
The Mechanism of Peach Gum Polysaccharide Preventing UVB-Induced Skin Photoaging by Regulating Matrix Metalloproteinanse and Oxidative Factors

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Properties of Peach Gum Polysaccharide
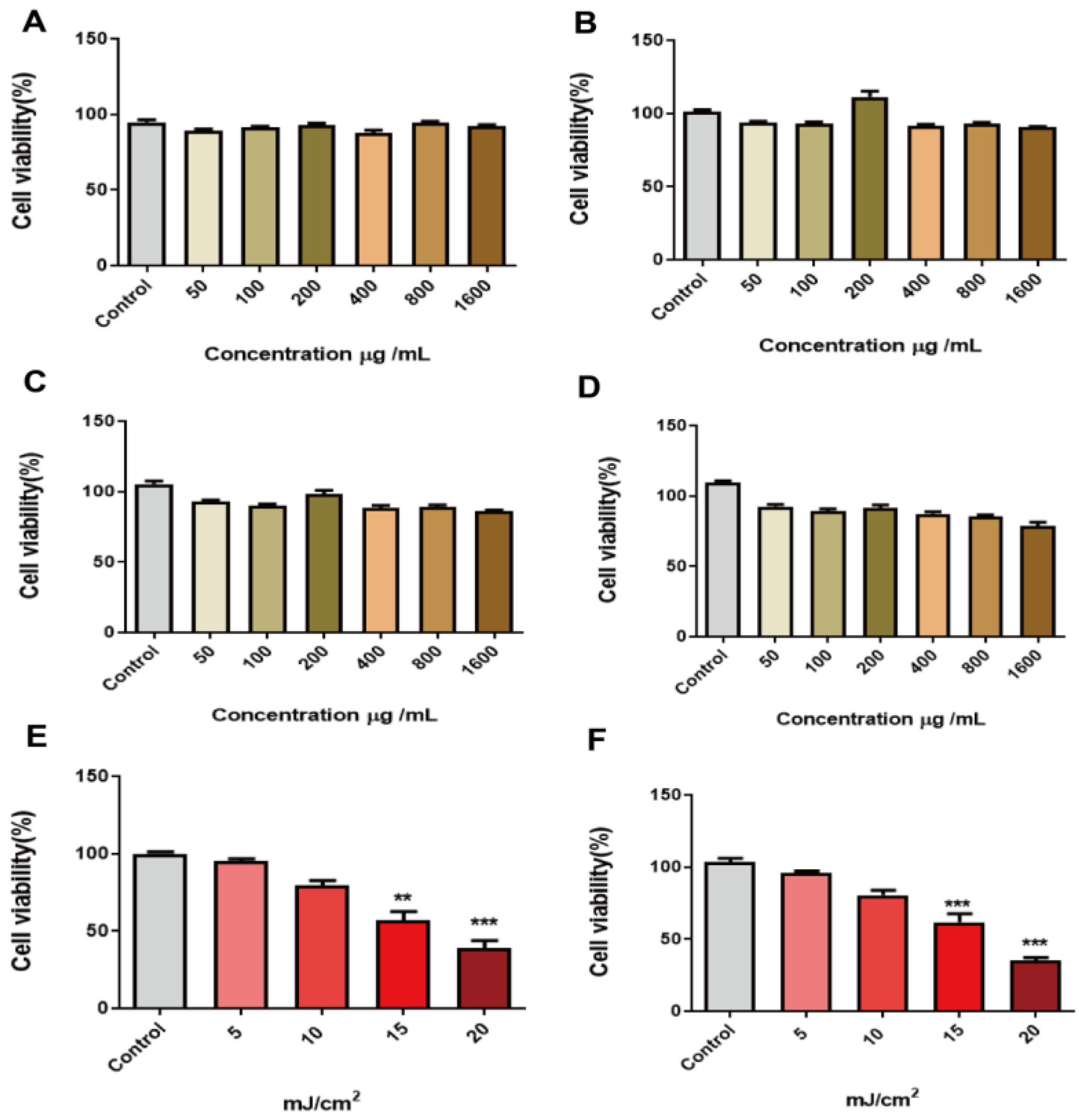
2.2. Effect of Peach Gum Polysaccharide on HaCaT Cells
2.3. UVB Radiation Dose Dependently Reduced the Viability of HaCaT Cells
2.4. Peach Gum Polysaccharide Protects HaCaT Cells from UVB-Induced Apoptosis
2.5. Effect of Peach Gum Polysaccharide on the Production of Oxidative Factors in HaCaT Cells
2.6. Effect of Peach Gum Polysaccharide on the Production of Antioxidant Factors in HaCaT Cells
2.7. Peach Gum Polysaccharide Reduced UVB-Induced Matrix Metalloproteinase Expression
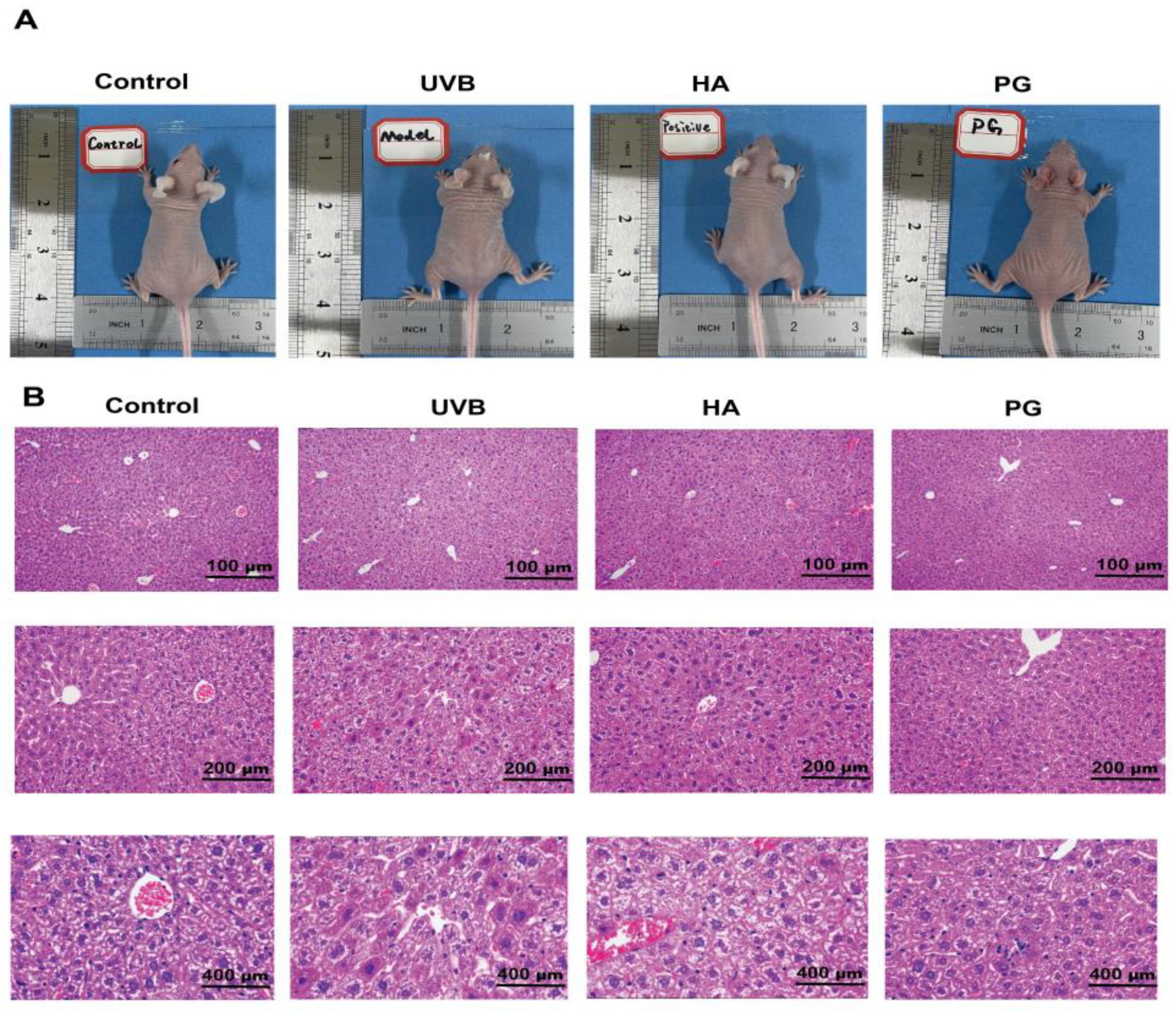
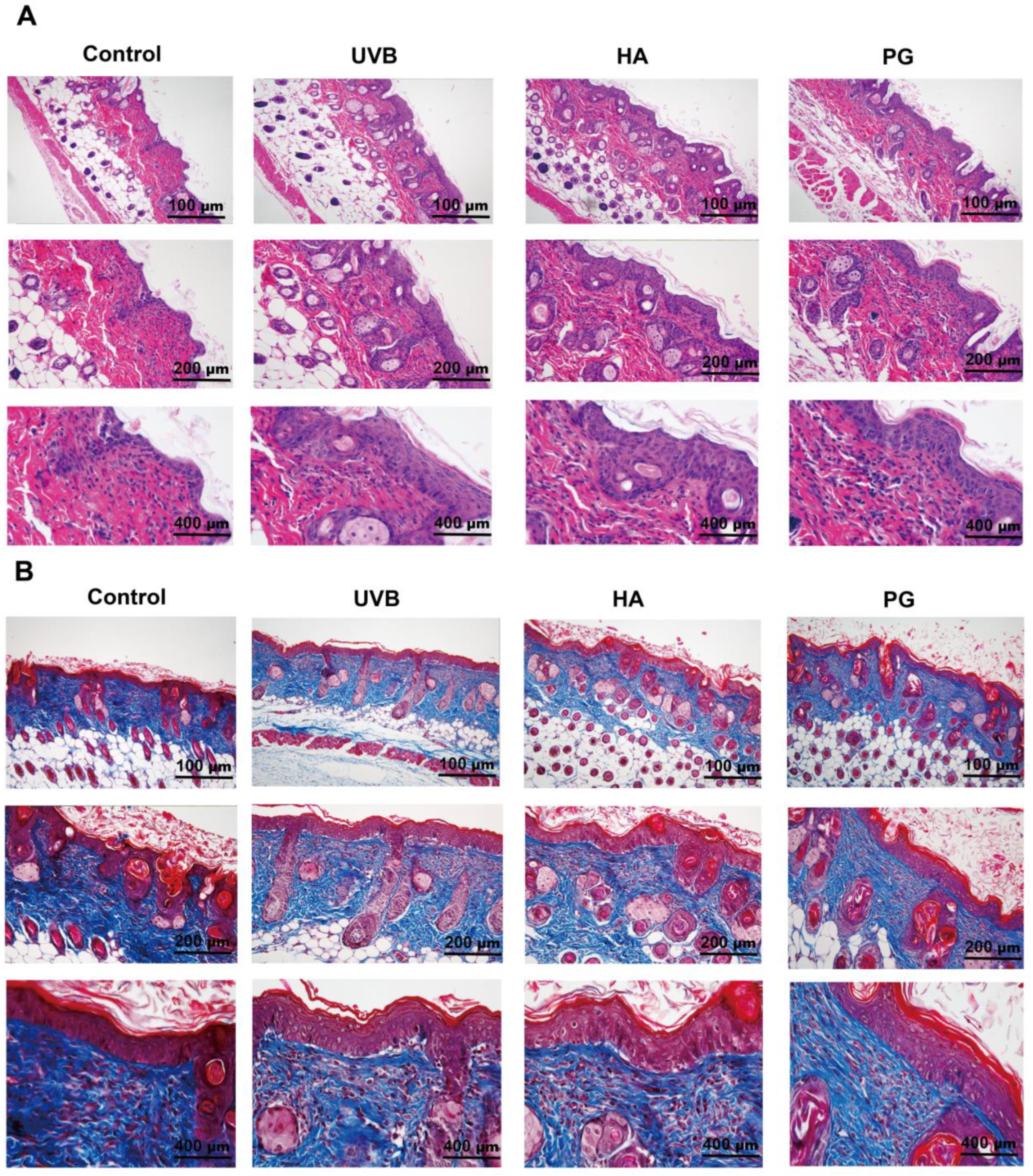
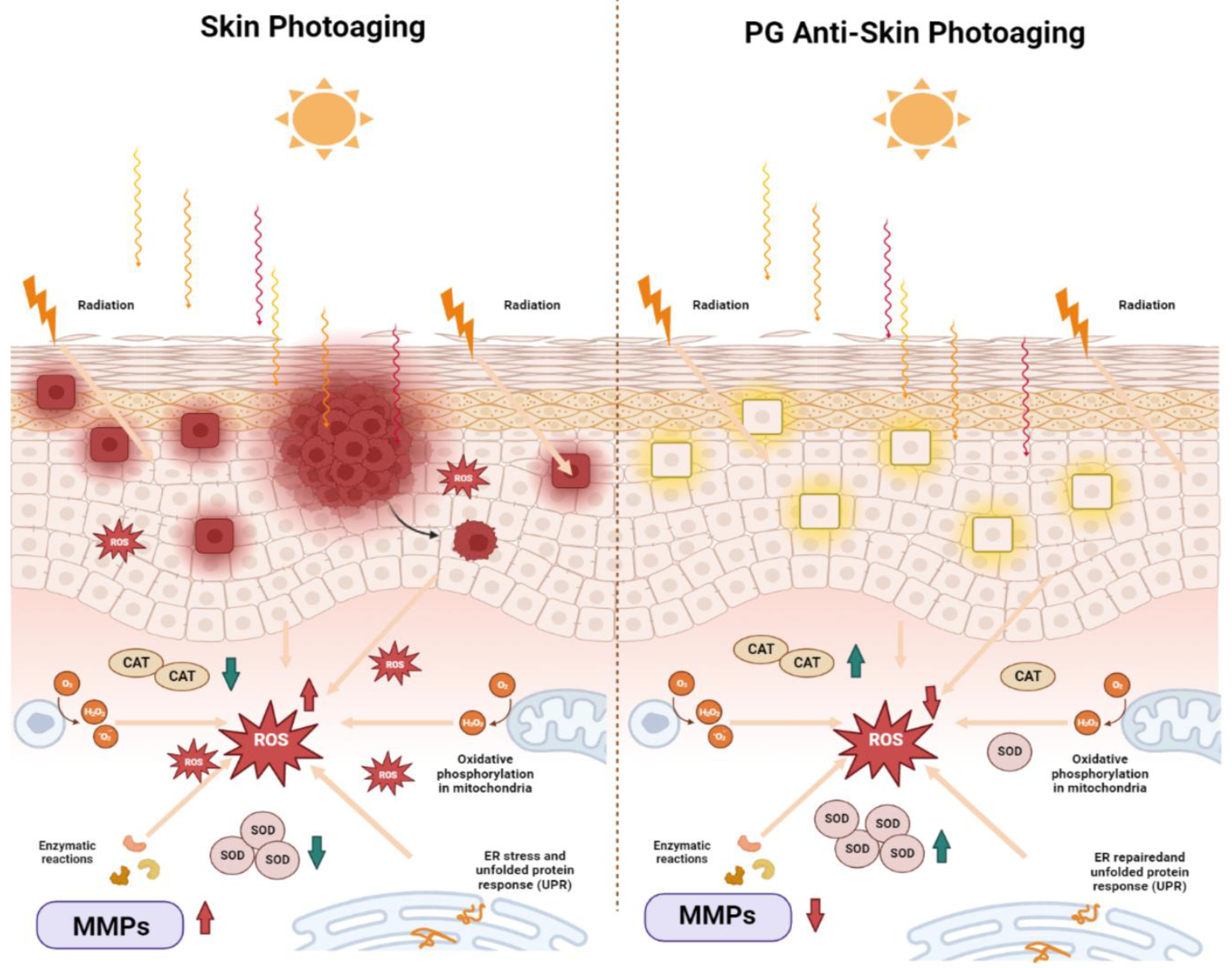
2.8. Peach Gum Polysaccharide Alleviates UVB-Induced Skin Photoaging Damage In Vivo
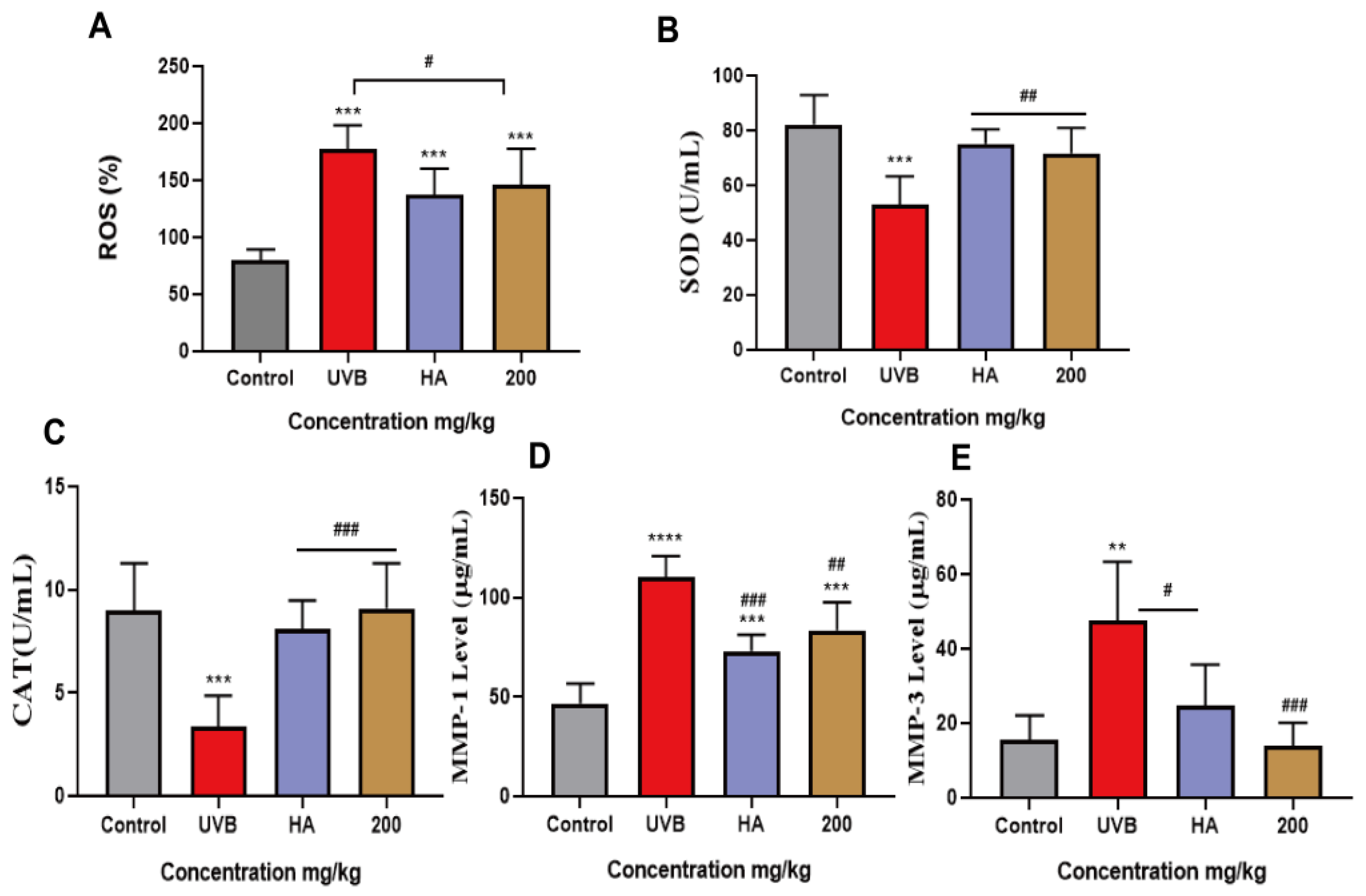
2.9. Peach Gum Polysaccharide Alleviated UVB-Induced Skin Photoaging In Vivo by Regulating MMPs and Oxidative Factors
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Reagents
4.2. Preparation of Polysaccharides from Peach Gum
4.3. Average Molecular Weight (Mw) Determination
4.4. Monosaccharide Composition Analysis
4.5. Determination of the Chemical Composition
4.6. Cell Line Culture and UVB Irradiation
4.7. MTT Cell Viability Assay
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. In Vivo Animal Photoaging Model
4.10. Skin Tissue Histological Analysis
4.11. Skin Tissue Masson Staining Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lee, H.; Hong, Y.; Kim, M. Structural and Functional Changes and Possible Molecular Mechanisms in Aged Skin. Int. J. Mol. Sci. 2021, 22, 12489. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.-J.; Kang, S.; Varani, J.; Csorgoet, Z.-B.; Wan, Y.-S.; Datta, S.; Voorhees, J.-J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.-P.; Han, J.-X.; Jiang, C.-P.; Zhang, Y. Biomarkers, oxidative stress and autophagy in skin aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- Song, X.