
Low-Dose Tacrolimus Promotes the Migration and Invasion and Nitric Oxide Production in the Human-Derived First Trimester Extravillous Trophoblast Cells In Vitro



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Test-Article Formulations
2.2. IncuCyte Scratch Wound Assay and In Vitro Live-Cell Migration Analysis
2.3. In Vitro Matrigel Invasion Assay
2.4. Western Blot Analysis
2.5. Detection of NO Production in the HTR-8/SVneo Cells
2.6. Fluorescent Microscopy
2.7. Detection of PGR mRNA in HTR8/SVneo Cells
2.8. Statistical Analyses
3. Results
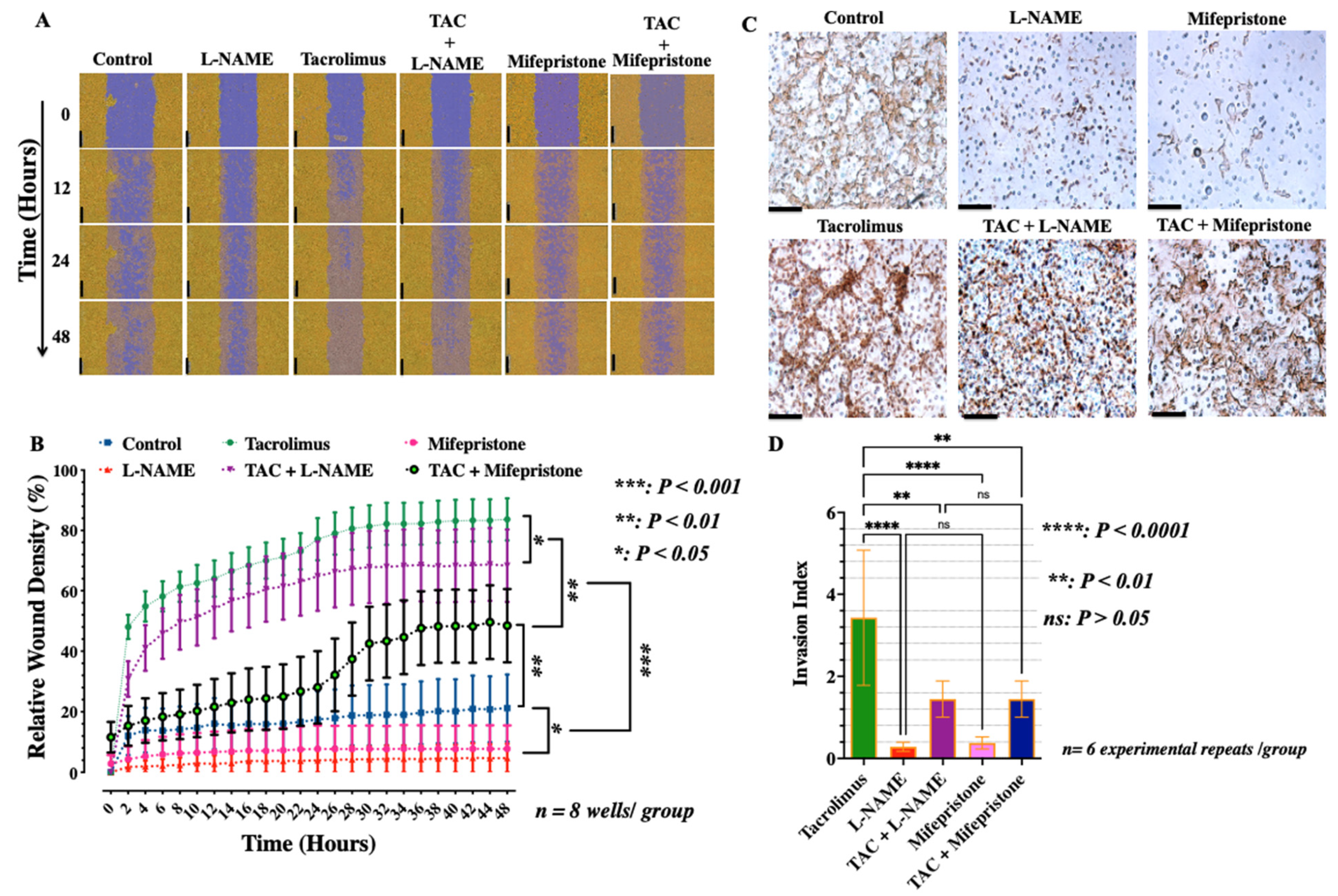
3.1. Low-Dose Tacrolimus Abrogates the Suppressive Effect of L-NAME and Promotes the Human-Derived First-Trimester Extravillous Trophoblast Cell Migration and Invasion In Vitro
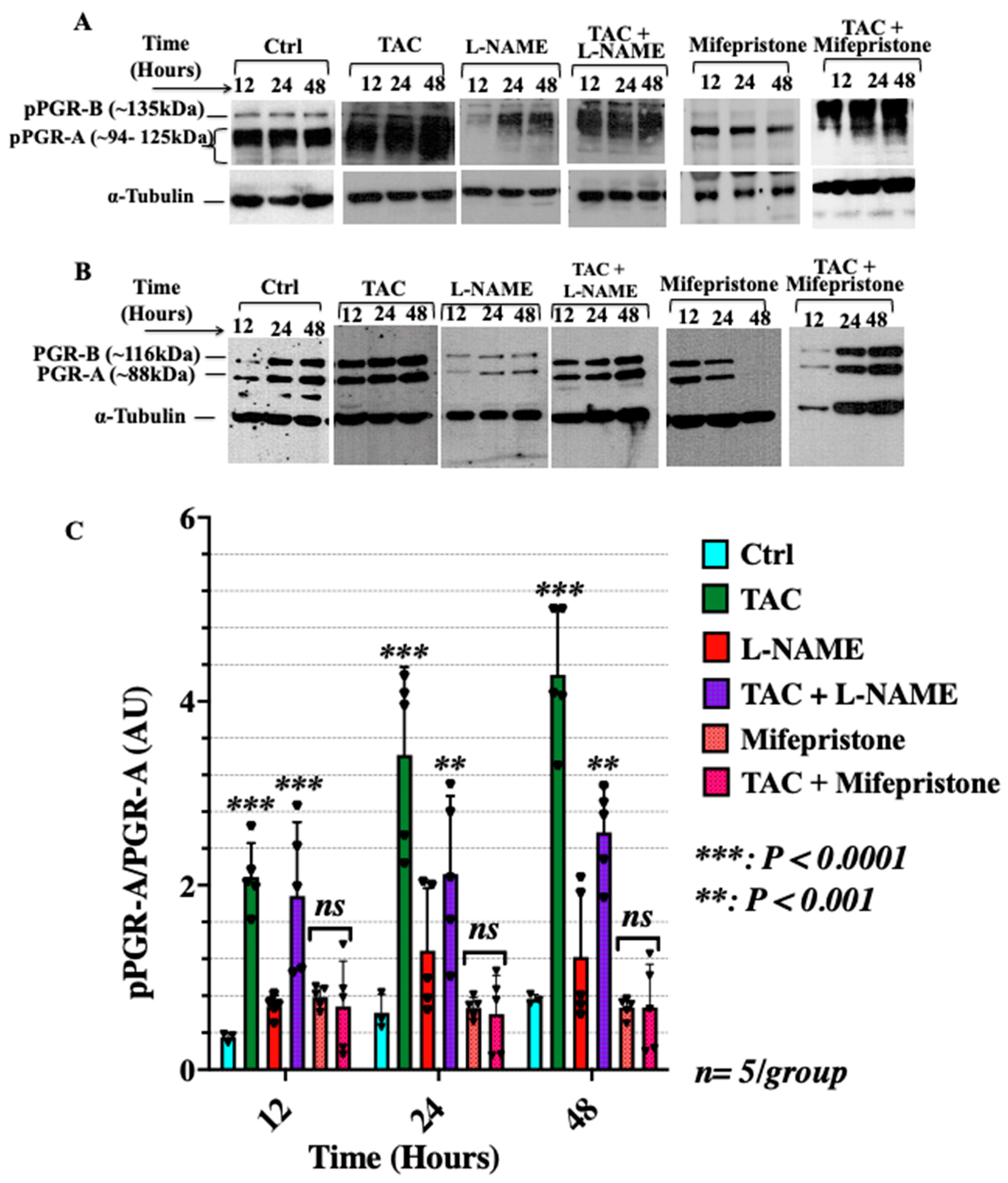
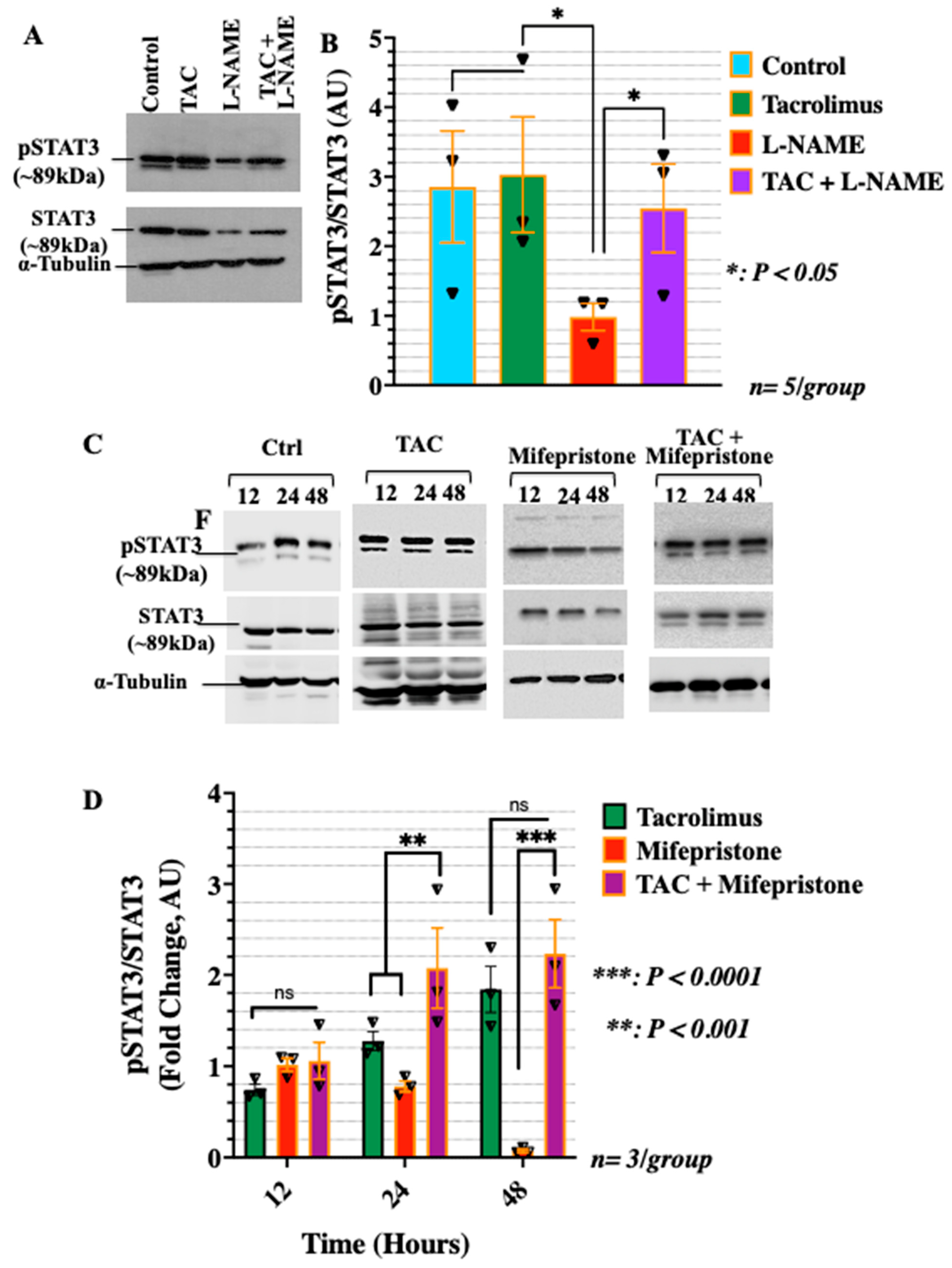
3.2. Low-Dose Tacrolimus Rescued the Expression of the Active Forms of the Cytosolic Progesterone Receptors and STAT3 in the HTR-8/SVneo Cells Cultured under Nitrosative Stresses In Vitro
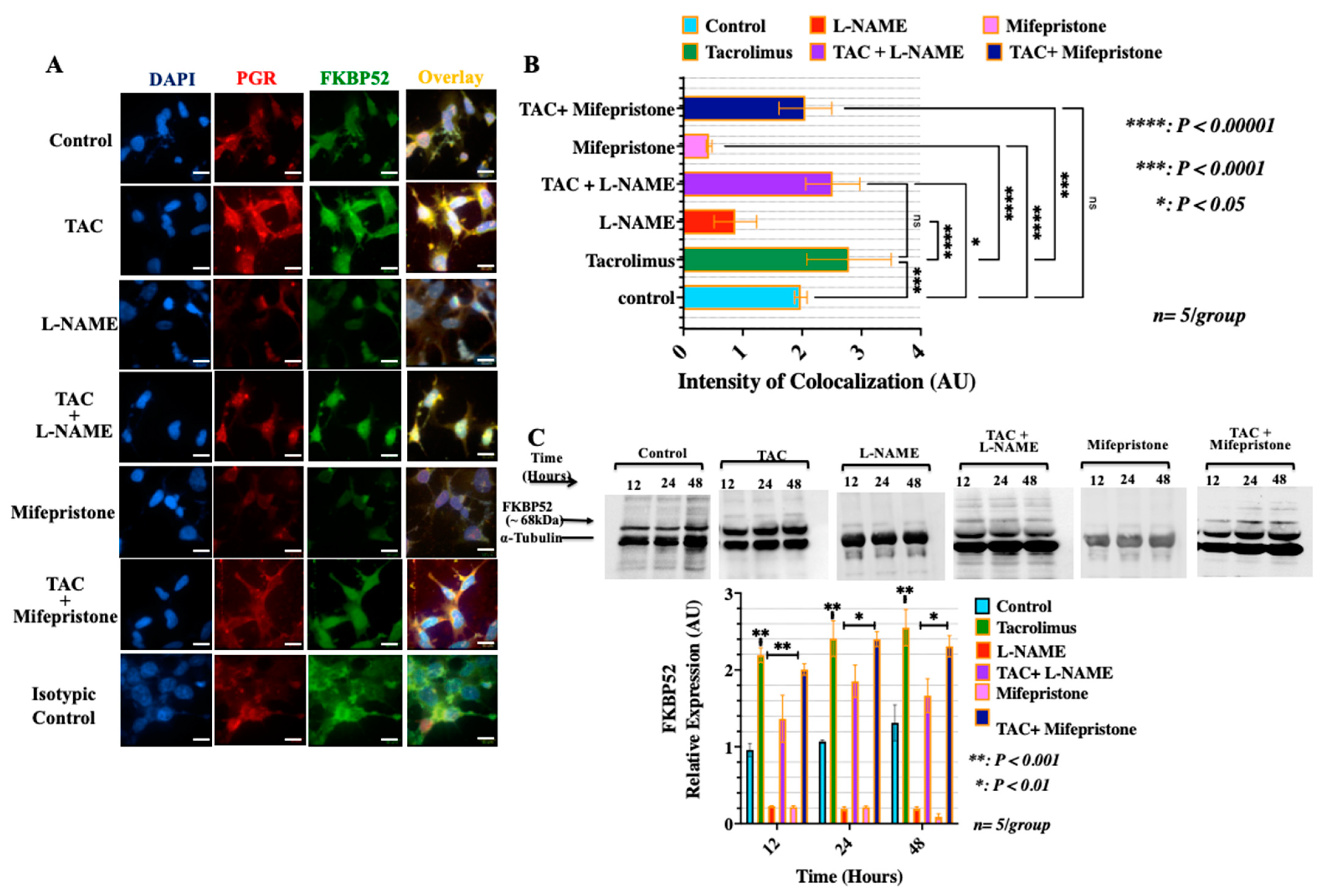
3.3. Treatment with Low-Dose Tacrolimus Increased the Expression of FKBP52 and Its Colocalization to the Cytosolic Progesterone Receptors in the HTR-8/SVneo Cells In Vitro
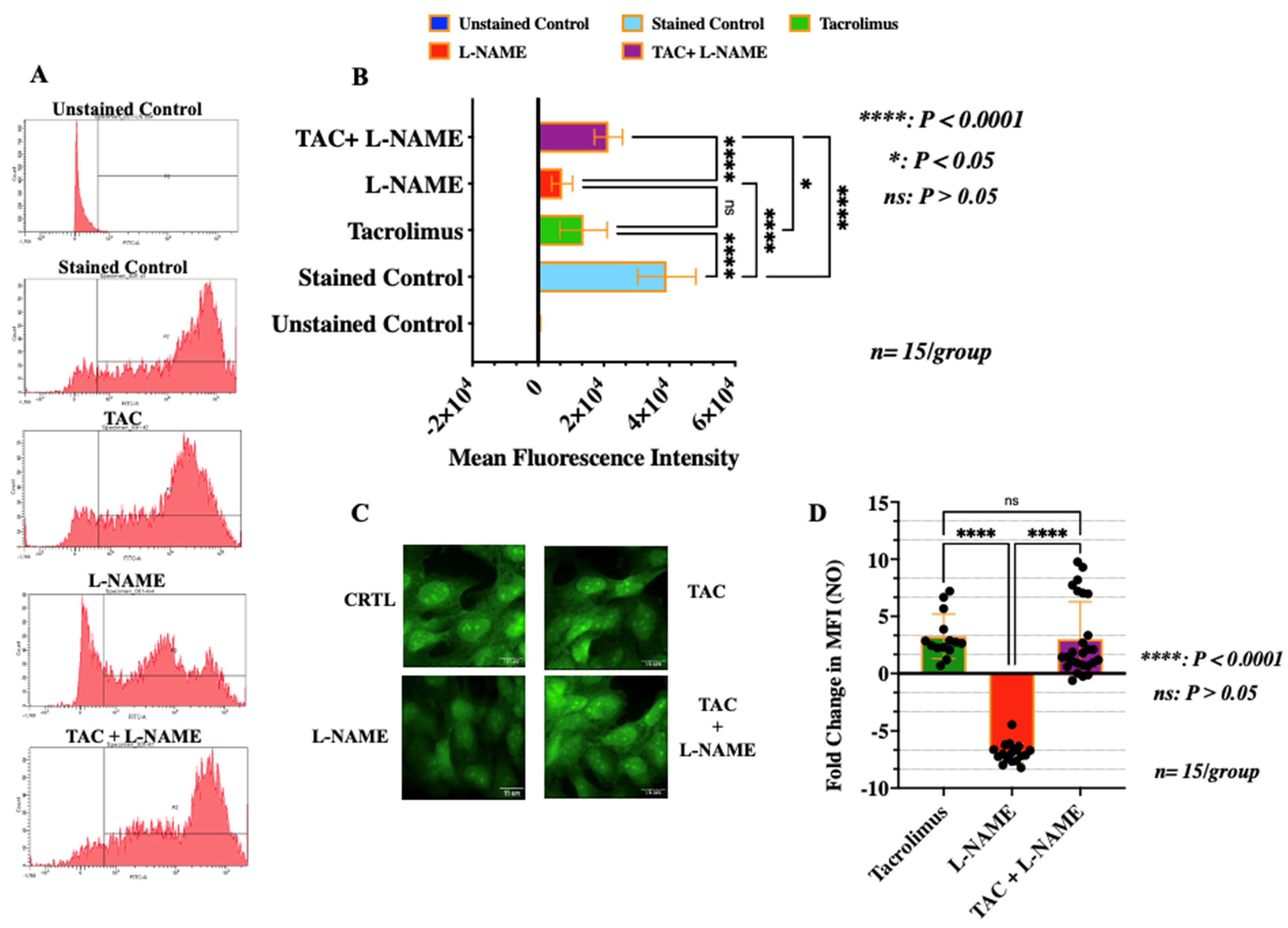
3.4. Treatment with Low-Dose Tacrolimus Rescued the Production of NO in HTR-8/SVneo Cells in the Presence of L-NAME
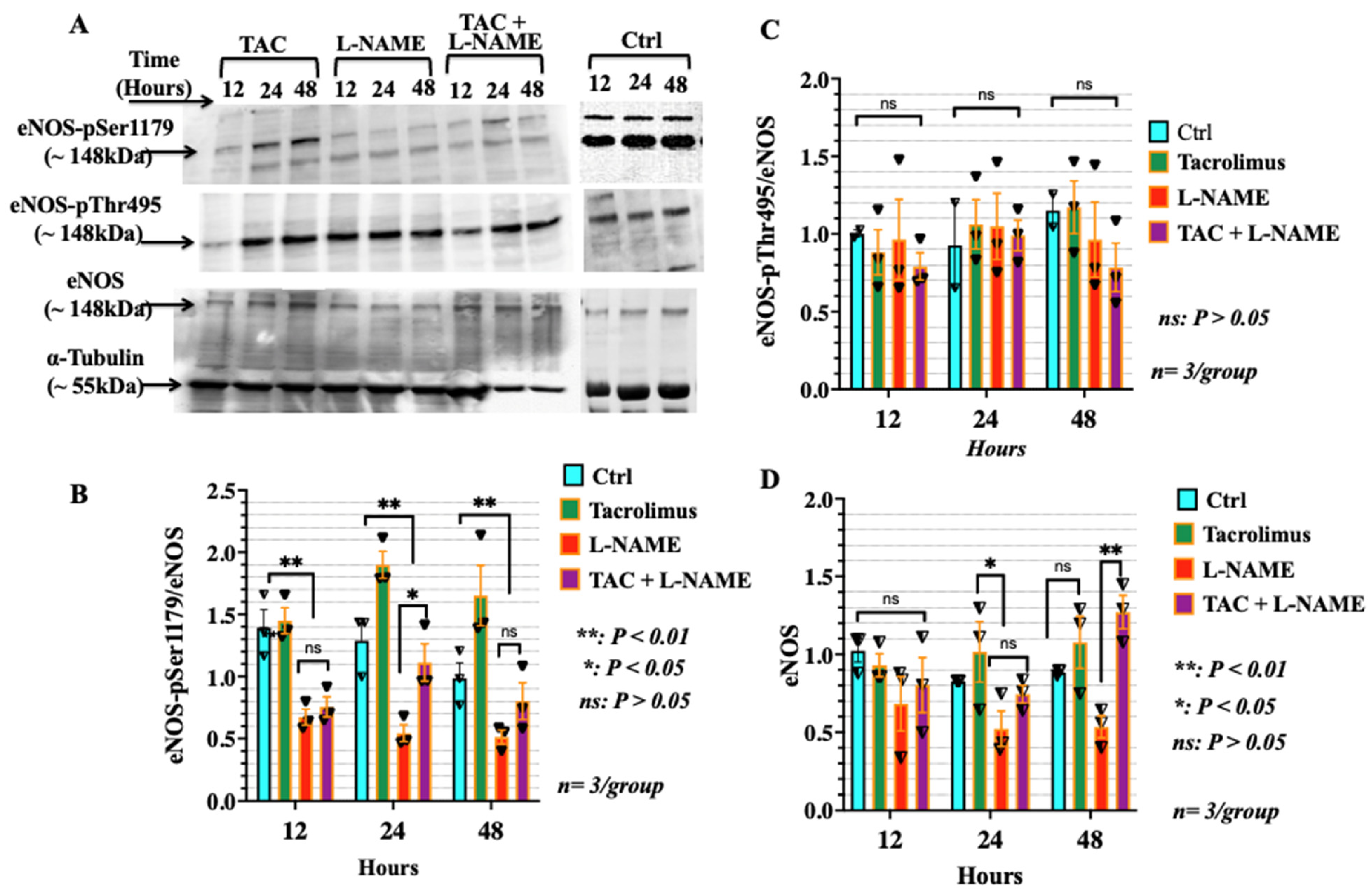
3.5. Low-Dose Tacrolimus Induced the Phosphorylation of the Stimulatory Domain of eNOS (p-Ser1179) in HTR-8/SVneo Cells Irrespective of the Presence of L-NAME
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anin, S.A.; Vince, G.; Quenby, S. Trophoblast invasion. Hum. Fertil. 2004, 7, 169–174. [Google Scholar] [CrossRef]
- Halasz, M.; Szekeres-Bartho, J. The role of progesterone in implantation and trophoblast invasion. J. Reprod. Immunol. 2013, 97, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tranguch, S.; Cheung-Flynn, J.; Daikoku, T.; Prapapanich, V.; Cox, M.B.; Xie, H.; Wang, H.; Das, S.K.; Smith, D.F.; Dey, S.K. Cochaperone immunophilin FKBP52 is critical to uterine receptivity for embryo implantation. Proc. Natl. Acad. Sci. USA 2005, 102, 14326–14331. [Google Scholar] [CrossRef] [Green Version]
- Albaghdadi, A.J.H.; Kan, F.W.K. Immunosuppression with tacrolimus improved implantation and rescued expression of uterine progesterone receptor and its co-regulators FKBP52 and PIASy at nidation in the obese and diabetic mice: Comparative studies with metformin. Mol. Cell Endocrinol. 2018, 460, 73–84. [Google Scholar] [CrossRef]
- Li, H.; Fidler, M.L.; Lim, C.S. Effect of initial subcellular localization of progesterone receptor on import kinetics and transcriptional activity. Mol. Pharm. 2005, 2, 509–518. [Google Scholar] [CrossRef]
- Durlej, M.; Tabarowski, Z.; Slomczynska, M. Immunohistochemical study on differential distribution of progesterone receptor A and progesterone receptor B within the porcine ovary. Anim. Reprod. Sci. 2010, 121, 167–173. [Google Scholar] [CrossRef]
- Myatt, L. Review: Reactive oxygen and nitrogen species and functional adaptation of the placenta. Placenta 2010, 31, S66–S69. [Google Scholar] [CrossRef] [Green Version]
