
A Novel Cell-Based Model for a Rare Disease: The Tks4-KO Human Embryonic Stem Cell Line as a Frank-Ter Haar Syndrome Model System

 ,
,  and
and 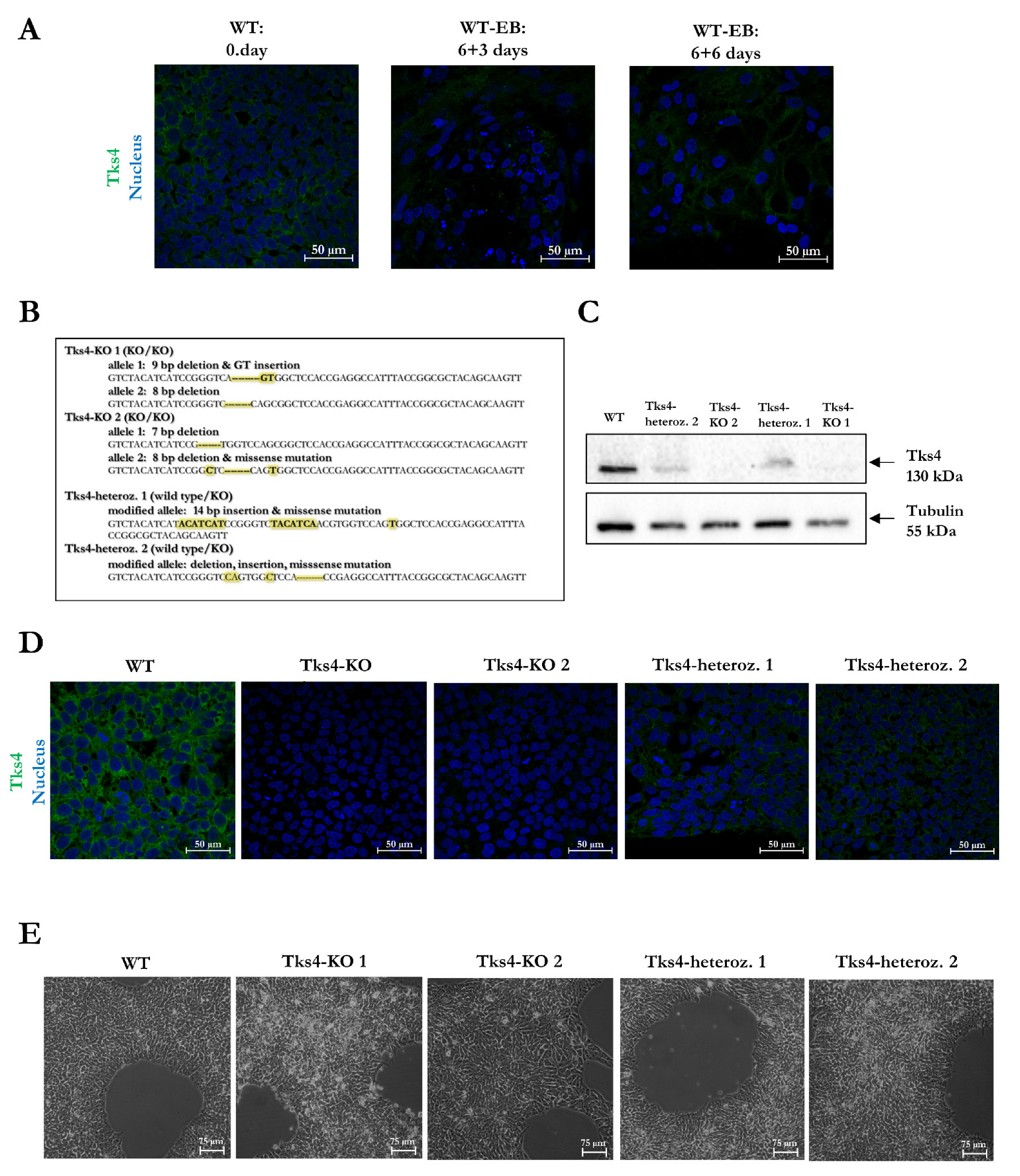{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
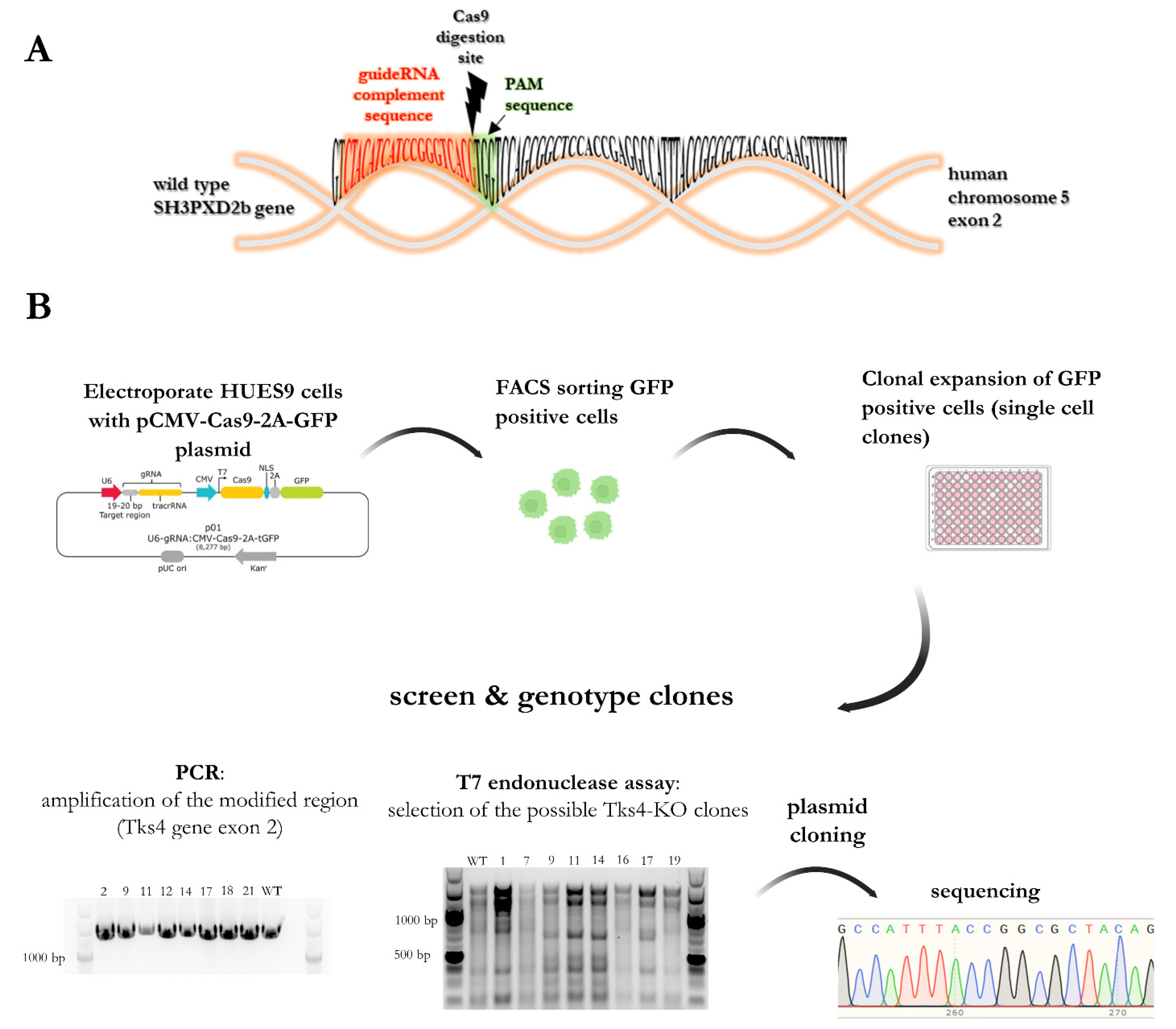
2.1. Generation of Tks4 Knockout HUES9 Cell Lines
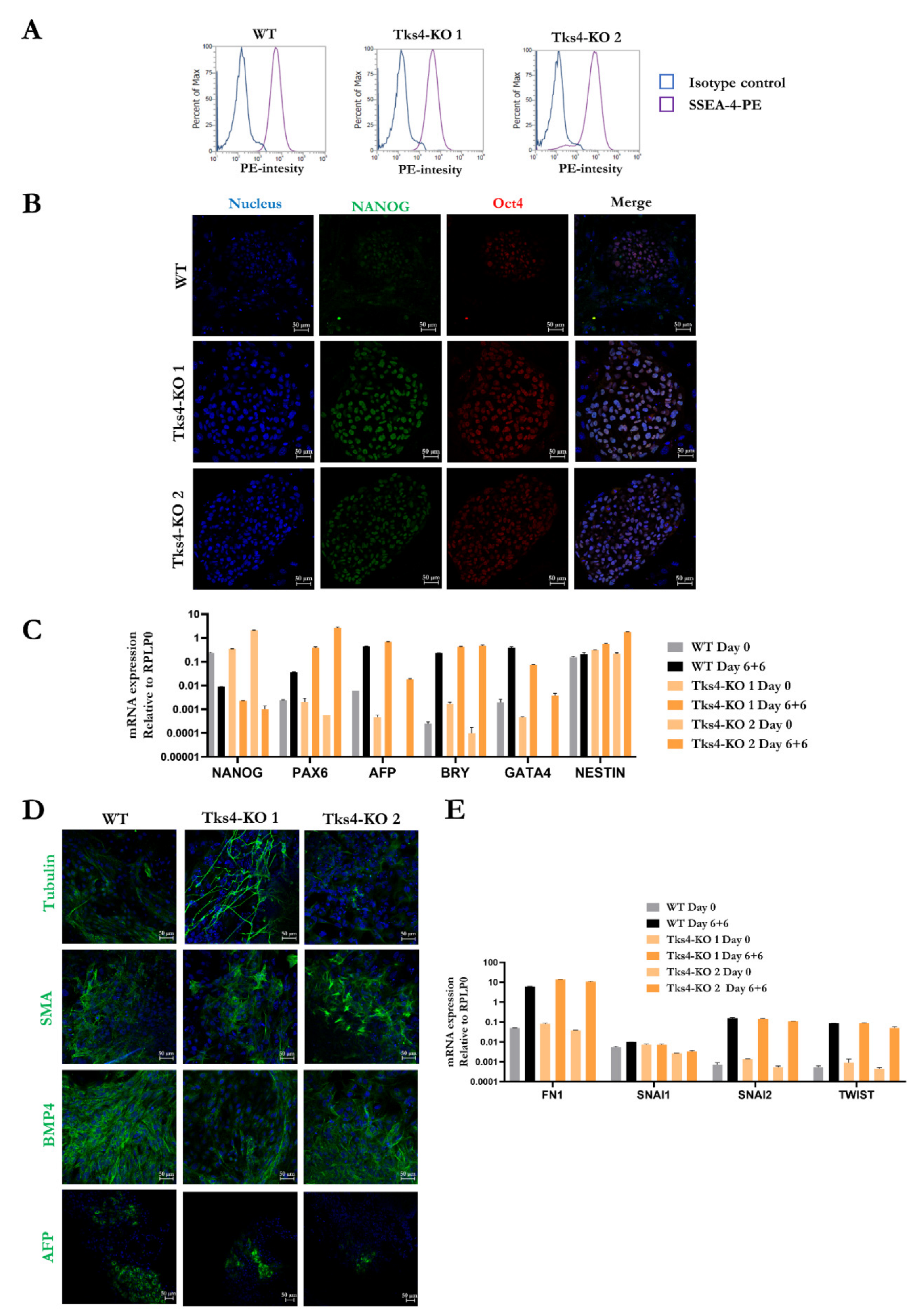
2.2. Validation of the Pluripotency of the Tks4-KO HUES9 Cells
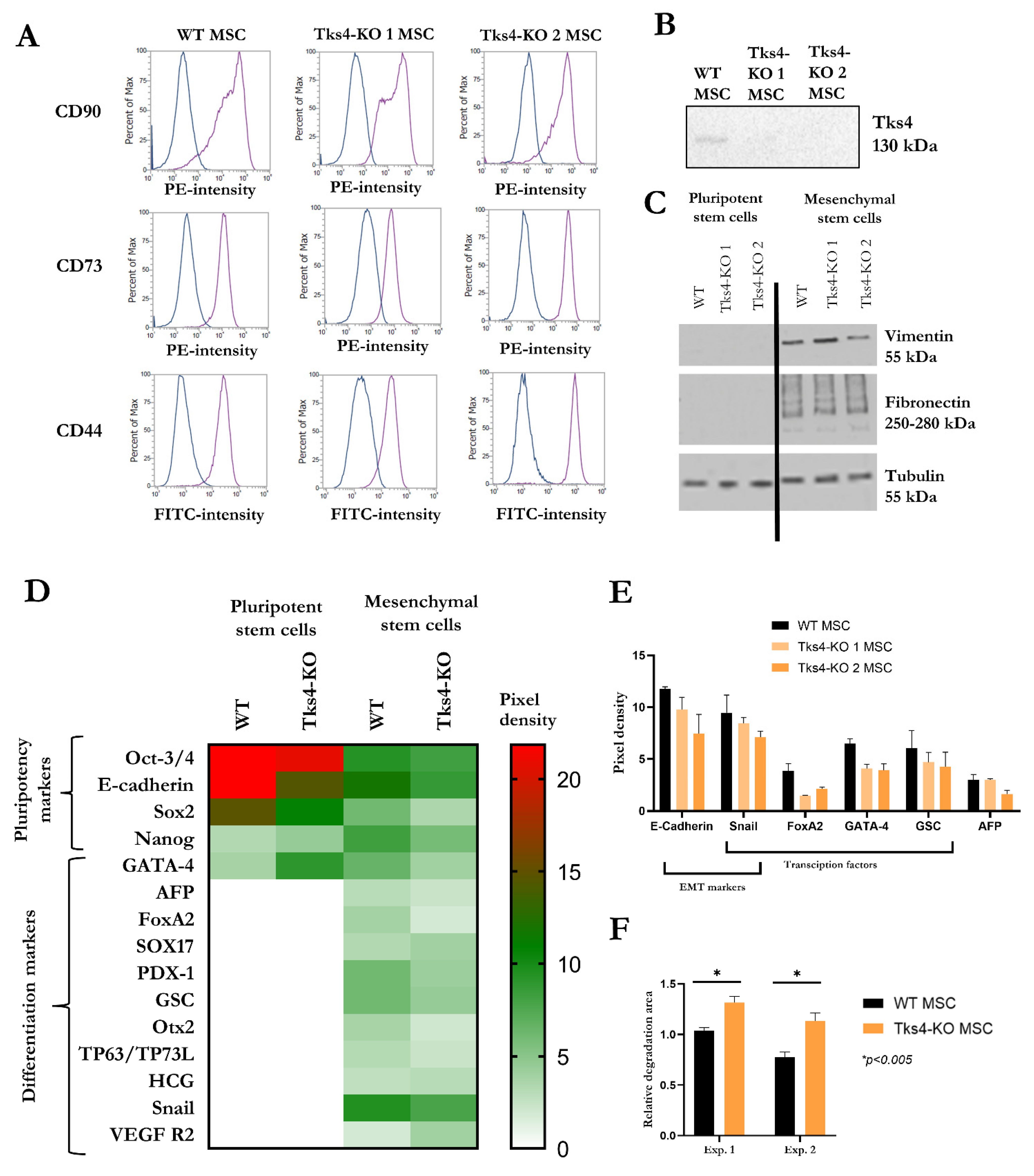
2.3. Directing the Differentiation of the Tks4-KO HUES9 Cells into Mesenchymal Stem Cells (MSCs)
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Cultures
4.2. In Vitro Spontaneous Differentiation via Embryoid Body Formation
4.3. MSC Differentiation Process
4.4. CRISPR-Cas9 Genome Editing Method to Generate Tks4-KO HUES9 Cell Line
4.5. Antibodies for Western Blotting, Immunocytochemistry, and Flow Cytometry
4.6. Western Blotting
4.7. Immunocytochemistry and Confocal Microscopy
4.8. Proteome Profiler Human Pluripotent Stem Cell Array Kit
4.9. Invadopodia Assay
4.10. RT-qPCR
4.11. Testing the Genetic Integrity of the Clones
4.12. Proliferation Rate Measurements
4.13. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dülk, M.; Szeder, B.; Glatz, G.; Merö, B.L.; Koprivanacz, K.; Kudlik, G.; Vas, V.; Sipeki, S.; Cserkaszky, A.; Radnai, L.; et al. EGF Regulates the Interaction of Tks4 with Src through Its SH2 and SH3 Domains. Biochemistry 2018, 57, 4186–4196. [Google Scholar] [CrossRef] [PubMed]
