
Catalytic Antibodies in Bipolar Disorder: Serum IgGs Hydrolyze Myelin Basic Protein


Abstract
:1. Introduction
2. Results
2.1. Characterization of Patients and Healthy Donors
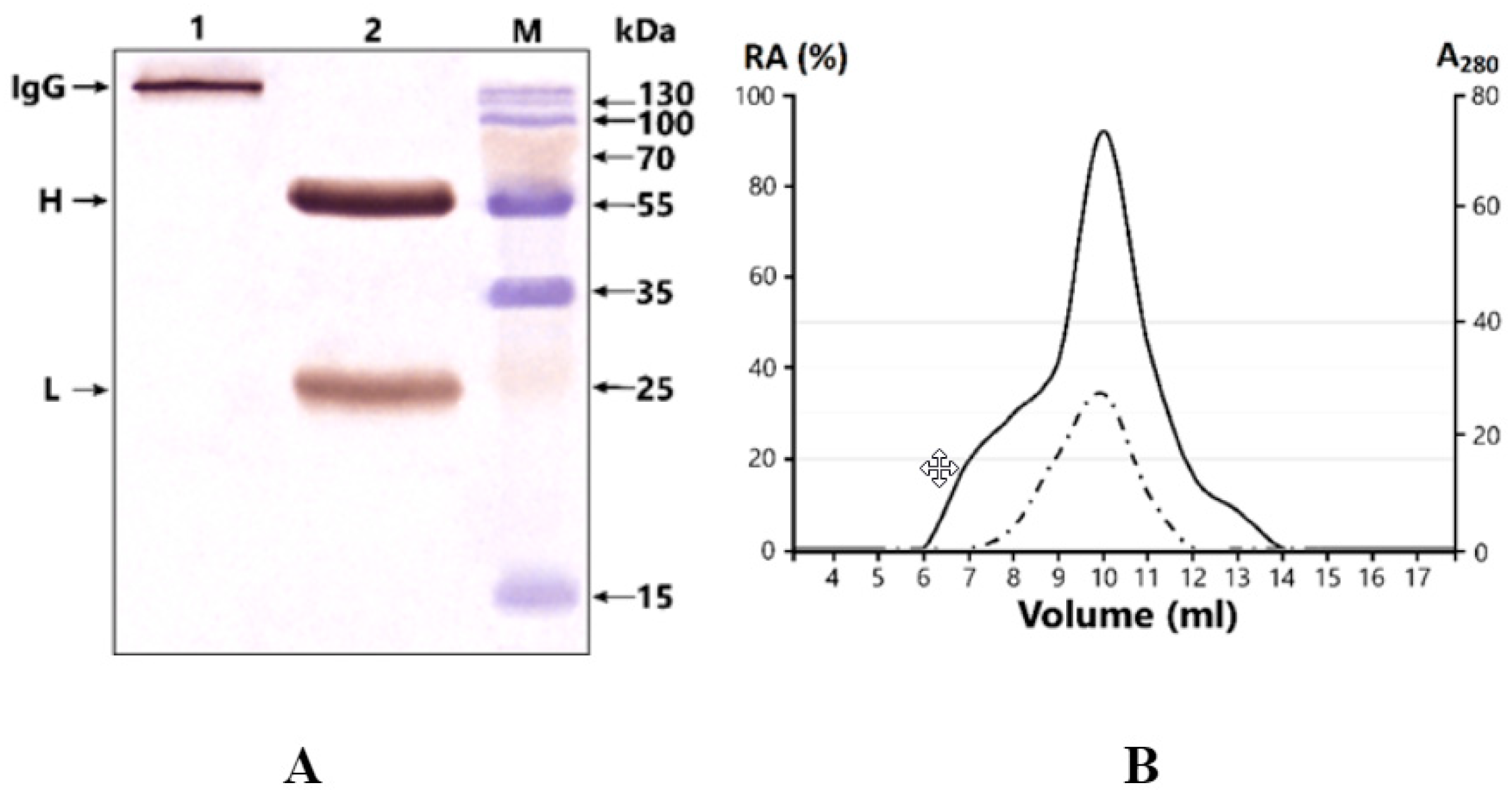
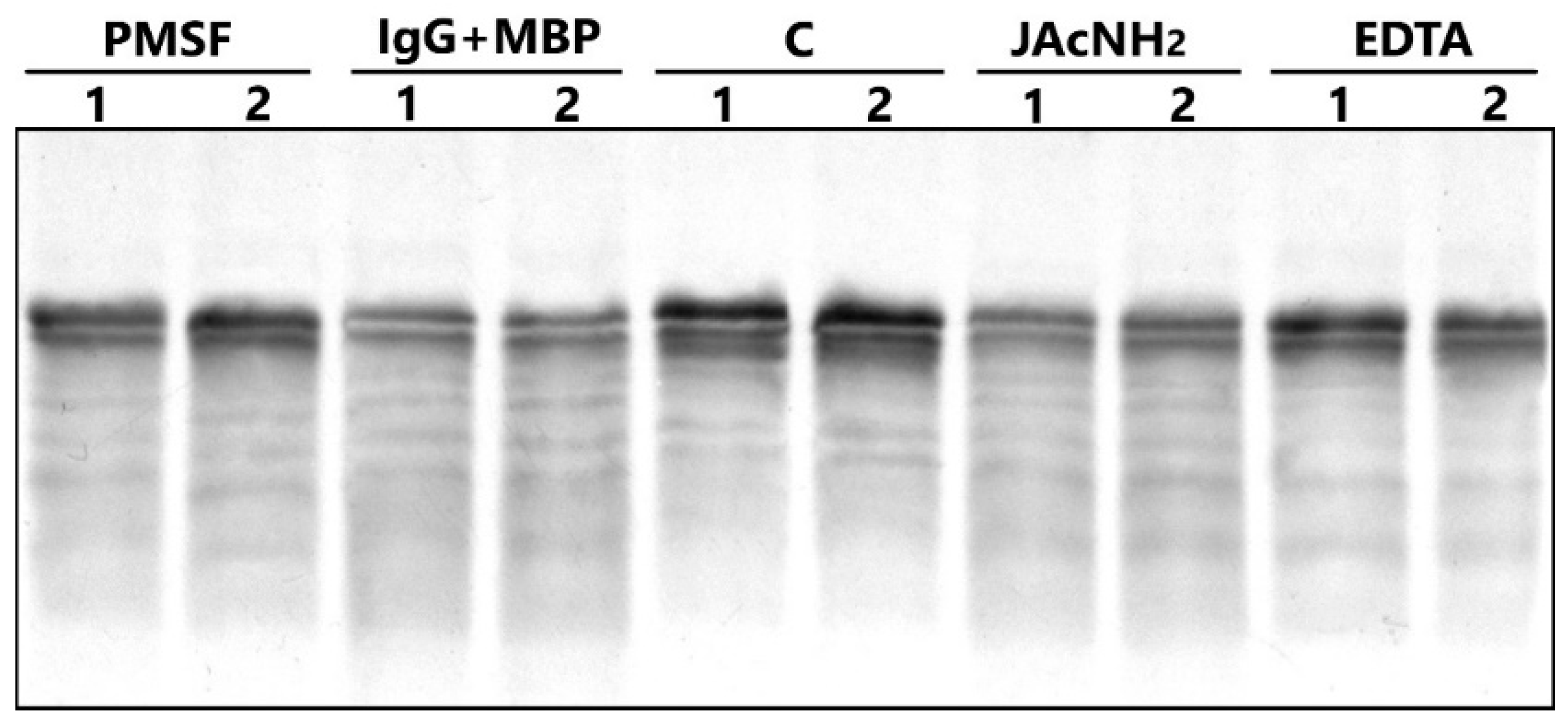
2.2. Antibody Purification and Application of Strict Criteria for IgGs with Proteolytic Activity
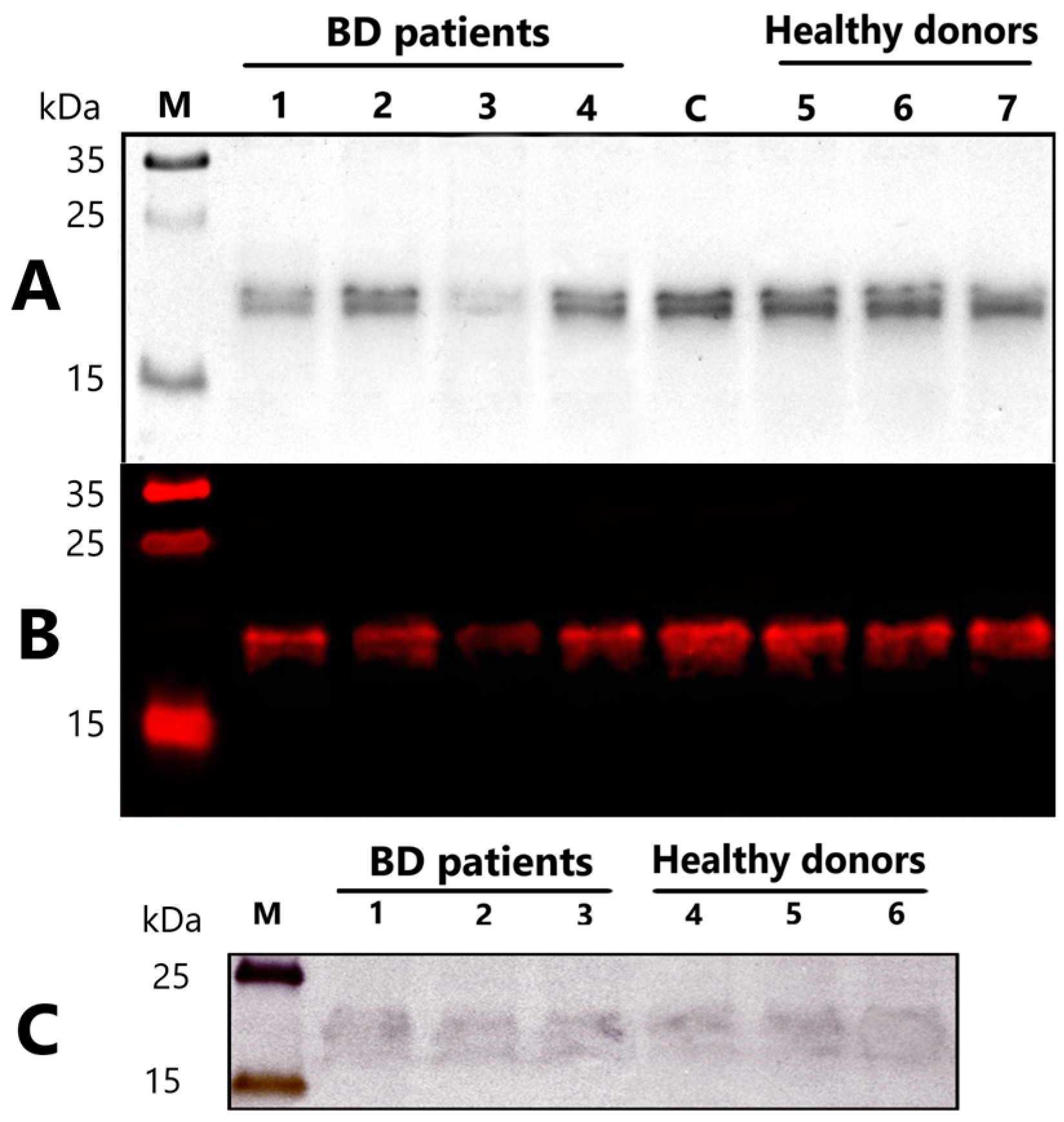
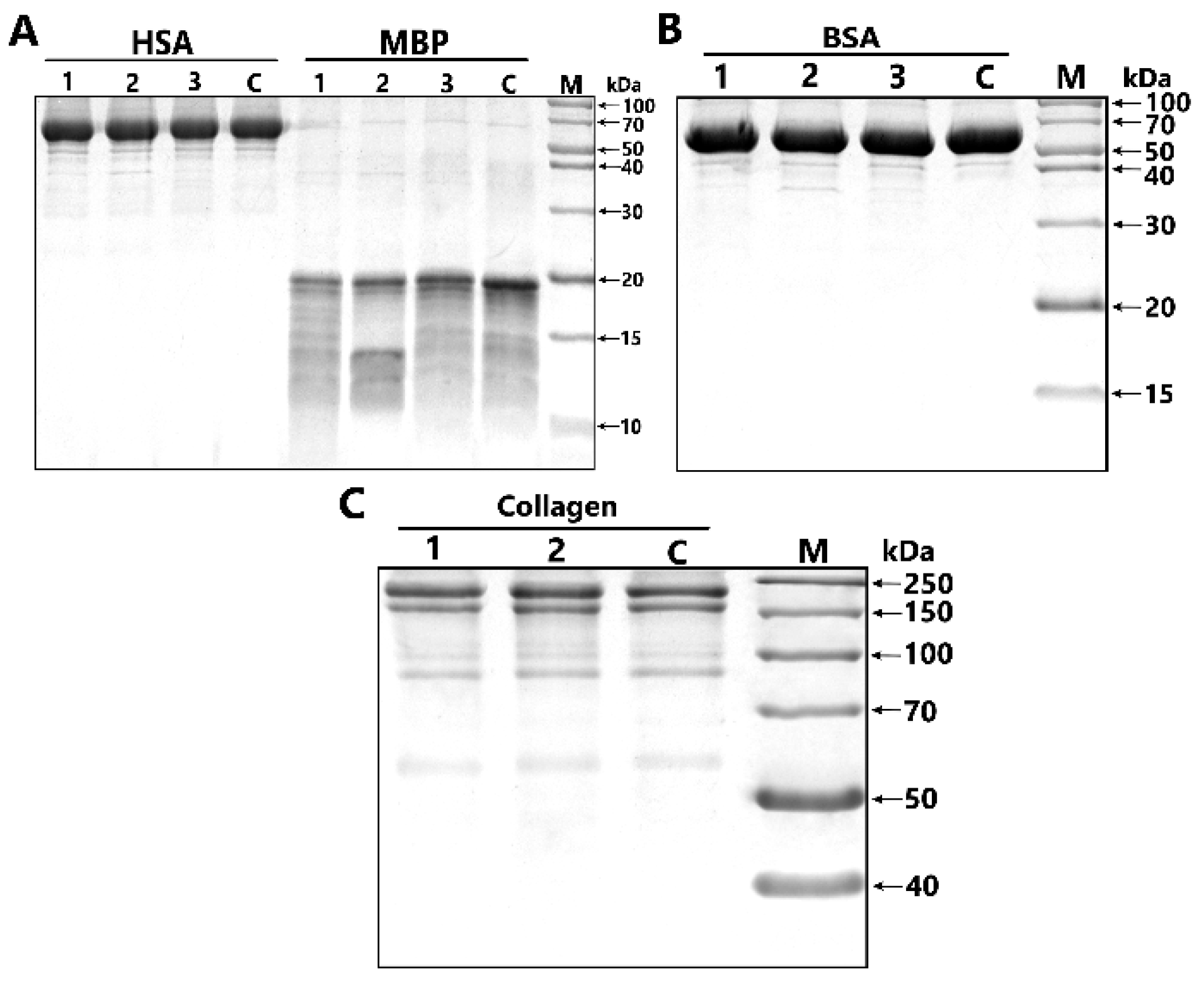
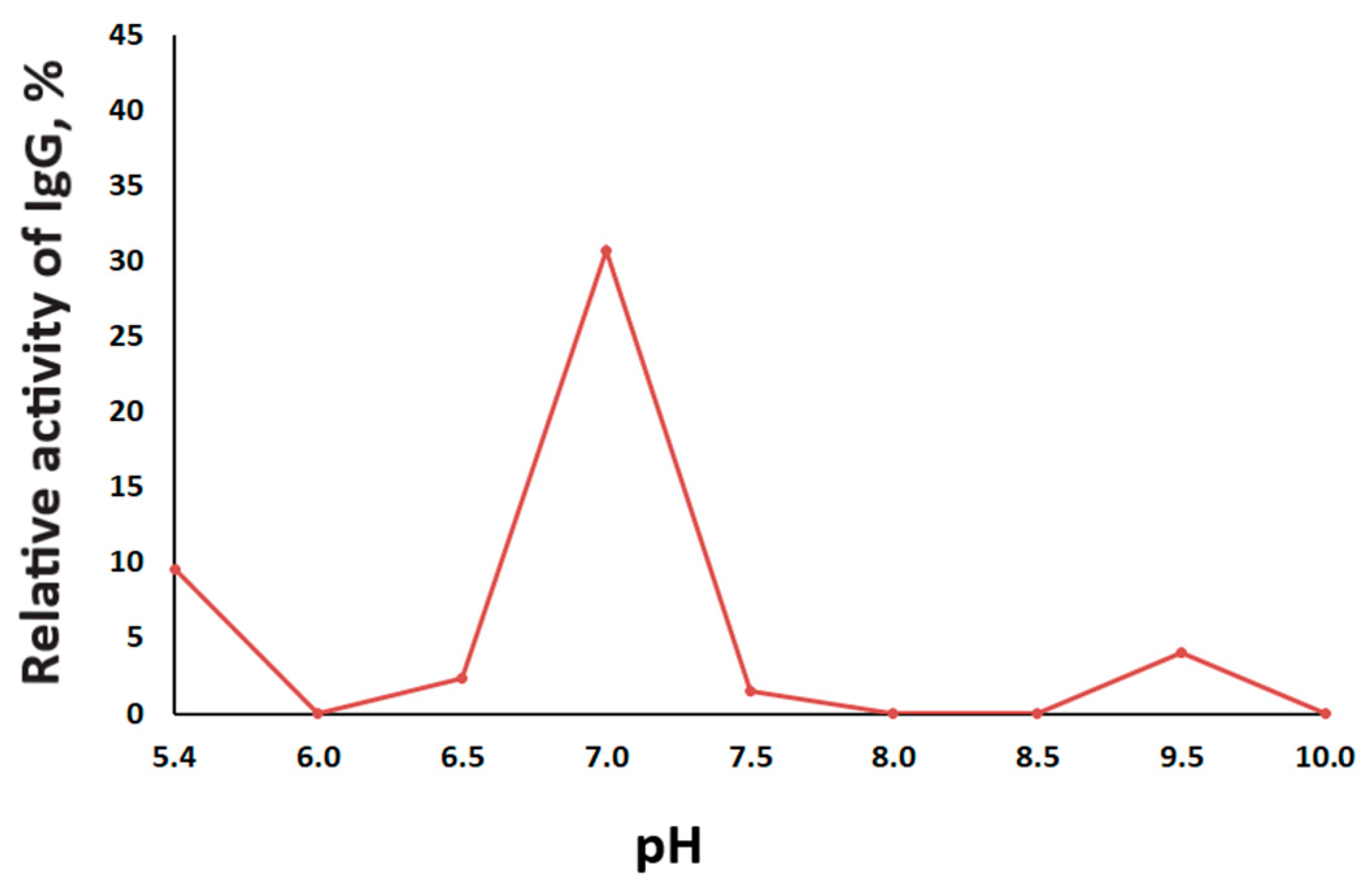
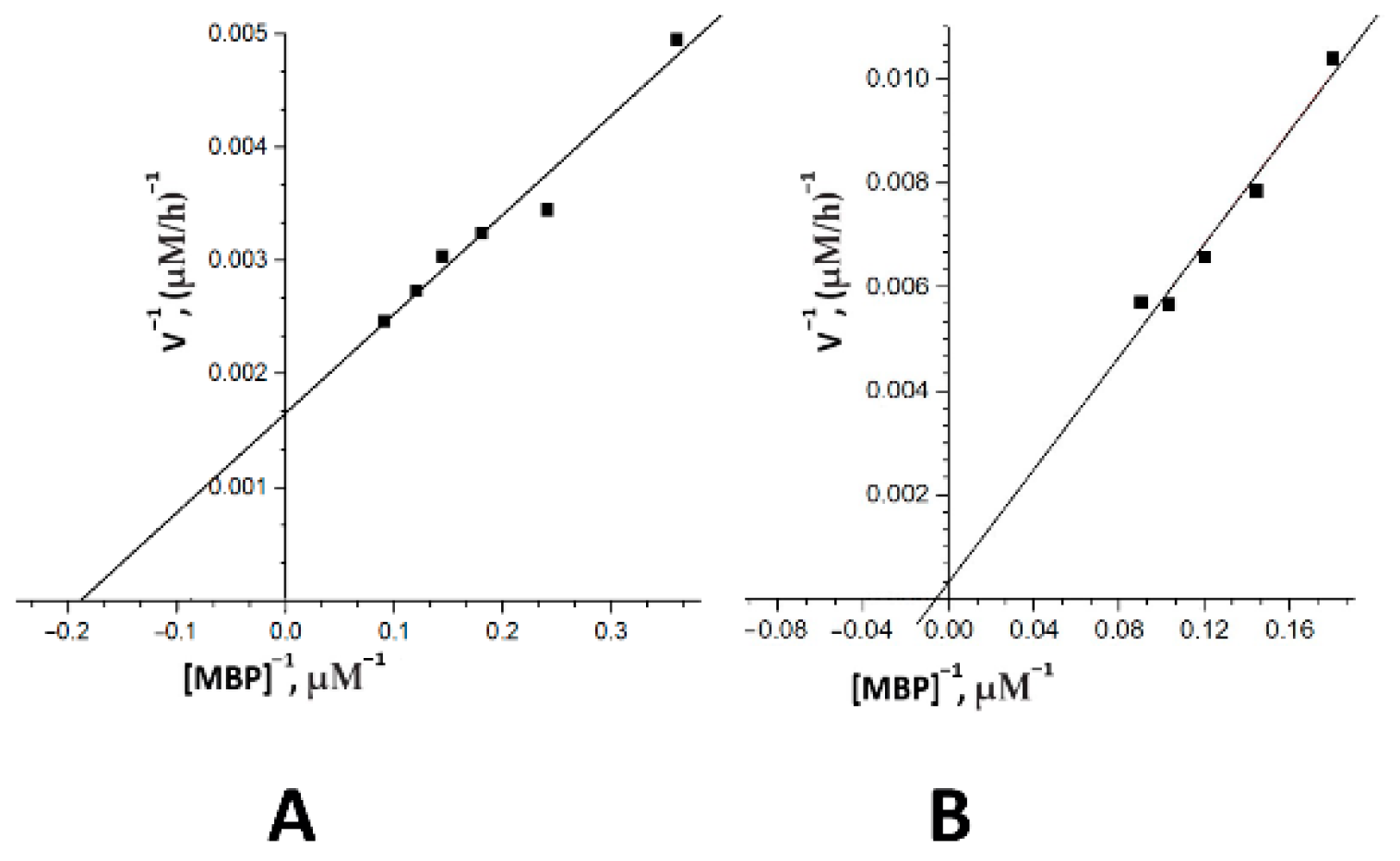
2.3. The Catalytic Properties of MBP-hydrolyzing IgGs in Bipolar Disorder
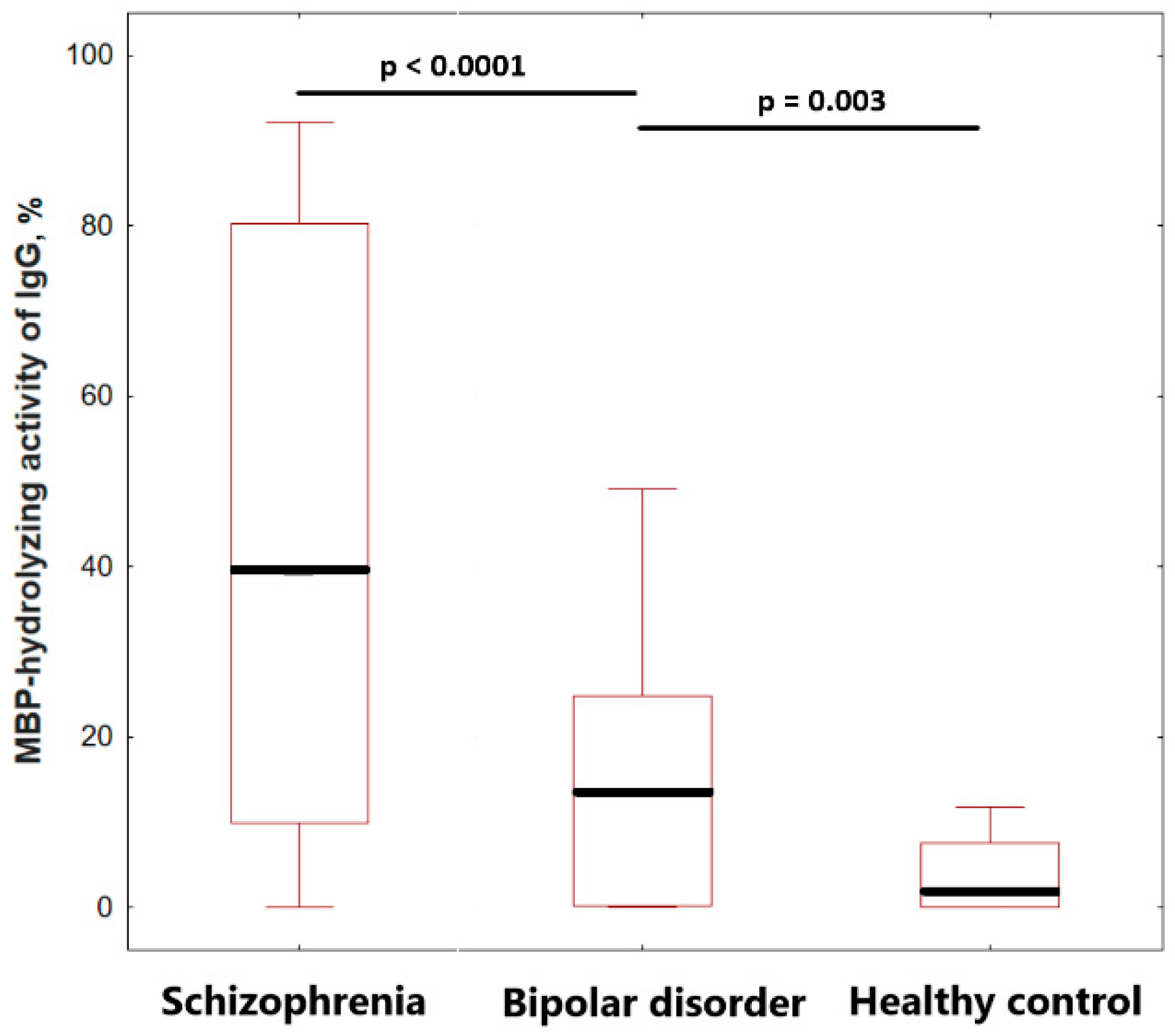
2.4. Comparison of IgGs MBP-Hydrolyzing Activity of Healthy Individuals and Patients
3. Discussion
4. Materials and Methods
4.1. Patients, Healthy Donors, and Biological Material
4.2. Purification of Serum IgGs
4.3. High-Performance Gel Filtration of Antibodies under “Acid Shock” Condition
4.4. Proteolytic Activity Assay of IgGs
4.5. Analysis of Substrate Specificity of Antibodies
4.6. The Effect of pH on the Proteolytic Activity of IgG
4.7. Inhibitory Assay of MBP-hydrolyzing Activity
4.8. Kinetic Parameter Analysis
4.9. Immunoblotting of Myelin Basic Protein
4.10. Western Blotting of MBP-Binding IgG
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Spearman Correlation Coefficient (r) * |
|---|---|
| Age, years | 0.045 |
| Age of manifistation, years | −0.003 |
| Duration of disorder | −0.193 |
| Number of episodes | −0.031 |
| CGI | 0.278 |
| SIGH-SAD total | −0.297 |
| HCL-32 | −0.035 |
References
- Rowland, T.; Marwaha, S. Epidemiology and risk factors for bipolar disorder. Ther. Adv. Psychopharmacol. 2018, 8, 251–269. [Google Scholar] [CrossRef]
- GBD 2019 Diseases and Injuries Collaborators. Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet. Psychiatry 2022, 9, 137–150. [Google Scholar] [CrossRef]
- Vieta, E.; Berk, M.; Schulze, T.G.; Carvalho, A.F.; Suppes, T.; Calabrese, J.R.; Gao, K.; Miskowiak, K.W.; Grande, I. Bipolar disorders. Nat. Rev. Dis. Primers 2018, 4, 18008. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Mitra, P. Bipolar Affective Disorder; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Hayes, J.F.; Miles, J.; Walters, K.; King, M.; Osborn, D.P.J. A systematic review and meta-analysis of premature mortality in bipolar affective disorder. Acta Psychiatr. Scand. 2015, 131, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Nguyen, L.; Hu, R.; Stavish, C.M.; Leibenluft, E.; Linke, J.O. The uncinate fasciculus in individuals with and at risk for bipolar disorder: A meta-analysis. J. Affect. Disord. 2022, 297, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Bi, W.; Liu, C.; Zhao, Y.; Zhang, D.; Yue, W. A hypothesis-driven pathway analysis reveals myelin-related pathways that contribute to the risk of schizophrenia and bipolar disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 51, 140–145. [Google Scholar] [CrossRef]
