
Directed Evolution of Phi Class Glutathione Transferases Involved in Multiple-Herbicide Resistance of Grass Weeds and Crops


Abstract
:1. Introduction
2. Results and Discussion
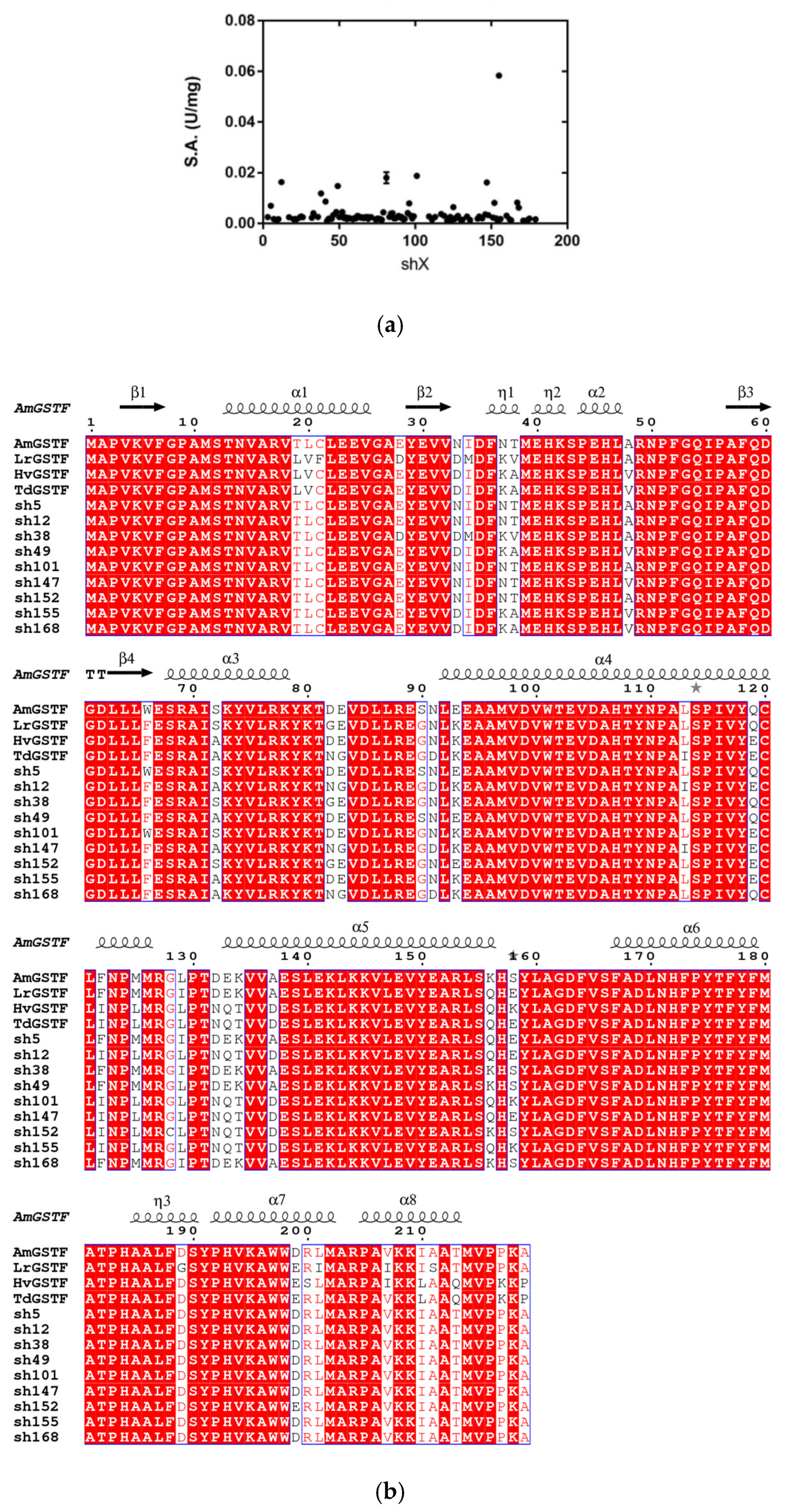
2.1. Shuffling of Parental GSTF Genes Encoded in A. myosuroides, L. rigidum, T. durum and H. vulgare and Activity Screening
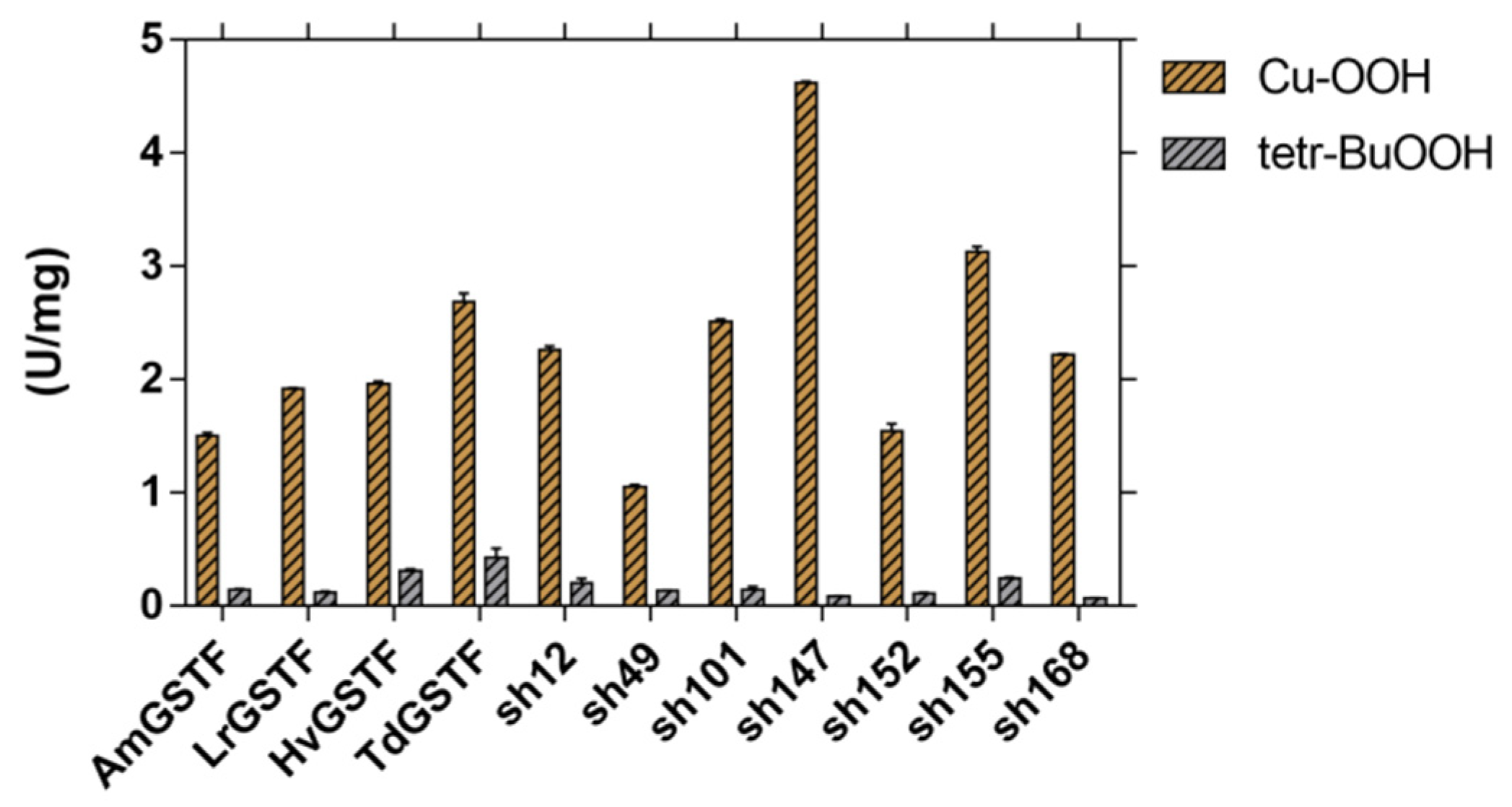
2.2. Kinetic Studies
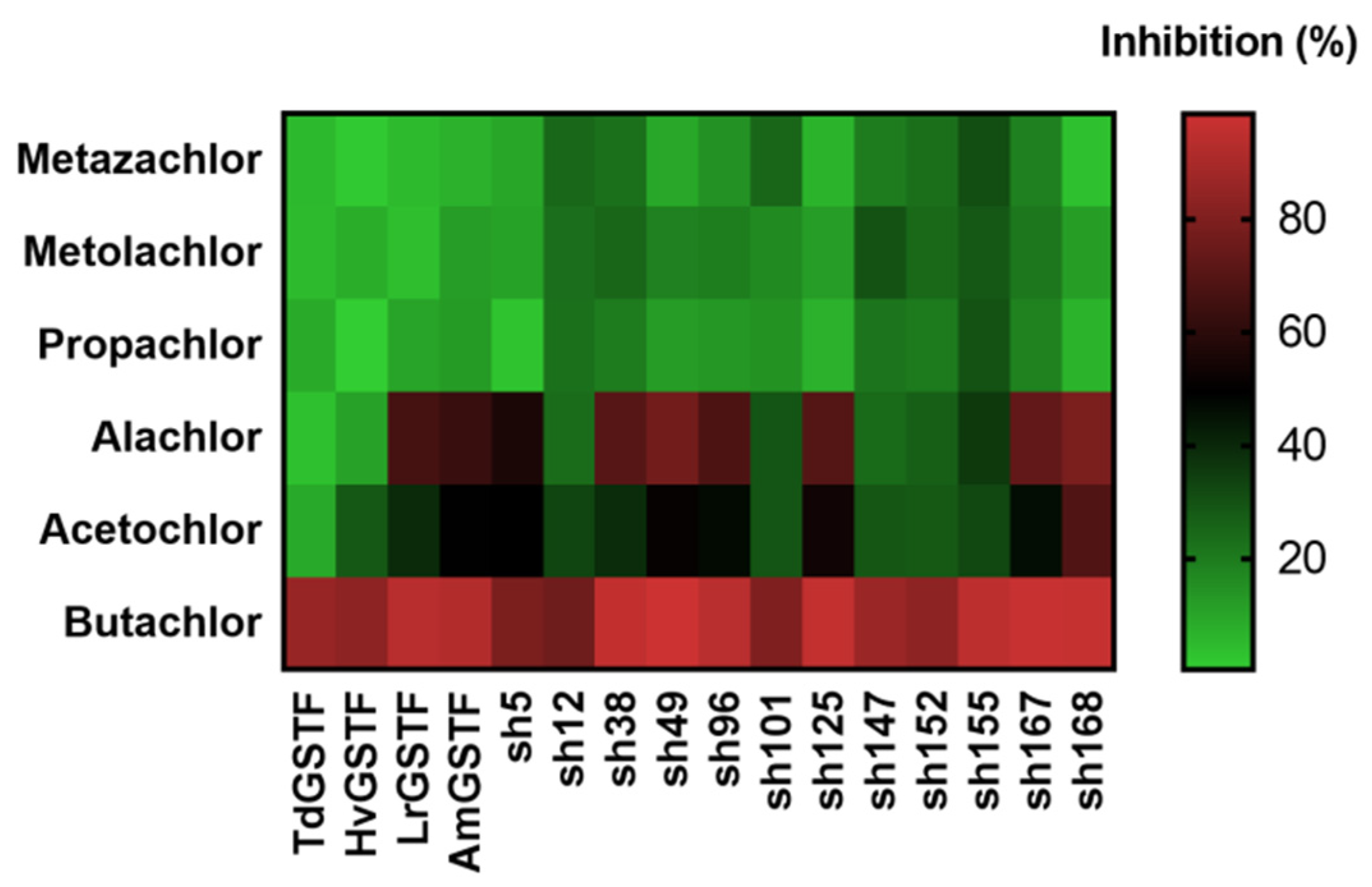
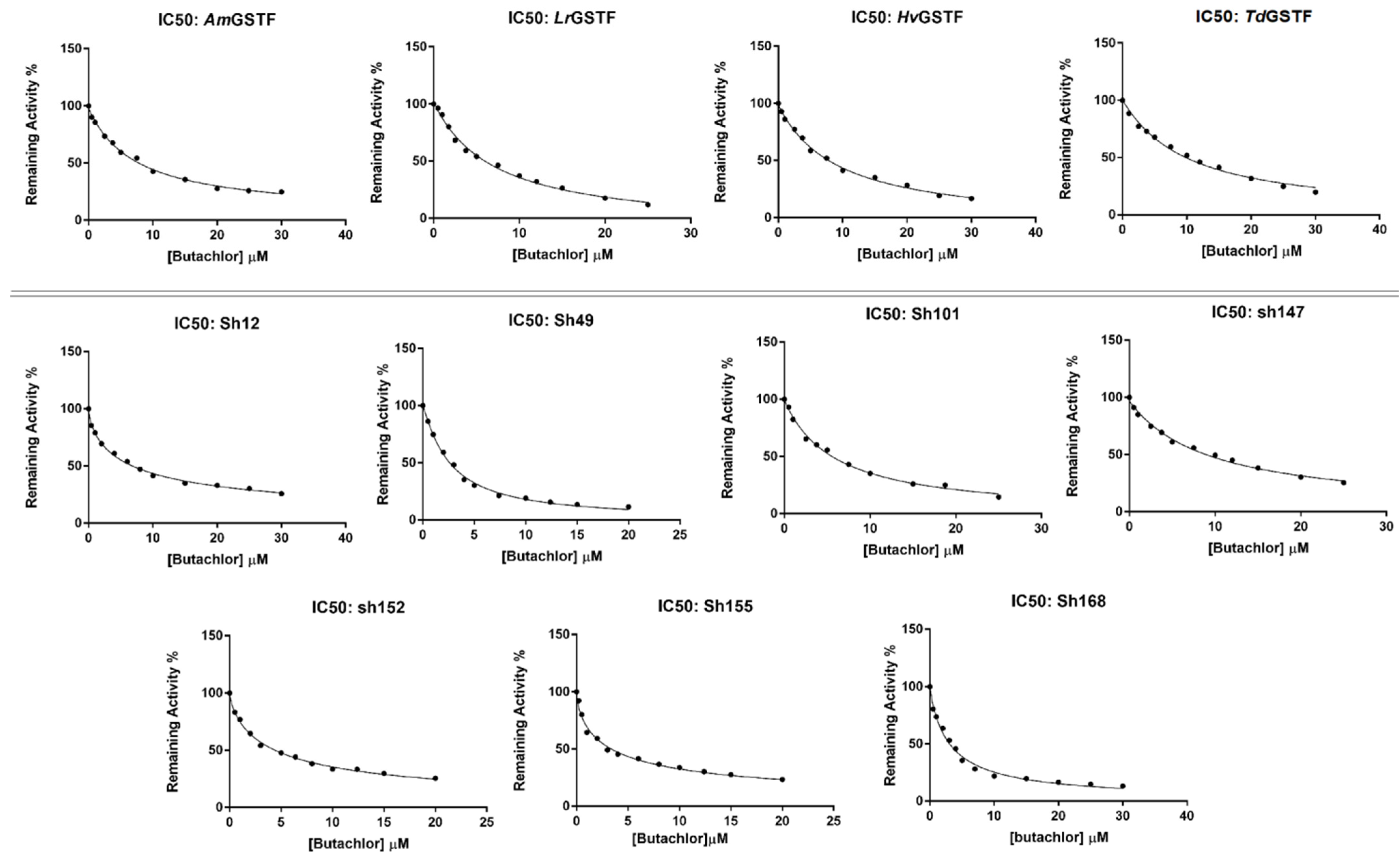
2.3. Determination of Butachlor’s Half-Maximal Inhibitory Concentration (IC50) for the GSTF Library