-Z.; Narzt, M.-S.; Nagelreiter, I.-M.; Hohensinner, P.; Zaniewicz, L.-T.; Tschachler, E.; Grillari, J.; Gruber, F. Autophagy deficient keratinocytes display increased DNA damage, senescence and aberrant lipid composition after oxidative stress in vitro and in vivo. Redox. Biol. 2017, 11, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.; Ren, R.; Jia, C.-L.; Lu, Y.-Z.; Yang, Q.-J.; Chen, L.; Wu, X.-Y.; Zhu, J.-J.; Guo, Y.; Yang, P.; et al. Rapamycin Protects Skin Fibroblasts from Ultraviolet B-Induced Photoaging by Suppressing the Production of Reactive Oxygen Species. Cell. Physiol. Biochem. 2018, 46, 1849–1860. [Google Scholar] [CrossRef]
- Lamore, S.-D.; Wondrak, G.-T. UVA causes dual inactivation of cathepsin B and L underlying lysosomal dysfunction in human dermal fibroblasts. J. Photochem. Photobiol. B Biol. 2013, 123, 1–12. [Google Scholar] [CrossRef]
- Yao, W.-Z.; Chen, X.-Y.; Li, X.; Chang, S.-Y.; Zhao, M.-M.; You, L.-J. Current trends in the anti-photoaging activities and mechanisms of dietary non-starch polysaccharides from natural resources. Crit. Rev. Food. Sci. Nutr. 2022, 62, 9021–9035. [Google Scholar] [CrossRef]
- Demirovic, D.; Nizard, C.; Rattan, S.-I. Basal level of autophagy is increased in aging human skin fibroblasts in vitro, but not in old skin. PLoS ONE 2015, 10, e0126546. [Google Scholar] [CrossRef]
- Csekes, E.; Račková, L. Skin Aging, Cellular Senescence and Natural Polyphenols. Int. J. Mol. Sci. 2021, 22, 12641. [Google Scholar] [CrossRef]
- Wang, Y.-J.; Ouyang, Q.-Q.; Chang, X.-F.; Yang, M.; He, J.-P.; Tian, Y.; Sheng, J. Anti-photoaging effects of flexible nanoliposomes encapsulated Moringa oleifera Lam. isothiocyanate in UVB-induced cell damage in HaCaT cells. Drug Deliv. 2022, 29, 871–881. [Google Scholar] [CrossRef]
- Choi, B.B.-R.; Choi, J.-.H.; Ji, J.; Song, K.-W.; Lee, H.-J.; Kim, G.-C. Increment of growth factors in mouse skin treated with non-thermal plasma. Int. J. Med. Sci. 2018, 15, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Qomaladewi, N.-P.; Kim, M.-Y.; Cho, J.-Y. Rottlerin Reduces cAMP/CREB-Mediated Melanogenesis via Regulation of Autophagy. Int. J. Mol. Sci. 2019, 20, 2081. