
Proof-of-Principle Study Suggesting Potential Anti-Inflammatory Activity of Butyrate and Propionate in Periodontal Cells


 , and
, and
Abstract
:1. Introduction
2. Results
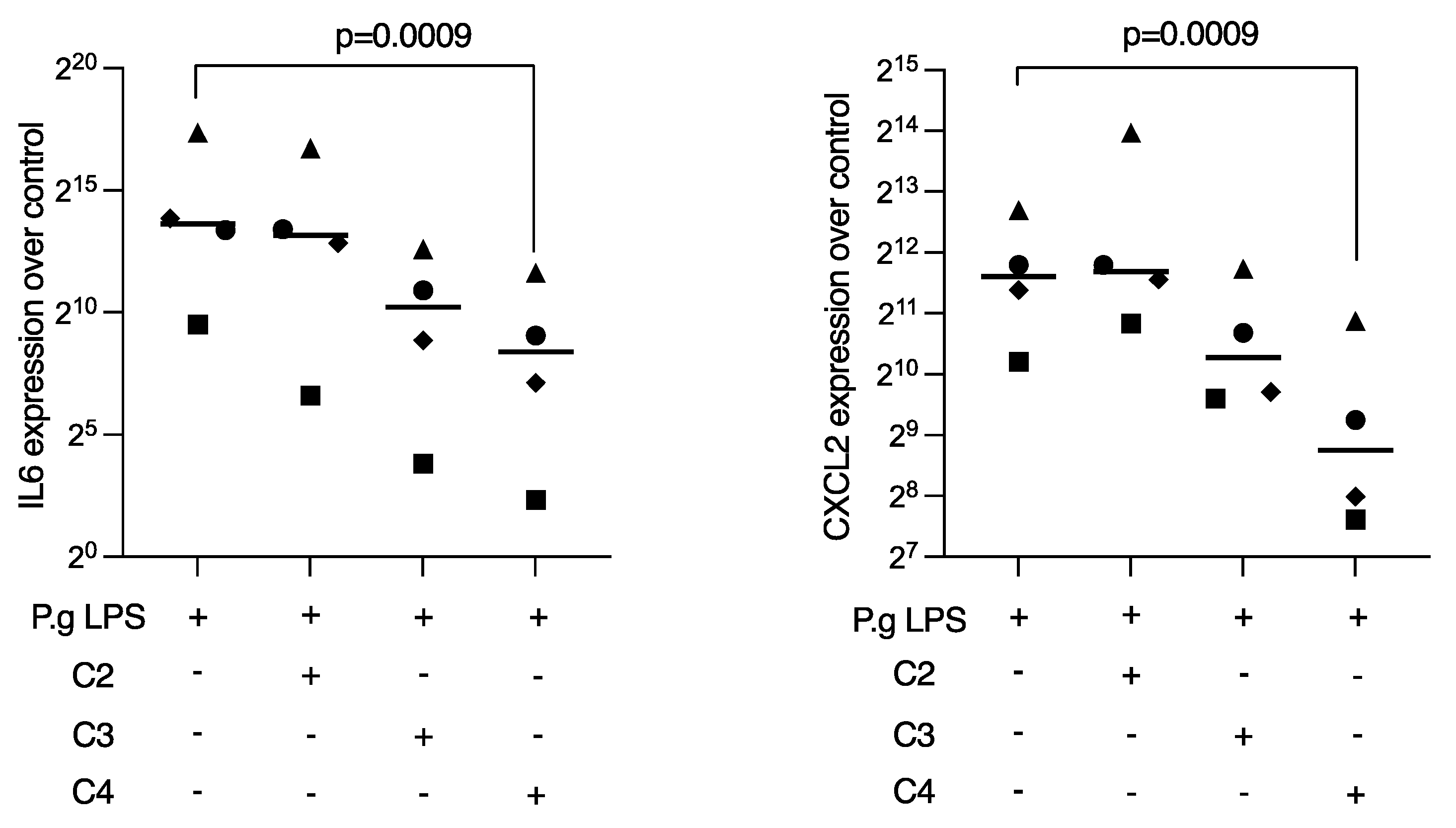
2.1. Butyrate Suppressed LPS-Induced IL6 and CXCL2 Expression in RAW 264.7 Cells
2.2. Butyrate Reduced NF-κB Signaling in RAW 264.7 Cells
2.3. Butyrate Suppressed Fimbriae and Saliva-Induced IL6 and CXCL2 Expression in Primary Macrophages
2.4. Butyrate Reduced Inflammatory Mediators in Fibroblasts Exposed to IL1β and TNFα
2.5. Propionate and Butyrate Reduced Inflammatory Mediators in HSC2 Cells Exposed to IL1β and TNFα
2.6. Butyrate Decreased the Translocation of p65 in Gingival Fibroblasts and HSC2 Cells
3. Discussion
4. Materials and Methods
4.1. Primary Macrophages and RAW 264.7 Macrophage-like Cells
4.2. Gingival Fibroblasts and HSC2 Oral Epithelial Cells
4.3. Viability Assay
4.4. Cell Stimulation
4.5. qRT-PCR Analysis and Immunoassay
4.6. Immunofluorescence
4.7. Western Blot
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nanci, A.; Bosshardt, D.D. Structure of periodontal tissues in health and disease. Periodontol. 2000 2006, 40, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Bosshardt, D.D. The periodontal pocket: Pathogenesis, histopathology and consequences. Periodontol. 2000 2018, 76, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Cochran, D.L. Inflammation and bone loss in periodontal disease. J. Periodontol. 2008, 79 (Suppl. S8), 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L.; Wolin, M.J. Pathways of acetate, propionate, and butyrate formation by the human fecal microbial flora. Appl. Environ. Microbiol. 1996, 62, 1589–1592. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Schonfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Guan, X.; Li, W.; Meng, H. A double-edged sword: Role of butyrate in the oral cavity and the gut. Mol. Oral Microbiol. 2021, 36, 121–131. [Google Scholar] [CrossRef]
- Singer, R.E.; Buckner, B.A. Butyrate and propionate: Important components of toxic dental plaque extracts. Infect. Immun. 1981, 32, 458–463. [Google Scholar] [CrossRef]
- Tonetti, M.; Eftimiadi, C.; Damiani, G.; Buffa, P.; Buffa, D.; Botta, G.A. Short chain fatty acids present in periodontal pockets may play a role in human periodontal diseases. J. Periodontal Res. 1987, 22, 190–191. [Google Scholar] [CrossRef]
- Eftimiadi, C.; Buzzi, E.; Tonetti, M.; Buffa, P.; Buffa, D.; van Steenbergen, M.T.; de Graaff, J.; Botta, G.A. Short-chain fatty acids produced by anaerobic bacteria alter the physiological responses of human neutrophils to chemotactic peptide. J. Infect. 1987, 14, 43–53. [Google Scholar] [CrossRef]