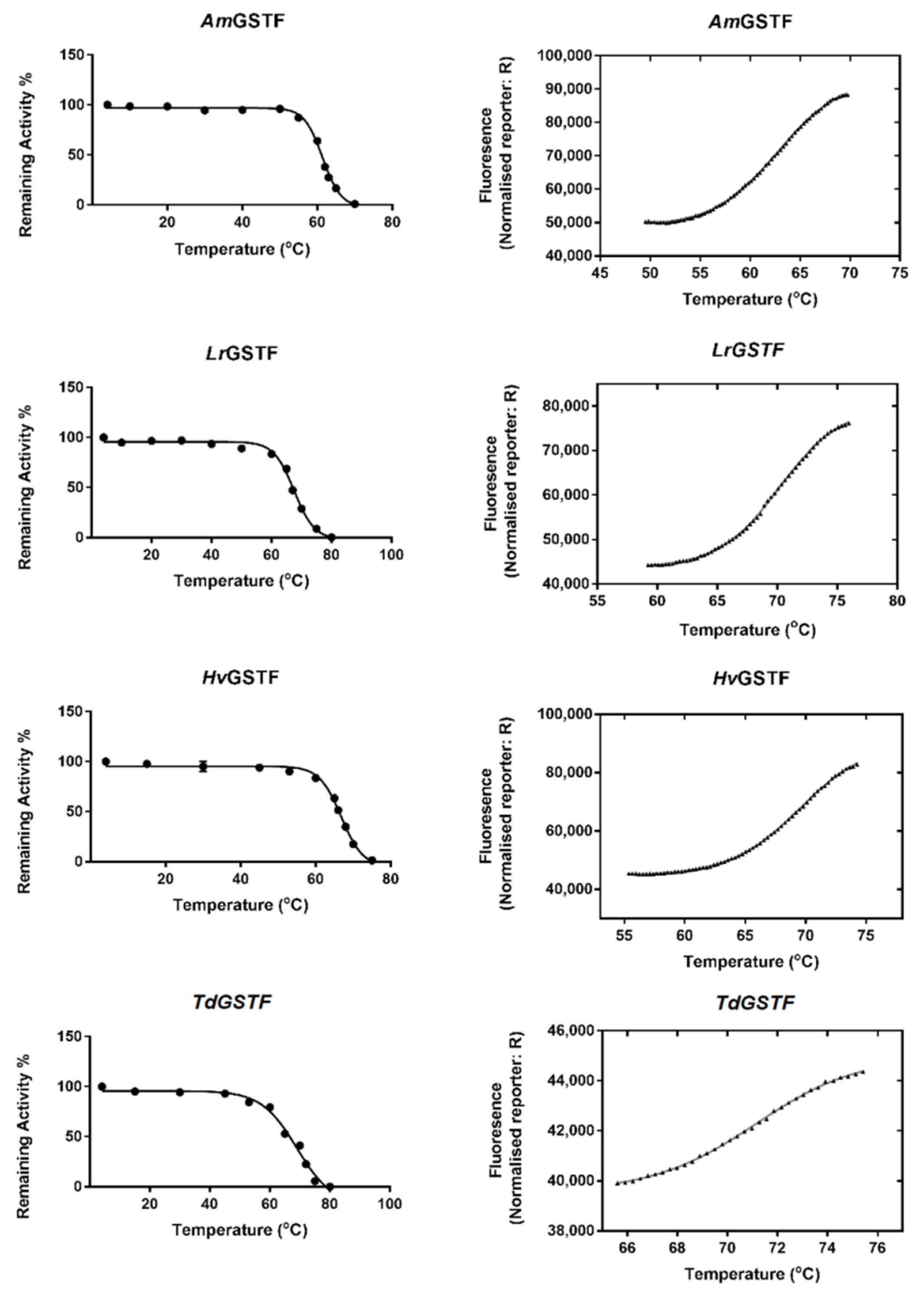
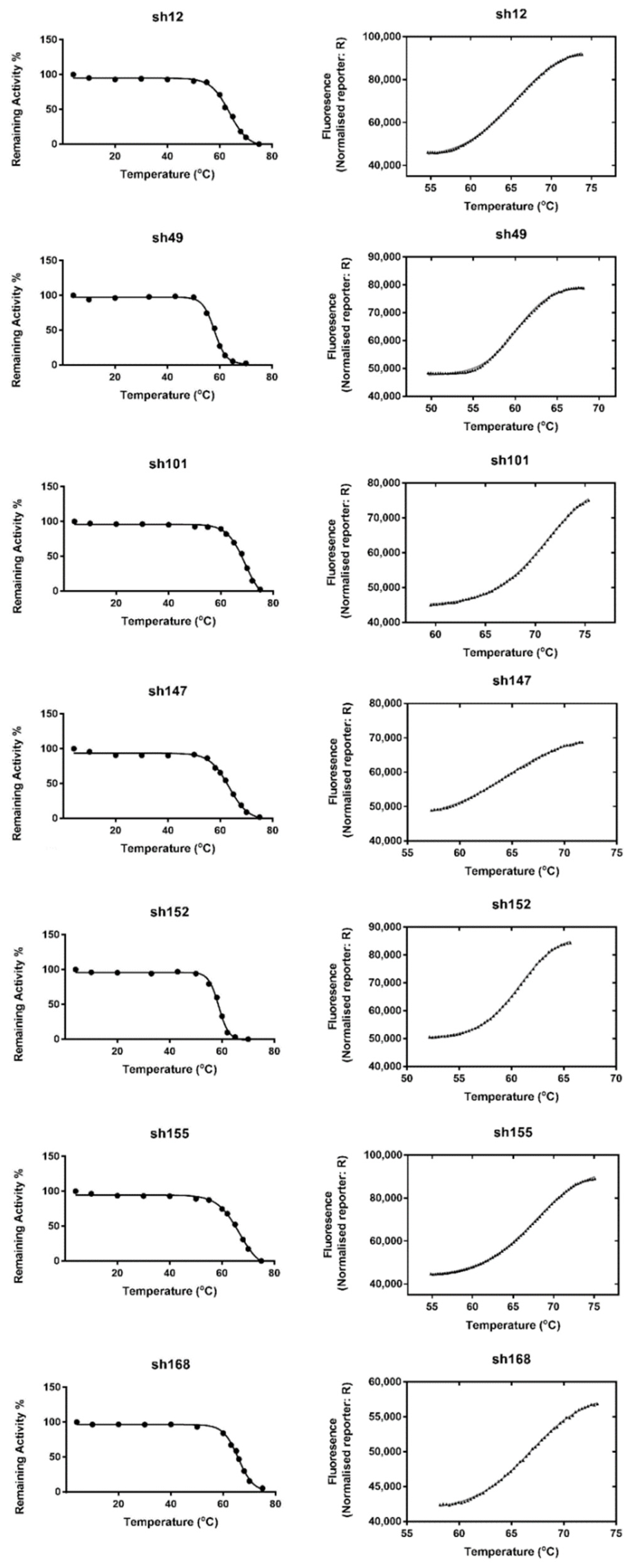
2.4. Evaluation of GSTF Thermal Stability
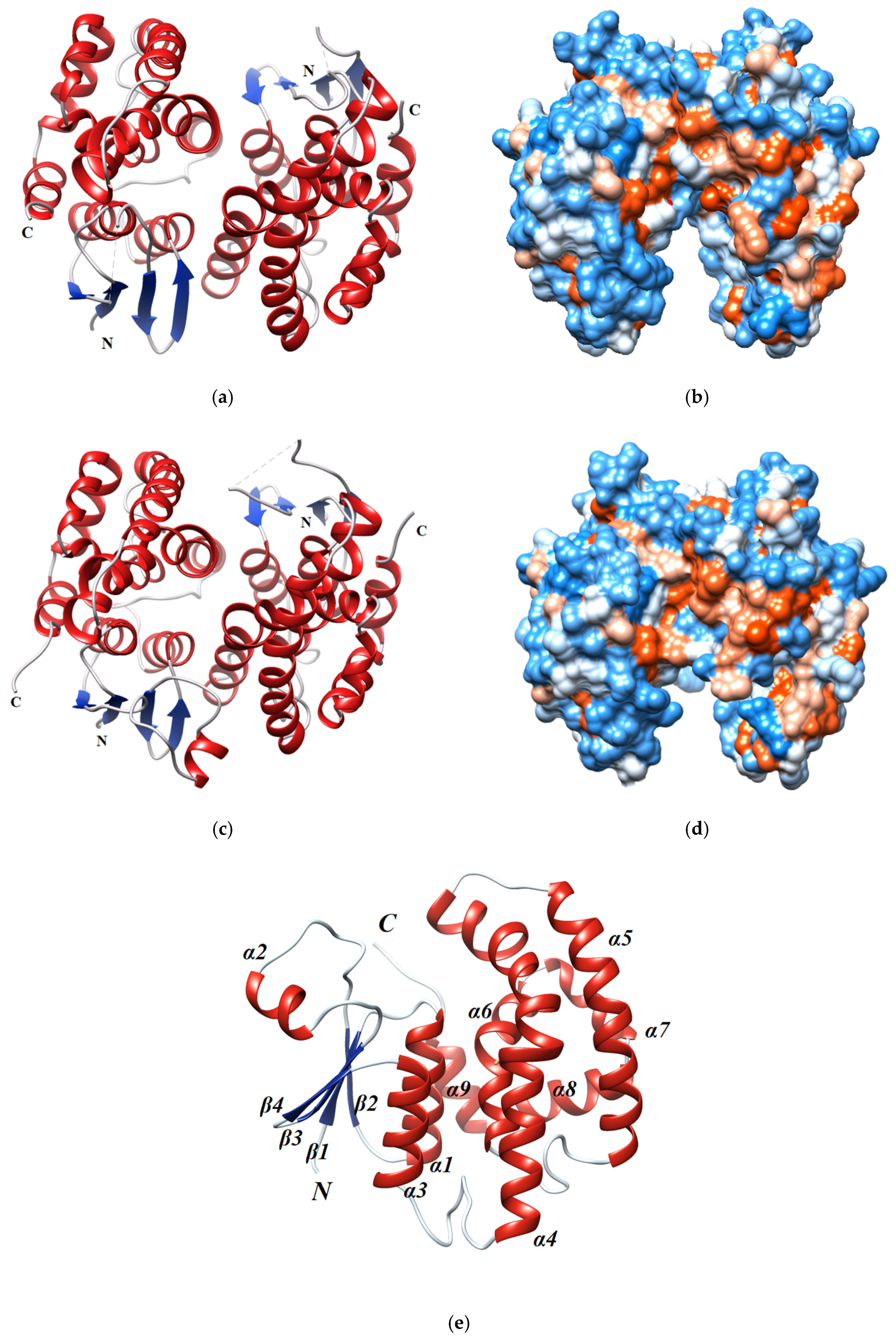
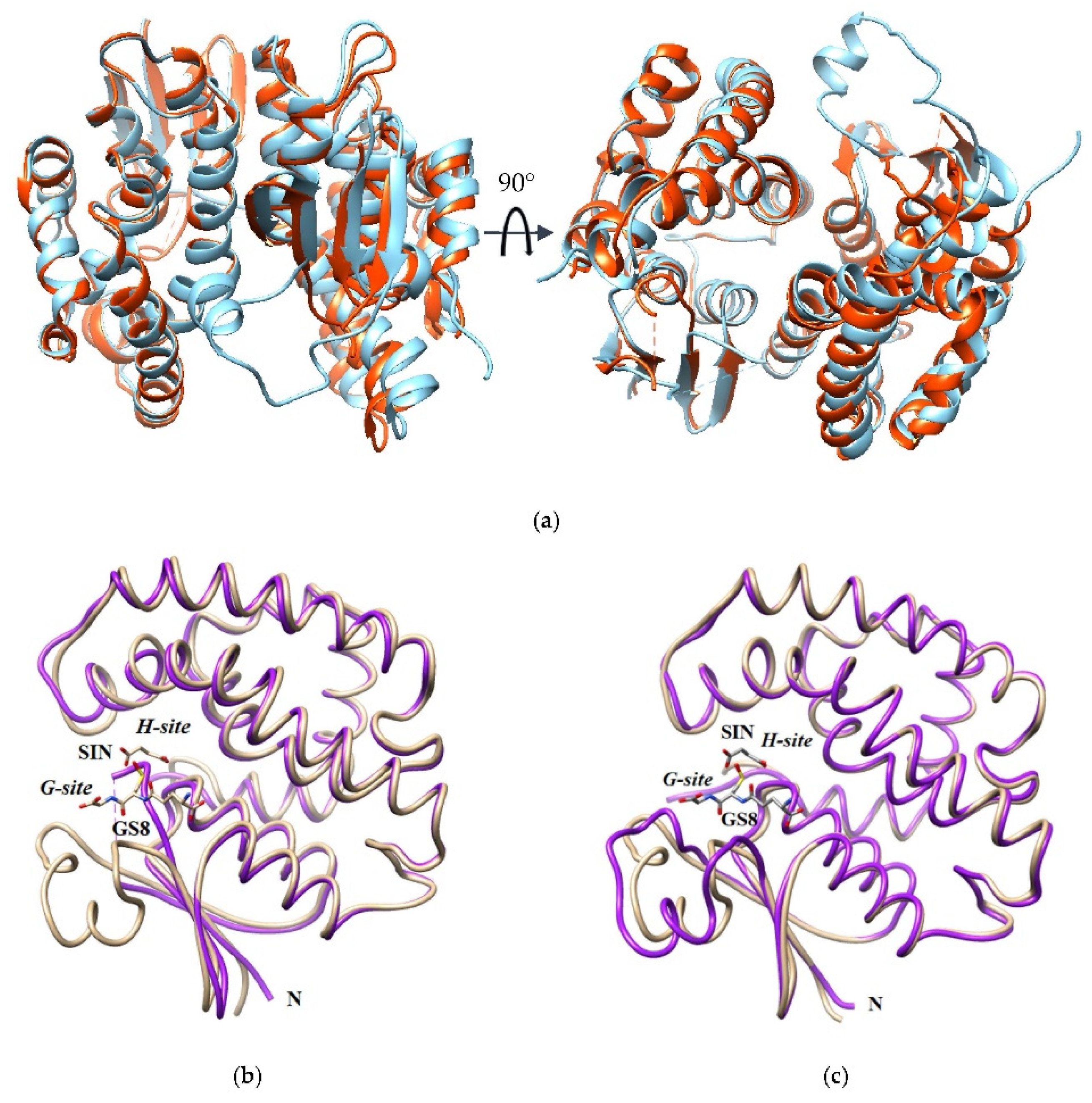
2.5. Overall Description of the Crystal Structure of sh101 and sh155 Enzyme Variants
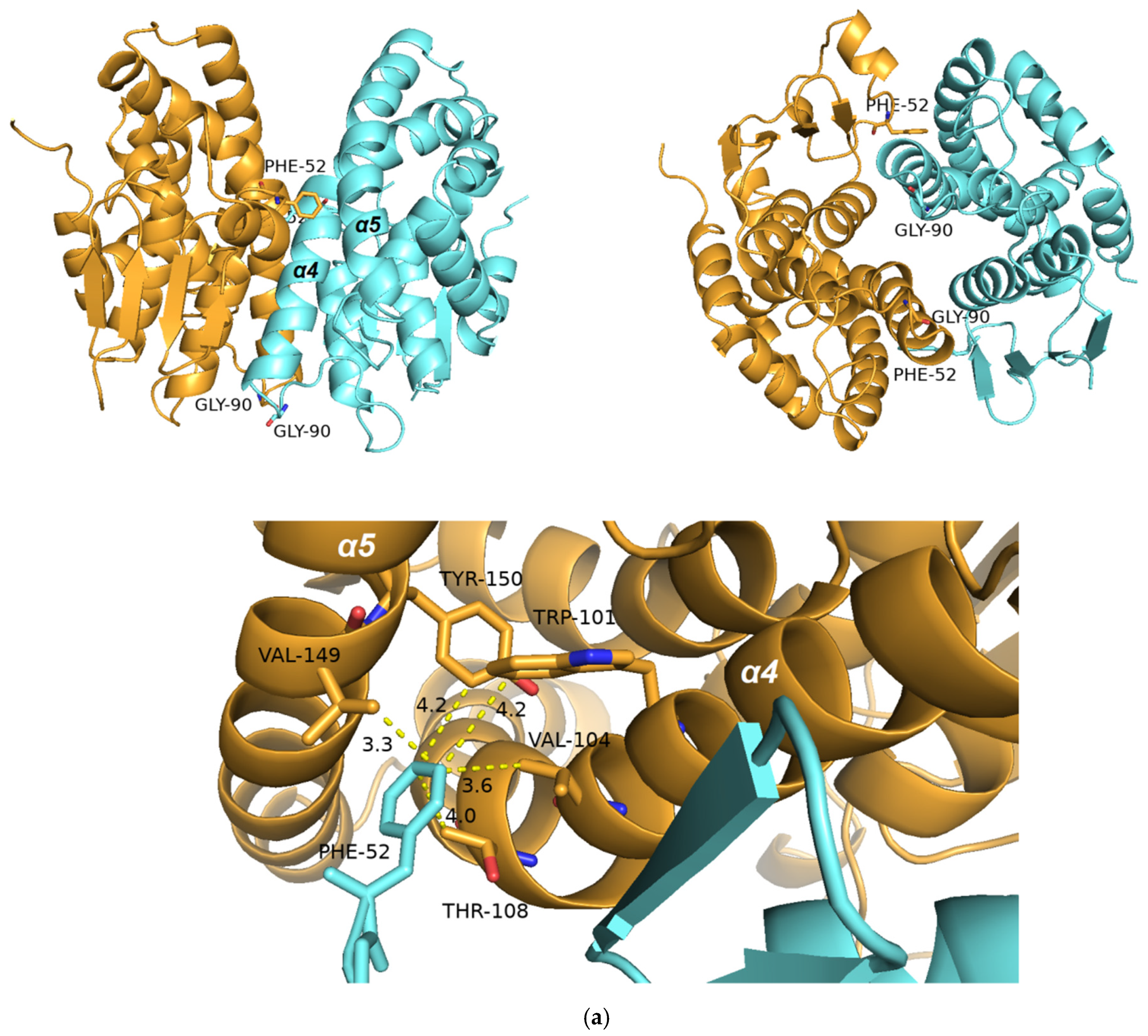
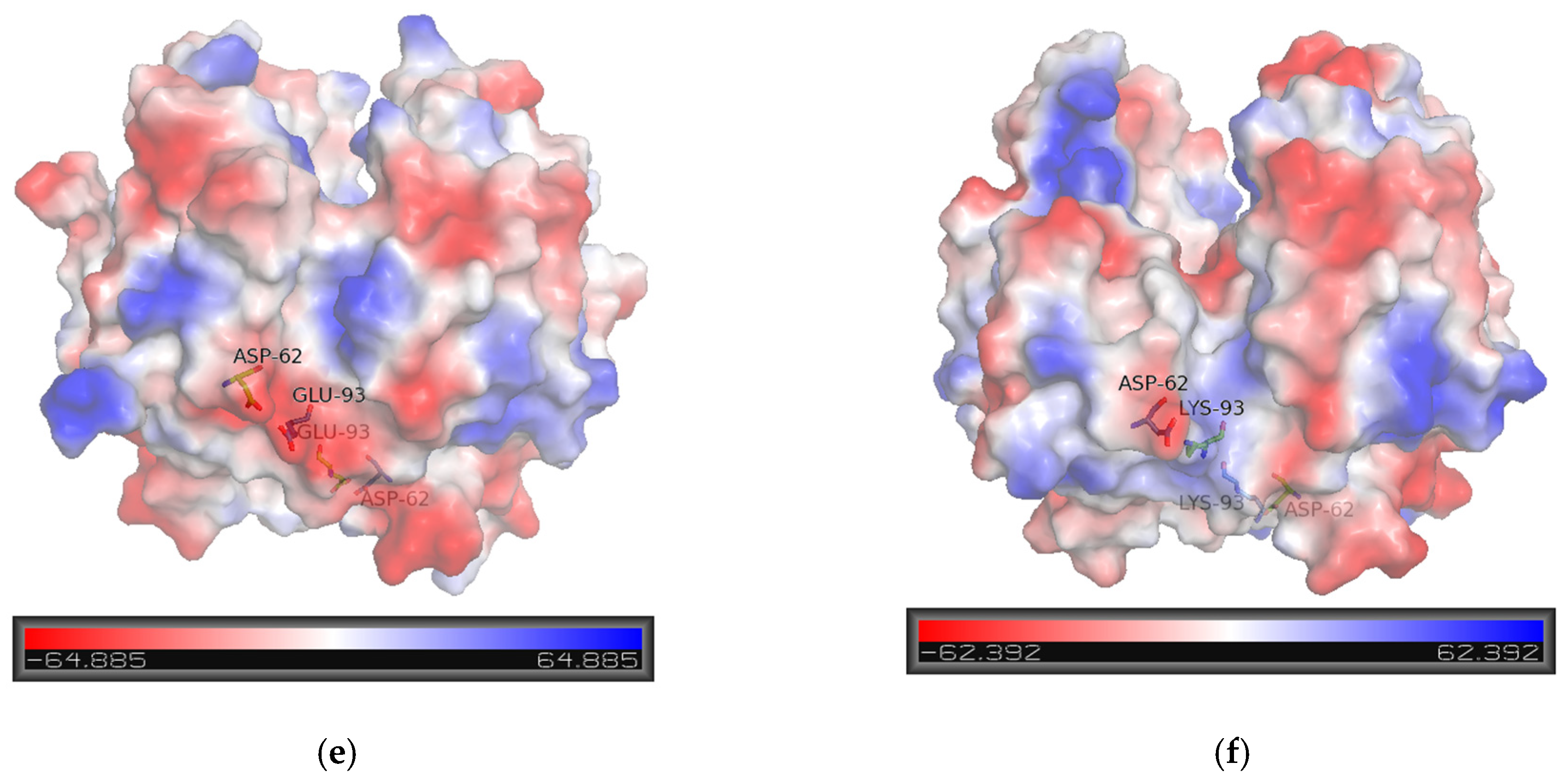
2.5.1. Structural Elements That Contribute to kcat Regulation

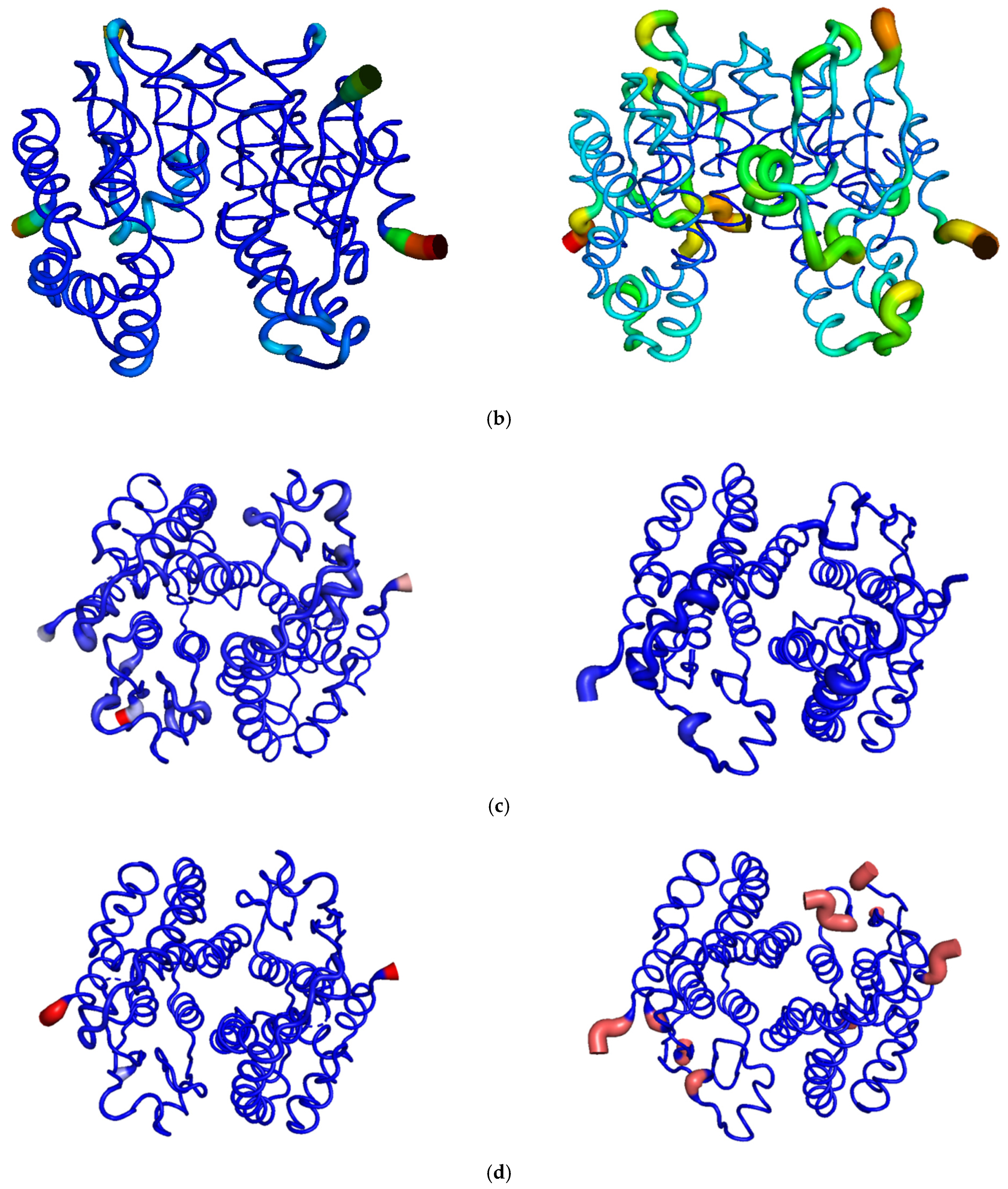
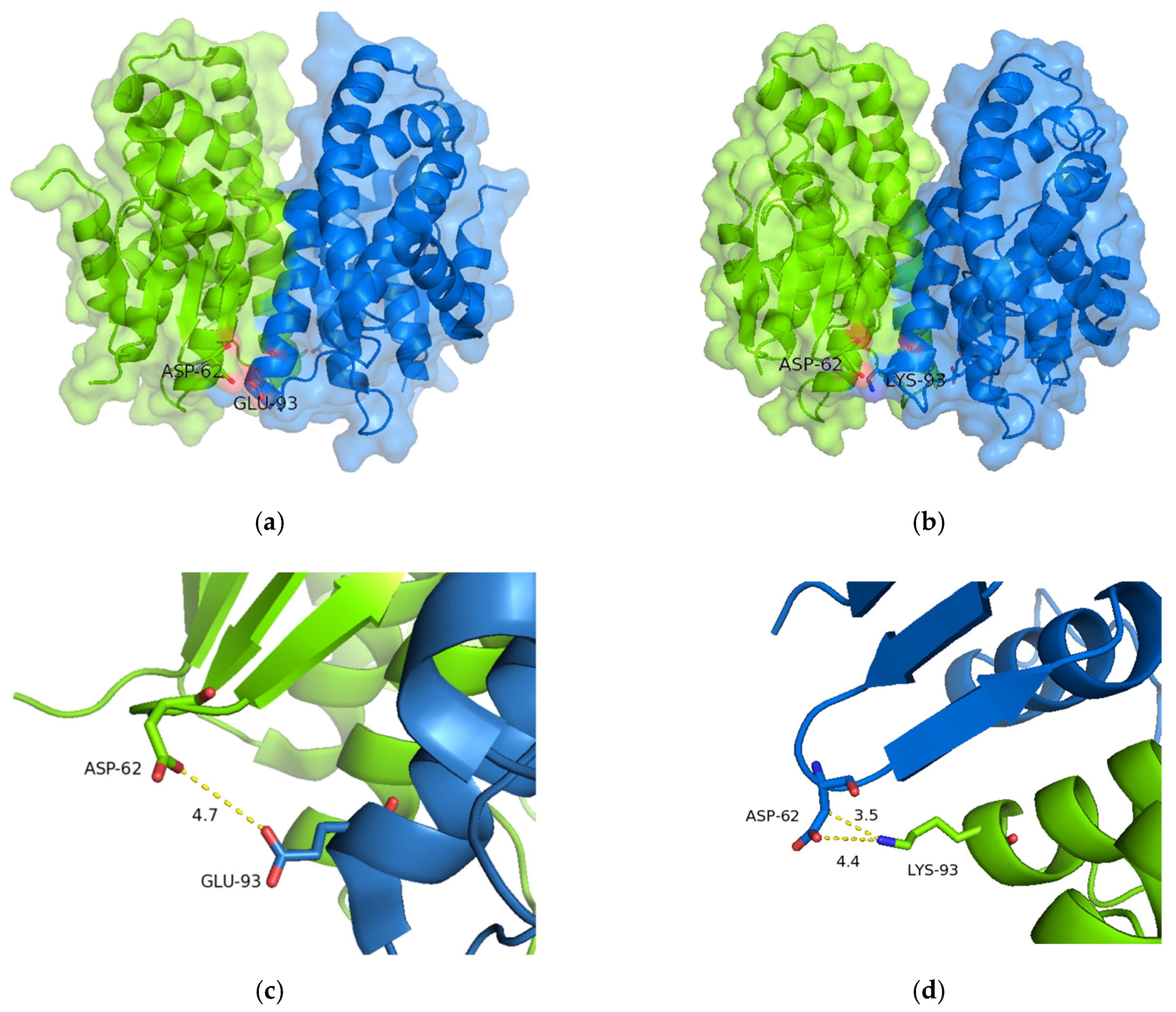
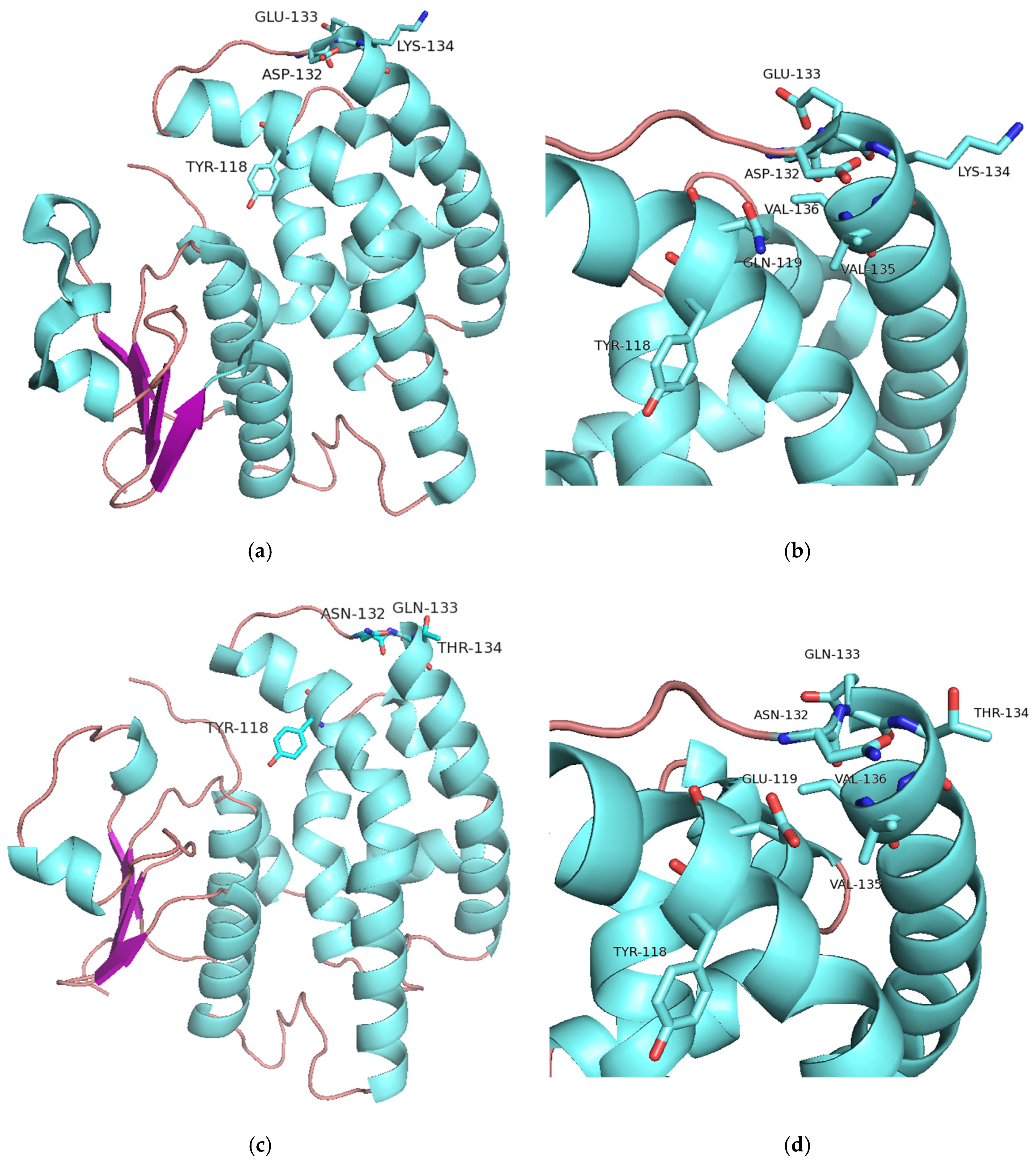