- Bögel, G.; Gujdár, A.; Geiszt, M.; Lányi, Á.; Fekete, A.; Sipeki, S.; Downward, J.; Buday, L. Frank-ter Haar Syndrome Protein Tks4 Regulates Epidermal Growth Factor-dependent Cell Migration. J. Biol. Chem. 2012, 287, 31321–31329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buday, L.; Tompa, P. Functional classification of scaffold proteins and related molecules. FEBS J. 2010, 277, 4348–4355. [Google Scholar] [CrossRef] [PubMed]
- Vas, V.; Háhner, T.; Kudlik, G.; Ernszt, D.; Kvell, K.; Kuti, D.; Kovács, K.J.; Tóvári, J.; Trexler, M.; Merő, B.L.; et al. Analysis of Tks4 Knockout Mice Suggests a Role for Tks4 in Adipose Tissue Homeostasis in the Context of Beigeing. Cells 2019, 8, 831. [Google Scholar] [CrossRef] [Green Version]
- Vas, V.; Kovács, T.; Körmendi, S.; Bródy, A.; Kudlik, G.; Szeder, B.; Mező, D.; Kállai, D.; Koprivanacz, K.; Merő, B.L.; et al. Significance of the Tks4 scaffold protein in bone tissue homeostasis. Sci. Rep. 2019, 9, 5781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudlik, G.; Takács, T.; Radnai, L.; Kurilla, A.; Szeder, B.; Koprivanacz, K.; Merő, B.L.; Buday, L.; Vas, V. Advances in Understanding TKS4 and TKS5: Molecular Scaffolds Regulating Cellular Processes from Podosome and Invadopodium Formation to Differentiation and Tissue Homeostasis. Int. J. Mol. Sci. 2020, 21, 8117. [Google Scholar] [CrossRef]
- Massadeh, S.; Alhabshan, F.; Alsudairi, H.N.; Alkwai, S.; Alswailm, M.; Kabbani, M.S.; Chaikhouni, F.; Alaamery, M. The Role of the Disrupted Podosome Adaptor Protein (SH3PXD2B) in Frank–Ter Haar Syndrome. Genes 2022, 13, 236. [Google Scholar] [CrossRef]