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Kim, J.-H.; Kim, C.-T.; Jeong, W.-S.; Kim, H.-M.; Sim, J.-H.; Kang, J.-S. Evaluation of Anti-Melanogenesis Activity of Enriched Pueraria lobata Stem Extracts and Characterization of Its Phytochemical Components Using HPLC-PDA-ESI-MS/MS. Int. J. Mol. Sci. 2021, 22, 8105. [Google Scholar] [CrossRef] [PubMed]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef]
- Liu, Y.; Chan, F.-X.; Sun, H.-M.; Yan, J.-H.; Fan, D.-Y.; Zhao, D.-Z.; An, J.; Zhou, D.-S. Resveratrol protects human keratinocytes HaCaT cells from UVA-induced oxidative stress damage by downregulating Keap1 expression. Eur. J. Pharmacol. 2011, 650, 130–137. [Google Scholar] [CrossRef]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.-V.-C.; Stuppner, H.; Jansen, D.-P. Plant extracts and natural compounds used against UVB-induced photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef]
- Cavinato, M.; Jansen, D.-P. Molecular mechanisms of UVB-induced senescence of dermal fibroblasts and its relevance for photoaging of the human skin. Exp. Gerontol. 2017, 94, 78–82. [Google Scholar] [CrossRef]
- Heng, M.-C. Signaling pathways targeted by curcumin in acute and chronic injury: Burns and photo-damaged skin. Int. J. Dermatol. 2013, 52, 531–543. [Google Scholar] [CrossRef]
- Christensen, L.; Suggs, A.; Baron, E. Ultraviolet Photobiology in Dermatology. Adv. Exp. Med. Biol. 2017, 996, 89–104. [Google Scholar]
- Mittal, M.; Siddiqui, M.-R.; Tran, K.; Reddy, S.-P.; Malik, A.-B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox. Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Wen, X.; Hao, D.; Zhang, N.; He, G.; Jiang, X. NF-κB signaling in skin aging. Mech. Ageing. Dev. 2019, 184, 111160. [Google Scholar] [CrossRef] [PubMed]
- Kambayashi, H.; Yamashita, M.; Odake, Y.; Takada, K.; Funasaka, Y.; Ichihashi, M. Epidermal changes caused by chronic low-dose UV irradiation induce wrinkle formation in hairless mouse. J. Dermatol. Sci. 2001, 27, S19–S25. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.W.; Ahn, S.I.; Jhoo, J.W.; Kim, G.Y. Antioxidant activity of yogurt fermented at low temperature and its anti-inflammatory effect on DSS-induced colitis in Mice. Food Sci. Anim. Resour. 