- Sklar, P.; Smoller, J.W.; Fan, J.; Ferreira, M.A.; Perlis, R.H.; Chambert, K.; Nimgaonkar, V.L.; McQueen, M.B.; Faraone, S.V.; Kirby, A.; et al. Whole-genome association study of bipolar disorder. Mol. Psychiatry 2008, 13, 558–569. [Google Scholar] [CrossRef]
- Ji, E.; Lejuste, F.; Sarrazin, S.; Houenou, J. From the microscope to the magnet: Disconnection in schizophrenia and bipolar disorder. Neurosci. Biobehav. Rev. 2019, 98, 47–57. [Google Scholar] [CrossRef]
- Uranova, N.; Orlovskaya, D.; Vikhreva, O.; Zimina, I.; Kolomeets, N.; Vostrikov, V.; Rachmanova, V. Electron microscopy of oligodendroglia in severe mental illness. Brain Res. Bull. 2001, 55, 597–610. [Google Scholar] [CrossRef]
- Hu, R.; Stavish, C.; Leibenluft, E.; Linke, J.O. White matter microstructure in individuals with and at risk for bipolar disorder: Evidence for an endophenotype from a voxel-based meta-analysis. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 1104–1113. [Google Scholar] [CrossRef]
- Hahn, C.; Lim, H.K.; Lee, C.U. Neuroimaging findings in late-onset schizophrenia and bipolar disorder. J. Geriatr. Psychiatry Neurol. 2014, 27, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Sehmbi, M.; Rowley, C.D.; Minuzzi, L.; Kapczinski, F.; Kwiecien, J.M.; Bock, N.A.; Frey, B.N. Age-related deficits in intracortical myelination in young adults with bipolar disorder type I. J. Psychiatry Neurosci. 2019, 44, 79–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitaker, J.N. Myelin basic protein in cerebrospinal fluid and other body fluids. Mult. Scler. 1998, 4, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Boggs, J.M. Myelin basic protein: A multifunctional protein. Cell. Mol. Life Sci. 2006, 63, 1945–1961. [Google Scholar] [CrossRef]
- Shibata, D.; Cain, K.; Tanzi, P.; Zierath, D.; Becker, K. Myelin basic protein autoantibodies, white matter disease and stroke outcome. J. Neuroimmunol. 2012, 252, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Rahman, H.A.; Salama, M.; Seham, A.; El-Hak, G.; El-Harouny, M.A.; ElKafrawy, P.; Abou-Donia, M.B. A Panel of Autoantibodies against Neural Proteins as Peripheral Biomarker for Pesticide-Induced. Neurotox. Res. 2018, 33, 316–336. [Google Scholar] [CrossRef]
- Martinsen, V.; Kursula, P. Multiple sclerosis and myelin basic protein: Insights into protein disorder and disease. Amino Acids. 2022, 54, 99–109. [Google Scholar] [CrossRef]
- Marchiori, P.E.; Dos Reis, M.; Quevedo, M.E.; Callegaro, D.; Hirata, M.T.; Scaff, M.; De Oliveira, R.M. Cerebrospinal fluid and serum antiphospholipid antibodies in multiple sclerosis, Guillain-Barré syndrome and systemic lupus erythematosus. Arq. Neuropsiquiatr. 1990, 48, 465–468. [Google Scholar] [CrossRef]
