
Effects of Four Extraction Methods on Structure and In Vitro Fermentation Characteristics of Soluble Dietary Fiber from Rape Bee Pollen
 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Extraction Yield, Monosaccharide Composition and Molecular Weight of Bee Pollen SDFs
2.2. Scanning Electron Microscopy (SEM)
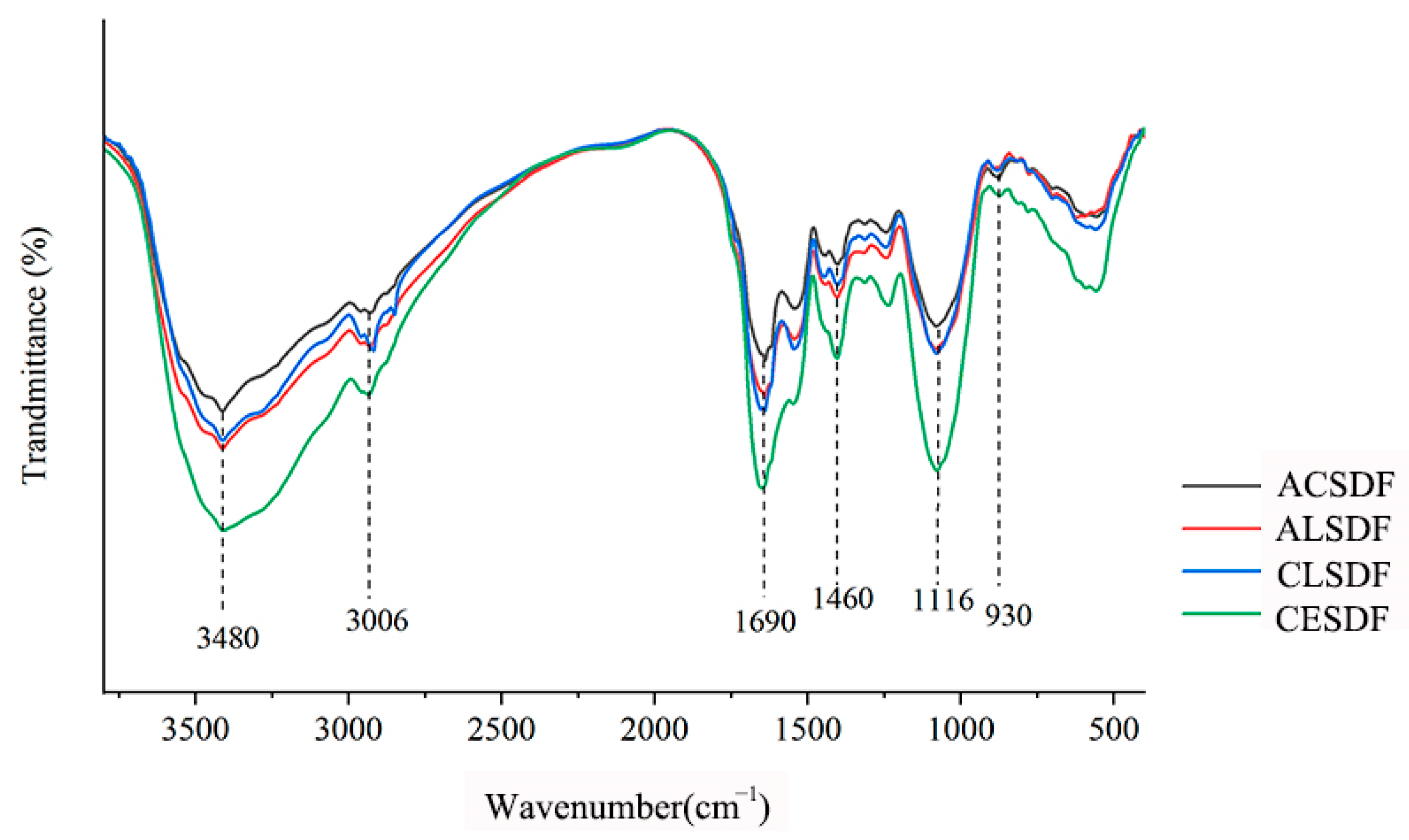
2.3. Fourier Transform Infrared Spectroscopy (FT-IR)
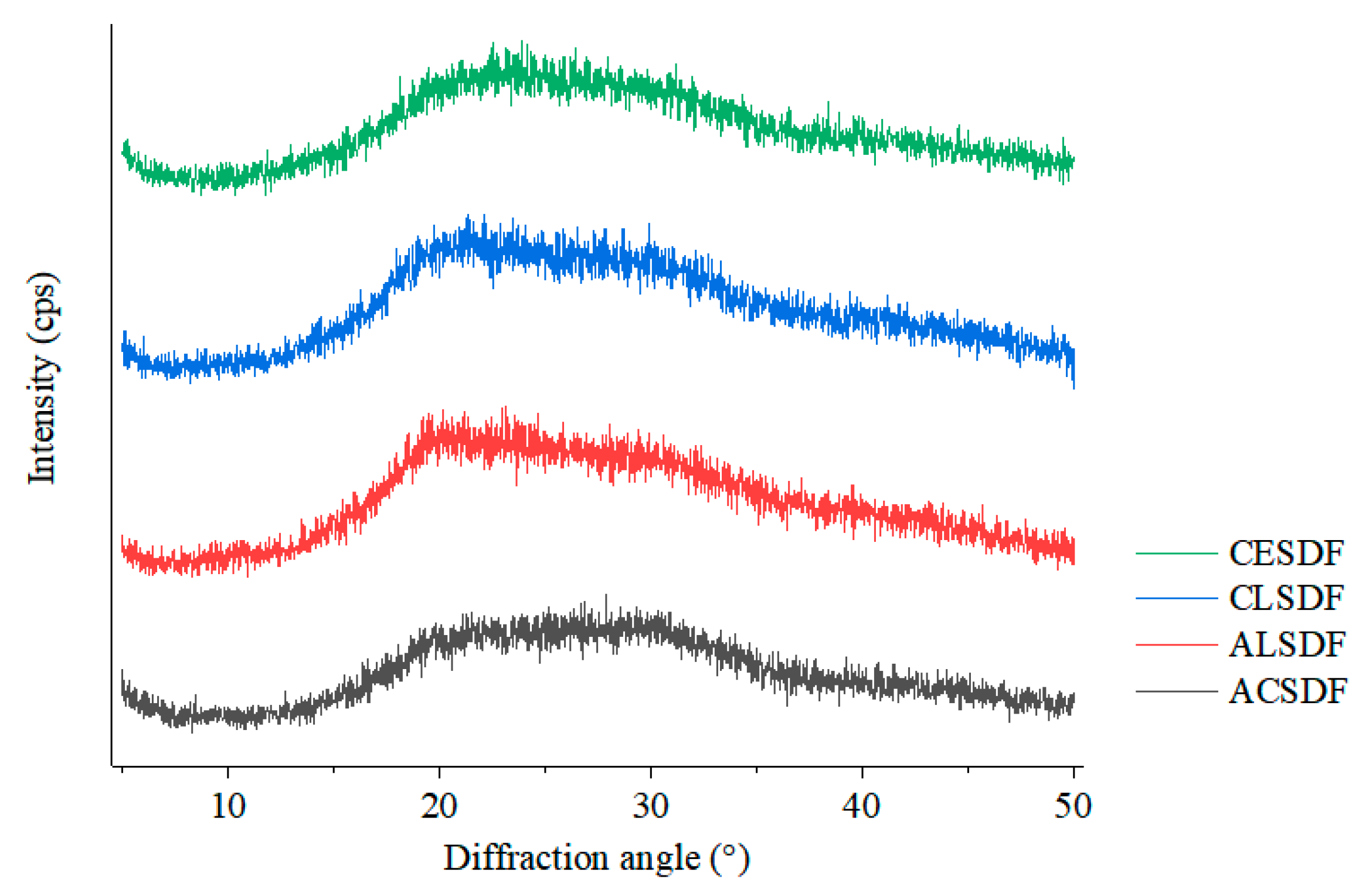
2.4. X-ray Diffraction (XRD)
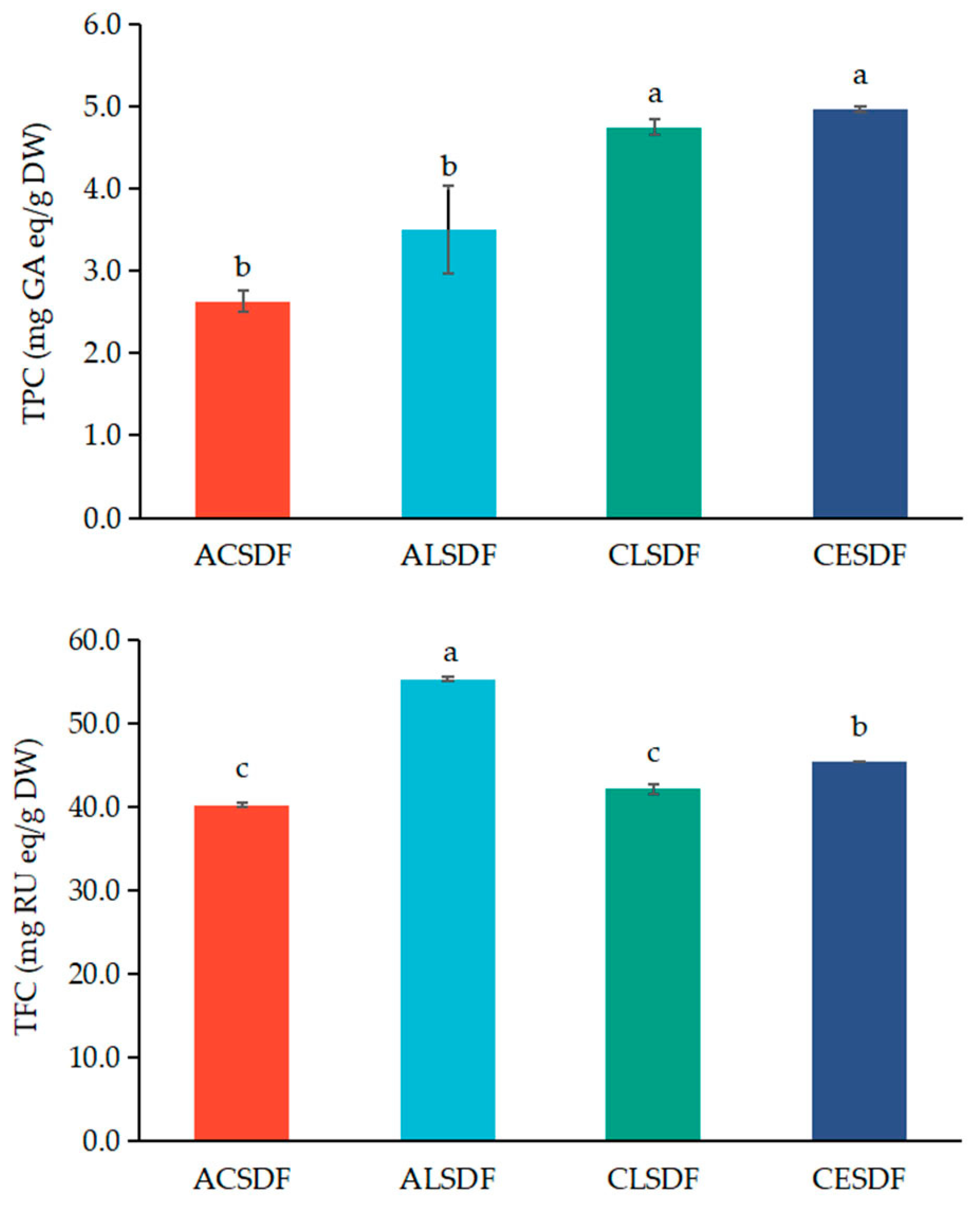
2.5. Phenolic Compounds Determination of Bee Pollen SDFs
2.6. SCFA Concentration after In Vitro Fermentation
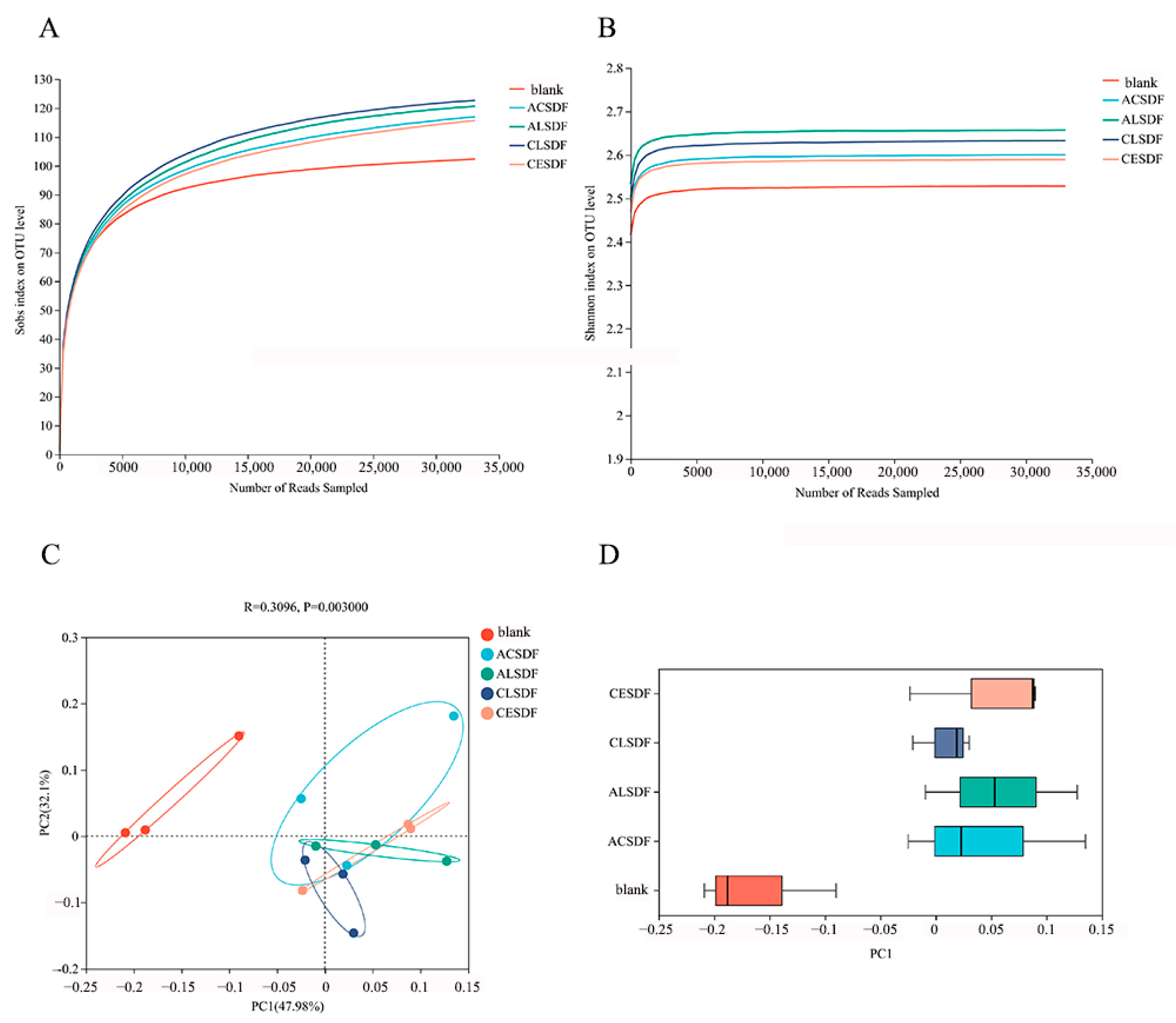
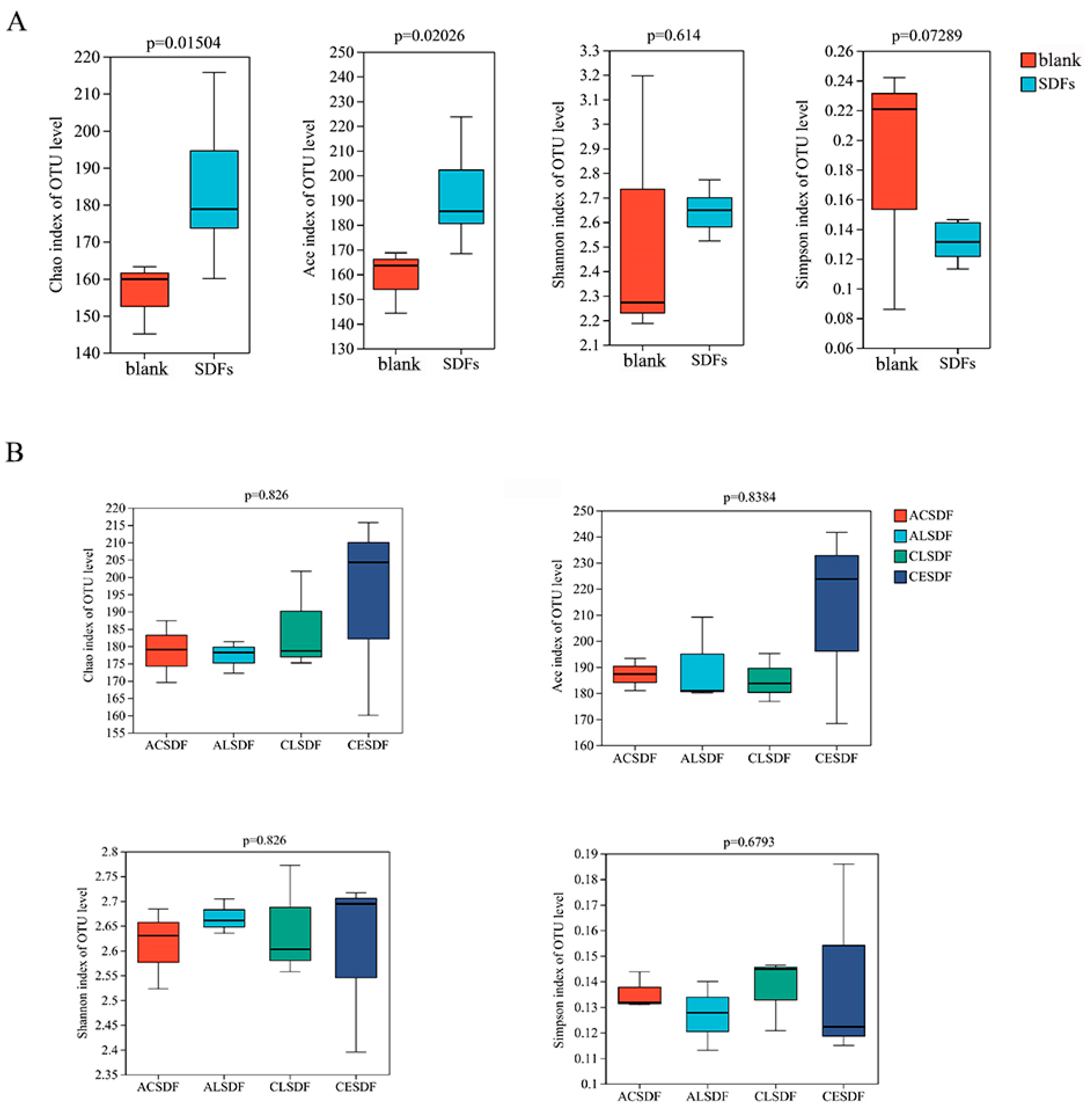
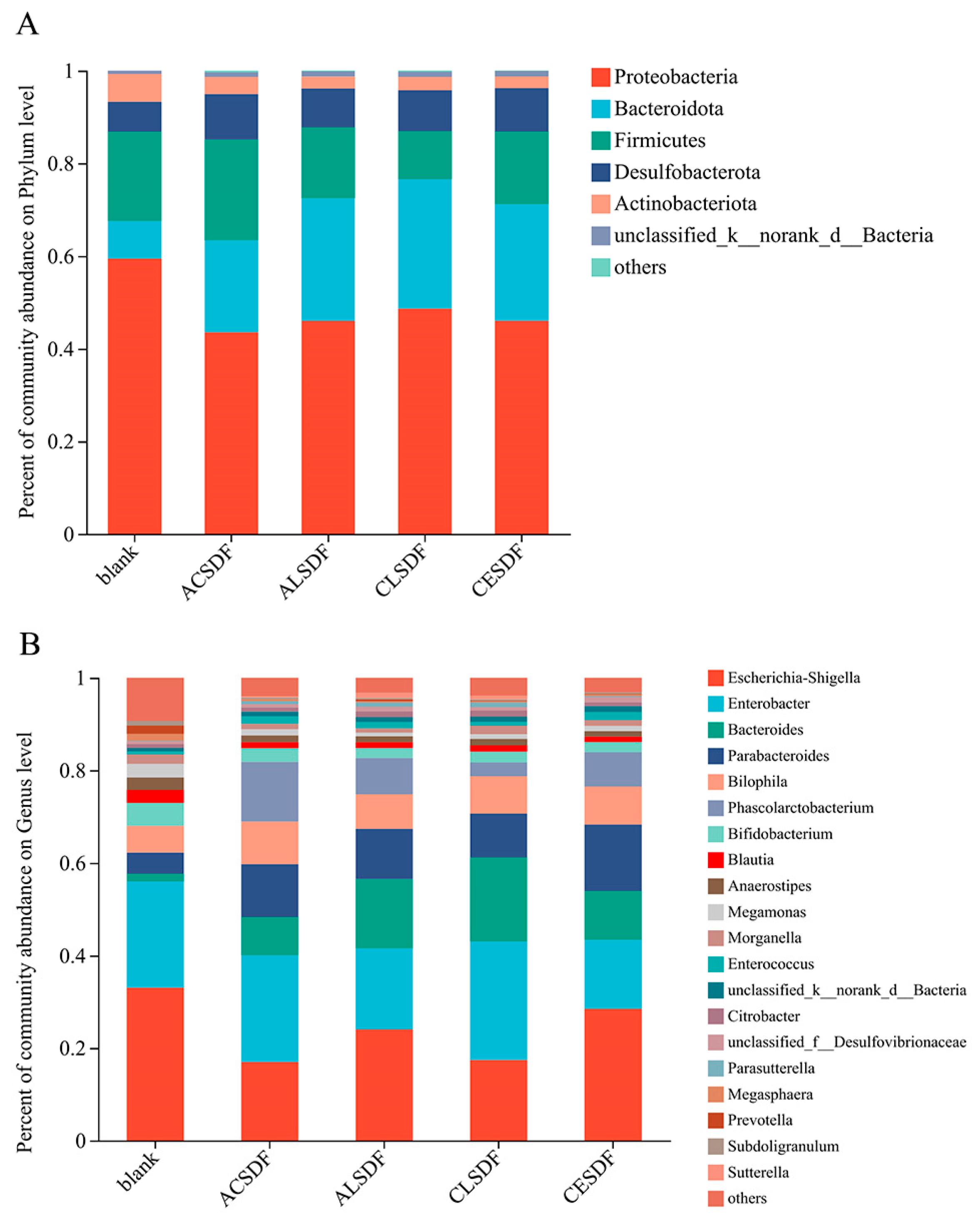
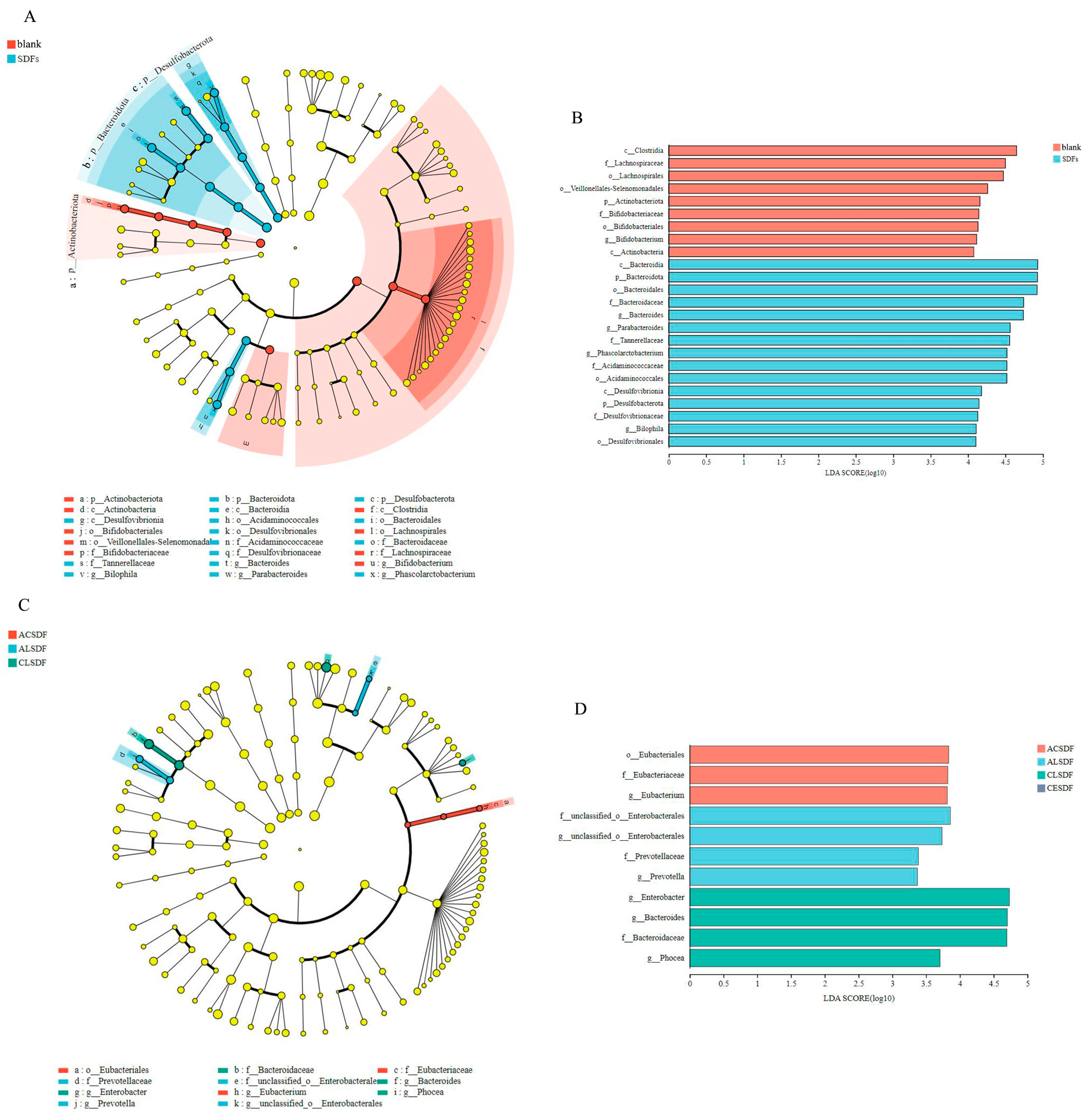
2.7. Effects of Bee Pollen SDFs In Vitro Fermentation on the Gut Microbiota
3. Materials and Methods
3.1. Materials
3.2. Extractions of Bee Pollen Soluble Dietary Fiber
3.2.1. Acid Extraction
3.2.2. Alkali Extraction
3.2.3. Cellulase Extraction
3.2.4. Complex Enzyme Extraction
3.3. Monosaccharide Composition Analysis
3.4. Molecular Weight Analysis
3.5. SEM
3.6. FT-IR
3.7. XRD
3.8. Phenolic Compounds’ Determination
3.8.1. Extraction of Phenolic Compounds
3.8.2. TPCs Determination
3.8.3. TFCs Determination
3.9. In Vitro Simulated Saliva-Gastrointestinal Digestion of Bee Pollen SDF
3.10. In Vitro Fermentation of Bee Pollen SDF
3.11. The SCFA Analysis
3.12. The Gut Microbiota Analysis
3.13. Statistic Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Makki, K.; Deehan, E.C.; Walter, J.; Bckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, S.; Beck, E.; Salman, H.; Tapsell, L. New horizons for the study of dietary fiber and health: A review. Plant Foods Hum. Nutr. 2016, 71, 1–12. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Flanagan, B.M.; Gidley, M.J. “Dietary fibre”: Moving beyond the “soluble/insoluble” classification for monogastric nutrition, with an emphasis on humans and pigs. J. Anim. Sci. Biotechnol. 2019, 10, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Wang, D.; Hu, R.; Long, Y.; Lv, L. Chemical composition, structural and functional properties of soluble dietary fiber obtained from coffee peel using different extraction methods. Food Res. Int. 2020, 136, 109497. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Ren, X.; Du, B.; Chen, L.; Yu, Z.; Yang, Y. Structure, Physicochemical Property, and Functional Activity of Dietary Fiber Obtained from Pear Fruit Pomace (Pyrus ussuriensis Maxim) via Different Extraction Methods. Foods 2022, 11, 2161. [Google Scholar] [CrossRef]