- Gagioti, S.; Scavone, C.; Bevilacqua, E. Participation of the mouse implanting trophoblast in nitric oxide production during pregnancy. Biol. Reprod. 2000, 62, 260–268. [Google Scholar] [CrossRef] [Green Version]
- Lyall, F.; Bulmer, J.N.; Kelly, H.; Duffie, E.; Robson, S.C. Human trophoblast invasion and spiral artery transformation: The role of nitric oxide. Am. J. Pathol. 1999, 154, 1105–1114. [Google Scholar] [CrossRef]
- Yamada, T.; Carson, A.R.; Caniggia, I.; Umebayashi, K.; Yoshimori, T.; Nakabayashi, K.; Scherer, S.W. Endothelial nitric-oxide synthase antisense (NOS3AS) gene encodes an autophagy-related protein (APG9-like2) highly expressed in trophoblast. J. Biol. Chem. 2005, 280, 18283–18290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwedhelm, E.; Maas, R.; Tan-Andresen, J.; Schulze, F.; Riederer, U.; Boger, R.H. High-throughput liquid chromatographic-tandem mass spectrometric determination of arginine and dimethylated arginine derivatives in human and mouse plasma. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2007, 851, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Krause, B.J.; Hanson, M.A.; Casanello, P. Role of nitric oxide in placental vascular development and function. Placenta 2011, 32, 797–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayling, L.J.; Whitley, G.S.J.; Aplin, J.D.; Cartwright, J.E. Dimethylarginine dimethylaminohydrolase (DDAH) regulates trophoblast invasion and motility through effects on nitric oxide. Hum. Reprod. 2006, 21, 2530–2537. [Google Scholar] [CrossRef] [Green Version]
- Harris, L.K.; McCormick, J.; Cartwright, J.E.; Whitley, G.S.; Dash, P.R. S-nitrosylation of proteins at the leading edge of migrating trophoblasts by inducible nitric oxide synthase promotes trophoblast invasion. Exp. Cell Res. 2008, 314, 1765–1776. [Google Scholar] [CrossRef]
- Heiss, E.H.; Dirsch, V.M. Regulation of eNOS enzyme activity by posttranslational modification. Curr. Pharm. Des. 2014, 20, 3503–3513. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I. Molecular mechanisms underlying the activation of eNOS. Pflügers Arch.-Eur. J. Physiol. 2010, 459, 793–806. [Google Scholar] [CrossRef]
- Fulton, D.; Gratton, J.P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 1999, 399, 597–601. [Google Scholar] [CrossRef]
- Quinta, H.R.; Galigniana, M.D. The neuroregenerative mechanism mediated by the Hsp90-binding immunophilin FKBP52 resembles the early steps of neuronal differentiation. Br. J. Pharmacol. 2012, 166, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Albaghdadi, A.J.; Hewitt, M.A.; Putos, S.M.; Wells, M.; Ozolins, T.R.; Kan, F.W. Tacrolimus in the prevention of adverse pregnancy outcomes and diabetes-associated embryopathies in obese and diabetic mice. J. Transl. Med. 2017, 15, 32. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, K.; Kwak-Kim, J.; Ota, K.; Kuroda, K.; Hisano, M.; Sugiyama, R.; Yamaguchi, K. Immunosuppression with Tacrolimus Improved Reproductive Outcome of Women with Repeated Implantation Failure and Elevated Peripheral Blood Th1/Th2 Cell Ratios. Am. J. Reprod. Immunol. 2014, 73, 353–361. [Google Scholar] [CrossRef]
- Nakagawa, K.; Kwak-Kim, J.; Kuroda, K.; Sugiyama, R.; Yamaguchi, K. Immunosuppressive treatment using tacrolimus promotes pregnancy outcome in infertile women with repeated implantation failures. Am. J. Reprod. Immunol. 2017, 78, e12682. [Google Scholar] [CrossRef]