2.5.2. Structural Elements That Contribute to Thermostability
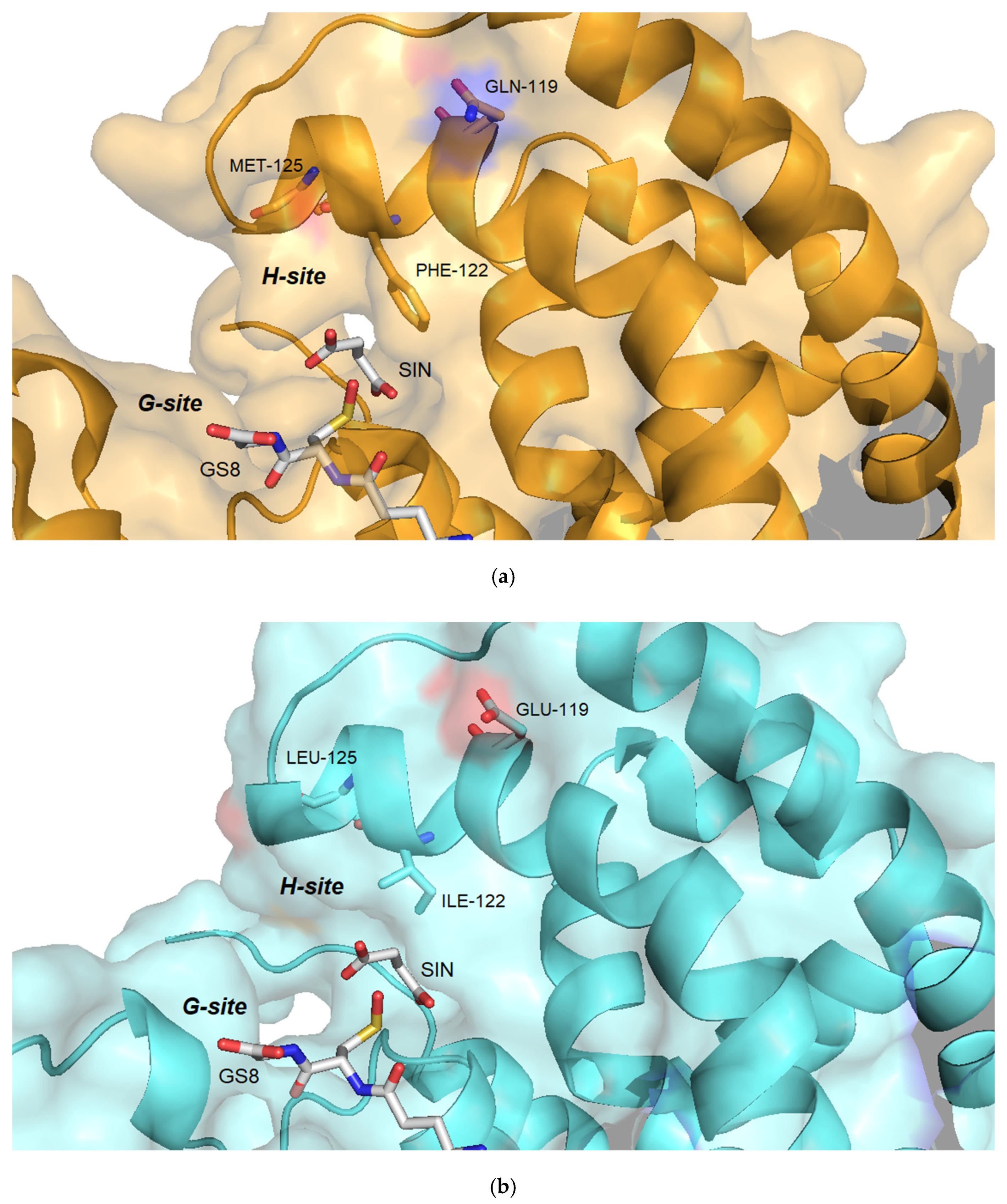
2.5.3. Structural Elements That Contribute to Inhibition Potency
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Preparation of DNA Shuffling and Construction of GSTF Library
3.2.2. GSTF Expression and Purification Methods
3.2.3. Enzyme Activity and Kinetic Analysis Assays
3.2.4. Protein Thermal Stability: T50 and Tm Determination
3.2.5. Crystallization and Structure Determination