- Shen, M.; Wang, W.H.; Cao, L.K. Soluble dietary fibers from black soybean hulls: Physical and enzymatic modification, structure, physical properties, and cholesterol binding capacity. J. Food Sci. 2020, 85, 1668–1674. [Google Scholar] [CrossRef]
- Hua, M.; Sun, Y.; Shao, Z.; Lu, J.; Lu, Y.; Liu, Z. Functional soluble dietary fiber from ginseng residue: Polysaccharide characterization, structure, antioxidant, and enzyme inhibitory activity. J. Food Biochem. 2020, 44, e13524. [Google Scholar] [CrossRef]
- Ji, X.L.; Cheng, Y.Q.; Tian, J.Y.; Zhang, S.Q.; Jing, Y.S.; Shi, M.M. Structural characterization of polysaccharide from jujube (Ziziphus jujuba Mill.) fruit. Chem. Biol. Technol. Agric. 2021, 8, 54. [Google Scholar] [CrossRef]
- Jiang, G.; Ramachandraiah, K.; Wu, Z.; Ameer, K. The Inflfluence of Different Extraction Methods on the Structure, Rheological, Thermal and Functional Properties of Soluble Dietary Fiber from Sanchi (Panax notoginseng) Flower. Foods 2022, 11, 1995. [Google Scholar] [CrossRef]
- Katarína, F.Š.; Janka, N.; Magda, M.; Miroslava, K. Biologically active antimicrobial and antioxidant substances in the Helianthus annuus L. bee pollen. J. Environ. Sci. Health Part B 2016, 51, 176–181. [Google Scholar]
- Matuszewska, E.; Plewa, S.; Pietkiewicz, D.; Kossakowski, K.; Matysiak, J.; Rosiński, G.; Matysiak, J. Mass Spectrometry-Based Identifification of Bioactive Bee Pollen Proteins: Evaluation of Allergy Risk after Bee Pollen Supplementation. Molecules 2022, 27, 7733. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Wang, K.; Marcucci, M.C.; Sawaya, A.C.H.F.; Alexandra, C.H.; Hu, L.; Xue, X.F.; Wu, L.M.; Hu, F.L. Nutrient-rich bee pollen: A treasure trove of active natural metabolites. J. Funct. Foods 2018, 49, 472–484. [Google Scholar] [CrossRef]
- Qiao, J.T.; Feng, Z.X.; Zhang, Y.; Xiao, X.Y.; Dong, J.; Haubruge, E.; Zhang, H.C. Phenolamide and flavonoid glycoside profiles of 20 types of monofloral bee pollen. Food Chem. 2023, 405, 134800. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, X.P.; You, Z.D.; Yin, Z.Y.; Chen, Z.Z. Analysis and evaluation of fiber composition of four kinds of bee pollens. J. Nutr. 2017, 39, 513–517. [Google Scholar]