- Bahrami-Asl, Z.; Farzadi, L.; Fattahi, A.; Yousefi, M.; Quinonero, A.; Hakimi, P.; Latifi, Z.; Nejabati, H.R.; Ghasemnejad, T.; Sadigh, A.R.; et al. Tacrolimus Improves the Implantation Rate in Patients with Elevated Th1/2 Helper Cell Ratio and Repeated Implantation Failure (RIF). Geburtshilfe Frauenheilkd. 2020, 80, 851–862. [Google Scholar] [CrossRef]
- Shen, Z.; Song, Q.; Chen, L.; Zhong, B.; Tang, S.; Hao, F. Bidirectional immunoregulation of calcineurin inhibitor tacrolimus on FOXP3 transcription? Med. Hypotheses 2011, 76, 178–180. [Google Scholar] [CrossRef]
- Banerji, S.S.; Parsons, J.N.; Tocci, M.J. The immunosuppressant FK-506 specifically inhibits mitogen-induced activation of the interleukin-2 promoter and the isolated enhancer elements NFIL-2A and NF-AT1. Mol. Cell Biol. 1991, 11, 4074–4087. [Google Scholar]
- Thomson, A.W.; Bonham, C.A.; Zeevi, A. Mode of action of tacrolimus (FK506): Molecular and cellular mechanisms. Ther. Drug Monit. 1995, 17, 584–591. [Google Scholar] [CrossRef]
- Shimamoto, S.; Kubota, Y.; Tokumitsu, H.; Kobayashi, R. S100 proteins regulate the interaction of Hsp90 with Cyclophilin 40 and FKBP52 through their tetratricopeptide repeats. FEBS Lett. 2010, 584, 1119–1125. [Google Scholar] [CrossRef] [Green Version]
- Pirkl, F.; Buchner, J. Functional analysis of the Hsp90-associated human peptidyl prolyl cis/trans isomerases FKBP51, FKBP52 and Cyp40. J. Mol. Biol. 2001, 308, 795–806. [Google Scholar] [CrossRef]
- Cox, M.B.; Riggs, D.L.; Hessling, M.; Schumacher, F.; Buchner, J.; Smith, D.F. FK506-binding protein 52 phosphorylation: A potential mechanism for regulating steroid hormone receptor activity. Mol. Endocrinol. 2007, 21, 2956–2967. [Google Scholar] [CrossRef] [Green Version]
- Cluning, C.; Ward, B.K.; Rea, S.L.; Arulpragasam, A.; Fuller, P.J.; Ratajczak, T. The helix 1-3 loop in the glucocorticoid receptor LBD is a regulatory element for FKBP cochaperones. Mol. Endocrinol. 2013, 27, 1020–1035. [Google Scholar] [CrossRef] [Green Version]
- Mazaira, G.I.; Echeverria, P.C.; Galigniana, M.D. Nucleocytoplasmic shuttling of the glucocorticoid receptor is influenced by tetratricopeptide repeat-containing proteins. J. Cell Sci. 2020, 133, jcs238873. [Google Scholar] [CrossRef]
- Silverstein, A.M.; Galigniana, M.D.; Kanelakis, K.C.; Radanyi, C.; Renoir, J.M.; Pratt, W.B. Different regions of the immunophilin FKBP52 determine its association with the glucocorticoid receptor, hsp90, and cytoplasmic dynein. J. Biol. Chem. 1999, 274, 36980–36986. [Google Scholar] [CrossRef] [Green Version]
- Can, C.; Erol, A.; Cinar, M.; Olukman, M.; Ulker, S.; Evinc, A. Therapeutic concentrations of tacrolimus do not interfere with endothelial nitric oxide synthesis in rat thoracic aortas and coronary arteries. J. Cardiovasc. Pharmacol. 2007, 50, 399–405. [Google Scholar] [CrossRef]
- Cook, L.G.; Chiasson, V.L.; Long, C.; Wu, G.Y.; Mitchell, B.M. Tacrolimus reduces nitric oxide synthase function by binding to FKBP rather than by its calcineurin effect. Kidney Int. 2009, 75, 719–726. [Google Scholar] [CrossRef] [Green Version]
- Tepperman, E.D.; Tumiati, L.C.; Ramzy, D.; Badiwala, M.V.; Sheshgiri, R.; Prodger, J.L.; Ross, H.J.; Rao, V. Tacrolimus preserves vasomotor function and maintains vascular homeostasis. J. Heart Lung Transplant. 2011, 30, 583–588. [Google Scholar] [CrossRef]