2019, 39, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Kim, D.; Kim, N.Y.; Kim, J.S.; Jang, A. Anti-wrinkle and anti-inflflammatory effects of a combination of topically applied horse oil and dietary enzyme hydrolysates from horse bone. Process Biochem. 2020, 90, 257–267. [Google Scholar] [CrossRef]
- Kim, J.-H.; Gao, D.; Jeong, W.-S.; Kim, C.-T.; Cho, C.-W.; Kim, H.-M.; Kang, J.-S. Anti-Wrinkle Effect of Isatis indigotica Leaf Extract: Evaluation of Antioxidant, Anti-Inflammation, and Clinical Activity. Antioxidants 2021, 10, 1339. [Google Scholar] [CrossRef]
- Kurz, C.; Carle, R.; Schieber, A. Characterisation of cell wall polysaccharide profiles of apricots (Prunus armeniaca L.), peaches (Prunus persica L.), and pumpkins (Cucurbita sp.) for the evaluation of fruit product authenticity. Food. Chem. 2008, 106, 421–430. [Google Scholar] [CrossRef]
- Ding, J.; Zhang, H.; Tian, Y.; Lai, P.-F.-H.; Xu, H.; Ai, L. Rheological properties of Prunus persica exudate: Potential effects of proteins and polyphenols. Int. J. Biol. Macromol. 2019, 133, 831–838. [Google Scholar] [CrossRef]
- Wang, Y.-T.; Lin, D.-B.; Wang, X.-L.; Zhu, W.; Ye, J.-L.; Li, G.-H.; Ma, Z.-C.; Deng, X.-X. The impact of a novel peach gum-derived polysaccharide on postprandial blood glucose control in streptozotocin-induced diabetic mice. Int. J. Biol. Macromol. 2017, 98, 379–386. [Google Scholar] [CrossRef]
- SimasTosin, F.-F.; Abud, A.-P.; de Oliveira, C.-C.; Gorin, P.; Sassaki, G.-L.; Bucchi, D.-F.; Iacominiet, M. Polysaccharides from peach pulp: Structure and effects on mouse peritoneal macrophages. Food Chem. 2012, 134, 2257–2260. [Google Scholar] [CrossRef]
- Wei, C.-Y.; Yao, L.; Zhang, Y.; Qiu, S.-Y.; Zeng, X.-Y.; Chen, S.-G.; Ye, X.-Q. Structural characterization of peach gum polysaccharide and its effects on the regulation of DSS-induced acute colitis. Int. J. Biol. Macromol. 2023, 225, 1224–1234. [Google Scholar] [CrossRef]
