
Functional Characterization of Human Induced Pluripotent Stem Cell-Derived Endothelial Cells

 , , , and
, , , and 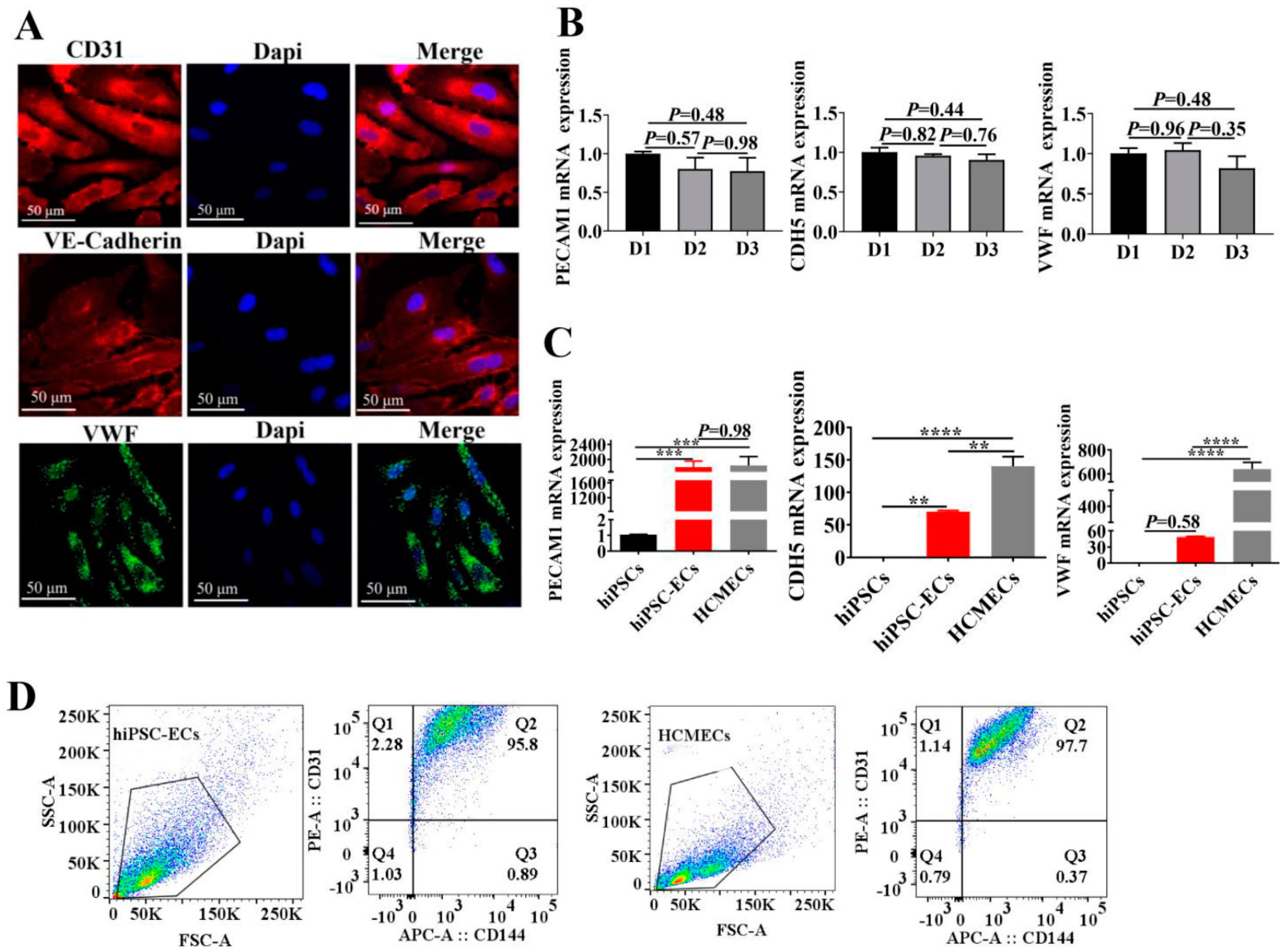{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
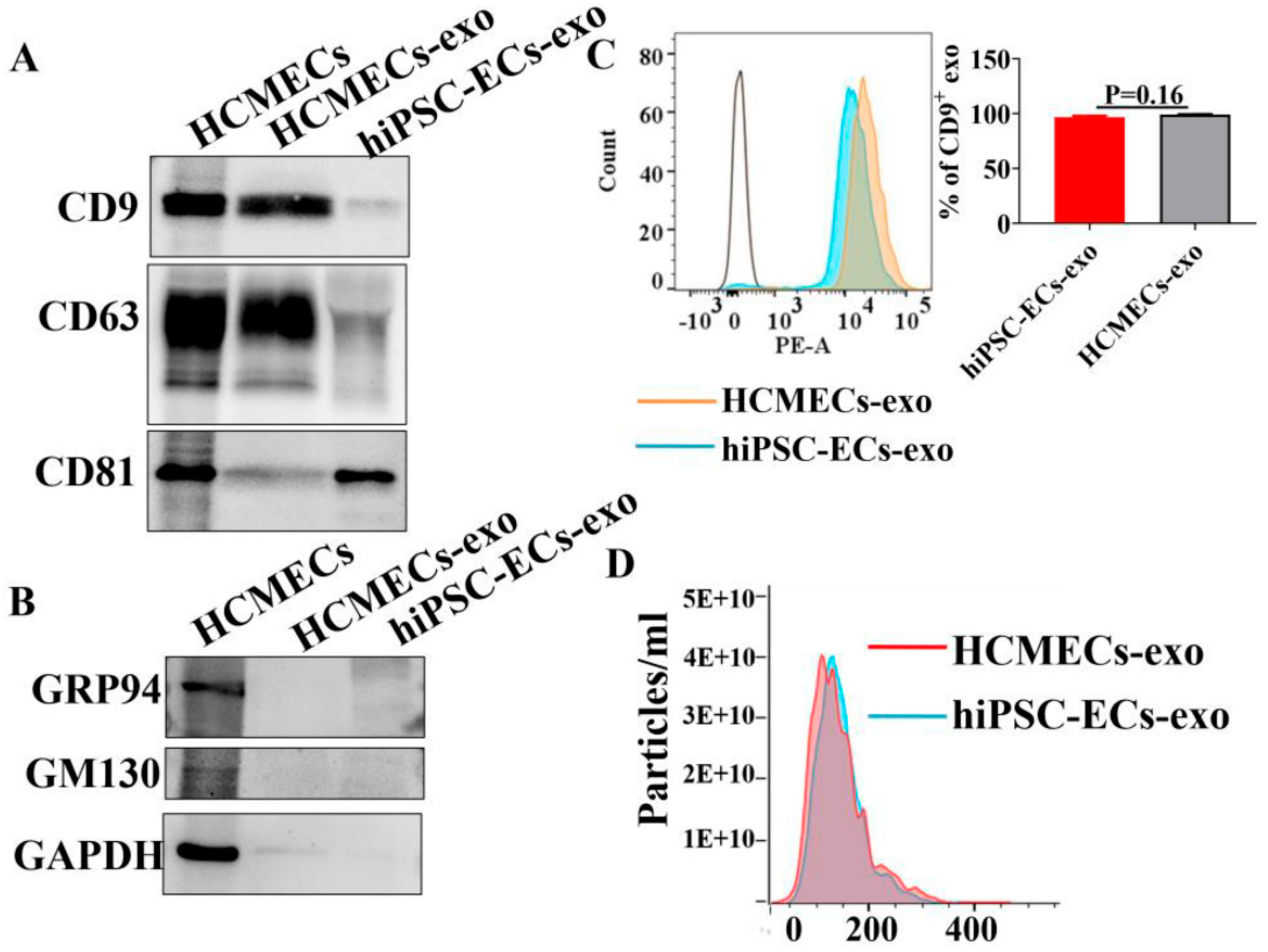
2.1. Characterizations of hiPSC-ECs
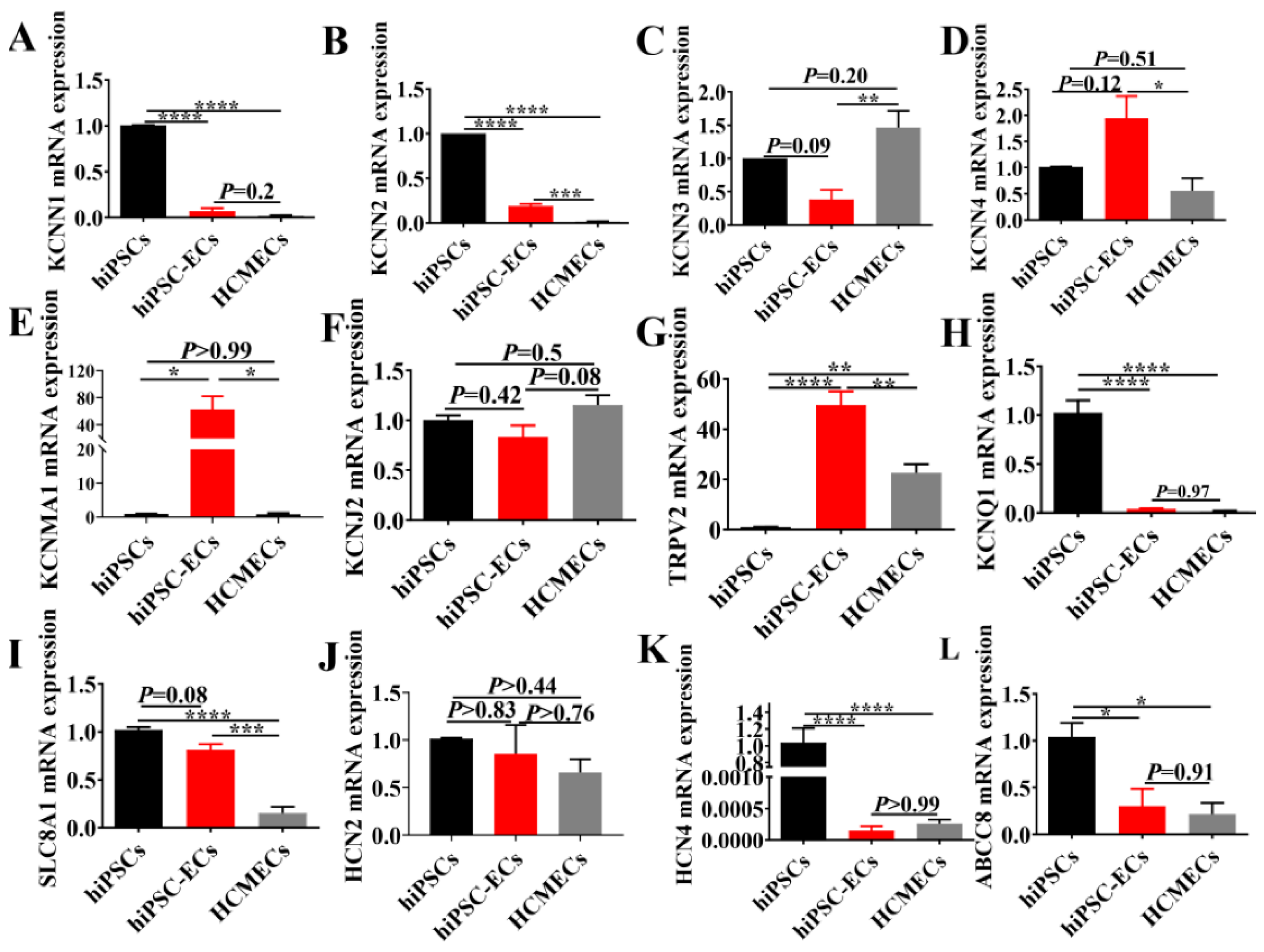
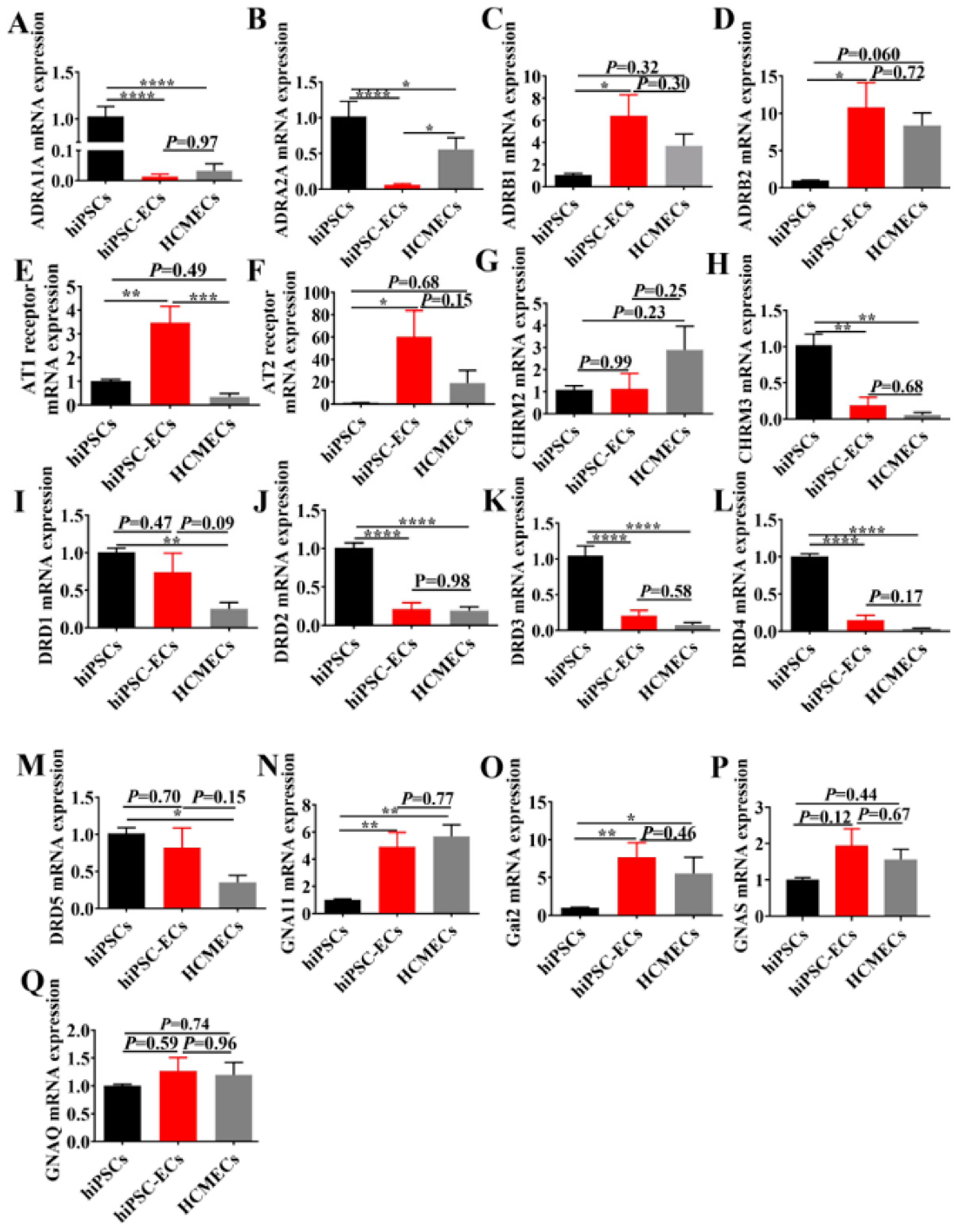
2.2. Gene Expression of Ion Channel and Membrane Receptors in hiPSC-ECs
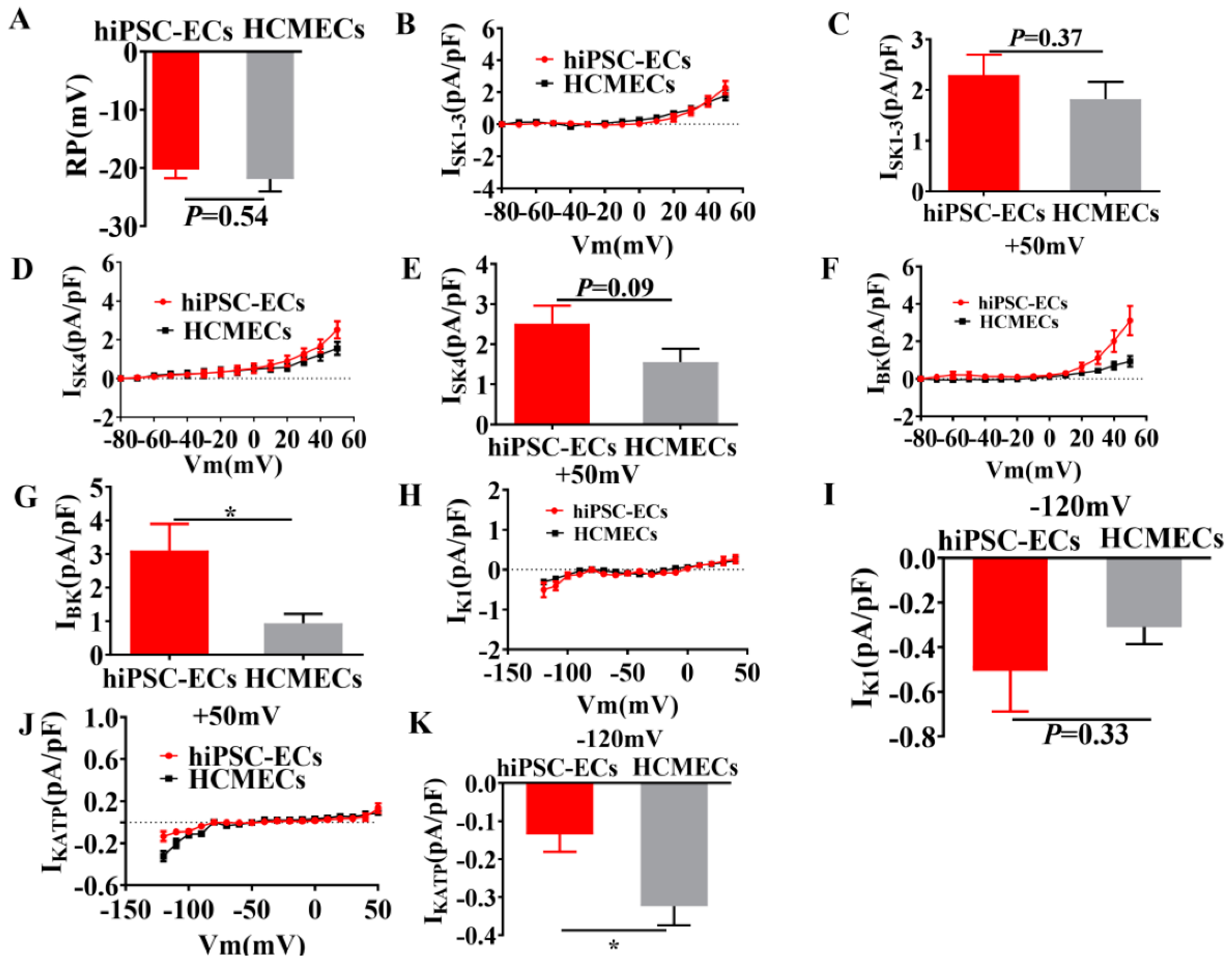
2.3. Membrane Potentials and Ion Channel Currents in hiPSC-ECs and HCMECs
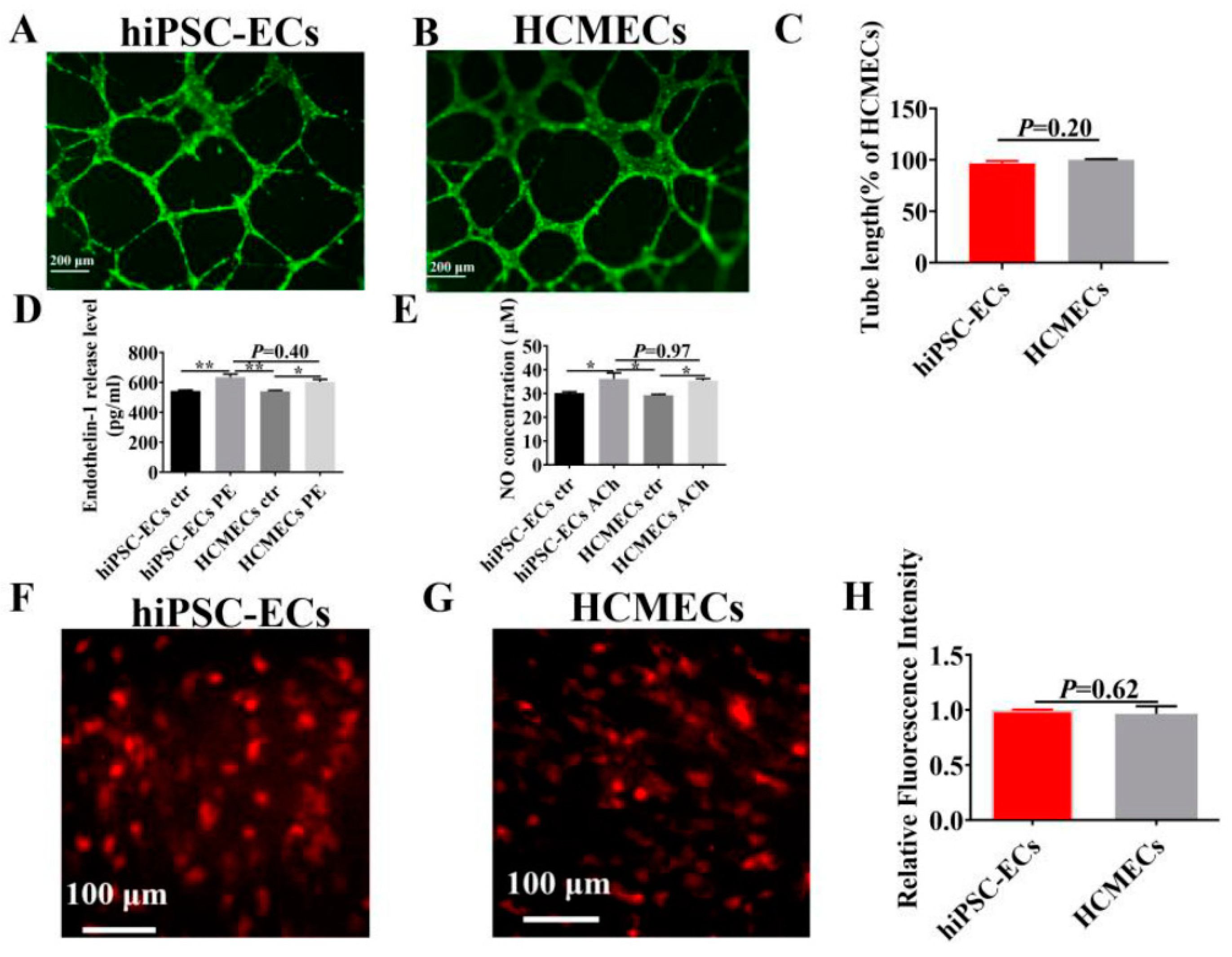
2.4. Functional Characterization of hiPSC-ECs
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Generation of Human-Induced Pluripotent Stem Cells
4.3. Cell Culture
4.4. Generation of hiPSC-ECs
4.5. Identification of HCMECS and hiPSC-ECs
4.6. Immunofluorescence (IF) Staining
4.7. Polymerase-Chain-Reaction Assays
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Vascular Tube Formation Assay
4.10. Low Density Lipoprotein (LDL) Uptake Assay
4.11. Patch-Clamp
4.12. Isolation and Quantification of Exosomes
4.13. Flow Cytometry Analysis for Exosomes Derived from HCMECS and hiPSC-ECs
4.14. Western Blot Analysis
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, Y.; Gil, C.H.; Yoder, M.C. Differentiation, Evaluation, and Application of Human Induced Pluripotent Stem Cell-Derived Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2014–2025. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.K.; Canfield, S.G.; Shusta, E.V.; Palecek, S.P. Concise review: Tissue-specific microvascular endothelial cells derived from human pluripotent stem cells. Stem Cells 2014, 32, 3037–3045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoder, M.C. Differentiation of pluripotent stem cells into endothelial cells. Curr. Opin. Hematol. 2015, 22, 252–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M. Efficient Differentiation of Human Pluripotent Stem Cells to Endothelial Cells. Curr. Protoc. Hum. Genet. 2018, 98, e64. [Google Scholar] [CrossRef]
- Zhang, J.; Chu, L.F.; Hou, Z.; Schwartz, M.P.; Hacker, T.; Vickerman, V.; Swanson, S.; Leng, N.; Nguyen, B.K.; Elwell, A.; et al. Functional characterization of human pluripotent stem cell-derived arterial endothelial cells. Proc. Natl. Acad. Sci. USA 2017, 114, E6072–E6078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.H.; Ong, S.G.; Zhou, Y.; Tian, L.; Bae, H.R.; Baker, N.; Whitlatch, A.; Mohammadi, L.; Guo, H.; Nadeau, K.C.; et al. Modeling Cardiovascular Risks of E-Cigarettes with Human-Induced Pluripotent Stem Cell-Derived Endothelial Cells. J. Am. Coll. Cardiol. 