- Salomon, C.; Yee, S.; Scholz-Romero, K.; Kobayashi, M.; Vaswani, K.; Kvaskoff, D.; Illanes, S.E.; Mitchell, M.D.; Rice, G.E. Extravillous trophoblast cells-derived exosomes promote vascular smooth muscle cell migration. Front. Pharmacol. 2014, 5, 175. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, M.; Mukosera, G.T.; Borchardt, D.; Li, Q.; Tipple, T.E.; Ishtiaq Ahmed, A.S.; Power, G.G.; Blood, A.B. L-NAME releases nitric oxide and potentiates subsequent nitroglycerin-mediated vasodilation. Redox Biol. 2019, 26, 101238. [Google Scholar] [CrossRef]
- Bottiger, Y.; Brattstrom, C.; Tyden, G.; Sawe, J.; Groth, C.G. Tacrolimus whole blood concentrations correlate closely to side-effects in renal transplant recipients. Br. J. Clin. Pharmacol. 1999, 48, 445–448. [Google Scholar] [CrossRef]
- Casanello, P.; Krause, B.; Torres, E.; Gallardo, V.; Gonzalez, M.; Prieto, C.; Escudero, C.; Farias, M.; Sobrevia, L. Reduced l-arginine transport and nitric oxide synthesis in human umbilical vein endothelial cells from intrauterine growth restriction pregnancies is not further altered by hypoxia. Placenta 2009, 30, 625–633. [Google Scholar] [CrossRef]
- Yallampalli, C.; Garfield, R.E. Inhibition of Nitric-Oxide Synthesis in Rats during Pregnancy Produces Signs Similar to Those of Preeclampsia. Am. J. Obstet. Gynecol. 1993, 169, 1316–1320. [Google Scholar] [CrossRef]
- Koh, Y.Q.; Peiris, H.N.; Vaswani, K.; Reed, S.; Rice, G.E.; Salomon, C.; Mitchell, M.D. Characterization of exosomal release in bovine endometrial intercaruncular stromal cells. Reprod. Biol. Endocrinol. 2016, 14, 78. [Google Scholar] [CrossRef] [Green Version]
- San Martin, S.; Poehlmann, T.; Codina-Canet, C.; Markert, U.R. Immunohistochemical analysis of trophoblastic cells invading through Matrigel. Placenta 2008, 29, 982–984. [Google Scholar] [CrossRef]
- Namin, S.M.; Nofallah, S.; Joshi, M.S.; Kavallieratos, K.; Tsoukias, N.M. Kinetic analysis of DAF-FM activation by NO: Toward calibration of a NO-sensitive fluorescent dye. Nitric Oxide 2013, 28, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; He, P. Improved measurements of intracellular nitric oxide in intact microvessels using 4,5-diaminofluorescein diacetate. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H108–H114. [Google Scholar] [CrossRef]
- Kumar, N.S.; Richer, J.; Owen, G.; Litman, E.; Horwitz, K.B.; Leslie, K.K. Selective down-regulation of progesterone receptor isoform B in poorly differentiated human endometrial cancer cells: Implications for unopposed estrogen action. Cancer Res. 1998, 58, 1860–1865. [Google Scholar]
- Nyholm, H.C.; Nielsen, A.L.; Norup, P. Endometrial cancer in postmenopausal women with and without previous estrogen replacement treatment: Comparison of clinical and histopathological characteristics. Gynecol. Oncol. 1993, 49, 229–235. [Google Scholar] [CrossRef]
- Chakraborty, C.; Gleeson, L.M.; McKinnon, T.; Lala, P.K. Regulation of human trophoblast migration and invasiveness. Can. J. Physiol. Pharmacol. 2002, 80, 116–124. [Google Scholar] [CrossRef]
- Halasz, M.; Polgar, B.; Berta, G.; Czimbalek, L.; Szekeres-Bartho, J. Progesterone-induced blocking factor differentially regulates trophoblast and tumor invasion by altering matrix metalloproteinase activity. Cell Mol. Life Sci. 2013, 70, 4617–4630. [Google Scholar] [CrossRef] [Green Version]
