
In Peripheral Blood Mononuclear Cells Helicobacter pylori Induces the Secretion of Soluble and Exosomal Cytokines Related to Carcinogenesis

 , , ,
, , , 
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
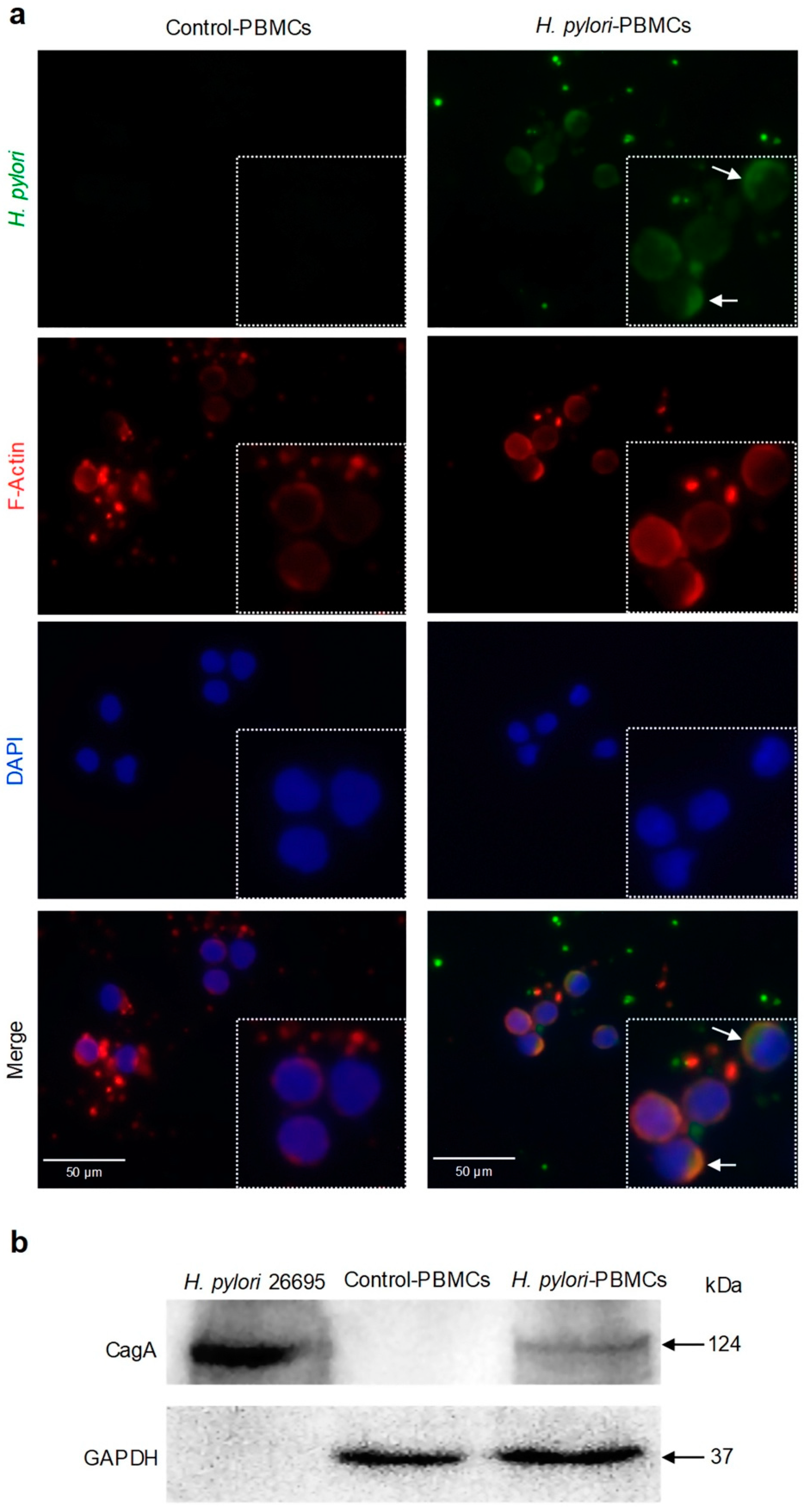
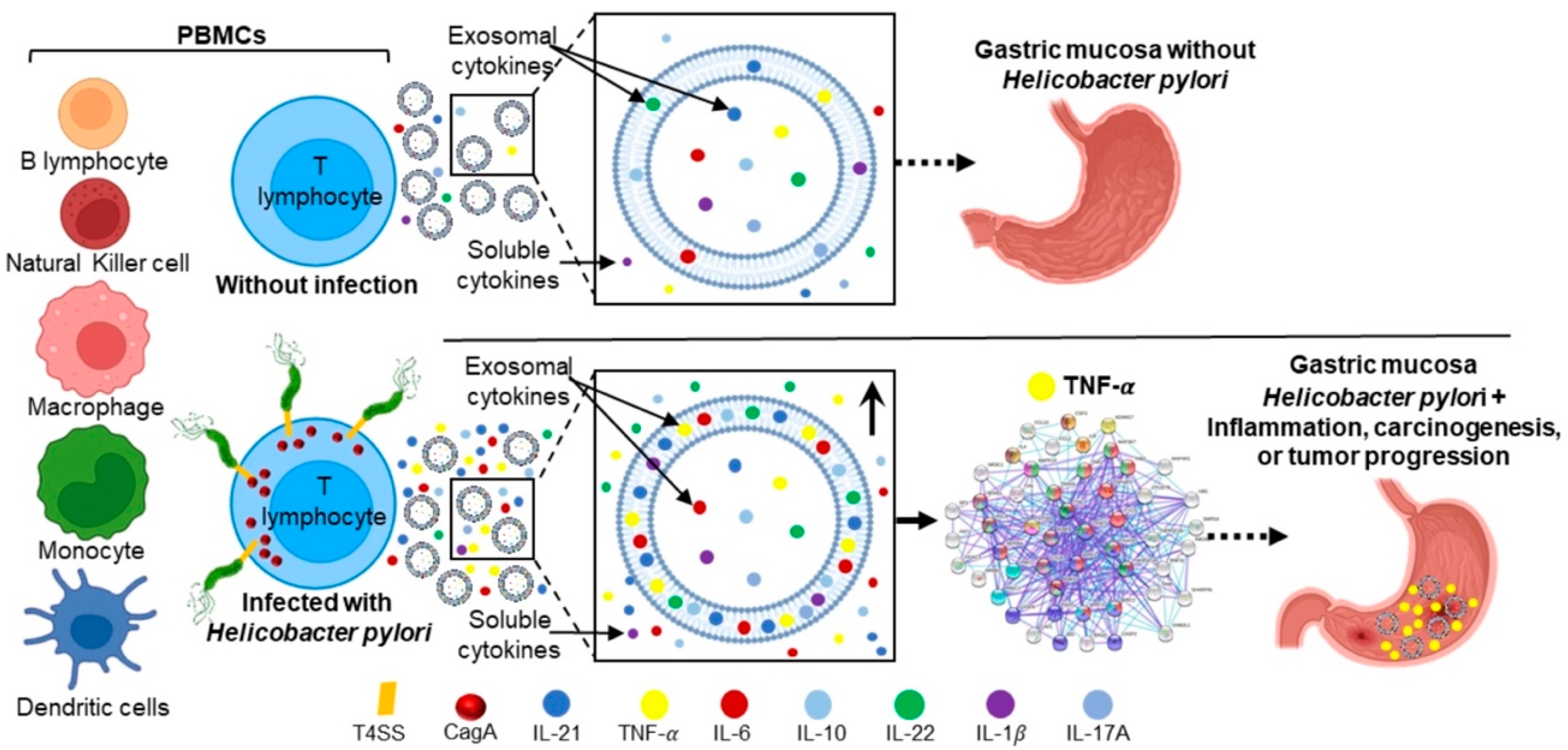
2.1. H. pylori Adheres to PBMCs and Translocates CagA
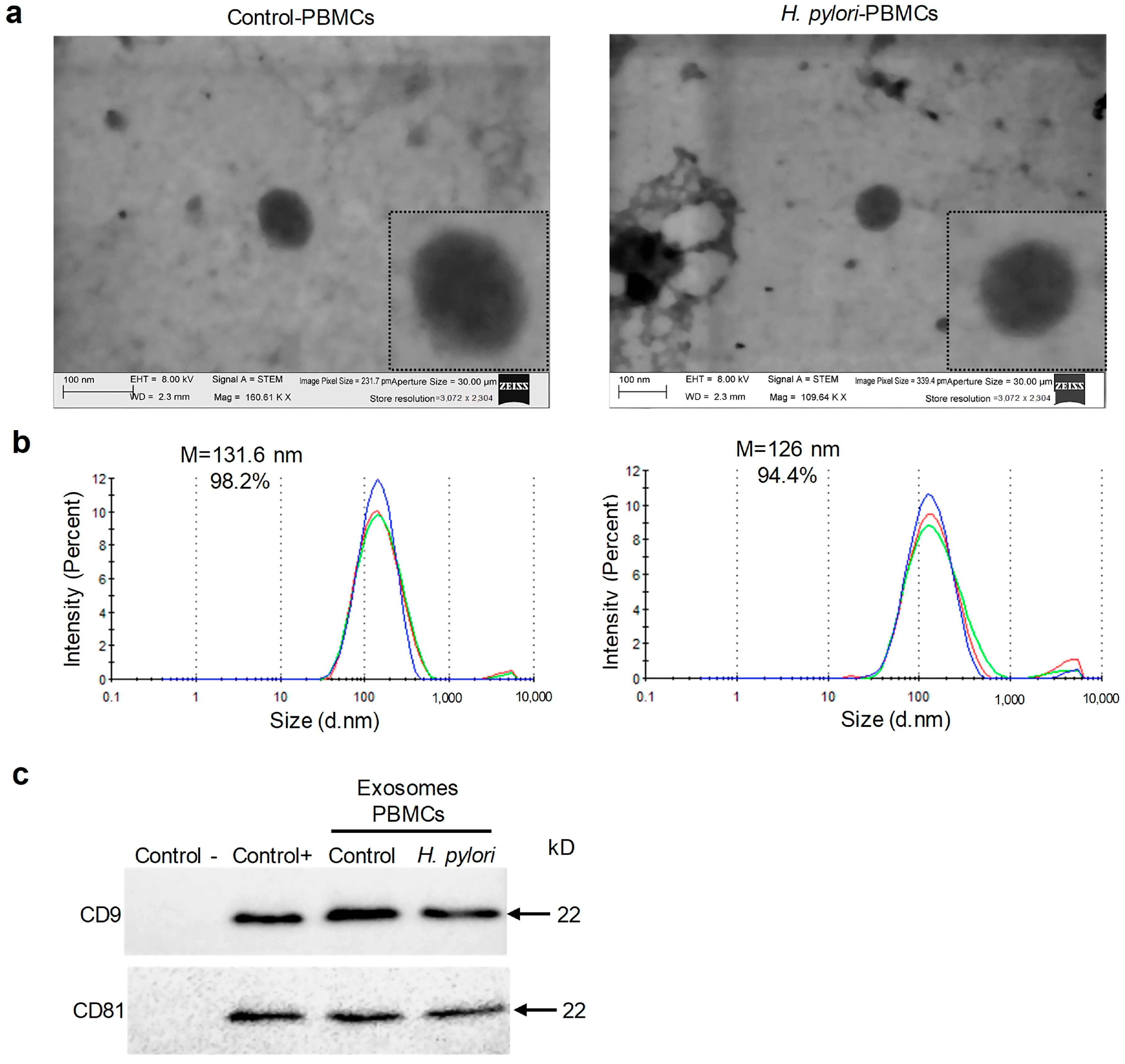
2.2. H. pylori Infected and Uninfected PBMCs Secrete Exosomes
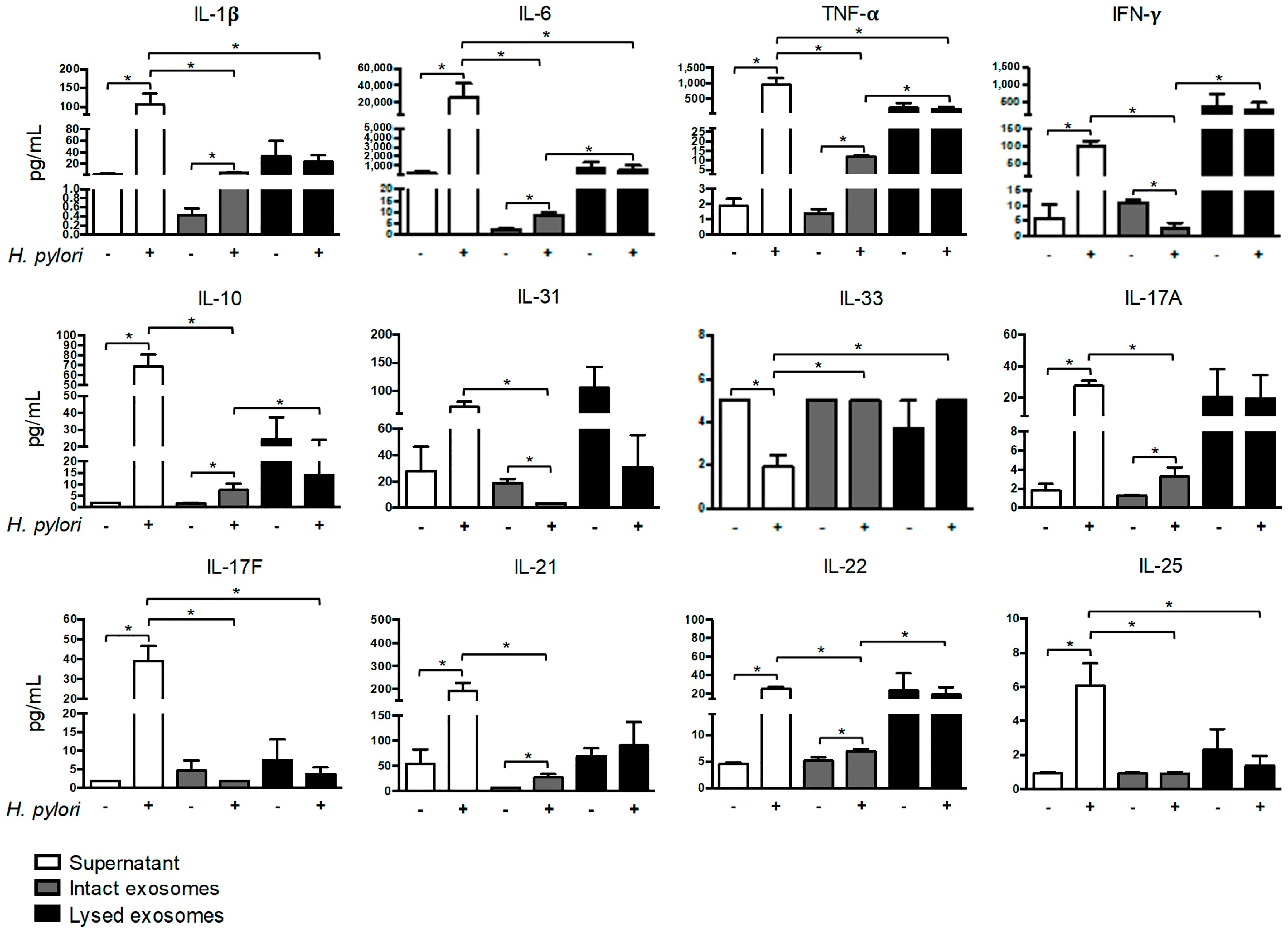
2.3. H. pylori Induces Soluble and Exosomal Cytokine Secretion by PBMCs
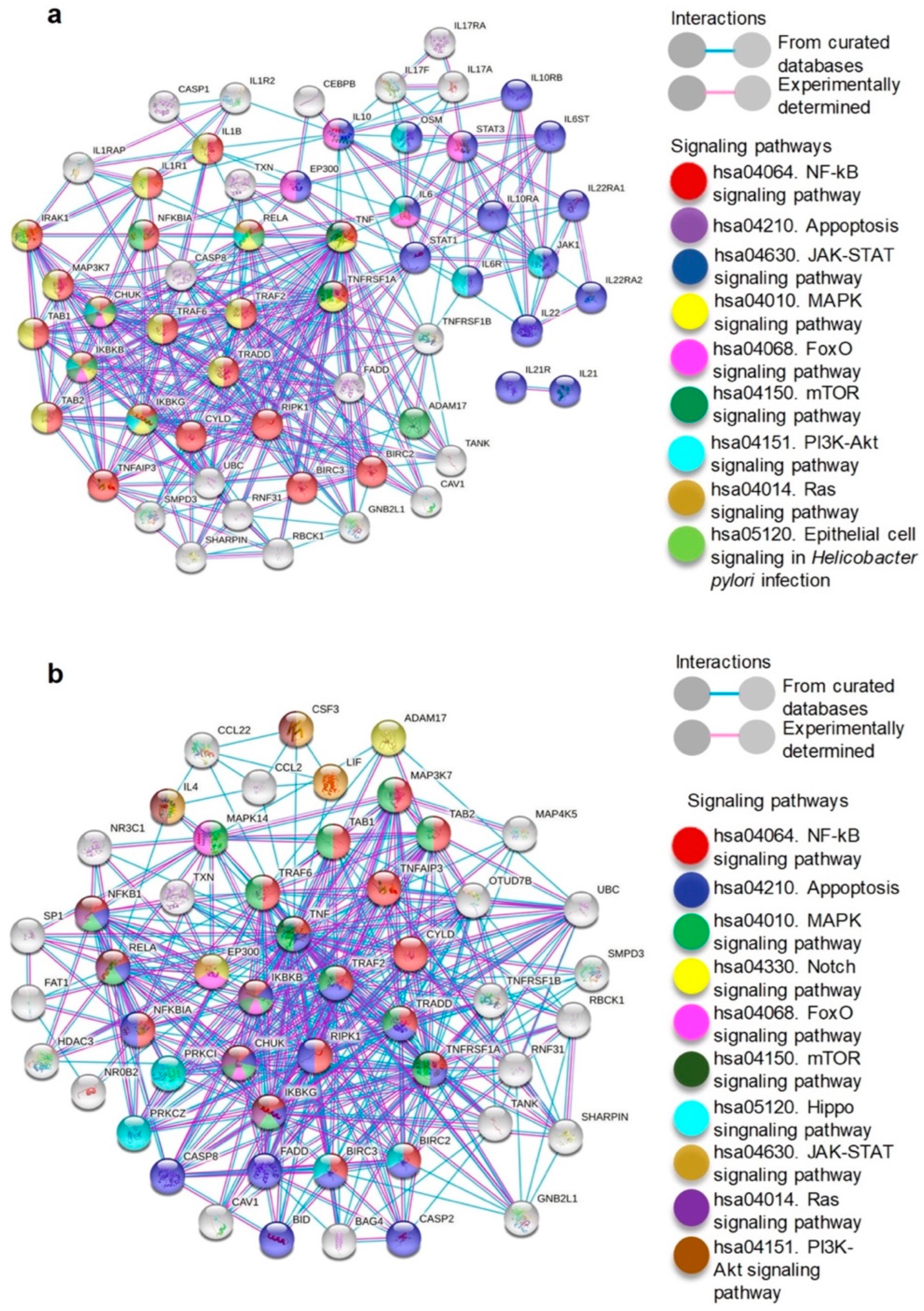
2.4. Cytokines Increased in Supernatant and Exosomes Secreted by H. pylori-PBMCs Are Associated with Signaling Pathways Involved in Carcinogenesis
3. Discussion
4. Materials and Methods
4.1. Isolation and Culture of PBMCs
4.2. H. pylori Culture and Infection of PBMCs
4.3. Immunofluorescence
4.4. Exosome Isolation and Identification
4.5. Western Blot
4.6. Multiplex Immunoassay
4.7. Protein-Protein Interaction Network Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zamani, M.; Ebrahimtabar, F.; Zamani, V.; Miller, W.H.; Alizadeh-Navaei, R.; Shokri-Shirvani, J.; Derakhshan, M.H. Systematic Review with Meta-Analysis: The Worldwide Prevalence of Helicobacter pylori Infection. Aliment. Pharmacol. Ther. 2018, 47, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, E.; Chua, W.; Ng, W.; Roberts, T.L. Peripheral Cytokine Levels as a Prognostic Indicator in Gastric Cancer: A Review of Existing Literature. Biomedicines 2021, 9, 1916. [Google Scholar] [CrossRef] [PubMed]
- Baj, J.; Forma, A.; Sitarz, M.; Portincasa, P.; Garruti, G.; Krasowska, D.; Maciejewski, R. Helicobacter pylori Virulence Factors—Mechanisms of Bacterial Pathogenicity in the Gastric Microenvironment. Cells 2020, 10, 27. [Google Scholar] [CrossRef]
- Bockerstett, K.A.; DiPaolo, R.J. Regulation of Gastric Carcinogenesis by Inflammatory Cytokines. Cell. Mol. Gastroenterol. Hepatol. 2017, 4, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Zhang, H.; Cao, A.; Cao, L.; Hu, X. Cytokine TNF-α Promotes Invasion and Metastasis of Gastric Cancer by down-Regulating Pentraxin3. J. Cancer 2020, 11, 1800–1807. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Lan, F.; Wang, X.; Yu, Y.; Ouyang, X.; Zheng, F.; Han, J.; Lin, Y.; Xie, Y.; Xie, F.; et al. IL-1β-Induced Activation of P38 Promotes Metastasis in Gastric Adenocarcinoma via Upregulation of AP-1/c-Fos, MMP2 and MMP9. Mol. Cancer 2014, 13, 18. [Google Scholar] [CrossRef]