- Zou, X.Q.; Xu, X.L.; Chao, Z.H.; Jiang, X.; Zheng, L.; Jiang, B.Z. Properties of plant-derived soluble dietary fibers for fiber-enriched foods: A comparative evaluation. Int. J. Biol. Macromol. 2022, 223, 1196–1207. [Google Scholar] [CrossRef]
- Tao, Y.X.; Zhou, E.N.; Li, F.K.; Meng, L.F.; Li, Q.Q.; Wu, L.M. Allergenicity alleviation of bee pollen by enzymatic hydrolysis: Regulation in mice allergic mediators, metabolism, and gut microbiota. Foods 2022, 11, 3454. [Google Scholar] [CrossRef]
- Pan, L.H.; Wang, J.; Ye, X.Q.; Zha, X.Q.; Luo, J.P. Enzyme-assisted extraction of polysaccharides from Dendrobium chrysotoxum and its functional properties and immunomodulatory activity. LWT Food Sci. Technol. 2015, 60, 1149–1154. [Google Scholar] [CrossRef]
- Gan, J.; Huang, Z.; Yu, Q.; Peng, G.Y.; Chen, Y.; Xie, J.H.; Nie, S.P.; Xie, M.Y. Microwave assisted extraction with three modifications on structural and functional properties of soluble dietary fibers from grapefruit peel. Food Hydrocoll. 2019, 101, 105549. [Google Scholar] [CrossRef]
- Wang, S.O.; Fang, Y.Q.; Xu, Y.B.; Zhu, B.; Piao, J.G.; Zhu, L.L.; Yao, L.M.; Liu, K.H.; Wang, S.C.; Zhang, Q.Y.; et al. The effects of different extraction methods on physicochemical, functional and physiological properties of soluble and insoluble dietary fiber from Rubus chingii Hu. fruits. J. Funct. Foods 2022, 93, 105081. [Google Scholar] [CrossRef]
- Zheng, H.; Chen, X.P.; You, Z.D.; Xia, D.X.; Hu, X. Enzymatic extraction of bee pollen soluble dietary fiber and its physicochemical analysis. Food Mach. 2016, 32, 184–188. [Google Scholar]
- Wang, K.L.; Li, M.; Wang, Y.X.; Liu, Z.Z.; Ni, Y.Y. Effects of extraction methods on the structural characteristics and functional properties of dietary fiber extracted from kiwifruit (Actinidia deliciosa). Food Hydrocoll. 2021, 110, 106162. [Google Scholar] [CrossRef]
- Li, N.; Feng, Z.Q.; Niu, Y.G.; Yu, L.L. Structural, rheological and functional properties of modified soluble dietary fiber from tomato peels. Food Hydrocoll. 2017, 110, 106162. [Google Scholar] [CrossRef]
- Yang, C.R.; Si, J.Y.; Chen, Y.; Xie, J.H.; Tian, S.L.; Cheng, Y.N.; Hu, X.B.; Yu, Q. Physicochemical structure and functional properties of soluble dietary fibers obtained by different modification methods from Mesona chinensis Benth. residue. Food Res. Int. 2022, 157, 111489. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.L.; Guo, J.H.; Ding, D.Q.; Gao, J.; Hao, L.R.; Guo, X.D.; Liu, Y.Q. Structural characterization and antioxidant activity of a novel high-molecular-weight polysaccharide from Ziziphus Jujuba cv. Muzao. J. Food Meas. Charact. 2022, 16, 2191–2200. [Google Scholar] [CrossRef]