- Türkyılmaz, A.; Sager, S.G.; Topcu, B.; Kaplan, A.T.; Günbey, H.P.; Akın, Y. Novel SH3PXD2B variant identified by whole-exome sequencing in a Turkish newborn with frank-ter haar Syndrome. Clin. Dysmorphol. 2022, 31, 45–49. [Google Scholar] [CrossRef]
- Durand, B.; Stoetzel, C.; Schaefer, E.; Calmels, N.; Scheidecker, S.; Kempf, N.; De Melo, C.; Guilbert, A.S.; Timbolschi, D.; Donato, L.; et al. A severe case of Frank-ter Haar syndrome and literature review: Further delineation of the phenotypical spectrum. Eur. J. Med. Genet. 2020, 63, 103857. [Google Scholar] [CrossRef]
- Chang, T.C.; Bauer, M.; Puerta, H.S.; Greenberg, M.B.; Cavuoto, K.M. Ophthalmic findings in Frank-ter Haar syndrome: Report of a sibling pair. J. AAPOS 2017, 21, 514–516. [Google Scholar] [CrossRef]
- Parker, K.; Pabla, R.; Hay, N.; Ayliffe, P. Common dental features and craniofacial development of three siblings with Ter Haar syndrome. Eur. Arch. Paediatr. Dent. 2014, 15, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Cejudo-Martin, P.; de Brouwer, A.; van der Zwaag, B.; Ruiz-Lozano, P.; Scimia, M.C.; Lindsey, J.D.; Weinreb, R.; Albrecht, B.; Megarbane, A.; et al. Disruption of the Podosome Adaptor Protein TKS4 (SH3PXD2B) Causes the Skeletal Dysplasia, Eye, and Cardiac Abnormalities of Frank-Ter Haar Syndrome. Am. J. Hum. Genet. 2010, 86, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buschman, M.D.; Bromann, P.A.; Cejudo-Martin, P.; Wen, F.; Pass, I.; Courtneidge, S.A. The novel adaptor protein Tks4 (SH3PXD2B) is required for functional podosome formation. Mol. Biol. Cell 2009, 20, 1302–1311. [Google Scholar] [CrossRef] [Green Version]