3.2.6. Structural Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Conte, S.S.; Lloyd, A.M. Exploring multiple drug and herbicide resistance in plants—Spotlight on transporter proteins. Plant Sci. 2011, 180, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Tétard-Jones, C.; Sabbadin, F.; Moss, S.; Hull, R.; Neve, P.; Edwards, R. Changes in the proteome of the problem weed blackgrass correlating with multiple-herbicide resistance. Plant J. 2018, 94, 709–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matzrafi, M.; Gerson, O.; Rubin, B.; Peleg, Z. Different Mutations Endowing Resistance to Acetyl-CoA Carboxylase Inhibitors Results in Changes in Ecological Fitness of Lolium rigidum Populations. Front. Plant Sci. 2017, 8, 1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthimidou, E.; Ntoanidou, S.; Madesis, P.; Eleftherohorinos, I. Mechanisms of Lolium rigidum multiple resistance to ALS- and ACCase-inhibiting herbicides and their impact on plant fitness. Pestic. Biochem. Physiol. 2019, 164, 65–72. [Google Scholar] [CrossRef]
- Perotti, V.E.; Larran, A.S.; Palmieri, V.E.; Martinatto, A.K.; Permingeat, H.R. Herbicide resistant weeds: A call to integrate conventional agricultural practices, molecular biology knowledge and new technologies. Plant Sci. 2020, 290, 110255. [Google Scholar] [CrossRef]
- Lombardo, L.; Coppola, G.; Zelasco, S. New Technologies for Insect-Resistant and Herbicide-Tolerant Plants. Trends Biotechnol. 2016, 34, 49–57. [Google Scholar] [CrossRef]
- Délye, C.; Jasieniuk, M.; Le Corre, V. Deciphering the evolution of herbicide resistance in weeds. Trends Genet. 2013, 29, 649–658. [Google Scholar] [CrossRef]
- Heap, I. The International Herbicide-Resistant Weed Database. 2022. Available online: http://www.weedscience.com/ (accessed on 22 March 2022).