- Corvinus, F.M.; Fitzgerald, J.S.; Friedrich, K.; Markert, U.R. Evidence for a correlation between trophoblast invasiveness and STAT3 activity. Am. J. Reprod. Immunol. 2003, 50, 316–321. [Google Scholar] [CrossRef]
- Choe, J.Y.; Park, K.Y.; Park, S.H.; Lee, S.I.; Kim, S.K. Regulatory effect of calcineurin inhibitor, tacrolimus, on IL-6/sIL-6R-mediated RANKL expression through JAK2-STAT3-SOCS3 signaling pathway in fibroblast-like synoviocytes. Arthritis Res. Ther. 2013, 15, R26. [Google Scholar] [CrossRef] [Green Version]
- Davies, T.H.; Ning, Y.M.; Sanchez, E.R. Differential control of glucocorticoid receptor hormone-binding function by tetratricopeptide repeat (TPR) proteins and the immunosuppressive ligand FK506. Biochemistry 2005, 44, 2030–2038. [Google Scholar] [CrossRef] [Green Version]
- Acar, N.; Ustunel, I. Expression of 52-kDa FK506-binding protein (FKBP52) in human placenta complicated by preeclampsia and intrauterine growth restriction. Anal. Quant. Cytopathol. Histopathol. 2015, 37, 87–95. [Google Scholar]
- Holcberg, G.; Kossenjans, W.; Miodovnik, M.; Myatt, L. The Interaction of Nitric-Oxide and Superoxide in the Human Fetal-Placental Vasculature. Am. J. Obstet. Gynecol. 1995, 173, 528–533. [Google Scholar] [CrossRef]
- Herr, F.; Baal, N.; Widmer-Teske, R.; McKinnon, T.; Zygmunt, M. How to study placental vascular development? Theriogenology 2010, 73, 817–827. [Google Scholar] [CrossRef]
- Burton, G.J.; Charnock-Jones, D.S.; Jauniaux, E. Regulation of vascular growth and function in the human placenta. Reproduction 2009, 138, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Bernatova, I.; Pechanova, O.; Simko, F. Captopril prevents NO-deficient hypertension and left ventricular hypertrophy without affecting nitric oxide synthase activity in rats. Physiol. Res. 1996, 45, 311–316. [Google Scholar]
- Babbedge, R.C.; Bland-Ward, P.A.; Hart, S.L.; Moore, P.K. Inhibition of rat cerebellar nitric oxide synthase by 7-nitro indazole and related substituted indazoles. Br. J. Pharmacol. 1993, 110, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Vitecek, J.; Lojek, A.; Valacchi, G.; Kubala, L. Arginine-based inhibitors of nitric oxide synthase: Therapeutic potential and challenges. Mediat. Inflamm. 2012, 2012, 318087. [Google Scholar] [CrossRef] [Green Version]
- Alderton, W.K.; Cooper, C.E.; Knowles, R.G. Nitric oxide synthases: Structure, function and inhibition. Biochem. J. 2001, 357, 593–615. [Google Scholar] [CrossRef]
- Kissinger, C.R.; Parge, H.E.; Knighton, D.R.; Lewis, C.T.; Pelletier, L.A.; Tempczyk, A.; Kalish, V.J.; Tucker, K.D.; Showalter, R.E.; Moomaw, E.W.; et al. Crystal structures of human calcineurin and the human FKBP12-FK506-calcineurin complex. Nature 1995, 378, 641–644. [Google Scholar] [CrossRef]
- Callsen, D.; Sandau, K.B.; Brune, B. Nitric oxide and superoxide inhibit platelet-derived growth factor receptor phosphotyrosine phosphatases. Free Radic. Biol. Med. 1999, 26, 1544–1553. [Google Scholar] [CrossRef]