- Papatheodorou, I.; Moreno, P.; Manning, J.; Fuentes, A.M.P.; George, N.; Fexova, S.; Fonseca, N.A.; Füllgrabe, A.; Green, M.; Huang, N.; et al. Expression Atlas update: From tissues to single cells. Nucleic Acids Res. 2020, 48, D77–D83. [Google Scholar] [CrossRef] [Green Version]
- Van De Leemput, J.; Boles, N.C.; Kiehl, T.R.; Corneo, B.; Lederman, P.; Menon, V.; Lee, C.; Martinez, R.A.; Levi, B.P.; Thompson, C.L.; et al. NeuroResource CORTECON: A Temporal Transcriptome Analysis of In Vitro Human Cerebral Cortex Development from Human Embryonic Stem Cells. Neuron 2014, 83, 51–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraiczy, J.; Nayak, K.M.; Howell, K.J.; Ross, A.; Forbester, J.; Salvestrini, C.; Mustata, R.; Perkins, S.; Andersson-Rolf, A.; Leenen, E.; et al. DNA methylation defines regional identity of human intestinal epithelial organoids and undergoes dynamic changes during development. Gut 2019, 68, 49–61. [Google Scholar] [CrossRef]
- Kondrashov, A.; Mohd Yusof, N.A.N.; Hasan, A.; Goulding, J.; Kodagoda, T.; Hoang, D.M.; Vo, N.T.N.; Melarangi, T.; Dolatshad, N.; Gorelik, J.; et al. CRISPR/Cas9-mediated generation and analysis of N terminus polymorphic models of β2AR in isogenic hPSC-derived cardiomyocytes. Mol. Ther. Methods Clin. Dev. 2021, 20, 39–53. [Google Scholar] [CrossRef]
