni-Giacobino, A.; Allemann, Y.; Cerny, D.; Dessen, P.; Sartori, C.; Scherrer, U.; Rimoldi, S.F. Prevention of vascular dysfunction and arterial hypertension in mice generated by assisted reproductive technologies by addition of melatonin to culture media. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1151–H1156. [Google Scholar] [CrossRef] [Green Version]
- Elton, T.S.; Stephan, C.C.; Taylor, G.R.; Kimball, M.G.; Martin, M.M.; Durand, J.N.; Oparil, S. Isolation of two distinct type I angiotensin II receptor genes. Biochem. Biophys. Res. Commun. 1992, 184, 1067–1073. [Google Scholar] [CrossRef]
- Iwai, N.; Inagami, T. Identification of two subtypes in the rat type I angiotensin II receptor. FEBS Lett. 1992, 298, 257–260. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Chen, Y.; Dirksen, W.P.; Morris, M.; Periasamy, M. AT1b receptor predominantly mediates contractions in major mouse blood vessels. Circ. Res. 2003, 93, 1089–1094. [Google Scholar] [CrossRef] [Green Version]
- Swafford, A.N.; Harrison-Bernard, L.M.; Dick, G.M. Knockout Mice Reveal That the Angiotensin II type 1B Receptor Links to Smooth Muscle Contraction. Am. J. Hypertens. 2007, 20, 335–337. [Google Scholar] [CrossRef]
- Chen, F.; Cao, K.; Zhang, H.; Yu, H.; Liu, Y.; Xue, Q. Maternal high-fat diet increases vascular contractility in adult offspring in a sex-dependent manner. Hypertens. Res. 2020, 44, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Dasgupta, C.; Li, Y.; Huang, X.; Zhang, L. Perinatal nicotine exposure increases angiotensin II receptor-mediated vascular contractility in adult offspring. PLoS ONE 2014, 9, e108161. [Google Scholar] [CrossRef] [PubMed]
- Pei, F.; Wang, X.; Yue, R.; Chen, C.; Huang, J.; Huang, J.; Li, X.; Zeng, C. Differential expression and DNA methylation of angiotensin type 1A receptors in vascular tissues during genetic hypertension development. Mol. Cell. Biochem. 2015, 402, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bogdarina, I.; Welham, S.; King, P.J.; Burns, S.P.; Clark, A.J.L. Epigenetic modification of the renin-angiotensin system in the fetal programming of hypertension. Circ. Res. 2007, 100, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, M.; Robker, R.L.; Robertson, S.A. Parenting from before conception. Science 2014, 345, 756–760. [Google Scholar] [CrossRef]
- Nickenig, G.; Harrison, D.G. The AT1-Type Angiotensin Receptor in Oxidative Stress and Atherogenesis. Circulation 2002, 105, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Nguyen Dinh Cat, A.; Montezano, A.C.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef]
- Rimoldi, S.F.; Sartori, C.; Rexhaj, E.; Bailey, D.M.; Marchi, S.F.d.; McEneny, J.; Arx, R.v.; Cerny, D.; Duplain, H.; Germond, M.; et al. Antioxidants improve vascular function in children conceived by assisted reproductive technologies: A randomized double-blind placebo-controlled trial. Eur. J. Prev. Cardiol. 2015, 22, 1399–1407. [Google Scholar] [CrossRef]
- Cerny, D.; Sartori, C.; Rimoldi, S.F.; Meister, T.; Soria, R.; Bouillet, E.; Scherrer, U.; Rexhaj, E. Assisted Reproductive Technologies Predispose to Insulin Resistance and Obesity in Male Mice Challenged With a High-Fat Diet. Endocrinology 2017, 158, 1152–1159. [Google Scholar] [CrossRef]
- Rose, C.; Schwegler, H.; Hanke, J.; Yilmazer-Hanke, D.M. Pregnancy rates, prenatal and postnatal survival of offspring, and litter sizes after reciprocal embryo transfer in DBA/2JHd, C3H/HeNCrl and NMRI mice. Theriogenology 2012, 77, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- Gollasch, M. Vasodilator signals from perivascular adipose tissue. Br. J. Pharm. 2012, 165, 633–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulvany, M.J.; Halpern, W. Contractile properties of small arterial resistance vessels in spontaneously hypertensive and normotensive rats. Circ. Res. 1977, 41, 19–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, D.D.; Palmer, R.M.; Schulz, R.; Hodson, H.F.; Moncada, S. Characterization of three inhibitors of endothelial nitric oxide synthase in vitro and in vivo. Br. J. Pharmacol. 1990, 101, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Kretz, M.; Mundy, A.L.; Widmer, C.C.; Barton, M. Early Aging and Anatomic Heterogeneity Determine Cyclooxygenase-mediated Vasoconstriction to Angiotensin II in Mice. J. Cardiovasc. Pharmacol. 2006, 48, 30–33. [Google Scholar] [CrossRef]
- Mehta, P.K.; Griendling, K.K. Angiotensin II cell signaling: Physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 2007, 292, C82–C97. [Google Scholar] [CrossRef]
- Stouder, C.; Deutsch, S.; Paoloni-Giacobino, A. Superovulation in mice alters the methylation pattern of imprinted genes in the sperm of the offspring. Reprod. Toxicol. 2009, 28, 536–541. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meister, T.A.; Soria, R.; Dogar, A.; Messerli, F.H.; Paoloni-Giacobino, A.; Stenz, L.; Scherrer, U.; Sartori, C.; Rexhaj, E. Increased Arterial Responsiveness to Angiotensin II in Mice Conceived by Assisted Reproductive Technologies. Int. J. Mol. Sci. 2022, 23, 13357. https://doi.org/10.3390/ijms232113357
Meister TA, Soria R, Dogar A, Messerli FH, Paoloni-Giacobino A, Stenz L, Scherrer U, Sartori C, Rexhaj E. Increased Arterial Responsiveness to Angiotensin II in Mice Conceived by Assisted Reproductive Technologies. International Journal of Molecular Sciences. 2022; 23(21):13357. https://doi.org/10.3390/ijms232113357
Chicago/Turabian StyleMeister, Theo Arthur, Rodrigo Soria, Afzal Dogar, Franz H. Messerli, Ariane Paoloni-Giacobino, Ludwig Stenz, Urs Scherrer, Claudio Sartori, and Emrush Rexhaj. 2022. "Increased Arterial Responsiveness to Angiotensin II in Mice Conceived by Assisted Reproductive Technologies" International Journal of Molecular Sciences 23, no. 21: 13357. https://doi.org/10.3390/ijms232113357