- Ruan, L.; Torres, C.M.; Buffett, R.J.; Kennard, S.; Fulton, D.; Venema, R.C. Calcineurin-mediated dephosphorylation of eNOS at serine 116 affects eNOS enzymatic activity indirectly by facilitating c-Src binding and tyrosine 83 phosphorylation. Vasc. Pharmacol. 2013, 59, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsuyama, T.; Hidaka, K.; Furuno, T.; Hara, N. Neutrophil-induced endothelial cell damage: Inhibition by a 14-membered ring macrolide through the action of nitric oxide. Int. Arch. Allergy Immunol. 1997, 114, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Balibrea, J.M.; Garcia-Martin, M.C.; Cuesta-Sancho, S.; Olmedilla, Y.; Arias-Diaz, J.; Fernandez-Sevilla, E.; Vara, E.; Balibrea, J.L. Tacrolimus modulates liver and pancreas nitric oxide synthetase and heme-oxygenase isoforms and cytokine production after endotoxemia. Nitric Oxide 2011, 24, 113–122. [Google Scholar] [CrossRef]
- Saura, M.; Zaragoza, C.; Bao, C.; Herranz, B.; Rodriguez-Puyol, M.; Lowenstein, C.J. Stat3 mediates interleukin-6 [correction of interelukin-6] inhibition of human endothelial nitric-oxide synthase expression. J. Biol. Chem. 2006, 281, 30057–30062. [Google Scholar] [CrossRef] [Green Version]
- Chiasson, V.L.; Talreja, D.; Young, K.J.; Chatterjee, P.; Banes-Berceli, A.K.; Mitchell, B.M. FK506 binding protein 12 deficiency in endothelial and hematopoietic cells decreases regulatory T cells and causes hypertension. Hypertension 2011, 57, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Kim, J.; Kim, J.H.; Lee, D.K.; Park, W.; Park, M.; Kim, S.; Hwang, J.Y.; Won, M.H.; Choi, Y.K.; et al. Carbon monoxide prevents TNF-alpha-induced eNOS downregulation by inhibiting NF-kappaB-responsive miR-155-5p biogenesis. Exp. Mol. Med. 2017, 49, e403. [Google Scholar] [CrossRef] [Green Version]
- Myatt, L.; Eis, A.L.; Brockman, D.E.; Kossenjans, W.; Greer, I.; Lyall, F. Inducible (type II) nitric oxide synthase in human placental villous tissue of normotensive, pre-eclamptic and intrauterine growth-restricted pregnancies. Placenta 1997, 18, 261–268. [Google Scholar] [CrossRef]
- Giannubilo, S.R.; Menegazzi, M.; Tedeschi, E.; Bezzeccheri, V.; Suzuki, H.; Tranquilli, A.L. Doppler analysis and placental nitric oxide synthase expression during fetal growth restriction. J. Matern.-Fetal Neonatal Med. 2008, 21, 617–622. [Google Scholar] [CrossRef]
- Wang, Y.P.; Gu, Y.; Zhang, Y.P.; Lewis, D.F. Evidence of endothetial dysfunction in preeclampsia: Decreased endothelial nitric oxide synthase expression is associated with increased cell permeability in endothelial cells from preeclampsia. Am. J. Obstet. Gynecol. 2004, 190, 817–824. [Google Scholar] [CrossRef]
- Rutherford, R.A.; McCarthy, A.; Sullivan, M.H.; Elder, M.G.; Polak, J.M.; Wharton, J. Nitric oxide synthase in human placenta and umbilical cord from normal, intrauterine growth-retarded and pre-eclamptic pregnancies. Br. J. Pharmacol. 1995, 116, 3099–3109. [Google Scholar] [CrossRef] [Green Version]
- Lyall, F.; Greer, I.A.; Young, A.; Myatt, L. Nitric oxide concentrations are increased in the feto-placental circulation in intrauterine growth restriction. Placenta 1996, 17, 165–168. [Google Scholar] [CrossRef]
- Tikvica, A.; Jukic, M.K.; Pintaric, I.; Medic, M.; Hudicek-Martincic, G.; Kosec, V.; Salihagic-Kadic, A. Nitric oxide synthesis in placenta is increased in intrauterine growth restriction and fetal hypoxia. Coll. Antropol. 2008, 32, 565–570. [Google Scholar]