- Li, S.; Hu, N.N.; Zhu, J.Y.; Zheng, M.Z.; Liu, H.M.; Liu, J.S. Influence of modification methods on physicochemical and structural properties of soluble dietary fiber from corn bran. Food Chem. X 2022, 14, 100298. [Google Scholar] [CrossRef] [PubMed]
- Speroni, C.S.; Bender, A.B.B.; Stiebe, J.; Ballus, C.A.; Ávila, P.F.; Goldbeckb, R.; Morisso, F.D.P.; Silva, L.P.; Emanuelli, T. Granulometric fractionation and micronization: A process for increasing soluble dietary fiber content and improving technological and functional properties of olive pomace. LWT Food Sci. Technol. 2020, 130, 109526. [Google Scholar] [CrossRef]
- Ago, M.; Endo, T.; Hirotsu, T. Crystalline transformation of native cellulose from cellulose I to cellulose II polymorph by a ball-milling method with a specific amount of water. Cellulose 2004, 11, 163–167. [Google Scholar] [CrossRef]
- Jia, M.Y.; Chen, J.J.; Liu, X.Z.; Xie, M.Y.; Nie, S.P.; Chen, Y.; Xie, J.H.; Yu, Q. Structural characteristics and functional properties of soluble dietary fiber from defatted rice bran obtained through trichoderma viride fermentation. Food Hydrocoll. 2019, 94, 468–474. [Google Scholar] [CrossRef]
- Zhang, H.F.; Zhu, X.L.; Huang, Q.; Zhang, L.; Liu, X.H.; Liu, R.; Lu, Q. Antioxidant and anti-inflammatory activities of rape bee pollen after fermentation and their correlation with chemical components by ultra-performance liquid chromatography-quadrupole time of flight mass spectrometry-based untargeted metabolomics. Food Chem. 2023, 409, 135342. [Google Scholar] [CrossRef]
- Gercek, Y.C.; Celik, S.; Bayram, S. Screening of plant pollen sources, polyphenolic compounds, fatty acids and antioxidant/sntimicrobial sctivity from bee pollen. Molecules 2022, 27, 117. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Li, F.; Wu, W. Effect of rice bran rancidity on the structure and antioxidant properties of rice bran soluble dietary fiber. J. Cereal Sci. 2022, 105, 103469. [Google Scholar] [CrossRef]
- Hu, Q.D.; Ma, R.Y.; Tang, X.X.; Long, R.; Wang, Q.; Fu, C.M.; Li, R.; Xiao, H. Self-assembled nanoparticles of curcuminoids and soluble dietary fibers: Characterization and mechanism. LWT Food Sci. Technol. 2023, 173, 114375. [Google Scholar] [CrossRef]