2019, 73, 2722–2737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Qi, J.; Xu, X.; Zeisberg, M.; Guan, K.; Zeisberg, E.M. Differentiation of functional endothelial cells from human induced pluripotent stem cells: A novel, highly efficient and cost effective method. Differentiation 2016, 92, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Guo, X.; Ni, Q.; Wang, H.; Ye, M.; Zhang, L. SREBP1 suppresses the differentiation and epithelial function of hiPSC-derived endothelial cells by inhibiting the microRNA199b-5p pathway. Stem Cell Res. 2021, 51, 102174. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Jun Koh, Y.; Jeon, J.; Cho, Y.H.; Jang, M.J.; Kang, Y.; Kim, M.J.; Choi, C.; Sook Cho, Y.; Chung, H.M.; et al. Efficient differentiation of human pluripotent stem cells into functional CD34+ progenitor cells by combined modulation of the MEK/ERK and BMP4 signaling pathways. Blood 2010, 116, 5762–5772. [Google Scholar] [CrossRef] [Green Version]
- Sahara, M.; Hansson, E.M.; Wernet, O.; Lui, K.O.; Spater, D.; Chien, K.R. Manipulation of a VEGF-Notch signaling circuit drives formation of functional vascular endothelial progenitors from human pluripotent stem cells. Cell Res. 2014, 24, 820–841. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Huang, W.; Cai, W.; Wang, L.; Guo, L.; Paul, C.; Yu, X.Y.; Wang, Y. Inhibition of microRNA-495 Enhances Therapeutic Angiogenesis of Human Induced Pluripotent Stem Cells. Stem Cells 2017, 35, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Dutton, J.R.; Su, L.; Zhang, J.; Ye, L. The influence of a spatiotemporal 3D environment on endothelial cell differentiation of human induced pluripotent stem cells. Biomaterials 2014, 35, 3786–3793. [Google Scholar] [CrossRef] [Green Version]
- Su, L.; Kong, X.; Lim, S.; Loo, S.; Tan, S.; Poh, K.; Dutton, J.; Stewart, C.; Cook, S.; Su, X.; et al. The prostaglandin H2 analog U-46619 improves the differentiation efficiency of human induced pluripotent stem cells into endothelial cells by activating both p38MAPK and ERK1/2 signaling pathways. Stem Cell Res. Ther. 2018, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Vila-Gonzalez, M.; Kelaini, S.; Magee, C.; Caines, R.; Campbell, D.; Eleftheriadou, M.; Cochrane, A.; Drehmer, D.; Tsifaki, M.; O’Neill, K.; et al. Enhanced Function of Induced Pluripotent Stem Cell-Derived Endothelial Cells Through