- Banerjee, A.; Periyasamy, S.; Wolf, I.M.; Hinds, T.D., Jr.; Yong, W.; Shou, W.; Sanchez, E.R. Control of glucocorticoid and progesterone receptor subcellular localization by the ligand-binding domain is mediated by distinct interactions with tetratricopeptide repeat proteins. Biochemistry 2008, 47, 10471–10480. [Google Scholar] [CrossRef] [Green Version]
- Barent, R.L.; Nair, S.C.; Carr, D.C.; Ruan, Y.; Rimerman, R.A.; Fulton, J.; Zhang, Y.; Smith, D.F. Analysis of FKBP51/FKBP52 chimeras and mutants for Hsp90 binding and association with progesterone receptor complexes. Mol. Endocrinol. 1998, 12, 342–354. [Google Scholar] [CrossRef]
- Storer, C.L.; Dickey, C.A.; Galigniana, M.D.; Rein, T.; Cox, M.B. FKBP51 and FKBP52 in signaling and disease. Trends Endocrinol. Metab. 2011, 22, 481–490. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, H.C.; Versteegh, J.E.; Uitdehaag, J.C. The X-ray structure of RU486 bound to the progesterone receptor in a destabilized agonistic conformation. J. Biol. Chem. 2009, 284, 19572–19579. [Google Scholar] [CrossRef] [Green Version]
- Cadepond, F.; Ulmann, A.; Baulieu, E.E. RU486 (mifepristone): Mechanisms of action and clinical uses. Annu. Rev. Med. 1997, 48, 129–156. [Google Scholar] [CrossRef] [Green Version]
- Terakawa, N.; Shimizu, I.; Tanizawa, O.; Matsumoto, K. RU486, a progestin antagonist, binds to progesterone receptors in a human endometrial cancer cell line and reverses the growth inhibition by progestins. J. Steroid Biochem. 1988, 31, 161–166. [Google Scholar] [CrossRef]
- Zong, S.S.; Li, C.Q.; Luo, C.F.; Zhao, X.; Liu, C.H.; Wang, K.; Jia, W.W.; Bai, M.L.; Yin, M.H.; Bao, S.H.; et al. Dysregulated expression of IDO may cause unexplained recurrent spontaneous abortion through suppression of trophoblast cell proliferation and migration. Sci. Rep. 2016, 6, 19916. [Google Scholar] [CrossRef] [Green Version]
- Stelzer, I.A.; Mori, M.; DeMayo, F.; Lydon, J.; Arck, P.C.; Solano, M.E. Differential mouse-strain specific expression of Junctional Adhesion Molecule (JAM)-B in placental structures. Cell Adhes. Migr. 2016, 10, 2–17. [Google Scholar] [CrossRef] [Green Version]
- Poehlmann, T.G.; Fitzgerald, J.S.; Meissner, A.; Wengenmayer, T.; Schleussner, E.; Friedrich, K.; Markert, U.R. Trophoblast invasion: Tuning through LIF, signalling via Stat3. Placenta 2005, 26 (Suppl. A), S37–S41. [Google Scholar] [CrossRef]
- Kim, T.J.; Kim, N.; Kang, H.J.; Kim, E.O.; Kim, S.T.; Ahn, H.S.; Bluestone, J.A.; Lee, K.M. FK506 causes cellular and functional defects in human natural killer cells. J. Leukoc. Biol. 2010, 88, 1089–1097. [Google Scholar] [CrossRef]
- Liang, H.; Venema, V.J.; Wang, X.; Ju, H.; Venema, R.C.; Marrero, M.B. Regulation of angiotensin II-induced phosphorylation of STAT3 in vascular smooth muscle cells. J. Biol. Chem. 1999, 274, 19846–19851. [Google Scholar] [CrossRef] [Green Version]
- Hirano, T.; Ishihara, K.; Hibi, M. Roles of STAT3 in mediating the cell growth, differentiation and survival signals relayed through the IL-6 family of cytokine receptors. Oncogene 2000, 19, 2548–2556. [Google Scholar] [CrossRef]
- Fu, A.K.; Fu, W.Y.; Ng, A.K.; Chien, W.W.; Ng, Y.P.; Wang, J.H.; Ip, N.Y. Cyclin-dependent kinase 5 phosphorylates signal transducer and activator of transcription 3 and regulates its transcriptional activity. Proc. Natl. Acad. Sci. USA 2004, 101, 6728–6733. [Google Scholar] [CrossRef] [Green Version]