- Chen, J.-X.; Zhou, M.; Liu, M.; Bi, J.-F. Physicochemical, rheological properties and in vitro hypoglycemic activities of polysaccharide fractions from peach gum. Carbohydr. Polym. 2022, 296, 119954. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.-L.; Peng, B.-X.; Ding, H.-H.; Cui, B.-B.; Nie, H.; Yan, Y.-Z. Purification, structure and biological activity of pumpkin (Cucurbita moschata) polysaccharides: A review. Food. Rev. Int. 2021, 39, 307–319. [Google Scholar] [CrossRef]
- Mellinger, C.-G.; Carbonero, E.-R.; Noleto, G.-R.; Cipriani, T.-R.; Oliveira, M.-B.-M.; Gorin, A.-J.-P.; Iacominiet, M. Chemical and biological properties of an arabinogalactan from Phyllanthusniruri. J. Nat. Prod. 2005, 68, 1479–1483. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.-L.; Guo, J.-H.; Cao, T.; Zhang, T.; Liu, Y.; Yan, Y. Review on mechanisms and structure-activity relationship of hypoglycemic effects of polysaccharides from natural resources. Food Sci. Hum. Wellness 2023, 12, 1969–1980. [Google Scholar] [CrossRef]
- Fernando, I.-P.-S.; Dias, M.K.H.M.; Madusanka, D.M.D.; Han, E.-J.; Kim, M.-J.; Jeon, Y.-J.; Lee, K.-H.; Cheong, S.-H.; Han, Y.-S.; Park, S.-R.; et al. Human Keratinocyte UVB-Protective Effects of a Low Molecular Weight Fucoidan from Sargassum horneri Purified by Step Gradient Ethanol Precipitation. Antioxidants 2020, 9, 340. [Google Scholar] [CrossRef]
- Zhang, Y.-T.; You, S.-Q.; Wang, D.-D.; Zhao, D.; Zhang, J.-C.; An, Q.; Li, M.; Wang, C.-T. Fermented Dendrobium officinale polysaccharides protect UVA-induced photoaging of human skin fibroblasts. Food. Sci. Nutr. 2022, 10, 1275–1288. [Google Scholar] [CrossRef]
- Ji, X.-L.; Cheng, Y.-Q.; Tian, J.-Y.; Zhang, S.-Q.; Jing, Y.-S.; Shi, M.-M. Structural characterization of polysaccharide from jujube (Ziziphus jujuba Mill.) fruit. Chem. Biol. Technol. Agric. 2021, 8, 54. [Google Scholar] [CrossRef]
- Ji, D.-S.; You, L.-J.; Ren, Y.-L.; Zheng, G.; Li, C. Protective effect of polysaccharides from Sargassum fusiforme against UVB-induced oxidative stress in HaCaT human keratinocytes. J. Funct. Foods 2017, 36, 332–340. [Google Scholar] [CrossRef]