- Xu, X.; Yang, C.; Chen, J.; Liu, J.; Li, P.; Shi, Y.; Yu, P. Interleukin-23 Promotes the Migration and Invasion of Gastric Cancer Cells by Inducing Epithelial-to-Mesenchymal Transition via the STAT3 Pathway. Biochem. Biophys. Res. Commun. 2018, 499, 273–278. [Google Scholar] [CrossRef]
- Yu, X.-X.; Hu, Z.; Shen, X.; Dong, L.-Y.; Zhou, W.-Z.; Hu, W.-H. IL-33 Promotes Gastric Cancer Cell Invasion and Migration via ST2–ERK1/2 Pathway. Dig. Dis. Sci. 2015, 60, 1265–1272. [Google Scholar] [CrossRef]
- Samanta, S.; Rajasingh, S.; Drosos, N.; Zhou, Z.; Dawn, B.; Rajasingh, J. Exosomes: New Molecular Targets of Diseases. Acta Pharmacol. Sin. 2018, 39, 501–513. [Google Scholar] [CrossRef]
- Jadli, A.S.; Ballasy, N.; Edalat, P.; Patel, V.B. Inside (Sight) of Tiny Communicator: Exosome Biogenesis, Secretion, and Uptake. Mol. Cell Biochem. 2020, 467, 77–94. [Google Scholar] [CrossRef]
- Barnes, B.J.; Somerville, C.C. Modulating Cytokine Production via Select Packaging and Secretion from Extracellular Vesicles. Front. Immunol. 2020, 11, 1040. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Zhu, W.; Li, H.; Ma, D.; Liu, W.; Yu, W.; Wang, L.; Cao, Y.; Jiang, Y. Association between Cytokines and Exosomes in Synovial Fluid of Individuals with Knee Osteoarthritis. Mod. Rheumatol. 2020, 30, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.; Kodidela, S.; Sinha, N.; Kumar, P.; Cory, T.J.; Kumar, S. Differential Packaging of Inflammatory Cytokines/ Chemokines and Oxidative Stress Modulators in U937 and U1 Macrophages-Derived Extracellular Vesicles upon Exposure to Tobacco Constituents. PLoS ONE 2020, 15, e0233054. [Google Scholar] [CrossRef]
- Kodidela, S.; Ranjit, S.; Sinha, N.; McArthur, C.; Kumar, A.; Kumar, S. Cytokine Profiling of Exosomes Derived from the Plasma of HIV-Infected Alcohol Drinkers and Cigarette Smokers. PLoS ONE 2018, 13, e0201144. [Google Scholar] [CrossRef] [Green Version]
- Tokarz, A.; Szuścik, I.; Kuśnierz-Cabala, B.; Kapusta, M.; Konkolewska, M.; Żurakowski, A.; Georgescu, A.; Stępień, E. Extracellular Vesicles Participate in the Transport of Cytokines and Angiogenic Factors in Diabetic Patients with Ocular Complications. Folia Med. Cracov. 2015, 55, 35–48. [Google Scholar]
- Im, K.; Baek, J.; Kwon, W.S.; Rha, S.Y.; Hwang, K.W.; Kim, U.; Min, H. The Comparison of Exosome and Exosomal Cytokines between Young and Old Individuals with or without Gastric Cancer. Int. J. Gerontol. 2018, 12, 233–238. [Google Scholar] [CrossRef]