- Singh, V.K.; Warren, R.P.; Odell, J.D.; Warren, W.L.; Cole, P. Antibodies to myelin basic protein in children with autistic behavior. Brain Behav. Immun. 1993, 7, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Kliushnik, T.P.; Siriachenko, T.M.; Sarmanova, Z.V.; Otman, I.N.; Dupin, A.M.; Sokolov, R.E. Changes of the level of serum antibodies to neuroantigens in patients with schizophrenia during the treatment. Zhurnal Nevrol. Psikhiatrii Im. SS Korsakova 2008, 108, 61–64. [Google Scholar]
- Altamura, A.C.; Buoli, M.; Pozzoli, S. Role of immunological factors in the pathophysiology and diagnosis of bipolar disorder: Comparison with schizophrenia. Psychiatry Clin. Neurosci. 2014, 68, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Muneer, A. Bipolar Disorder: Role of Inflammation and the Development of Disease Biomarkers. Psychiatry Investig. 2016, 13, 18–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dazzan, P.; Fusté, M.; Davies, W. Do defective immune system-mediated myelination processes increase postpartum psychosis risk? Trends. Mol. Med. 2018, 24, 942–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, J.P.; Frey, B.N. Disruption in the blood-brain barrier: The missing link between brain and body inflammation in bipolar disorder? Neural Plast. 2015, 2015, 708306. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.V.; Passos, I.C.; Librenza-Garcia, D.; Marcon, G.; Schneider, M.A.; Conte, J.H.; da Silva, J.P.A.; Lima, L.P.; Quincozes-Santos, A.; Kauer-Sant’Anna, M.; et al. Neuron-glia interaction as a possible pathophysiological mechanism of bipolar disorder. Curr. Neuropharmacol. 2018, 16, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Sæther, S.G.; Rø, A.D.B.; Larsen, J.B.; Vaaler, A.; Kondziella, D.; Reitan, S.K. Biomarkers of autoimmunity in acute psychiatric disorders. J. Neuropsychiatry Clin. Neurosci. 2019, 31, 246–253. [Google Scholar] [CrossRef]
- Gan, Z.; Wu, X.; Chen, Z.; Liao, Y.; Wu, Y.; He, Z.; Yang, Z.; Zhang, Q. Rapid cycling bipolar disorder is associated with antithyroid antibodies, instead of thyroid dysfunction. BMC Psychiatry 2019, 19, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Choe, C.U.; Karamatskos, E.; Schattling, B.; Leypoldt, F.; Liuzzi, G.; Gerloff, C.; Friese, M.A.; Mulert, C. A clinical and neurobiological case of IgM NMDA receptor antibody associated encephalitis mimicking bipolar disorder. Psychiatry Res. 2013, 208, 194–196. [Google Scholar] [CrossRef]
- Hammer, C.; Stepniak, B.; Schneider, A.; Papiol, S.; Tantra, M.; Begemann, M.; Siren, A.L.; Pardo, L.A.; Sperling, S.; Mohd Jofrry, S.; et al. Neuropsychiatric disease relevance of circulating anti-nmda receptor autoantibodies depends on blood-brain barrier integrity. Mol. Psychiatry 2014, 19, 1143–1149. [Google Scholar] [CrossRef]