- Klementieva, N.; Goliusova, D.; Krupinova, J.; Yanvarev, V.; Panova, A.; Mokrysheva, N.; Kiselev, S.L. A Novel Isogenic Human Cell-Based System for MEN1 Syndrome Generated by CRISPR/Cas9 Genome Editing. Int. J. Mol. Sci. 2021, 22, 12054. [Google Scholar] [CrossRef]
- Szeder, B.; Tárnoki-Zách, J.; Lakatos, D.; Vas, V.; Kudlik, G.; Merő, B.; Koprivanacz, K.; Bányai, L.; Hámori, L.; Róna, G.; et al. Absence of the Tks4 Scaffold Protein Induces Epithelial-Mesenchymal Transition-Like Changes in Human Colon Cancer Cells. Cells 2019, 8, 1343. [Google Scholar] [CrossRef] [Green Version]
- Varga, N.; Veréb, Z.; Rajnavölgyi, É.; Német, K.; Uher, F.; Sarkadi, B.; Apáti, Á. Mesenchymal stem cell like (MSCl) cells generated from human embryonic stem cells support pluripotent cell growth. Biochem. Biophys. Res. Commun. 2011, 414, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Pérez, J.A.; Wakamiya, M.; Behringer, R.R. Goosecoid acts cell autonomously in mesenchyme-derived tissues during craniofacial development. Development 1999, 126, 3811–3821. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.B.; Pence, J.; Suthon, S.; Lin, J.; Miranda-Carboni, G.A.; Krum, S.A. GATA4 regulates mesenchymal stem cells via direct transcriptional regulation of the WNT signalosome. Bone 2021, 144, 115819. [Google Scholar] [CrossRef] [PubMed]