- Eftimiadi, C.; Tonetti, M.; Cavallero, A.; Sacco, O.; Rossi, G.A. Short-chain fatty acids produced by anaerobic bacteria inhibit phagocytosis by human lung phagocytes. J. Infect. Dis. 1990, 161, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Eftimiadi, C.; Stashenko, P.; Tonetti, M.; Mangiante, P.E.; Massara, R.; Zupo, S.; Ferrarini, M. Divergent effect of the anaerobic bacteria by-product butyric acid on the immune response: Suppression of T-lymphocyte proliferation and stimulation of interleukin-1 beta production. Oral Microbiol. Immunol. 1991, 6, 17–23. [Google Scholar] [CrossRef]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.Y.; Naleway, C.; Robinson, P.; Kent, R. The relationship of gingival crevicular fluid short chain carboxylic acid concentration to gingival inflammation. J. Clin. Periodontol. 1996, 23, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.Y.; Robinson, P.; Naleway, C. Short-chain carboxylic acid concentration in human gingival crevicular fluid. J. Dent. Res. 1997, 76, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Meng, H.; Gao, X.; Xu, L.; Feng, X. Effect of non-surgical periodontal treatment on short chain fatty acid levels in gingival crevicular fluid of patients with generalized aggressive periodontitis. J. Periodontal Res. 2014, 49, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Sasaki, M.; Tsujikawa, T.; Fujiyama, Y.; Bamba, T.; Kusunoki, M. Preventive efficacy of butyrate enemas and oral administration of Clostridium butyricum M588 in dextran sodium sulfate-induced colitis in rats. J. Gastroenterol. 2000, 35, 341–346. [Google Scholar] [CrossRef]
- Yue, M.; Tao, Y.; Fang, Y.; Lian, X.; Zhang, Q.; Xia, Y.; Wei, Z.; Dai, Y. The gut microbiota modulator berberine ameliorates collagen-induced arthritis in rats by facilitating the generation of butyrate and adjusting the intestinal hypoxia and nitrate supply. FASEB J. 2019, 33, 12311–12323. [Google Scholar] [CrossRef]
- Jia, X.; Jia, L.; Mo, L.; Yuan, S.; Zheng, X.; He, J.; Chen, V.; Guo, Q.; Zheng, L.; Yuan, Q.; et al. Berberine Ameliorates Periodontal Bone Loss by Regulating Gut Microbiota. J. Dent. Res. 2019, 98, 107–116. [Google Scholar] [CrossRef]
- Li, L.; Bao, J.; Chang, Y.; Wang, M.; Chen, B.; Yan, F. Gut Microbiota May Mediate the Influence of Periodontitis on Prediabetes. J. Dent. Res. 2021, 100, 1387–1396. [Google Scholar] [CrossRef]
- Cheng, X.; Zhou, X.; Liu, C.; Xu, X. Oral Osteomicrobiology: The Role of Oral Microbiota in Alveolar Bone Homeostasis. Front. Cell Infect. Microbiol. 2021, 11, 751503. [Google Scholar] [CrossRef] [PubMed]
- Irie, K.; Novince, C.M.; Darveau, R.P. Impact of the Oral Commensal Flora on Alveolar Bone Homeostasis. J. Dent. Res. 2014, 93, 801–806. [Google Scholar] [CrossRef]
- Shirasugi, M.; Nakagawa, M.; Nishioka, K.; Yamamoto, T.; Nakaya, T.; Kanamura, N. Relationship between periodontal disease and butyric acid produced by periodontopathic bacteria. Inflamm. Regen. 2018, 38, 23. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Kim, B.G.; Kim, J.H.; Chun, J.; Im, J.P.; Kim, J.S. Sodium butyrate inhibits the NF-kappa B signaling pathway and histone deacetylation, and attenuates experimental colitis in an IL-10 independent manner. Int. Immunopharmacol. 2017, 51, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Chakravortty, D.; Koide, N.; Kato, Y.; Sugiyama, T.; Mu, M.M.; Yoshida, T.; Yokochi, T. The inhibitory action of butyrate on lipopolysaccharide-induced nitric oxide production in RAW 264.7 murine macrophage cells. J. Endotoxin. Res. 2000, 6, 243–247. [Google Scholar] [CrossRef]