- Paul, S.; Volle, D.J.; Beach, C.M.; Johnson, D.R.; Powell, M.J.; Massey, R.J. Catalytic hydrolysis of vasoactive intestinal peptide by human autoantibody. Science 1989, 244, 1158–1162. [Google Scholar] [CrossRef]
- Belogurov, A., Jr.; Kozyr, A.; Ponomarenko, N.; Gabibov, A. Catalytic antibodies: Balancing between Dr. Jekyll and Mr. Hyde. Bioessays 2009, 31, 1161–1171. [Google Scholar] [CrossRef] [PubMed]
- Mahendra, A.; Sharma, M.; Rao, D.N.; Peyron, I.; Planchais, C.; Dimitrov, J.D.; Kaveri, S.V.; Lacroix-Desmazes, S. Antibody-mediated catalysis: Induction and therapeutic relevance. Autoimmun. Rev. 2013, 12, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Polosukhina, D.I.; Kanyshkova, T.G.; Doronin, B.M.; Tyshkevich, O.B.; Buneva, V.N.; Boiko, A.N.; Gusev, E.I.; Favorova, O.O.; Nevinsky, G.A. Hydrolysis of myelin basic protein by polyclonal catalytic IgGs from the sera of patients with multiple sclerosis. J. Cell Mol. Med. 2004, 8, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Bezuglova, A.V.; Konenkova, L.P.; Doronin, B.M.; Buneva, V.N.; Nevinsky, G.A. Affinity and catalytic heterogeneity and metal-dependence of polyclonal myelin basic protein-hydrolyzing IgGs from sera of patients with systemic lupus erythematosus. J. Mol. Recognit. 2011, 24, 960–974. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gronow, M.; Cuchacovich, M.; Francos, R.; Cuchacovich, S.; Blanco, A.; Sandoval, R.; Gomez, C.F.; Valenzuela, J.A.; Ray, R.; Pizzo, S.V. Catalytic autoantibodies against myelin basic protein (MBP) isolated from serum of autistic children impair in vitro models of synaptic plasticity in rat hippocampus. J. Neuroimmunol. 2015, 287, 1–8. [Google Scholar] [CrossRef]
- Parshukova, D.; Smirnova, L.P.; Ermakov, E.A.; Bokhan, N.A.; Semke, A.V.; Ivanova, S.A.; Buneva, V.N.; Nevinsky, G.A. Autoimmunity and immune system dysregulation in schizophrenia: IgGs from sera of patients hydrolyze myelin basic protein. J. Mol. Recognit. 2019, 32, e2759. [Google Scholar] [CrossRef]
- Parshukova, D.A.; Smirnova, L.P.; Kornetova, E.G.; Semke, A.V.; Buneva, V.N.; Ivanova, S.A. IgG-Dependent hydrolysis of myelin basic protein of patients with different courses of schizophrenia. J. Immunol. Res. 2020, 2020, 8986521. [Google Scholar] [CrossRef]
- Ermakov, E.A.; Parshukova, D.A.; Nevinsky, G.A.; Buneva, V.N. Natural Catalytic IgGs Hydrolyzing Histones in Schizophrenia: Are They the Link between Humoral Immunity and Inflammation? Int. J. Mol. Sci. 2020, 21, 7238. [Google Scholar] [CrossRef]
- Nevinsky, G.A.; Baranova, S.V.; Buneva, V.N.; Dmitrenok, P.S. Multiple Sclerosis: Enzymatic Cross Site-Specific Hydrolysis of H1 Histone by IgGs against H1, H2A, H2B, H3, H4 Histones, and Myelin Basic Protein. Biomolecules 2021, 11, 1140. [Google Scholar] [CrossRef]