- Eastham, A.M.; Spencer, H.; Soncin, F.; Ritson, S.; Merry, C.L.R.; Stern, P.L.; Ward, C.M. Epithelial-Mesenchymal Transition Events during Human Embryonic Stem Cell Differentiation. Cancer Res. 2007, 67, 11254–11262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zrhidri, A.; Jaouad, I.C.; Lyahyai, J.; Raymond, L.; Egéa, G.; Taoudi, M.; El Mouatassim, S.; Sefiani, A. Identification of two novel SH3PXD2B gene mutations in Frank-Ter Haar syndrome by exome sequencing: Case report and review of the literature. Gene 2017, 628, 190–193. [Google Scholar] [CrossRef]
- Ratukondla, B.; Prakash, S.; Reddy, S.; Puthuran, G.V.; Kannan, N.B.; Pillai, M.R. A Rare Case Report of Frank ter Haar Syndrome in a Sibling Pair Presenting with Congenital Glaucoma. J. Glaucoma 2020, 29, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Thedens, D.R.; Bo, A.E.; Ae, C.; Ae, B.S.H.; Yin, Q.; Ae, Z.; Johnson, K.R.; Leah, A.E.; Donahue, R.; et al. The podosomal-adaptor protein SH3PXD2B is essential for normal postnatal development. Mamm. Genome 2009, 20, 462–475. [Google Scholar] [CrossRef] [Green Version]
- Dülk, M.; Kudlik, G.; Fekete, A.; Ernszt, D.; Kvell, K.; Pongrácz, J.E.; Merő, B.L.; Szeder, B.; Radnai, L.; Geiszt, M.; et al. The scaffold protein Tks4 is required for the differentiation of mesenchymal stromal cells (MSCs) into adipogenic and osteogenic lineages. Sci. Rep. 2016, 6, 34280. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Cui, K.; Yi, Q.; Yu, L.; Xu, Y. DNA repair mechanisms in embryonic stem cells. Cell. Mol. Life Sci. 2016, 74, 487–493. [Google Scholar] [CrossRef]
- Yao, Y.; Dai, W. Genomic Instability and Cancer. J. Carcinog. Mutagen. 2014, 5, 1000165. [Google Scholar] [CrossRef]