- Ji, X.-L.; Hou, C.-Y.; Yan, Y.-Z.; Shi, M.-M.; Liu, Y.-Q. Comparison of structural characterization and antioxidant activity of polysaccharides from jujube (Ziziphus jujuba Mill.) fruit. Int. J. Biol. Macromol. 2020, 149, 1008–1018. [Google Scholar] [CrossRef]
- Choi, S.-I.; Lee, J.-H.; Kim, J.-M.; Jung, T.-D.; Cho, B.-Y.; Choi, S.-H.; Lee, D.-W.; Kim, J.-K.; Kim, J.-Y.; Lee, O.-H. Ulmus macrocarpa Hance Extracts Attenuated H2O2 and UVB-Induced Skin Photo-Aging by Activating Antioxidant Enzymes and Inhibiting MAPK Pathways. Int. J. Mol. Sci. 2017, 18, 1200. [Google Scholar] [CrossRef]
- Dai, J.-R.; Ma, H.-Y.; Fan, J.; Li, Y.-Z.; Wang, J.-G.; Ni, H.-M.; Xia, G.-M.; Chen, S.-Y. Crude polysaccharide from an anti-UVB cell clone of Bupleurum scorzonerifolium protect HaCaT cells against UVB-induced oxidative stress. Cytotechnology 2011, 63, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.-Z.; Li, D.-D.; Luo, H.; Li, W.-J.; Huang, Y.-M.; Li, J.-C.; Hu, Z.; Huang, N.; Guo, M.-H.; Chen, Y.; et al. Anti-photoaging effects of chitosan oligosaccharide in ultraviolet-irradiated hairless mouse skin. Exp. Gerontol. 2018, 103, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.-S.; Meng, X.-Q.; Wang, Z.-B.; Liu, J.-L.; Kuang, H.-X.; Wang, Q.-H. Polysaccharide from Ephedra sinica Stapf inhibits inflammation expression by regulating Factor-β1/Smad2 signaling. Int. J. Biol. Macromol. 2018, 106, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-Y.; Huang, T.; Lan, C.; Ding, H.; Yan, C.-S.; Dou, Y.-L. Protective effect of polysaccharide from Sophora japonica L. flower buds against UVB radiation in a human keratinocyte cell line (HaCaT cells). J. Photochem. Photobiol. 2019, 191, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-P.; Li, Z.-J.; Peng, L.-Q.; Jiang, N.; Liu, Q.; Zhang, E.; Liang, B.-H.; Li, R.-X.; Zhu, H.-L. Lycium barbarum polysaccharide protects human keratinocytes against UVB-induced photo-damage. Free Radic. Res. 2017, 51, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Z.; Liu, P.-P.; You, S.-Q.; Zhao, D.; An, Q.; Wang, D.-D.; Zhang, J.-C.; Li, M.; Wang, C.-T. Anti-Inflammatory Effects of Opuntia Milpa Alta Polysaccharides Fermented by Lactic Acid Bacteria in Human Keratinocyte HaCaT Cells. Chem. Biodivers. 2022, 19, e202100923. [Google Scholar] [CrossRef] [PubMed]