- Magorivska, I.B.; Bilyy, R.O.; Havrylyuk, A.M.; Chop’yak, V.V.; Stoika, R.S.; Kit, Y.Y. Anti-histone H1 IgGs from blood serum of systemic lupus erythematosus patients are capable of hydrolyzing histone H1 and myelin basic protein. J. Mol. Recognit. 2010, 23, 495–502. [Google Scholar] [CrossRef]
- Baranova, S.V.; Mikheeva, E.V.; Buneva, V.N.; Nevinsky, G.A. Antibodies from the sera of multiple sclerosis patients efficiently hydrolyze five histones. Biomolecules 2019, 9, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, R.; Sundaresh, A.; Rajkumar, R.P.; Negi, V.S.; Vijayalakshmi, M.A.; Krishnamoorthy, R.; Tamouza, R.; Leboyer, M.; Kamalanathan, A.S. DNA hydrolysing IgG catalytic antibodies: An emerging link between psychoses and autoimmunity. NPJ Schizophr. 2021, 7, 1–7. [Google Scholar] [CrossRef]
- Ganesh, C.; Shah, A.N.; Swaminathan, C.P.; Surolia, A.; Varadarajan, R. Thermodynamic characterization of the reversible, two-state unfolding of maltose binding protein, a large two-domain protein. Biochemistry 1997, 36, 5020–5028. [Google Scholar] [CrossRef] [PubMed]
- Nevinsky, G.A. Natural catalytic antibodies in norm and in autoimmune diseases. In Autoimmune Diseases: Symptoms, Diagnosis and Treatment; Brenner, K.J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2010; pp. 1–107. [Google Scholar]
- Nevinsky, G.A.; Buneva, V.N. Autoantibodies and Natural Catalytic Antibodies in Health, Multiple Sclerosis, and Some Other Diseases. Adv. Neuroimmune Biol. 2012, 3, 157–182. [Google Scholar] [CrossRef]
- Belov, S.; Buneva, V.N.; Nevinsky, G.A. How human IgGs against myelin basic protein (MBP) recognize oligopeptides and MBP. J. Mol. Recognit. 2017, 30, e2637. [Google Scholar] [CrossRef]
- Rao, M.B.; Tanksale, A.M.; Ghatge, M.S.; Deshpande, V.V. Molecular and biotechnological aspects of microbial proteases. Microbiol. Mol. Biol. Rev. 1998, 62, 597–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legostaeva, G.A.; Polosukhina, D.I.; Bezuglova, A.M.; Doronin, B.M.; Buneva, V.N.; Nevinsky, G.A. Affinity and catalytic heterogeneity of polyclonal myelin basic protein-hydrolyzing IgGs from sera of patients with multiple sclerosis. J. Cell Mol. Med. 2010, 14, 699–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta 2012, 1824, 68–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanman, L.E.; van der Linden, W.A.; Verdoes, M.; Bogyo, M. Bifunctional Probes of Cathepsin Protease Activity and pH Reveal Alterations in Endolysosomal pH during Bacterial Infection. Cell Chem. Biol. 2016, 23, 793–804. [Google Scholar] [CrossRef] [Green Version]
- Erra Díaz, F.; Dantas, E.; Geffner, J. Unravelling the Interplay between Extracellular Acidosis and Immune Cells. Mediat. Inflamm. 2018, 2018, 1218297. [Google Scholar] [CrossRef]