- Che, Y.; Geng, B.; Xu, Y.; Miao, X.; Chen, L.; Mu, X.; Pan, J.; Zhang, C.; Zhao, T.; Wang, C.; et al. Helicobacter pylori-Induced Exosomal MET Educates Tumour-Associated Macrophages to Promote Gastric Cancer Progression. J. Cell. Mol. Med. 2018, 22, 5708–5719. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Zhang, L.; Chi, J.; Li, H.; Liu, X.; Hu, T.; Li, R.; Guo, Y.; Zhang, X.; Wang, H.; et al. Helicobacter pylori Infection Impairs Endothelial Function through an Exosome-Mediated Mechanism. JAHA 2020, 9, e014120. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Xia, Y.; Luo, X.; Chen, S.; Li, B.; Ye, Z.; Chen, S.; Mao, L.; Jin, H.; Li, Y.; et al. Exosomal CagA Derived from Helicobacter pylori-Infected Gastric Epithelial Cells Induces Macrophage Foam Cell Formation and Promotes Atherosclerosis. J. Mol. Cell. Cardiol. 2019, 135, 40–51. [Google Scholar] [CrossRef]
- Wang, J.; Deng, Z.; Wang, Z.; Wu, J.; Gu, T.; Jiang, Y.; Li, G. MicroRNA-155 in Exosomes Secreted from Helicobacter pylori Infection Macrophages Immunomodulates Inflammatory Response. Am. J. Transl. Res. 2016, 8, 3700–3709. [Google Scholar]
- Li, N.; Liu, S.; Dong, K.; Zhang, G.; Huang, J.; Wang, Z.; Wang, T. Exosome-Transmitted MiR-25 Induced by H. pylori Promotes Vascular Endothelial Cell Injury by Targeting KLF2. Front. Cell. Infect. Microbiol. 2019, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Díaz, P.; Valenzuela Valderrama, M.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and Gastric Cancer: Adaptive Cellular Mechanisms Involved in Disease Progression. Front. Microbiol. 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, P.; Kemppainen, E.; Orešič, M. Perspectives on Systems Modeling of Human Peripheral Blood Mononuclear Cells. Front. Mol. Biosci. 2018, 4, 96. [Google Scholar] [CrossRef] [Green Version]
- Doohan, D.; Rezkitha, Y.A.A.; Waskito, L.A.; Yamaoka, Y.; Miftahussurur, M. Helicobacter pylori BabA–SabA Key Roles in the Adherence Phase: The Synergic Mechanism for Successful Colonization and Disease Development. Toxins 2021, 13, 485. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, V.; Schimpl, M.; Overman, R.C.; Stolnik, S.; Chen, Y.; Nguyen, L.; Winkler, G.S.; Gellert, P.; Klassen, J.S.; Falcone, F.H. Structural and Binding Characterization of the LacdiNAc-Specific Adhesin (LabA; HopD) Exodomain from Helicobacter pylori. Curr. Res. Struct. Biol. 2021, 3, 19–29. [Google Scholar] [CrossRef]