- Poon, F.; Kang, S.; Chien, A.L. Mechanisms and treatments of photoaging. Photoimmunol. Photomed. 2015, 31, 65–74. [Google Scholar] [CrossRef]
- Roh, E.; Kim, J.-E.; Kwon, J.-Y.; Park, J.-S.; Bode, A.-M.; Dong, Z.-G.; Lee, K.-W. Molecular mechanisms of green tea polyphenols with protective effects against skin photoaging. Crit. Rev. Food. Sci. Nutr. 2017, 57, 1631–1637. [Google Scholar] [CrossRef]
- Zeng, S.-H.; Long, J.-W.; Sun, J.-H.; Wang, G.; Zhou, L. A review on peach gum polysaccharide: Hydrolysis, structure, properties and applications. Carbohydr. Polym. 2022, 279, 119015. [Google Scholar] [CrossRef]
- Tang, J.; Xiong, L.-D.; Shu, X.-H.; Chen, W.; Li, W.-J.; Li, J.; Ma, L.-Z.; Xiao, Y.; Li, L. Antioxidant effects of bioactive compounds isolated from cordyceps and their protective effects against UVB-irradiated HaCaT cells. J. Cosmet. Dermatol. 2019, 18, 1899–1906. [Google Scholar] [CrossRef]
- Radjei, S.; Friguet, B.; Nizard, C.; Petropoulos, I. Prevention of dicarbonyl-mediated advanced glycation by glyoxalases: Implication in skin aging. Biochem. Soc. Trans. 2014, 42, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Li, X.; Sun-Waterhouse, D.-X.; Zhu, B.-Y.; You, L.-J.; Hileuskaya, K. Polysaccharides from Sargassum fusiforme after UV/H2O2 degradation effectively ameliorate dextran sulfate sodium-induced colitis. Food. Funct. 2021, 12, 11747–11759. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.-L.; Guo, J.-H.; Ding, D.-Q.; Gao, J.; Hao, L.-R.; Guo, X.-D.; Liu, Y.-Q. Structural characterization and antioxidant activity of a novel high-molecular-weight polysaccharide from Ziziphus Jujuba cv. Muzao. J. Food Meas. Charact. 2022, 16, 2191–2200. [Google Scholar] [CrossRef]
- Dairi, N.; Ferfera-Harrar, H.; Ramos, M.; Garrigós, M.C. Cellulose acetate/AgNPs-organoclay and/or thymol nano-biocomposite films with combined antimicrobial/antioxidant properties for active food packaging use. Int. J. Biol. Macromol. 2019, 121, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.-C.; Cao, Y.; Wu, S.-J. Antioxidant activity and antibacterial activity of peach gum derived oligosaccharides. Int. J. Biol. Macromol. 2013, 62, 1–3. [Google Scholar] [CrossRef]