- Edlow, D.W.; Sheldon, W.H. The pH of inflammatory exudates. Proc. Soc. Exp. Biol. Med. 1971, 137, 1328–1332. [Google Scholar] [CrossRef] [PubMed]
- Kocks, C.; Rajewsky, K. Stepwise intraclonal maturation of antibody affinity through somatic hypermutation. Proc. Natl. Acad. Sci. USA 1988, 85, 8206–8210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, H.N. Affinity enhancement of antibodies: How low-affinity antibodies produced early in immune responses are followed by high-affinity antibodies later and in memory B-cell responses. Cancer Immunol. Res. 2014, 2, 381–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevinsky, G.A.; Buneva, V.N. Catalytic antibodies in healthy humans and patients with autoimmune and viral pathologies. J. Cell Mol. Med. 2003, 7, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Jiang, Q.; Zhang, L. The prevalence of bipolar disorder in autoimmune disease: A systematic review and meta-analysis. Ann. Palliat. Med. 2021, 10, 350–361. [Google Scholar] [CrossRef]
- Jakobsson, J.; Bjerke, M.; Ekman, C.J.; Sellgren, C.; Johansson, A.G.; Zetterberg, H.; Blennow, K.; Landén, M. Elevated Concentrations of Neurofilament Light Chain in the Cerebrospinal Fluid of Bipolar Disorder Patients. Neuropsychopharmacology 2014, 39, 2349–2356. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Gildengers, A.G.; Hsu, J.L.; Chung, K.H.; Chen, P.H.; Huang, Y.J. Inflammation associated with volume reduction in the gray matter and hippocampus of older patients with bipolar disorder. J. Affect. Disord. 2019, 244, 60–66. [Google Scholar] [CrossRef]
- Smirnova, L.P.; Yarnykh, V.L.; Parshukova, D.A.; Kornetova, E.G.; Semke, A.V.; Usova, A.V.; Pishchelko, A.O.; Khodanovich, M.Y.; Ivanova, S.A. Global Hypomyelination of the Brain White and Gray Matter in Schizophrenia: Quantitative Imaging Using Macromolecular Proton Fraction. Transl. Psychiatry 2021, 11, 365. [Google Scholar] [CrossRef]
- Hidese, S.; Hattori, K.; Sasayama, D.; Tsumagari, T.; Miyakawa, T.; Matsumura, R.; Yokota, Y.; Ishida, I.; Matsuo, J.; Yoshida, S.; et al. Cerebrospinal fluid inflammatory cytokine levels in patients with major psychiatric disorders: A multiplex immunoassay study. Front. Pharmacol. 2021, 11, 2474. [Google Scholar] [CrossRef]
- Lyu, N.; Xing, G.; Yang, J.; Zhu, X.; Zhao, X.; Zhang, L.; Wang, G. Comparison of inflammatory, nutrient, and neurohormonal indicators in patients with schizophrenia, bipolar disorder and major depressive disorder. J. Psychiat. Res. 2021, 137, 401–408. [Google Scholar] [CrossRef]