- Wang, F.; Liu, J.; Weng, T.; Shen, K.; Chen, Z.; Yu, Y.; Huang, Q.; Wang, G.; Liu, Z.; Jin, S. The Inflammation Induced by Lipopolysaccharide can be Mitigated by Short-chain Fatty Acid, Butyrate, through Upregulation of IL-10 in Septic Shock. Scand. J. Immunol. 2017, 85, 258–263. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, E.J.; Lee, J.C.; Kim, W.K.; Kim, H.S. Anti-inflammatory effects of short chain fatty acids in IFN-gamma-stimulated RAW 264.7 murine macrophage cells: Involvement of NF-kappaB and ERK signaling pathways. Int. Immunopharmacol. 2007, 7, 70–77. [Google Scholar] [CrossRef]
- Takeshita, A.; Murakami, Y.; Yamashita, Y.; Ishida, M.; Fujisawa, S.; Kitano, S.; Hanazawa, S. Porphyromonas gingivalis fimbriae use beta2 integrin (CD11/CD18) on mouse peritoneal macrophages as a cellular receptor, and the CD18 beta chain plays a functional role in fimbrial signaling. Infect. Immun. 1998, 66, 4056–4060. [Google Scholar] [CrossRef]
- Saito, A.; Sojar, H.T.; Genco, R.J. Interleukin-1 gene expression in macrophages induced by surface protein components of Porphyromonas gingivalis: Role of tyrosine kinases in signal transduction. Oral Microbiol. Immunol. 1997, 12, 135–140. [Google Scholar] [CrossRef]
- Hanazawa, S.; Murakami, Y.; Hirose, K.; Amano, S.; Ohmori, Y.; Higuchi, H.; Kitano, S. Bacteroides (Porphyromonas) gingivalis fimbriae activate mouse peritoneal macrophages and induce gene expression and production of interleukin-1. Infect. Immun. 1991, 59, 1972–1977. [Google Scholar] [CrossRef] [Green Version]
- Pourgonabadi, S.; Muller, H.D.; Mendes, J.R.; Gruber, R. Saliva initiates the formation of pro-inflammatory macrophages in vitro. Arch. Oral Biol. 2017, 73, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.H.; Chan, C.P.; Ho, Y.S.; Lan, W.H.; Hsieh, C.C.; Chang, M.C. Effects of butyrate and propionate on the adhesion, growth, cell cycle kinetics, and protein synthesis of cultured human gingival fibroblasts. J. Periodontol. 1999, 70, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Tsai, Y.L.; Chen, Y.W.; Chan, C.P.; Huang, C.F.; Lan, W.C.; Lin, C.C.; Lan, W.H.; Jeng, J.H. Butyrate induces reactive oxygen species production and affects cell cycle progression in human gingival fibroblasts. J. Periodontal Res. 2013, 48, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Kurita-Ochiai, T.; Seto, S.; Suzuki, N.; Yamamoto, M.; Otsuka, K.; Abe, K.; Ochiai, K. Butyric acid induces apoptosis in inflamed fibroblasts. J. Dent. Res. 2008, 87, 51–55. [Google Scholar] [CrossRef]