ESM1 Signaling. Stem Cells 2019, 37, 226–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Shi, X.; Lin, Y.; Yuan, Y.; Kural, M.H.; Wang, J.; Ellis, M.W.; Anderson, C.W.; Zhang, S.M.; Riaz, M.; et al. Efficient Differentiation of Human Induced Pluripotent Stem Cells into Endothelial Cells under Xenogeneic-free Conditions for Vascular Tissue Engineering. Acta Biomater. 2021, 119, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Tiemeier, G.L.; Wang, G.; Dumas, S.J.; Sol, W.; Avramut, M.C.; Karakach, T.; Orlova, V.V.; van den Berg, C.W.; Mummery, C.L.; Carmeliet, P.; et al. Closing the Mitochondrial Permeability Transition Pore in hiPSC-Derived Endothelial Cells Induces Glycocalyx Formation and Functional Maturation. Stem Cell Rep. 2019, 13, 803–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufaihah, A.J.; Huang, N.F.; Jame, S.; Lee, J.C.; Nguyen, H.N.; Byers, B.; De, A.; Okogbaa, J.; Rollins, M.; Reijo-Pera, R.; et al. Endothelial cells derived from human iPSCS increase capillary density and improve perfusion in a mouse model of peripheral arterial disease. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e72–e79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Wang, L.; Wei, Y.; Krishnamurthy, P.; Walcott, G.P.; Menasche, P.; Zhang, J. Exosomes secreted by hiPSC-derived cardiac cells improve recovery from myocardial infarction in swine. Sci. Transl. Med. 2020, 12, eaay1318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, L.; Li, Y.; Liu, W.; Duan, F.; Huang, R.; Chen, X.; Chang, S.C.; Du, Y.; Na, J. Optimizing mesoderm progenitor selection and three-dimensional microniche culture allows highly efficient endothelial differentiation and ischemic tissue repair from human pluripotent stem cells. Stem Cell Res. Ther. 2017, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, H.; Macklin, B.L.; Lin, Y.Y.; Zhou, L.; Lai, M.J.; Lee, G.; Gerecht, S.; Duh, E.J. iPSC-derived endothelial cell response to hypoxia via SDF1a/CXCR4 axis facilitates incorporation to revascularize ischemic retina. JCI Insight 2020, 5, e131828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, M.; Ni, Q.; Qi, H.; Qian, X.; Chen, J.; Guo, X.; Li, M.; Zhao, Y.; Xue, G.; Deng, H.; et al. Exosomes Derived from Human Induced Pluripotent Stem Cells-Endothelia Cells Promotes Postnatal Angiogenesis in Mice Bearing Ischemic Limbs. Int. J. Biol. Sci. 2019, 15, 158–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.; Aggarwal, P.; Matter, A.; Olson, B.; Gu, C.C.; Hunt, S.C.; Lewis, C.E.; Arnett, D.K.; Lorier, R.; Broeckel, U. Donor-specific phenotypic variation in hiPSC cardiomyocyte-derived exosomes impacts endothelial cell function. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H954–H968. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Gao, Y.; Xu, L.; Dang, W.; Yan, H.; Zou, D.; Zhu, Z.; Luo, L.; Tian, N.; Wang, X.; et al. Exosomes from high glucose-treated glomerular endothelial cells trigger the epithelial-mesenchymal transition and dysfunction of podocytes. Sci. Rep. 2017, 7, 9371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kefaloyianni, E.; Coetzee, W.A. Transcriptional remodeling of ion channel subunits by flow adaptation in human coronary artery endothelial cells. J. Vasc. Res. 2011, 48, 357–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xie, A.; Singh, A.K.; Ehsan, A.; Choudhary, G.; Dudley, S.; Sellke, F.W.; Feng, J. Inactivation of Endothelial Small/Intermediate Conductance of Calcium-Activated Potassium Channels Contributes to Coronary Arteriolar Dysfunction in Diabetic Patients. J. Am. Heart Assoc. 2015, 4, e002062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Shi, G.; Liu, Y.; Xing, H.; Kabakov, A.Y.; Zhao, A.S.; Agbortoko, V.; Kim, J.; Singh, A.K.; Koren, G.; et al. Coronary endothelial dysfunction prevented by small-conductance calcium-activated potassium channel activator in mice and patients with diabetes. J. Thorac. Cardiovasc. Surg. 2020, 160, e263–e280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wulff, H.; Kohler, R. Endothelial small-conductance and intermediate-conductance KCa channels: An update on their pharmacology and usefulness as cardiovascular targets. J. Cardiovasc. Pharmacol. 2013, 61, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Huang, J.H.; Man, Y.B.; Yao, X.Q.; He, G.W. Use of intermediate/small conductance calcium-activated potassium-channel activator for endothelial protection. J. Thorac. Cardiovasc. Surg. 2011, 141, 501–510.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comerma-Steffensen, S.G.; Carvacho, I.; Hedegaard, E.R.; Simonsen, U. Small and Intermediate Calcium-Activated Potassium Channel Openers Improve Rat Endothelial and Erectile Function. Front. Pharmacol. 2017, 8, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zyrianova, T.; Lopez, B.; Liao, A.; Gu, C.; Wong, L.; Ottolia, M.; Olcese, R.; Schwingshackl, A. BK Channels Regulate LPS-induced CCL-2 Release from Human Pulmonary Endothelial Cells. Am. J. Respir. Cell Mol. Biol. 2021, 64, 224–234. [Google Scholar] [CrossRef]