- Senkovich, O.A.; Yin, J.; Ekshyyan, V.; Conant, C.; Traylor, J.; Adegboyega, P.; McGee, D.J.; Rhoads, R.E.; Slepenkov, S.; Testerman, T.L. Helicobacter pylori AlpA and AlpB Bind Host Laminin and Influence Gastric Inflammation in Gerbils. Infect. Immun. 2011, 79, 3106–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zargari, S.; Bahari, A.; Goodarzi, M.T.; Mahmoodi, M.; Valadan, R. TLR2 and TLR4 Signaling Pathways and Gastric Cancer: Insights from Transcriptomics and Sample Validation. Iran Biomed. J. 2022, 26, 36–43. [Google Scholar] [PubMed]
- Lin, W.-C.; Tsai, H.-F.; Kuo, S.-H.; Wu, M.-S.; Lin, C.-W.; Hsu, P.-I.; Cheng, A.-L.; Hsu, P.-N. Translocation of Helicobacter pylori CagA into Human B Lymphocytes, the Origin of Mucosa-Associated Lymphoid Tissue Lymphoma. Cancer Res. 2010, 70, 5740–5748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odenbreit, S.; Gebert, B.; Puls, J.; Fischer, W.; Haas, R. Interaction of Helicobacter pylori with Professional Phagocytes: Role of the Cag Pathogenicity Island and Translocation, Phosphorylation and Processing of CagA. Cell Microbiol. 2001, 3, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Yoshida, M.; Nishiumi, S.; Ohnishi, N.; Kobayashi, K.; Yamamoto, K.; Fujita, T.; Hatakeyama, M.; Azuma, T. The CagA Protein of Helicobacter pylori Suppresses the Functions of Dendritic Cell in Mice. Arch. Biochem. Biophys. 2010, 498, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas-Moruno, C.; Fraioli, R.; Rechenmacher, F.; Neubauer, S.; Kapp, T.G.; Kessler, H. αvβ3- or α5β1-Integrin-Selective Peptidomimetics for Surface Coating. Angew. Chem. Int. Ed. 2016, 55, 7048–7067. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Soto, L.F.; Kutter, S.; Sewald, X.; Ertl, C.; Weiss, E.; Kapp, U.; Rohde, M.; Pirch, T.; Jung, K.; Retta, S.F.; et al. Helicobacter pylori Type IV Secretion Apparatus Exploits Β1 Integrin in a Novel RGD-Independent Manner. PLoS Pathog. 2009, 5, e1000684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, M.; Thiel, M.; Hogg, N. Leukocyte Integrins. Curr. Opin. Cell Biol. 1995, 7, 690–696. [Google Scholar] [CrossRef]
- Tegtmeyer, N.; Lind, J.; Schmid, B.; Backert, S. Helicobacter pylori CagL Y58/E59 Mutation Turns-Off Type IV Secretion-Dependent Delivery of CagA into Host Cells. PLoS ONE 2014, 9, e97782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavares, R.; Pathak, S.K. Induction of TNF, CXCL8 and IL-1β in Macrophages by Helicobacter pylori Secreted Protein HP1173 Occurs via MAP-Kinases, NF-κB and AP-1 Signaling Pathways. Microb. Pathog. 2018, 125, 295–305. [Google Scholar] [CrossRef]