- Gabant, P.; Forrester, L.; Nichols, J.; Van Reeth, T.; De Mees, C.; Pajack, B.; Watt, A.; Smitz, J.; Alexandre, H.; Szpirer, C.; et al. Alpha-fetoprotein, the major fetal serum protein, is not essential for embryonic development but is required for female fertility. Proc. Natl. Acad. Sci. USA 2002, 99, 12865–12870. [Google Scholar] [CrossRef] [Green Version]
- Reé, D.; Borsy, A.; Fóthi, Á.; Orbán, T.I.; Várady, G.; Erdei, Z.; Sarkadi, B.; Réthelyi, J.; Varga, N.; Apáti, Á. Establishing a human embryonic stem cell clone with a heterozygous mutation in the DGCR8 gene. Stem Cell Res. 2021, 50, 102134. [Google Scholar] [CrossRef]
- Erdei, Z.; Lorincz, R.; Szebényi, K.; Péntek, A.; Varga, N.; Likó, I.; Várady, G.; Szakács, G.; Orbán, T.I.; Sarkadi, B.; et al. Expression pattern of the human ABC transporters in pluripotent embryonic stem cells and in their derivatives. Cytom. Part B Clin. Cytom. 2014, 86, 299–310. [Google Scholar] [CrossRef]
- Lányi, Á.; Baráth, M.; Péterfi, Z.; Bögel, G.; Orient, A.; Simon, T.; Petrovszki, E.; Kis-Tóth, K.; Sirokmány, G.; Rajnavölgyi, É.; et al. The Homolog of the Five SH3-Domain Protein (HOFI/SH3PXD2B) Regulates Lamellipodia Formation and Cell Spreading. PLoS ONE 2011, 6, e23653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambacurta, A.; Merlini, G.; Ruggiero, C.; Diedenhofen, G.; Battista, N.; Bari, M.; Balsamo, M.; Piccirillo, S.; Valentini, G.; Mascetti, G.; et al. Human osteogenic differentiation in Space: Proteomic