- Joachim, A.; Ye, S.; Zhao, M.T. Generation of Cardiomyocytes and Endothelial Cells from Human iPSCs by Chemical Modulation of Wnt Signaling. Methods Mol. Biol. 2022, 2549, 335–344. [Google Scholar] [PubMed]
- Lv, L.; Qi, H.; Guo, X.; Ni, Q.; Yan, Z.; Zhang, L. Long Noncoding RNA uc001pwg.1 Is Downregulated in Neointima in Arteriovenous Fistulas and Mediates the Function of Endothelial Cells Derived from Pluripotent Stem Cells. Stem Cells Int. 2017, 2017, 4252974. [Google Scholar] [CrossRef] [Green Version]
- Iring, A.; Jin, Y.J.; Albarran-Juarez, J.; Siragusa, M.; Wang, S.; Dancs, P.T.; Nakayama, A.; Tonack, S.; Chen, M.; Kunne, C.; et al. Shear stress-induced endothelial adrenomedullin signaling regulates vascular tone and blood pressure. J. Clin. Investig. 2019, 129, 2775–2791. [Google Scholar] [CrossRef] [Green Version]
- Nilius, B.; Droogmans, G. Ion channels and their functional role in vascular endothelium. Physiol. Rev. 2001, 81, 1415–1459. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, D.; Aoyama, M.; Ohya, S.; Muraki, K.; Asai, K.; Imaizumi, Y. Novel functions of small conductance Ca2+-activated K+ channel in enhanced cell proliferation by ATP in brain endothelial cells. J. Biol. Chem. 2006, 281, 38430–38439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnham, M.P.; Bychkov, R.; Feletou, M.; Richards, G.R.; Vanhoutte, P.M.; Weston, A.H.; Edwards, G. Characterization of an apamin-sensitive small-conductance Ca(2+)-activated K(+) channel in porcine coronary artery endothelium: Relevance to EDHF. Br. J. Pharmacol. 2002, 135, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Sultan, S.; Gosling, M.; Abu-Hayyeh, S.; Carey, N.; Powell, J.T. Flow-dependent increase of ICAM-1 on saphenous vein endothelium is sensitive to apamin. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H22–H28. [Google Scholar] [CrossRef] [Green Version]
- Nam, Y.W.; Kong, D.; Wang, D.; Orfali, R.; Sherpa, R.T.; Totonchy, J.; Nauli, S.M.; Zhang, M. Differential modulation of SK channel subtypes by phosphorylation. Cell Calcium. 2021, 94, 102346. [Google Scholar] [CrossRef]
- Sorensen, C.M.; Braunstein, T.H.; Holstein-Rathlou, N.H.; Salomonsson, M. Role of vascular potassium channels in the regulation of renal hemodynamics. Am. J. Physiol. Renal. Physiol. 2012, 302, F505–F518. [Google Scholar] [CrossRef]
- Ma, W.; Compan, V.; Zheng, W.; Martin, E.; North, R.A.; Verkhratsky, A.; Surprenant, A. Pannexin 1 forms an anion-selective channel. Pflugers Arch. 2012, 463, 585–592. [Google Scholar] [CrossRef]