- Morningstar-Wright, L.; Czinn, S.J.; Piazuelo, M.B.; Banerjee, A.; Godlewska, R.; Blanchard, T.G. The TNF-Alpha Inducing Protein Is Associated with Gastric Inflammation and Hyperplasia in a Murine Model of Helicobacter pylori Infection. Front. Pharmacol. 2022, 13, 817237. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, N.; Salimzadeh, L.; Shirzad, H. The Role of T Helper 1-Cell Response in Helicobacter pylori-Infection. Microb. Pathog. 2018, 123, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chen, L.; Sun, H.; Yang, W.; Hu, J.; He, Y.; Wei, S.; Zhao, Z.; Zhang, J.; Li, H.; et al. Immunodominant Epitope-Specific Th1 but Not Th17 Responses Mediate Protection against Helicobacter pylori Infection Following UreB Vaccination of BALB/c Mice. Sci. Rep. 2015, 5, 14793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr-Esfahani, M.; Doosti, A.; Sazegar, H. Evaluation of the Immune Response against Helicobacter pylori in Infused BALB/c Mice by PcDNA3.1(+)-UreA. Folia Med. 2020, 62, 37–45. [Google Scholar] [CrossRef]
- Paydarnia, N.; Mansoori, B.; Esmaeili, D.; Kazemi, T.; Aghapour, M.; Hajiasgharzadeh, K.; Alizadeh, N.; Baradaran, B. Helicobacter pylori Recombinant CagA Regulates Th1/Th2 Balance in a BALB/c Murine Model. Adv. Pharm. Bull. 2020, 10, 264. [Google Scholar] [CrossRef]
- Adamsson, J.; Ottsjö, L.S.; Lundin, S.B.; Svennerholm, A.-M.; Raghavan, S. Gastric Expression of IL-17A and IFNγ in Helicobacter pylori Infected Individuals Is Related to Symptoms. Cytokine 2017, 99, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Arachchi, P.S.; Fernando, N.; Weerasekera, M.M.; Senevirathna, B.; Weerasekera, D.D.; Gunasekara, C.P. Proinflammatory Cytokine IL-17 Shows a Significant Association with Helicobacter pylori Infection and Disease Severity. Gastroenterol. Res. Pract. 2017, 2017, 6265150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagheri, N.; Azadegan-Dehkordi, F.; Shirzad, M.; Zamanzad, B.; Rahimian, G.; Taghikhani, A.; Rafieian-Kopaei, M.; Shirzad, H. Clinical Immunology Mucosal Interleukin-21 MRNA Expression Level Is High in Patients with Helicobacter pylori and Is Associated with the Severity of Gastritis. Central Eur. J. Immunol. 2015, 1, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Davarpanah, E.; Jafarzadeh, A.; Nemati, M.; Bassagh, A.; Abasi, M.H.; Khosravimashizi, A.; Kazemipoor, N.; Ghazizadeh, M.; Mirzaee, M. Circulating Concentration of Interleukin-37 in Helicobacter pylori-Infected Patients with Peptic Ulcer: Its Association with IL-37 Related Gene Polymorphisms and Bacterial Virulence Factor CagA. Cytokine 2020, 126, 154928. [Google Scholar] [CrossRef]