- Wang, M.M.; Wichienchot, S.; He, X.W.; Fu, X.; Huang, Q.; Zhang, B. In vitro colonic fermentation of dietary fifibers: Fermentation rate, short-chain fatty acid production and changes in microbiota. Trends Food Sci. Technol. 2019, 88, 1–9. [Google Scholar] [CrossRef]
- Guo, Y.X.; Chen, X.F.; Gong, P.; Wang, M.R.; Yao, W.B.; Yang, W.J.; Chen, F.X. In vitro digestion and fecal fermentation of Siraitia grosvenorii polysaccharide and its impact on human gut microbiota. Food Funct. 2022, 13, 9443–9458. [Google Scholar] [CrossRef]
- Maruta, H.; Yamashita, H. Acetic acid stimulates G-protein-coupled receptor GPR43 and induces intracellular calcium influx in L6 myotube cells. PLoS ONE 2020, 15, 0239428. [Google Scholar] [CrossRef]
- Ma, J.L.; Liu, Z.K.; Gao, X.X.; Bao, Y.Y.; Hong, Y.; He, X.F.; Zhu, W.Z.; Li, Y.; Huang, W.J.; Zheng, N.N.; et al. Gut microbiota remodeling improves natural aging-related disorders through Akkermansia muciniphila and its derived acetic acid. Pharmacol. Res. 2022, 189, 106687. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.T.; Nie, X.R.; Gan, R.Y.; Guo, H.; Qin, W. In vitro digestion and fecal fermentation behaviors of a pectic polysaccharide from okra (Abelmoschus esculentus) and its impacts on human gut microbiota. Food Hydrocoll. 2021, 114, 106577. [Google Scholar] [CrossRef]
- Negatu, D.A.; Gengenbacher, M.; Dartois, V.; Dick, T. Indole propionic acid, an unusual antibiotic produced by the gut microbiota, with anti-inflammatory and antioxidant properties. Front. Microbiol. 2020, 11, 575586. [Google Scholar] [CrossRef]
- Duscha, A.; Gisevius, B.; Hirschberg, S.; Yissachar, N.; Stangl, G.I.; Eilers, E.; Bader, V.; Haase, S.; Kaisler, J.; David, C.; et al. Propionic acid shapes the multiple sclerosis disease course by an immunomodulatory mechanism. Cell 2020, 180, 1067–1080. [Google Scholar] [CrossRef]
- Onyszkiewicz, M.; Gawry-Kopczynska, M.; Konopelski, P.; Aleksandrowicz, M.; Sawicka, A.; Koźniewska, E.; Samborowska, E.; Ufnal, M. Butyric acid, a gut bacteria metabolite, lowers arterial blood pressure via colon-vagus nerve signaling and GPR41/43 receptors. Pflügers Arch. Eur. J. Physiol. 2019, 471, 1441–1453. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.M.; Huang, H.L.; Xu, J.; He, J.; Zhao, C.; Peng, Y.; Zhao, H.L.; Huang, W.Q.; Cao, Y.; Zhou, Y.J.; et al. Cross-talk between butyric acid and gut microbiota in ulcerative colitis following fecal microbiota transplantation. Front. Microbiol. 2021, 12, 658292. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Wan, P.; Chen, G.J.; Xie, M.H.; Sun, Y.; Zeng, X.X.; Liu, Z.H. Simulated digestion and fermentation in vitro by human gut microbiota of intra- and extra-cellular polysaccharides from Aspergillus cristatus. LWT Food Sci. Technol. 2019, 116, 108508. [Google Scholar] [CrossRef]
- Afrizal, L.; Hitch, T.C.A.; Viehof, A.; Treichel, N.; Riede, T.; Abt, B.; Buhl, E.M.; Kohlheyer, D.; Overmann, J.; Clavel, T. Anaerobic single-cell dispensing facilitates the cultivation of human gut bacteria. Environ. Microbiol. 2022, 24, 3861–3881. [Google Scholar] [CrossRef] [PubMed]
- Pascal, L.; Lombard, V.; Drula, E.; Terrapon, N.; Henrissat, B. Bacteroidetes use thousands of enzyme combinations to break down glycans. Nat. Commun. 2019, 10, 2043. [Google Scholar]
- Chang, C.J.; Lin, C.S.; Liu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Hu, B.; Liu, C.; Jiang, W.H.; Zhu, H.K.; Zhang, H.; Qian, H.; Zhang, W.G. Chronic in vitro fermentation and in vivo metabolism: Extracellular polysaccharides from sporidiobolus pararoseus regulate the intestinal microbiome of humans and mice. Int. J. Biol. Macromol. 2021, 192, 398–406. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, M.; Ning, X.X.; Qin, Y.L.; Wang, Y.W.; Yu, Z.X.; Dong, R.J.; Zhang, Y.M.; Sun, S.R. Expansion of Escherichia-Shigella in gut is associated with the onset and response to immunosuppressive therapy of IgA nephropathy. Clin. Res. 2022, 33, 2276–2292. [Google Scholar] [CrossRef]
- Johnson, E.L.; Heaver, S.L.; Walters, W.A.; Ley, R.E. Microbiome and metabolic disease: Revisiting the bacterial phylum bacteroidetes. J. Mol. Med. 2017, 95, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.L.; Zhang, L.S.; Wang, X.; Yi, Y.L.; Shan, Y.Y.; Liu, B.F.; Zhou, Y.; Lüa, X. Roles of intestinal Parabacteroides in human health an diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef]
- Li, L.; Su, Q.; Xie, B.; Duan, L.; Zhao, W.; Hu, D.; Wu, R.; Liu, H. Gut microbes in correlation with mood: Case study in a closed experimental human life support system. Neurogastroenterol. Motil. 2016, 28, 1233–1240. [Google Scholar] [CrossRef]