- Taylor, M.S.; Bonev, A.D.; Gross, T.P.; Eckman, D.M.; Brayden, J.E.; Bond, C.T.; Adelman, J.P.; Nelson, M.T. Altered expression of small-conductance Ca2+-activated K+ (SK3) channels modulates arterial tone and blood pressure. Circ. Res. 2003, 93, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, A.I.; Drachuk, K.; Panasiuk, O.; Sagach, V.; Deak, A.T.; Malli, R.; Graier, W.F. N-Arachidonoyl glycine suppresses Na(+)/Ca(2)(+) exchanger-mediated Ca(2)(+) entry into endothelial cells and activates BK(Ca) channels independently of GPCRs. Br. J. Pharmacol. 2013, 169, 933–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soloviev, A.I.; Kizub, I.V. Mechanisms of vascular dysfunction evoked by ionizing radiation and possible targets for its pharmacological correction. Biochem. Pharmacol. 2019, 159, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, A.I.; Panasiuk, O.; Okhai, I.; Montecucco, F.; Brandt, K.J.; Mach, F. Ca(2+)-dependent potassium channels and cannabinoid signaling in the endothelium of apolipoprotein E knockout mice before plaque formation. J. Mol. Cell Cardiol. 2018, 115, 54–63. [Google Scholar] [CrossRef]
- Qu, L.; Yu, L.; Wang, Y.; Jin, X.; Zhang, Q.; Lu, P.; Yu, X.; Zhong, W.; Zheng, X.; Cui, N.; et al. Inward Rectifier K+ Currents Are Regulated by CaMKII in Endothelial Cells of Primarily Cultured Bovine Pulmonary Arteries. PLoS ONE 2015, 10, e0145508. [Google Scholar] [CrossRef]
- Lee, C.H.; Dai, Z.K.; Yen, C.T.; Hsieh, S.L.; Wu, B.N. KMUP-1 protects against streptozotocin-induced mesenteric artery dysfunction via activation of ATP-sensitive potassium channels. Pharmacol. Rep. 2018, 70, 746–752. [Google Scholar] [CrossRef]
- Cheng, Z.; Shen, X.; Jiang, X.; Shan, H.; Cimini, M.; Fang, P.; Ji, Y.; Park, J.Y.; Drosatos, K.; Yang, X.; et al. Hyperhomocysteinemia potentiates diabetes-impaired EDHF-induced vascular relaxation: Role of insufficient hydrogen sulfide. Redox Biol. 2018, 16, 215–225. [Google Scholar] [CrossRef]
- Peng, G.Y.; Lin, Y.; Li, J.J.; Wang, Y.; Huang, H.Y.; Shen, Z.Y. The Application of Induced Pluripotent Stem Cells in Pathogenesis Study and Gene Therapy for Vascular Disorders: Current Progress and Future Challenges. Stem Cells Int. 2019, 2019, 9613258. [Google Scholar] [CrossRef]