- Powles, S.B.; Yu, Q. Evolution in Action: Plants Resistant to Herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [Green Version]
- Peterson, M.A.; Collavo, A.; Ovejero, R.; Shivrain, V.; Walsh, M.J. The challenge of herbicide resistance around the world: A current summary. Pest Manag. Sci. 2018, 74, 2246–2259. [Google Scholar] [CrossRef]
- Cummins, I.; Wortley, D.J.; Sabbadin, F.; He, Z.; Coxon, C.R.; Straker, H.E.; Sellars, J.D.; Knight, K.; Edwards, L.; Hughes, D.; et al. Key role for a glutathione transferase in multiple-herbicide resistance in grass weeds. Proc. Natl. Acad. Sci. USA 2013, 110, 5812–5817. [Google Scholar] [CrossRef] [Green Version]
- Nakka, S.; Jugulam, M.; Peterson, D.; Asif, M. Herbicide resistance: Development of wheat production systems and current status of resistant weeds in wheat cropping systems. Crop J. 2019, 7, 750–760. [Google Scholar] [CrossRef]
- Han, H.; Yu, Q.; Vila-Aiub, M.; Powles, S. Genetic inheritance of cytochrome P450-mediated metabolic resistance to chlorsulfuron in a multiple herbicide resistant Lolium rigidum population. Crop Prot. 2014, 65, 57–63. [Google Scholar] [CrossRef]
- Labrou, N.E.; Papageorgiou, A.C.; Pavli, O.; Flemetakis, E. Plant GSTome: Structure and functional role in xenome network and plant stress response. Curr. Opin. Biotechnol. 2015, 32, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.; McEwen, A.G.; Lapthorn, A.; Edwards, R. Forced Evolution of a Herbicide Detoxifying Glutathione Transferase. J. Biol. Chem. 2003, 278, 23930–23935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pégeot, H.; Mathiot, S.; Perrot, T.; Gense, F.; Hecker, A.; Didierjean, C.; Rouhier, N. Structural plasticity among glutathione transferase Phi members: Natural combination of catalytic residues confers dual biochemical activities. FEBS J. 2017, 284, 2442–2463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrs, K.A. The Functions and Regulation of Glutathione S-Transferases in Plants. Annu. Rev. Plant Biol. 1996, 47, 127–158. [Google Scholar] [CrossRef] [PubMed]
- Hasan, S.; Islam, S.; Hasan, N.; Das Sajib, S.; Ahmed, S.; Islam, T.; Ghosh, A. Genome-wide analysis and transcript profiling identify several abiotic and biotic stress-responsive Glutathione S-transferase genes in soybean. Plant Gene 2020, 23, 100239. [Google Scholar] [CrossRef]
- Hasan, S.; Singh, V.; Islam, S.; Islam, S.; Ahsan, R.; Kaundal, A.; Islam, T.; Ghosh, A. Genome-wide identification and expression profiling of glutathione S-transferase family under multiple abiotic and biotic stresses in Medicago truncatula L. PLoS ONE 2021, 16, e0247170. [Google Scholar] [CrossRef]
- Davies, J.; Caseley, J.C. Herbicide safeners: A review. Pestic. Sci. 1999, 55, 1043–1058. [Google Scholar] [CrossRef]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant glutathione transferases. Genome Biol. 2002, 3, reviews3004.1. [Google Scholar] [CrossRef] [Green Version]
- Shimabukuro, R.H.; Frear, D.S.; Swanson, H.R.; Walsh, W.C. Glutathione Conjugation. Plant Physiol. 1971, 47, 10–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, I.; Moss, S.; Cole, D.J.; Edwards, R. Glutathione transferases in herbicide-resistant and herbicide-susceptible black-grass (Alopecurus myosuroides). Pestic. Sci. 1997, 51, 244–250. [Google Scholar] [CrossRef]
- Cummins, I.; Cole, D.J.; Edwards, R. A role for glutathione transferases functioning as glutathione peroxidases in resistance to multiple herbicides in black-grass. Plant J. 1999, 18, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Hatton, P.J.; Cummins, I.; Price, L.J.; Cole, D.J.; Edwards, R. Glutathione transferases and herbicide detoxification in suspension-cultured cells of giant foxtail (Setaria faberi). Pestic. Sci. 1998, 53, 209–216. [Google Scholar] [CrossRef]
- Letouzé, A.; Gasquez, J. Enhanced activity of several herbicide-degrading enzymes: A suggested mechanism responsible for multiple resistance in blackgrass (Alopecurus myosuroides Huds.). Agronomie 2003, 23, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Gaines, T.A.; Lorentz, L.; Figge, A.; Herrmann, J.; Maiwald, F.; Ott, M.; Han, H.; Busi, R.; Yu, Q.; Powles, S.B.; et al. RNA-Seq transcriptome analysis to identify genes involved in metabolism-based diclofop resistance in Lolium rigidum. Plant J. 2014, 78, 865–876. [Google Scholar] [CrossRef]
- Gardin, J.A.C.; Gouzy, J.; Carrère, S.; Délye, C. ALOMYbase, a resource to investigate non-target-site-based resistance to herbicides inhibiting acetolactate-synthase (ALS) in the major grass weed Alopecurus myosuroides (black-grass). BMC Genom. 2015, 16, 590. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.; Dixon, D.P. The role of glutathione transferases in herbicide metabolism. In Herbicides and Their Mechanisms of Action; Cobb, A.H., Kirkwood, R.C., Eds.; Academic Press: Sheffield, UK, 2000; pp. 38–71. [Google Scholar]
- Georgakis, N.; Poudel, N.; Papageorgiou, A.; Labrou, N.E. Comparative structural and functional analysis of phi class glutathione transferases involved in multiple-herbicide resistance of grass weeds and crops. Plant Physiol. Biochem. 2020, 149, 266–276. [Google Scholar] [CrossRef]
- Georgakis, N.; Poudel, N.; Vlachakis, D.; Papageorgiou, A.C.; Labrou, N.E. Phi class glutathione transferases as molecular targets towards multiple-herbicide resistance: Inhibition analysis and pharmacophore design. Plant Physiol. Biochem. 2020, 158, 342–352. [Google Scholar] [CrossRef]
- Dücker, R.; Parcharidou, E.; Beffa, R. Flufenacet activity is affected by GST inhibitors in blackgrass (Alopecurus myosuroides) populations with reduced flufenacet sensitivity and higher expression levels of GSTs. Weed Sci. 2020, 68, 451–459. [Google Scholar] [CrossRef]
- Schwarz, M.; Eno, R.F.M.; Freitag-Pohl, S.; Coxon, C.R.; Straker, H.E.; Wortley, D.J.; Hughes, D.J.; Mitchell, G.; Moore, J.; Cummins, I.; et al. Flavonoid-based inhibitors of the Phi-class glutathione transferase from black-grass to combat multiple herbicide resistance. Org. Biomol. Chem. 2021, 19, 9211–9222. [Google Scholar] [CrossRef] [PubMed]
- Dücker, R.; Zöllner, P.; Lümmen, P.; Ries, S.; Collavo, A.; Beffa, R. Glutathione transferase plays a major role in flufenacet resistance of ryegrass (Lolium spp.) field populations. Pest Manag. Sci. 2019, 75, 3084–3092. [Google Scholar] [CrossRef]
- Dücker, R.; Zöllner, P.; Parcharidou, E.; Ries, S.; Lorentz, L.; Beffa, R. Enhanced metabolism causes reduced flufenacet sensitivity in black-grass (Alopecurus myosuroides Huds.) field populations. Pest Manag. Sci. 2019, 75, 2996–3004. [Google Scholar] [CrossRef] [PubMed]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, Structural and Biochemical Features of Plant Serinyl-Glutathione Transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef] [PubMed]
- Gulati, K.; Poluri, K.M.; Gualti, K. An Overview of Computational and Experimental Methods for Designing Novel Proteins. Recent Patents Biotechnol. 2016, 10, 235–263. [Google Scholar] [CrossRef]
- FPerperopoulou, F.; Pouliou, F.; Labrou, N.E. Recent advances in protein engineering and biotechnological applications of glutathione transferases. Crit. Rev. Biotechnol. 2017, 38, 511–528. [Google Scholar] [CrossRef]
- Axarli, I.; Muleta, A.W.; Vlachakis, D.; Kossida, S.; Kotzia, G.; Maltezos, A.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Directed evolution of Tau class glutathione transferases reveals a site that regulates catalytic efficiency and masks co-operativity. Biochem. J. 2016, 473, 559–570. [Google Scholar] [CrossRef]
- Axarli, I.; Muleta, A.W.; Chronopoulou, E.G.; Papageorgiou, A.C.; Labrou, N.E. Directed evolution of glutathione transferases towards a selective glutathione-binding site and improved oxidative stability. Biochim. Biophys. Acta 2017, 1861, 3416–3428. [Google Scholar] [CrossRef]
- Chronopoulou, E.G.; Papageorgiou, A.C.; Ataya, F.; Nianiou-Obeidat, I.; Madesis, P.; Labrou, N.E. Expanding the Plant GSTome Through Directed Evolution: DNA Shuffling for the Generation of New Synthetic Enzymes with Engineered Catalytic and Binding Properties. Front. Plant Sci. 2018, 9, 1737. [Google Scholar] [CrossRef]
- Perperopoulou, F.; Fragoulaki, M.; Papageorgiou, A.; Labrou, N. Directed Evolution of a Glutathione Transferase for the Development of a Biosensor for Alachlor Determination. Symmetry 2021, 13, 461. [Google Scholar] [CrossRef]
- Traut, T. Allosteric Regulatory Enzymes; Springer: Boston, MA, USA, 2008. [Google Scholar]
- Chronopoulou, E.; Madesis, P.; Asimakopoulou, B.; Platis, D.; Tsaftaris, A.; Labrou, N.E. Catalytic and structural diversity of the fluazifop-inducible glutathione transferases from Phaseolus vulgaris. Planta 2011, 235, 1253–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cummins, I.; O’Hagan, D.; Jablonkai, I.; Cole, D.J.; Hehn, A.; Werck-Reichhart, D.; Edwards, R. Cloning, characterization and regulation of a family of phi class glutathione transferases from wheat. Plant Mol. Biol. 2003, 52, 591–603. [Google Scholar] [CrossRef]
- Cho, H.-Y.; Kong, K.-H. Molecular cloning, expression, and characterization of a phi-type glutathione S-transferase from Oryza sativa. Pestic. Biochem. Physiol. 2005, 83, 29–36. [Google Scholar] [CrossRef]
- Jo, H.J.; Kong, J.N.; Lim, J.K.; Kong, K.H. Site-directed mutagenesis of evolutionarily conserved serine residues in the N-terminal domain of rice Phi-class glutathione S-transferase F5. J. Mol. Catal. B Enzym. 2014, 106, 71–75. [Google Scholar] [CrossRef]
- Pégeot, H.; Koh, C.S.; Petre, B.; Mathiot, S.; Duplessis, S.; Hecker, A.; Didierjean, C.; Rouhier, N.; Pégeot, H. The poplar Phi class glutathione transferase: Expression, activity and structure of GSTF1. Front. Plant Sci. 2014, 5, 712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Georgakis, N.; Ioannou, E.; Varotsou, C.; Premetis, G.; Chronopoulou, E.G.; Labrou, N.E. Determination of half-maximal inhibitory concentration of an enzyme inhibitor. In Targeting Enzymes for Pharmaceutical Development, Methods in Molecular Biology; Labrou, N., Ed.; Humana: New York, NY, USA, 2020; Volume 2089, pp. 41–46. [Google Scholar] [CrossRef]
- Axarli, I.; Georgiadou, C.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Investigation of the role of conserved residues Ser13, Asn48 and Pro49 in the catalytic mechanism of the tau class glutathione transferase from Glycine max. Biochim. Biophys. Acta 2010, 1804, 662–667. [Google Scholar] [CrossRef]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.G.; Pavli, O.; Flemetakis, E.; Skaracis, G.N.; Labrou, N.E. Characterization and functional analysis of a recombinant tau class glutathione transferase GmGSTU2-2 from Glycine max. Int. J. Biol. Macromol. 2016, 94, 802–812. [Google Scholar] [CrossRef]
- Wei, T.; Wang, C.-L.; Kao, H.-N.; Yang, H.-L.; Wang, X.-R.; Zeng, Q.-Y. Molecular and catalytic characterization of a phi class glutathione transferase from Cathaya argyrophylla. Biochem. Syst. Ecol. 2012, 40, 75–85. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinemer, P.; Prade, L.; Hof, P.; Neuefeind, T.; Huber, R.; Zettl, R.; Palme, K.; Schell, J.; Koelln, I.; Bartunik, H.D.; et al. Three-dimensional structure of glutathione S-transferase from Arabidopsis thaliana at 2.2 Å resolution: Structural characterization of herbicide-conjugating plant glutathione S-transferases and a novel active site architecture. J. Mol. Biol. 1996, 255, 289–309. [Google Scholar] [CrossRef] [PubMed]
- Thom, R.; Cummins, I.; Dixon, D.P.; Edwards, R.; Cole, A.D.J.; Lapthorn, A.J. Structure of a Tau Class Glutathione S-Transferase from Wheat Active in Herbicide Detoxification. Biochemistry 2002, 41, 7008–7020. [Google Scholar] [CrossRef]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.; Labrou, N.E. Catalytic features and crystal structure of a tau class glutathione transferase from Glycine max specifically upregulated in response to soybean mosaic virus infections. Biochim. Biophys. Acta (BBA) 2015, 1854, 166–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labrou, N.E.; Mello, L.V.; Clonis, Y.D. Functional and structural roles of the glutathione-binding residues in maize (Zea mays) glutathione S-transferase I. Biochem. J. 2001, 358, 101–110. [Google Scholar] [CrossRef]
- Ricci, G.; Caccuri, A.M.; Bello, M.L.; Rosato, N.; Mei, G.; Nicotra, M.; Chiessi, E.; Mazzetti, A.P.; Federici, G. Structural flexibility modulates the activity of human glutathione transferase P1-1. Role of helix 2 flexibility in the catalytic mechanism. J. Biol. Chem. 1996, 271, 16187–16192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger, L.; DeLano, W. The PyMOL Molecular Graphics System, Version 2.0; Schrödinger LLC: New York, NY, USA, 2020; Available online: http://www.pymol.org/pymol (accessed on 22 March 2022).