- Ikeyama, N.; Murakami, T.; Toyoda, A.; Mori, H.; Lion, T.; Ohkuma, M.; Sakamoto, M. Microbial interaction between the succinate-utilizing bacterium Phascolarctobacterium faecium and the gut commensal Bacteroides thetaiotaomicron. MicrobiologyOpen 2020, 9, e1111. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Arriaga, B.; Sandoval-Jiménez, S.; Rodríguez-Silverio, J.; Lizeth Alcaráz-Estrada, S.; Cortés-Espinosa, T.; Pérez-Cabeza de Vaca, R.; Licona-Cassani, C.; Gámez-Valdez, J.S.; Shaw, J.; Mondragón-Terán, P.; et al. Gut dysbiosis and clinical phases of pancolitis in patients with ulcerative colitis. MicrobiologyOpen 2012, 10, 1181. [Google Scholar] [CrossRef] [PubMed]
- Kivenson, V.; Giovannoni, S.J. An expanded genetic code enables trimethylamine metabolism in human gut bacteria. mSystems 2020, 5, e00413-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Mao, B.Y.; Gu, J.Y.; Wu, J.Y.; Cui, S.M.; Wang, G.; Zhao, J.X.; Zhang, H.; Chen, W. Blautia—A new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- He, B.L.; Xiong, Y.; Hu, T.G.; Zong, M.H.; Wu, H. Bifidobacterium spp. as functional foods: A review of current status, challenges, and strategies. Crit. Rev. Food Sci. Nutr. 2022, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Lucie, E.M.; Adeline, S.; Pascale, M.; Courtin, C.M.; Tom, V.W.; Stéphanie, B.D. Tripartite relationship between gut microbiota, intestinal mucus and dietary fibers: Towards preventive strategies against enteric infections. FEMS Microbiol. Rev. 2020, 45, fuaa052. [Google Scholar]
- Gu, M.; Fang, H.; Gao, Y.; Su, T.; Niu, Y.; Yu, L.L. Characterization of enzymatic modified soluble dietary fiber from tomato peels with high release of lycopene. Food Hydrocoll. 2019, 99, 105321. [Google Scholar] [CrossRef]
- Ma, Y.N.; Wang, Z.; Arifeen, M.Z.U.; Xue, Y.; Yuan, S.; Liu, C.H. Structure and bioactivity of polysaccharide from a subseafloor strain of schizophyllum commune 20R-7-F01. Int. J. Biol. Macromol. 2022, 222, 610–619. [Google Scholar] [CrossRef]
- Lin, P.; Chen, L.; Huang, X.; Xiao, F.; Fu, L.; Jing, D.; Wang, J.; Zhang, H.; Sun, L.; Wu, Y. Structural characteristics of polysaccharide GP2a in Gardenia jasminoides and Its Immunomodulatory Effect on Macrophages. Int. J. Mol. Sci. 2022, 23, 11279. [Google Scholar] [CrossRef] [PubMed]
- Li, H.M.; Zheng, T.; Zeng, Y.Q.; Fan, Y.B.; Li, W.Z.; Yan, J.Y.; Zhao, L.P.; Zheng, H.; Yang, Y. Comprehensive optimization of modified technology of polygonatum odoratum whole powder by AHP-CRITIC weighted evaluation. Food Ferment Ind. 2021, 47, 138–146. [Google Scholar]