- Krasilnikov, M.; Ivanov, V.N.; Dong, J.; Ronai, Z. ERK and PI3K negatively regulate STAT-transcriptional activities in human melanoma cells: Implications towards sensitization to apoptosis. Oncogene 2003, 22, 4092–4101. [Google Scholar] [CrossRef] [Green Version]
- Beguelin, W.; Diaz Flaque, M.C.; Proietti, C.J.; Cayrol, F.; Rivas, M.A.; Tkach, M.; Rosemblit, C.; Tocci, J.M.; Charreau, E.H.; Schillaci, R.; et al. Progesterone receptor induces ErbB-2 nuclear translocation to promote breast cancer growth via a novel transcriptional effect: ErbB-2 function as a coactivator of Stat3. Mol. Cell Biol. 2010, 30, 5456–5472. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.L.; Guan, Y.J.; Chatterjee, D.; Chin, Y.E. Stat3 dimerization regulated by reversible acetylation of a single lysine residue. Science 2005, 307, 269–273. [Google Scholar] [CrossRef]
- Mertens, C.; Darnell Jr, J.E. Snapshot: Jak-stat signaling. Cell 2007, 131, 612–612.e611. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, X.; Wang, J.; Zhang, L. The decreased expression of Stat3 and p-Stat3 in preeclampsia-like rat placenta. J. Mol. Histol. 2018, 49, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Park, M.N.; Park, K.H.; Lee, J.E.; Shin, Y.Y.; An, S.M.; Kang, S.S.; Cho, W.S.; An, B.S.; Kim, S.C. The expression and activation of sex steroid receptors in the preeclamptic placenta. Int. J. Mol. Med. 2018, 41, 2943–2951. [Google Scholar] [CrossRef] [PubMed]
- Weber, M.; Knoefler, I.; Schleussner, E.; Markert, U.R.; Fitzgerald, J.S. HTR-8/SVneo cells display trophoblast progenitor cell-like characteristics indicative of self-renewal, repopulation activity, and expression of “stemness-” associated transcription factors. Biomed. Res. Int. 2013, 2013, 243649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastuschek, J.; Nonn, O.; Gutierrez-Samudio, R.N.; Murrieta-Coxca, J.M.; Muller, J.; Sanft, J.; Huppertz, B.; Markert, U.R.; Groten, T.; Morales-Prieto, D.M. Molecular characteristics of established trophoblast-derived cell lines. Placenta 2021, 108, 122–133. [Google Scholar] [CrossRef]
- Akgun, S.; Tekeli, A.; Kurtkaya, O.; Civelek, A.; Isbir, S.C.; Ak, K.; Arsan, S.; Sav, A. Neuroprotective effects of FK-506, L-carnitine and azathioprine on spinal cord ischemia-reperfusion injury. Eur. J. Cardio-Thorac. Surg. 2004, 25, 105–110. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albaghdadi, A.J.H.; Coyle, K.; Kan, F.W.K. Low-Dose Tacrolimus Promotes the Migration and Invasion and Nitric Oxide Production in the Human-Derived First Trimester Extravillous Trophoblast Cells In Vitro. Int. J. Mol. Sci. 2022, 23, 8426. https://doi.org/10.3390/ijms23158426
Albaghdadi AJH, Coyle K, Kan FWK. Low-Dose Tacrolimus Promotes the Migration and Invasion and Nitric Oxide Production in the Human-Derived First Trimester Extravillous Trophoblast Cells In Vitro. International Journal of Molecular Sciences. 2022; 23(15):8426. https://doi.org/10.3390/ijms23158426
Chicago/Turabian StyleAlbaghdadi, Ahmad J. H., Kassandra Coyle, and Frederick W. K. Kan. 2022. "Low-Dose Tacrolimus Promotes the Migration and Invasion and Nitric Oxide Production in the Human-Derived First Trimester Extravillous Trophoblast Cells In Vitro" International Journal of Molecular Sciences 23, no. 15: 8426. https://doi.org/10.3390/ijms23158426