- Figueroa, X.F.; Duling, B.R. Gap junctions in the control of vascular function. Antioxid. Redox Signal. 2009, 11, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.; Collin de l’Hortet, A.; Soto-Gutierrez, A. Induced Pluripotent Stem Cell-Derived Endothelial Cells: Overview, Current Advances, Applications, and Future Directions. Am. J. Pathol. 2019, 189, 502–512. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.A.; Armstrong, M.L.; Piegors, D.J.; Heistad, D.D. Vascular responses to endothelin-1 in atherosclerotic primates. Arteriosclerosis 1990, 10, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Blixt, F.W.; Haanes, K.A.; Ohlsson, L.; Tolstrup Christiansen, A.; Warfvinge, K.; Edvinsson, L. Increased endothelin-1-mediated vasoconstriction after organ culture in rat and pig ocular arteries can be suppressed with MEK/ERK1/2 inhibitors. Acta Ophthalmol. 2018, 96, e619–e625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skuratovskaia, D.; Vulf, M.; Khaziakhmatova, O.; Malashchenko, V.; Komar, A.; Shunkin, E.; Gazatova, N.; Litvinova, L. Exosome Limitations in the Treatment of Inflammatory Diseases. Curr. Pharm. Des. 2021, 27, 3105–3121. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Takahashi, Y.; Takakura, Y. Possibility of Exosome-Based Therapeutics and Challenges in Production of Exosomes Eligible for Therapeutic Application. Biol. Pharm. Bull. 2018, 41, 835–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Rourke, S.T.; Vanhoutte, P.M. Adrenergic and cholinergic regulation of bronchial vascular tone. Am. Rev. Respir. Dis. 1992, 146, S11–S14. [Google Scholar] [CrossRef] [PubMed]
- Grimsey, N.J.; Lin, Y.; Narala, R.; Rada, C.C.; Mejia-Pena, H.; Trejo, J. G protein-coupled receptors activate p38 MAPK via a non-canonical TAB1-TAB2- and TAB1-TAB3-dependent pathway in endothelial cells. J. Biol. Chem. 2019, 294, 5867–5878. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, M.; Sekizuka, E.; Yamaguchi, N.; Nakadate, H.; Terao, S.; Granger, D.N.; Minamitani, H. Angiotensin II type 1 receptor signaling contributes to platelet-leukocyte-endothelial cell interactions in the cerebral microvasculature. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H2306–H2315. [Google Scholar] [CrossRef]
- Jacques, D.; Provost, C.; Normand, A.; Abou Abdallah, N.; Al-Khoury, J.; Bkaily, G. Angiotensin II induces apoptosis of human right and left ventricular endocardial endothelial cells by activating the AT2 receptor (1). Can. J. Physiol. Pharmacol. 2019, 97, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, C.; Ganju, R.K.; Pompili, V.J.; Chakroborty, D. Enhanced peripheral dopamine impairs post-ischemic healing by suppressing angiotensin receptor type 1 expression in endothelial cells and inhibiting angiogenesis. Angiogenesis 2017, 20, 97–107. [Google Scholar] [CrossRef]