- Rodrigues, C.H.M.; Pires, D.E.V.; Ascher, D.B. DynaMut: Predicting the impact of mutations on protein conformation, flexibility and stability. Nucleic Acids Res. 2018, 46, W350–W355. [Google Scholar] [CrossRef] [PubMed]
- Stemmer, W.P. DNA shuffling by random fragmentation and reassembly: In vitro recombination for molecular evolution. Proc. Natl. Acad. Sci. USA 1994, 91, 10747–10751. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H. Optimization of DNA shuffling for high fidelity recombination. Nucleic Acids Res. 1997, 25, 1307–1308. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.J.; Ellefson, J.W.; Ellington, A.D. Library Generation by Gene Shuffling. Curr. Protoc. Mol. Biol. 2014, 105, 15.12.1–15.12.7. [Google Scholar] [CrossRef] [Green Version]
- Georgakis, N.D.; Karagiannopoulos, D.A.; Thireou, T.N.; Eliopoulos, E.E.; Labrou, N.E.; Tsoungas, P.G.; Koutsilieris, M.N.; Clonis, Y.D. Concluding the trilogy: The interaction of 2,2′-dihydroxy-benzophenones and their carbonyl N-analogues with human glutathione transferase M1-1 face to face with the P1-1 and A1-1 isoenzymes involved in MDR. Chem. Biol. Drug Des. 2017, 90, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Axarli, I.; Labrou, N.; Petrou, C.; Rassias, N.; Cordopatis, P.; Clonis, Y. Sulphonamide-based bombesin prodrug analogues for glutathione transferase, useful in targeted cancer chemotherapy. Eur. J. Med. Chem. 2009, 44, 2009–2016. [Google Scholar] [CrossRef] [PubMed]
- Skopelitou, K.; Muleta, A.W.; Pavli, O.; Skaracis, G.N.; Flemetakis, E.; Papageorgiou, A.C.; Labrou, N.E. Overlapping protective roles for glutathione transferase gene family members in chemical and oxidative stress response in Agrobacterium tumefaciens. Funct. Integr. Genom. 2011, 12, 157–172. [Google Scholar] [CrossRef]
- Lawrence, R.A.; Burk, R.F. Glutathione peroxidase activity in selenium-deficient rat liver. Biochem. Biophys. Res. Commun. 1976, 71, 952–958. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Huynh, K.; Partch, C.L. Analysis of Protein Stability and Ligand Interactions by Thermal Shift Assay. Curr. Protoc. Protein Sci. 2015, 79, 28.9.1–28.9.14. [Google Scholar] [CrossRef]
- McCoy, A.J. Solving structures of protein complexes by molecular replacement with Phaser. Acta Crystallogr. Sect. D Biol. Crystallogr. 2006, 63, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Km (mM) | S0.5 (mM) | nH | kcat (min−1) | kcat/Km (min−1 mM−1) | kcat/S0.5 (min−1 mM−1) | |
|---|---|---|---|---|---|---|
| GSH | CDNB | GSH | CDNB | |||
| AmGSTF | 1.78 ± 0.04 | 0.65 ± 0.03 | 1.5 ± 0.05 | 35.9 ± 1.7 | 20.17 | 55.23 |
| LrGSTF | 1.36 ± 0.04 | 0.76 ± 0.04 | 1.41 ± 0.05 | 28.35 ± 1.02 | 20.85 | 37.30 |
| HvGSTF | 0.8 ± 0.017 | 0.52 ± 0.01 | 1.46 ± 0.03 | 110 ± 3.05 | 137.5 | 211.54 |
| TdGSTF | 0.9 ± 0.03 | 0.51 ± 0.02 | 1.43 ± 0.06 | 141.3 ± 5.08 | 157 | 277.06 |
| sh12 | 1.4 ± 0.04 | 0.35 ± 0.01 | 1.16 ± 0.03 | 151.2 ± 1.9 | 108 | 432.00 |
| sh49 | 1.5 ± 0.05 | 0.61 ± 0.02 | 1.41 ± 0.05 | 34.8 ± 0.5 | 23.2 | 57.05 |
| sh101 | 1.37 ± 0.05 | 0.41 ± 0.02 | 1.1 ± 0.02 | 185.9 ± 1.8 | 135.69 | 453.4 |
| sh147 | 1.4 ± 0.03 | 0.46 ± 0.03 | 1.05 ± 0.03 | 149.6 ± 1.8 | 106.86 | 325.20 |
| sh155 | 1.43 ± 0.04 | 0.77 ± 0.06 | 1.02 ± 0.03 | 197.7 ± 3.18 | 138.25 | 256.75 |
| sh152 | 1.3 ± 0.05 | 0.38 ± 0.02 | 1.13 ± 0.04 | 140.4 ± 2.1 | 108.00 | 369.47 |
| sh168 | 1.7 ± 0.05 | 0.85 ± 0.04 | 1.28 ± 0.04 | 34.05 ± 0.9 | 20.03 | 40.10 |
| Parent GSTFs | |||||||
| TdGSTF | HvGSTF | LrGSTF | AmGSTF | ||||
| Butachlor (μM) | 9.9 ± 0.2 | 9.2 ± 0.6 | 7.1 ± 0.5 | 7.3 ± 0.4 | |||
| Shuffled enzyme variants | |||||||
| sh12 | sh49 | sh101 | sh147 | sh152 | sh155 | sh168 | |
| Butachlor (μM) | 6.8 ± 0.1 | 2.6 ± 0.04 | 5.9 ± 0.4 | 9.4 ± 0.7 | 4.4 ± 0.07 | 3.4 ± 0.1 | 3.1 ± 0.08 |
| T50 (°C) | Tm (°C) | Tm − T50 (°C) | |
|---|---|---|---|
| AmGSTF | 61.4 ± 0.2 | 62.8 ± 0.04 | 1.4 |
| TdGSTF | 69 ± 0.9 | 71.1 ± 0.05 | 2.1 |
| HvGSTF | 66.8 ± 0.2 | 69.5 ± 0.05 | 2.7 |
| LrGSTF | 67.5 ± 0.2 | 70.2 ± 0.04 | 2.7 |
| sh12 | 63.7 ± 0.2 | 65.4 ± 0.04 | 1.7 |
| sh49 | 58 ± 0.1 | 60.1 ± 0.04 | 2.1 |
| sh101 | 69.3 ± 0.3 | 71.1 ± 0.06 | 1.8 |
| sh147 | 63.3 ± 0.3 | 64.5 ± 0.05 | 1.2 |
| sh152 | 58.8 ± 0.1 | 60.8 ± 0.03 | 2 |
| sh155 | 66.6 ± 0.3 | 68.0 ± 0.05 | 1.4 |
| sh168 | 65.8 ± 0.2 | 66.8 ± 0.06 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioannou, E.; Papageorgiou, A.C.; Labrou, N.E. Directed Evolution of Phi Class Glutathione Transferases Involved in Multiple-Herbicide Resistance of Grass Weeds and Crops. Int. J. Mol. Sci. 2022, 23, 7469. https://doi.org/10.3390/ijms23137469
Ioannou E, Papageorgiou AC, Labrou NE. Directed Evolution of Phi Class Glutathione Transferases Involved in Multiple-Herbicide Resistance of Grass Weeds and Crops. International Journal of Molecular Sciences. 2022; 23(13):7469. https://doi.org/10.3390/ijms23137469
Chicago/Turabian StyleIoannou, Elisavet, Anastassios C. Papageorgiou, and Nikolaos E. Labrou. 2022. "Directed Evolution of Phi Class Glutathione Transferases Involved in Multiple-Herbicide Resistance of Grass Weeds and Crops" International Journal of Molecular Sciences 23, no. 13: 7469. https://doi.org/10.3390/ijms23137469