- Nakagawa, M.; Shirasugi, M.; Yamamoto, T.; Nakaya, T.; Kanamura, N. Long-term exposure to butyric acid induces excessive production of matrix metalloproteases in human gingival fibroblasts. Arch. Oral Biol. 2021, 123, 105035. [Google Scholar] [CrossRef] [PubMed]
- Shirasugi, M.; Nishioka, K.; Yamamoto, T.; Nakaya, T.; Kanamura, N. Normal human gingival fibroblasts undergo cytostasis and apoptosis after long-term exposure to butyric acid. Biochem. Biophys. Res. Commun. 2017, 482, 1122–1128. [Google Scholar] [CrossRef]
- Friscic, J.; Durholz, K.; Chen, X.; Engdahl, C.; Moller, L.; Schett, G.; Zaiss, M.M.; Hoffmann, M.H. Dietary Derived Propionate Regulates Pathogenic Fibroblast Function and Ameliorates Experimental Arthritis and Inflammatory Tissue Priming. Nutrients 2021, 13, 1643. [Google Scholar] [CrossRef]
- Shao, F.; Panahipour, L.; Sordi, M.B.; Tang, F.; Liu, R.; Gruber, R. Heartwood of Dalbergia cochinchinensis: 4,7,2′-Trihydroxy-4′-methoxyisoflavanol and 6,4′-Dihydroxy-7-methoxyflavane Reduce Cytokine and Chemokine Expression In Vitro. Molecules 2022, 27, 1321. [Google Scholar] [CrossRef]
- Magrin, G.L.; Strauss, F.J.; Benfatti, C.A.M.; Maia, L.C.; Gruber, R. Effects of Short-Chain Fatty Acids on Human Oral Epithelial Cells and the Potential Impact on Periodontal Disease: A Systematic Review of In Vitro Studies. Int. J. Mol. Sci. 2020, 21, 4895. [Google Scholar] [CrossRef]
- Magrin, G.L.; Di Summa, F.; Strauss, F.J.; Panahipour, L.; Mildner, M.; Magalhaes Benfatti, C.A.; Gruber, R. Butyrate Decreases ICAM-1 Expression in Human Oral Squamous Cell Carcinoma Cells. Int. J. Mol. Sci. 2020, 21, 1679. [Google Scholar] [CrossRef] [Green Version]
- Zang, W.; Liu, J.; Geng, F.; Liu, D.; Zhang, S.; Li, Y.; Pan, Y. Butyrate promotes oral squamous cell carcinoma cells migration, invasion and epithelial-mesenchymal transition. PeerJ 2022, 10, e12991. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Mukae, S.; Murakami, M.; Ishikawa, Y.; Ishii, T.; Ohki, H.; Matsumoto, M.; Komiyama, K. Butyrate inhibits oral cancer cell proliferation and regulates expression of secretory phospholipase A2-X and COX-2. Anticancer Res. 2007, 27, 1493–1502. [Google Scholar] [PubMed]
- Wang, A.; Zeng, R.; Huang, H. Retinoic acid and sodium butyrate as cell cycle regulators in the treatment of oral squamous carcinoma cells. Oncol. Res. 2008, 17, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Meng, H.; Yu, J.; Lu, H.; Li, W.; Lu, R.; Zhao, Y.; Li, Q.; Su, L. Butyrate rather than LPS subverts gingival epithelial homeostasis by downregulation of intercellular junctions and triggering pyroptosis. J. Clin. Periodontol. 2019, 46, 894–907. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, T.; Matsumoto, N.; Matsue, Y.; Okudera, M.; Nishikawa, Y.; Abiko, Y.; Komiyama, K. Sodium butyrate, a histone deacetylase inhibitor, regulates Lymphangiogenic factors in oral cancer cell line HSC-3. Anticancer Res. 2014, 34, 1701–1708. [Google Scholar] [PubMed]