- Michalkiewicz, J.; Helmin-Basa, A.; Grzywa, R.; Czerwionka-Szaflarska, M.; Szaflarska-Poplawska, A.; Mierzwa, G.; Marszalek, A.; Bodnar, M.; Nowak, M.; Dzierzanowska-Fangrat, K. Innate Immunity Components and Cytokines in Gastric Mucosa in Children with Helicobacter pylori Infection. Mediat. Inflamm. 2015, 2015, 176726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahid-Samiei, M.; Rahimian, G.; Shafigh, M.; Taheri, F.; Karami-Hurestani, M.; Sanaei, M.; Heshmati, M.; Bagheri, N. Enhanced Frequency of CD19+IL-10+B Cells in Human Gastric Mucosa Infected by Helicobacter pylori. Am. J. Med. Sci. 2020, 359, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Outlioua, A.; Badre, W.; Desterke, C.; Echarki, Z.; El Hammani, N.; Rabhi, M.; Riyad, M.; Karkouri, M.; Arnoult, D.; Khalil, A.; et al. Gastric IL-1β, IL-8, and IL-17A Expression in Moroccan Patients Infected with Helicobacter pylori May Be a Predictive Signature of Severe Pathological Stages. Cytokine 2020, 126, 154893. [Google Scholar] [CrossRef] [PubMed]
- Zabaglia, L.M.; Sallas, M.L.; Santos, M.P.D.; Orcini, W.A.; Peruquetti, R.L.; Constantino, D.H.; Chen, E.; Smith, M.D.A.C.; Payão, S.-L.M.; Rasmussen, L.T. Expression of MiRNA-146a, MiRNA-155, IL-2, and TNF-α in Inflammatory Response to Helicobacter pylori Infection Associated with Cancer Progression. Ann. Hum. Genet. 2018, 82, 135–142. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X.; Yu, Y.; Xiao, Y.; Huang, J.; Yao, Z.; Chen, X.; Zhou, T.; Li, P.; Xu, C. Serum Exosomes of Chronic Gastritis Patients Infected with Helicobacter pylori Mediate IL-1α Expression via IL-6 Trans-Signalling in Gastric Epithelial Cells: Exosomes Mediate IL-1α by SIL-6R. Clin. Exp. Immunol. 2018, 194, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Beer, L.; Zimmermann, M.; Mitterbauer, A.; Ellinger, A.; Gruber, F.; Narzt, M.-S.; Zellner, M.; Gyöngyösi, M.; Madlener, S.; Simader, E.; et al. Analysis of the Secretome of Apoptotic Peripheral Blood Mononuclear Cells: Impact of Released Proteins and Exosomes for Tissue Regeneration. Sci. Rep. 2015, 5, 16662. [Google Scholar] [CrossRef]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.-L.T.; Schmittgen, T.D.; et al. Detection of MicroRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Park, H.; Kim, H.; Mun, D.; Park, H.; Yun, N.; Joung, B. Human Peripheral Blood-derived Exosomes for MicroRNA Delivery. Int. J. Mol. Med. 2019, 43, 2319–2328. [Google Scholar] [CrossRef] [Green Version]