- Taguchi, H.; Planque, S.; Nishiyama, Y.; Szabo, P.; Weksler, M.E.; Friedland, R.P.; Paul, S. Catalytic antibodies to amyloid β peptide in defense against Alzheimer disease. Autoimmun. Rev. 2008, 7, 391–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranova, S.V.; Dmitrienok, P.S.; Buneva, V.N.; Nevinsky, G.A. Autoantibodies in HIV-infected patients: Cross site-specific hydrolysis of H1 histone and myelin basic protein. Biofactors 2019, 45, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Platt, M.P.; Agalliu, D.; Cutforth, T. Hello from the Other Side: How Autoantibodies Circumvent the Blood–Brain Barrier in Autoimmune Encephalitis. Front. Immunol. 2017, 8, 442. [Google Scholar] [CrossRef] [Green Version]
- Fries, G.R.; Walss-Bass, C.; Bauer, M.E.; Teixeira, A.L. Revisiting inflammation in bipolar disorder. Pharmacol. Biochem. Behav. 2019, 177, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Rosenblat, J.D.; McIntyre, R.S. Bipolar Disorder and Inflammation. Psychiatr. Clin. N. Am. 2016, 39, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Vadodaria, K.C.; Mendes, A.P.D.; Mei, A.; Racha, V.; Erikson, G.; Shokhirev, M.N.; Oefner, R.; Heard, K.J.; McCarthy, M.J.; Eyler, L.; et al. Altered neuronal support and inflammatory response in bipolar disorder patient-derived astrocytes. Stem Cell Rep. 2021, 16, 825–835. [Google Scholar] [CrossRef]
- Pereira, A.C.; Oliveira, J.; Silva, S.; Madeira, N.; Pereira, C.M.; Cruz, M.T. Inflammation in bipolar disorder (BD): Identification of new therapeutic targets. Pharmacol. Res. 2021, 163, 105325. [Google Scholar] [CrossRef]
- Misiak, B.; Bartoli, F.; Carrà, G.; Małecka, M.; Samochowiec, J.; Jarosz, K.; Banik, A.; Stańczykiewicz, B. Chemokine alterations in bipolar disorder: A systematic review and meta-analysis. BrainBehav. Immun. 2020, 88, 870–877. [Google Scholar] [CrossRef]
- Snijders, G.; Brouwer, R.; Kemner, S.; Bootsman, F.; Drexhage, H.A.; Hillegers, M.H.J. Genetic and environmental influences on circulating NK and T cells and their relation to bipolar disorder. Int. J. Bipolar. Disord. 2019, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Benedetti, F.; Aggio, V.; Pratesi, M.L.; Greco, G.; Furlan, R. Neuroinflammation in bipolar depression. Front. Psychiatry 2020, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- Giridharan, V.V.; Sayana, P.; Pinjari, O.F.; Ahmad, N.; da Rosa, M.I.; Quevedo, J.; Barichello, T. Postmortem evidence of brain inflammatory markers in bipolar disorder: A systematic review. Mol. Psychiatry 2020, 25, 94–113. [Google Scholar] [CrossRef] [PubMed]
- Becking, K.; Haarman, B.C.; van der Lek, R.F.; Grosse, L.; Nolen, W.A.; Claes, S.; Drexhage, H.A.; Schoevers, R.A. Inflammatory monocyte gene expression: Trait or state marker in bipolar disorder? Int. J. Bipolar. Disord. 2015, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isgren, A.; Sellgren, C.; Ekman, C.J.; Holmén-Larsson, J.; Blennow, K.; Zetterberg, H.; Jakobsson, J.; Landén, M. Markers of neuroinflammation and neuronal injury in bipolar disorder: Relation to prospective clinical outcomes. Brain Behav. Immun. 2017, 65, 195–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedetti, F.; Poletti, S.; Hoogenboezem, T.A.; Locatelli, C.; de Wit, H.; Wijkhuijs, A.J.M.; Colombo, C.; Drexhage, H.A. Higher Baseline Proinflammatory Cytokines Mark Poor Antidepressant Response in Bipolar Disorder. J. Clin. Psychiatry 2017, 78, e986–e993. [Google Scholar] [CrossRef] [PubMed]