- Tsuda, H.; Ochiai, K.; Suzuki, N.; Otsuka, K. Butyrate, a bacterial metabolite, induces apoptosis and autophagic cell death in gingival epithelial cells. J. Periodontal Res. 2010, 45, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, Y.; Murofushi, T.; Koshi, R.; Mikami, Y.; Tsuda, H. Reactive oxygen species-dependent release of damage-associated molecular patterns from human gingival epithelial Ca9-22 cells during butyrate or propionate exposure. J. Oral Sci. 2021, 63, 195–197. [Google Scholar] [CrossRef]
- Stadnyk, A.W. Cytokine production by epithelial cells. FASEB J. 1994, 8, 1041–1047. [Google Scholar] [CrossRef]
- Kargarpour, Z.; Nasirzade, J.; Panahipour, L.; Miron, R.J.; Gruber, R. Platelet-Rich Fibrin Decreases the Inflammatory Response of Mesenchymal Cells. Int. J. Mol. Sci. 2021, 22, 11333. [Google Scholar] [CrossRef]
- Rutting, S.; Xenaki, D.; Malouf, M.; Horvat, J.C.; Wood, L.G.; Hansbro, P.M.; Oliver, B.G. Short-chain fatty acids increase TNFalpha-induced inflammation in primary human lung mesenchymal cells through the activation of p38 MAPK. Am. J. Physiol. Lung Cell Mol. Physiol. 2019, 316, L157–L174. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Guo, H.L.; Deng, X.; Zhu, T.T.; Xiong, J.F.; Xu, Y.H.; Xu, Y. Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide. Exp. Clin. Endocrinol. Diabetes 2017, 125, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Moylan, H.E.C.; Nguyen-Ngo, C.; Lim, R.; Lappas, M. The short-chain fatty acids butyrate and propionate protect against inflammation-induced activation of mediators involved in active labor: Implications for preterm birth. Mol. Hum. Reprod. 2020, 26, 452–468. [Google Scholar] [CrossRef] [PubMed]
- Ohira, H.; Fujioka, Y.; Katagiri, C.; Mamoto, R.; Aoyama-Ishikawa, M.; Amako, K.; Izumi, Y.; Nishiumi, S.; Yoshida, M.; Usami, M.; et al. Butyrate attenuates inflammation and lipolysis generated by the interaction of adipocytes and macrophages. J. Atheroscler Thromb. 2013, 20, 425–442. [Google Scholar] [CrossRef]
- Zhou, Y.; Ji, X.; Chen, J.; Fu, Y.; Huang, J.; Guo, R.; Zhou, J.; Cen, J.; Zhang, Q.; Chu, A.; et al. Short-chain fatty acid butyrate: A novel shield against chronic gastric ulcer. Exp. Ther. Med. 2021, 21, 329. [Google Scholar] [CrossRef]
- Ali, I.; Li, C.; Kuang, M.; Shah, A.U.; Shafiq, M.; Ahmad, M.A.; Abdalmegeed, D.; Li, L.; Wang, G. Nrf2 Activation and NF-Kb & caspase/bax signaling inhibition by sodium butyrate alleviates LPS-induced cell injury in bovine mammary epithelial cells. Mol. Immunol. 2022, 148, 54–67. [Google Scholar] [PubMed]