- Lugini, L.; Cecchetti, S.; Huber, V.; Luciani, F.; Macchia, G.; Spadaro, F.; Paris, L.; Abalsamo, L.; Colone, M.; Molinari, A.; et al. Immune Surveillance Properties of Human NK Cell-Derived Exosomes. J. Immunol. 2012, 189, 2833–2842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazrati, A.; Soudi, S.; Malekpour, K.; Mahmoudi, M.; Rahimi, A.; Hashemi, S.M.; Varma, R.S. Immune Cells-Derived Exosomes Function as a Double-Edged Sword: Role in Disease Progression and Their Therapeutic Applications. Biomark. Res. 2022, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Zhang, L.; Wu, H.; Chen, F.; Liu, X.; Xu, H.; Cui, Y.; Zhu, Q.; Wang, M.; Hao, H.; et al. CagA+ Helicobacter pylori, Not CagA− Helicobacter pylori, Infection Impairs Endothelial Function Through Exosomes-Mediated ROS Formation. Front. Cardiovasc. Med. 2022, 9, 881372. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, L.; Liang, X.; Li, S.; Ma, L.; Zheng, L.; Li, T.; Yu, H.; Chan, H.; Chen, C.; et al. Helicobacter pylori-induced YAP1 Nuclear Translocation Promotes Gastric Carcinogenesis by Enhancing IL-1β Expression. Cancer Med. 2019, 8, 3965–3980. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Zhang, Y.; Qian, L.; Wang, P. Emerging Strategies to Target RAS Signaling in Human Cancer Therapy. J. Hematol. Oncol. 2021, 14, 116. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R.; Llambi, F. Cell Death Signaling. Cold Spring Harb. Perspect. Biol. 2015, 7, a006080. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK Signalling Pathway and Tumorigenesis (Review). Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Kodani, N.; Nakae, J. Tissue-Specific Metabolic Regulation of FOXO-Binding Protein: FOXO Does Not Act Alone. Cells 2020, 9, 702. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/MTOR Pathway and Its Role in Cancer Therapeutics: Are We Making Headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef] [PubMed]
- Rah, B.; Rather, R.A.; Bhat, G.R.; Baba, A.B.; Mushtaq, I.; Farooq, M.; Yousuf, T.; Dar, S.B.; Parveen, S.; Hassan, R.; et al. JAK/STAT Signaling: Molecular Targets, Therapeutic Opportunities, and Limitations of Targeted Inhibitions in Solid Malignancies. Front. Pharmacol. 2022, 13, 821344. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tao, T.; Li, H.; Zhu, X. MTOR Signaling Pathway and MTOR Inhibitors in Cancer: Progress and Challenges. Cell Biosci. 2020, 10, 31. [Google Scholar] [CrossRef]
- Oshima, H.; Ishikawa, T.; Yoshida, G.J.; Naoi, K.; Maeda, Y.; Naka, K.; Ju, X.; Yamada, Y.; Minamoto, T.; Mukaida, N.; et al. TNF-α/TNFR1 Signaling Promotes Gastric Tumorigenesis through Induction of Noxo1 and Gna14 in Tumor Cells. Oncogene 2014, 33, 3820–3829. [Google Scholar] [CrossRef]