and epigenetic clues to better understand osteoporosis. Sci. Rep. 2019, 9, 8343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobis-Wozowicz, S.; Kmiotek, K.; Sekula, M.; Kedracka-Krok, S.; Kamycka, E.; Adamiak, M.; Jankowska, U.; Madetko-Talowska, A.; Sarna, M.; Bik-Multanowski, M.; et al. Human Induced Pluripotent Stem Cell-Derived Microvesicles Transmit RNAs and Proteins to Recipient Mature Heart Cells Modulating Cell Fate and Behavior. Stem Cells 2015, 33, 2748–2761. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
László, L.; Maczelka, H.; Takács, T.; Kurilla, A.; Tilajka, Á.; Buday, L.; Vas, V.; Apáti, Á. A Novel Cell-Based Model for a Rare Disease: The Tks4-KO Human Embryonic Stem Cell Line as a Frank-Ter Haar Syndrome Model System. Int. J. Mol. Sci. 2022, 23, 8803. https://doi.org/10.3390/ijms23158803
László L, Maczelka H, Takács T, Kurilla A, Tilajka Á, Buday L, Vas V, Apáti Á. A Novel Cell-Based Model for a Rare Disease: The Tks4-KO Human Embryonic Stem Cell Line as a Frank-Ter Haar Syndrome Model System. International Journal of Molecular Sciences. 2022; 23(15):8803. https://doi.org/10.3390/ijms23158803
Chicago/Turabian StyleLászló, Loretta, Hédi Maczelka, Tamás Takács, Anita Kurilla, Álmos Tilajka, László Buday, Virag Vas, and Ágota Apáti. 2022. "A Novel Cell-Based Model for a Rare Disease: The Tks4-KO Human Embryonic Stem Cell Line as a Frank-Ter Haar Syndrome Model System" International Journal of Molecular Sciences 23, no. 15: 8803. https://doi.org/10.3390/ijms23158803