- Li, M.; van Esch, B.; Henricks, P.A.J.; Garssen, J.; Folkerts, G. IL-33 Is Involved in the Anti-Inflammatory Effects of Butyrate and Propionate on TNFalpha-Activated Endothelial Cells. Int. J. Mol. Sci. 2021, 22, 2447. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.; Yang, M.; Zhang, M.; Xiao, M.; Li, X. Butyrate mitigates TNF-alpha-induced attachment of monocytes to endothelial cells. J. Bioenerg. Biomembr. 2020, 52, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Farkas, O.; Matis, G.; Paszti-Gere, E.; Palocz, O.; Kulcsar, A.; Petrilla, J.; Csiko, G.; Neogrady, Z.; Galfi, P. Effects of Lactobacillus plantarum 2142 and sodium n-butyrate in lipopolysaccharide-triggered inflammation: Comparison of a porcine intestinal epithelial cell line and primary hepatocyte monocultures with a porcine enterohepatic co-culture system. J. Anim. Sci. 2014, 92, 3835–3845. [Google Scholar] [CrossRef]
- Korsten, S.; Peracic, L.; van Groeningen, L.M.B.; Diks, M.A.P.; Vromans, H.; Garssen, J.; Willemsen, L.E.M. Butyrate Prevents Induction of CXCL10 and Non-Canonical IRF9 Expression by Activated Human Intestinal Epithelial Cells via HDAC Inhibition. Int. J. Mol. Sci. 2022, 23, 3980. [Google Scholar] [CrossRef]
- Llama-Palacios, A.; Potupa, O.; Sanchez, M.C.; Figuero, E.; Herrera, D.; Sanz, M. Proteomic analysis of Fusobacterium nucleatum growth in biofilm versus planktonic state. Mol. Oral Microbiol. 2020, 35, 168–180. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| wo | P.g LPS | P.g LPS + C4 | |
|---|---|---|---|
| Experiment 1 | 167 | 880 | 221 |
| Experiment 2 | 113 | 789 | 167 |
| wo | Fimbriae | Fimbriae + C4 | Saliva | Saliva + C4 | |
|---|---|---|---|---|---|
| Experiment 1 | 481 | 4872 | 706 | 15,469 | 1234 |
| Experiment 2 | 320 | 1691 | 565 | 1827 | 442 |
| Primer | Sequence Forward | Sequence Reverse |
|---|---|---|
| IL6 | gctaccaaactggatataatcagga | ccaggtagctatggtactccagaa |
| CXCL2 | catccagagcttgagtgtgacg | ggcttcagggtcaaggcaaac |
| GAPDH | aactttggcattgtggaagg | ggatgcagggatgatgttct |
| Primer | Sequence Forward | Sequence Reverse |
|---|---|---|
| IL8 | aacttctccacaaccctctg | ttggcagccttcctgatttc |
| CXCL1 | tcctgcatcccccatagtta | cttcaggaacagccaccagt |
| CXCL2 | cccatggttaagaaaatcatcg | cttcaggaacagccaccaat |
| GAPDH | aagccacatcgctcagacac | gcccaatacgaccaaatcc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.F.P.; Cervantes, L.C.C.; Panahipour, L.; Souza, F.Á.; Gruber, R. Proof-of-Principle Study Suggesting Potential Anti-Inflammatory Activity of Butyrate and Propionate in Periodontal Cells. Int. J. Mol. Sci. 2022, 23, 11006. https://doi.org/10.3390/ijms231911006
Santos AFP, Cervantes LCC, Panahipour L, Souza FÁ, Gruber R. Proof-of-Principle Study Suggesting Potential Anti-Inflammatory Activity of Butyrate and Propionate in Periodontal Cells. International Journal of Molecular Sciences. 2022; 23(19):11006. https://doi.org/10.3390/ijms231911006
Chicago/Turabian StyleSantos, Ana Flávia Piquera, Lara Cristina Cunha Cervantes, Layla Panahipour, Francisley Ávila Souza, and Reinhard Gruber. 2022. "Proof-of-Principle Study Suggesting Potential Anti-Inflammatory Activity of Butyrate and Propionate in Periodontal Cells" International Journal of Molecular Sciences 23, no. 19: 11006. https://doi.org/10.3390/ijms231911006