- Wang, X.; Lin, Y. Tumor Necrosis Factor and Cancer, Buddies or Foes? Acta Pharmacol. Sin. 2008, 29, 1275–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jafarzadeh, A.; Nemati, M.; Jafarzadeh, S. The Important Role Played by Chemokines Influence the Clinical Outcome of Helicobacter pylori Infection. Life Sci. 2019, 231, 116688. [Google Scholar] [CrossRef] [PubMed]
- Charitos, I.A.; D’Agostino, D.; Topi, S.; Bottalico, L. 40 Years of Helicobacter pylori: A Revolution in Biomedical Thought. Gastroenterol. Insights 2021, 12, 111–135. [Google Scholar] [CrossRef]
- Uhl, B.; Prochazka, K.T.; Fechter, K.; Pansy, K.; Greinix, H.T.; Neumeister, P.; Deutsch, A.J. Impact of the Microenvironment on the Pathogenesis of Mucosa-Associated Lymphoid Tissue Lymphomas. World J. Gastrointest. Oncol. 2022, 14, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Mello, C.; Sevéri, E.; Ricci, E.; Marangoni, A.; Coelho, L.; Ribeiro, D.; Poppi, R.J. Fast Differentiation of Bacteria Causing Pharyngitis by Low Resolution Raman Spectroscopy and PLS-Discriminant Analysis. J. Braz. Chem. Soc. 2008, 19, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3.22.1–3.22.29. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein–Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atrisco-Morales, J.; Ramírez, M.; Castañón-Sánchez, C.A.; Román-Román, A.; Román-Fernández, I.V.; Martínez-Carrillo, D.N.; García-Arellano, S.; Muñoz-Valle, J.F.; Rodríguez-Ruiz, H.A.; Fernández-Tilapa, G. In Peripheral Blood Mononuclear Cells Helicobacter pylori Induces the Secretion of Soluble and Exosomal Cytokines Related to Carcinogenesis. Int. J. Mol. Sci. 2022, 23, 8801. https://doi.org/10.3390/ijms23158801
Atrisco-Morales J, Ramírez M, Castañón-Sánchez CA, Román-Román A, Román-Fernández IV, Martínez-Carrillo DN, García-Arellano S, Muñoz-Valle JF, Rodríguez-Ruiz HA, Fernández-Tilapa G. In Peripheral Blood Mononuclear Cells Helicobacter pylori Induces the Secretion of Soluble and Exosomal Cytokines Related to Carcinogenesis. International Journal of Molecular Sciences. 2022; 23(15):8801. https://doi.org/10.3390/ijms23158801
Chicago/Turabian StyleAtrisco-Morales, Josefina, Mónica Ramírez, Carlos Alberto Castañón-Sánchez, Adolfo Román-Román, Ilce Valeria Román-Fernández, Dinorah Nashely Martínez-Carrillo, Samuel García-Arellano, José Francisco Muñoz-Valle, Hugo Alberto Rodríguez-Ruiz, and Gloria Fernández-Tilapa. 2022. "In Peripheral Blood Mononuclear Cells Helicobacter pylori Induces the Secretion of Soluble and Exosomal Cytokines Related to Carcinogenesis" International Journal of Molecular Sciences 23, no. 15: 8801. https://doi.org/10.3390/ijms23158801