- Smirnova, L.P.; Mednova, I.A.; Krotenko, N.M.; Alifirova, V.M.; Ivanova, S.A. IgG-Dependent Dismutation of Superoxide in Patients with Different Types of Multiple Sclerosis and Healthy Subjects. Oxid. Med. Cell Longev. 2020, 2020, 8171020. [Google Scholar] [CrossRef]
- Grodzki, A.C.; Berenstein, E. Antibody Purification: Affinity Chromatography—Protein A and Protein G Sepharose. In Immunocytochemical Methods and Protocols; Humana Press: New York, NY, USA, 2010. [Google Scholar] [CrossRef]
- Sapparapu, G.; Planque, S.A.; Nishiyama, Y.; Foung, S.K.; Paul, S. Antigen-specific proteolysis by hybrid antibodies containing promiscuous proteolytic light chains paired with an antigen-binding heavy chain. J. Biol. Chem. 2009, 284, 24622–24633. [Google Scholar] [CrossRef] [Green Version]







| Parameter | Healthy Controls | Bipolar Disorder | Schizophrenia |
|---|---|---|---|
| Number of participants, persons | 20 | 25 | 30 |
| Sex (male/female), % | 42/58 | 29/71 | 31/69 |
| Age, years | 34.96 ± 10.13 | 37.1 ± 14.6 | 39.42 ± 11.5 |
| BD I type/BD II type, persons | - | 20/5 | - |
| Duration of disorder, years | - | 12.1 ± 9.0 | 13.6 ± 7.59 |
| Number of episodes | - | 6.7 ± 5.0 | - |
| CGI | - | 4.6 ± 0.9 | 4.8 ± 0.7 |
| SIGH-SAD total | - | 24.4 ± 7.8 | - |
| HCL-32 | - | 17.7 ± 4.7 | - |
| PANSS total score | - | - | 89.00 (80.00; 105.00) |
| Kinetics Parameters | IgG-1 1 | IgG-2 1 |
|---|---|---|
| Km, mkM | 5.2 ± 0.43 | 150 ± 10.6 |
| kcat, min−1 | 7.7 ± 0.35 | (3.85 ± 0.9) × 10−6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamaeva, D.A.; Smirnova, L.P.; Vasilieva, S.N.; Kazantseva, D.V.; Vasilieva, A.R.; Ivanova, S.A. Catalytic Antibodies in Bipolar Disorder: Serum IgGs Hydrolyze Myelin Basic Protein. Int. J. Mol. Sci. 2022, 23, 7397. https://doi.org/10.3390/ijms23137397
Kamaeva DA, Smirnova LP, Vasilieva SN, Kazantseva DV, Vasilieva AR, Ivanova SA. Catalytic Antibodies in Bipolar Disorder: Serum IgGs Hydrolyze Myelin Basic Protein. International Journal of Molecular Sciences. 2022; 23(13):7397. https://doi.org/10.3390/ijms23137397
Chicago/Turabian StyleKamaeva, Daria A., Liudmila P. Smirnova, Svetlana N. Vasilieva, Daria V. Kazantseva, Alisa R. Vasilieva, and Svetlana A. Ivanova. 2022. "Catalytic Antibodies in Bipolar Disorder: Serum IgGs Hydrolyze Myelin Basic Protein" International Journal of Molecular Sciences 23, no. 13: 7397. https://doi.org/10.3390/ijms23137397