- Yao, X.-C.; Chang, C.-F.; Wu, S.-J. Effect of peach gum polysaccharides on quality changes of white shrimp. Int. J. Biol. Macromol. 2015, 72, 1076–1080. [Google Scholar] [CrossRef]
- Li, C.-Z.; Tao, J.; Zhang, H.-X. Peach gum polysaccharides-based edible coatings extend shelf life of cherry tomatoes. 3 Biotech 2017, 7, 168. [Google Scholar] [CrossRef]
- Li, C.; Wang, X.-J.; Meng, D.-Y.; Zhou, L. Facile synthesis of low-cost magnetic biosorbent from peach gum polysaccharide for selective and efficient removal of cationic dyes. Int. J. Biol. Macromol. 2018, 107, 1871–1878. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Composition | PG |
|---|---|
| Total sugars | 82.34% |
| Protein | 0.18% |
| Uronic acids (g/100 g) | 12.50 |
| Mp (g/mol) | 4.17 × 106 |
| Mn (g/mol) | 4.05 × 106 |
| Mw (g/mol) | 4.10 × 106 |
| Monosaccharide | PG (mg/g) |
|---|---|
| Man | 21.96 |
| Rha | 7.82 |
| Glu | 0.81 |
| Gal | 348.69 |
| Xyl | 70.63 |
| Ara | 362.71 |
| Rib | 0.91 |
| Gul A | 0.40 |
| Glu A | 65.91 |
| Gal A | 0.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Tao, L.; Wang, Z.; Li, L.; Luo, J.; Pai, K.; Li, W.; Zhao, C.; Sheng, J.; Tian, Y. The Mechanism of Peach Gum Polysaccharide Preventing UVB-Induced Skin Photoaging by Regulating Matrix Metalloproteinanse and Oxidative Factors. Molecules 2023, 28, 4104. https://doi.org/10.3390/molecules28104104
Yang M, Tao L, Wang Z, Li L, Luo J, Pai K, Li W, Zhao C, Sheng J, Tian Y. The Mechanism of Peach Gum Polysaccharide Preventing UVB-Induced Skin Photoaging by Regulating Matrix Metalloproteinanse and Oxidative Factors. Molecules. 2023; 28(10):4104. https://doi.org/10.3390/molecules28104104
Chicago/Turabian StyleYang, Min, Liang Tao, Zilin Wang, Lingfei Li, Junyi Luo, Kuannu Pai, Weitong Li, Cunchao Zhao, Jun Sheng, and Yang Tian. 2023. "The Mechanism of Peach Gum Polysaccharide Preventing UVB-Induced Skin Photoaging by Regulating Matrix Metalloproteinanse and Oxidative Factors" Molecules 28, no. 10: 4104. https://doi.org/10.3390/molecules28104104