- Rhoden, A.; Speiser, J.; Geertz, B.; Uebeler, J.; Schmidt, K.; de Wit, C.; Eschenhagen, T. Preserved cardiovascular homeostasis despite blunted acetylcholine-induced dilation in mice with endothelial muscarinic M3 receptor deletion. Acta Physiol. 2019, 226, e13262. [Google Scholar] [CrossRef]
- El-Battrawy, I.; Lan, H.; Cyganek, L.; Zhao, Z.; Li, X.; Buljubasic, F.; Lang, S.; Yucel, G.; Sattler, K.; Zimmermann, W.H.; et al. Modeling Short QT Syndrome Using Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes. J. Am. Heart Assoc. 2018, 7, e007394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattler, K.; El-Battrawy, I.; Cyganek, L.; Lang, S.; Lan, H.; Li, X.; Zhao, Z.; Utikal, J.; Wieland, T.; Borggrefe, M.; et al. TRPV1 activation and internalization is part of the LPS-induced inflammation in human iPSC-derived cardiomyocytes. Sci. Rep. 2021, 11, 14689. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, L.; Li, Y.; Chen, L.; Wang, X.; Guo, W.; Zhang, X.; Qin, G.; He, S.H.; Zimmerman, A.; et al. Exosomes/microvesicles from induced pluripotent stem cells deliver cardioprotective miRNAs and prevent cardiomyocyte apoptosis in the ischemic myocardium. Int. J. Cardiol. 2015, 192, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, K.; Haslbeck, M.; Buchner, J. The heat shock response: Life on the verge of death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, X.; Cyganek, L.; Nitschke, K.; Uhlig, S.; Nuhn, P.; Bieback, K.; Duerschmied, D.; El-Battrawy, I.; Zhou, X.; Akin, I. Functional Characterization of Human Induced Pluripotent Stem Cell-Derived Endothelial Cells. Int. J. Mol. Sci. 2022, 23, 8507. https://doi.org/10.3390/ijms23158507
Fan X, Cyganek L, Nitschke K, Uhlig S, Nuhn P, Bieback K, Duerschmied D, El-Battrawy I, Zhou X, Akin I. Functional Characterization of Human Induced Pluripotent Stem Cell-Derived Endothelial Cells. International Journal of Molecular Sciences. 2022; 23(15):8507. https://doi.org/10.3390/ijms23158507
Chicago/Turabian StyleFan, Xuehui, Lukas Cyganek, Katja Nitschke, Stefanie Uhlig, Philipp Nuhn, Karen Bieback, Daniel Duerschmied, Ibrahim El-Battrawy, Xiaobo Zhou, and Ibrahim Akin. 2022. "Functional Characterization of Human Induced Pluripotent Stem Cell-Derived Endothelial Cells" International Journal of Molecular Sciences 23, no. 15: 8507. https://doi.org/10.3390/ijms23158507