- Zheng, H.; Sun, Y.; Zheng, T.; Zeng, Y.; Fu, L.; Zhou, T.; Jia, F.; Xu, Y.; He, K.; Yang, Y. Effects of shear emulsifying/ball milling/autoclave modifification on structure, physicochemical properties, phenolic compounds, and antioxidant capacity of lotus (Nelumbo) leaves dietary fifiber. Front. Nutr. 2023, 10, 1064662. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Wu, Y.; Wang, L.; Tan, B.; Shen, W.Y.; Li, X.N.; Liu, Y.X.; Tian, X.H.; Zhang, D.Q. Comparison of six modification methods on the chemical composition, functional properties and antioxidant capacity of wheat bran. LWT Food Sci. Technol. 2021, 149, 111996. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Amorim, C.; Silvério, S.C.; Cardoso, B.B.; Alves, J.I.; Pereira, M.A.; Rodrigues, L.R. In vitro fermentation of raffinose to unravel its potential as prebiotic ingredient. LWT Food Sci. Technol. 2020, 126, 109322. [Google Scholar] [CrossRef]
- Liu, Y.T.; Li, Y.W.; Ke, Y.; Li, C.; Zhang, Z.Q.; Wu, Y.L.; Hu, B.; Liu, A.; Luo, Q.Y.; Wu, W.J. In vitro saliva-gastrointestinal digestion and fecal fermentation of Oudemansiella radicata polysaccharides reveal its digestion profile and effect on the modulation of the gut microbiota. Carbohydr. Polym. 2021, 251, 117041. [Google Scholar] [CrossRef]
- Yang, F.; Yang, J.; Ruan, Z.; Wang, Z. Fermentation of dietary fibers modified by an enzymatic-ultrasonic treatment and evaluation of their impact on gut microbiota in mice. J. Food Process. Preserv. 2021, 45, e15337. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | ACSDF | ALSDF | CLSDF | CESDF |
|---|---|---|---|---|
| Extraction yield (%) | 3.61 | 3.98 | 6.01 | 5.05 |
| Monosaccharide composition (molar ratio %) | ||||
| Fucose | 1.20 | 1.42 | 0.71 | 0.46 |
| Arabinose | 33.83 | 40.99 | 33.06 | 29.98 |
| Rhamnose | 0.82 | 3.56 | 0.69 | 3.45 |
| Galactose | 16.35 | 20.07 | 17.44 | 15.08 |
| Glucose | 32.16 | 13.53 | 22.57 | 17.79 |
| Xylose | 6.49 | 13.75 | 2.38 | 8.37 |
| Mannose | 6.90 | 4.83 | 20.48 | 19.00 |
| Ribose | ND | ND | ND | 3.47 |
| Glucuronic acid | 2.25 | 1.85 | 2.66 | 2.33 |
| Molecular weight | ||||
| Mw (kDa) | 77.17 | 56.50 | 143.51 | 1050.27 |
| Mn (kDa) | 20.91 | 15.27 | 32.23 | 18.75 |
| D (Mw/Mn) | 3.69 | 3.70 | 4.45 | 56.02 |
| SCFAs (mmol/L) | Blank | ACSDF | ALSDF | CLSDF | CESDF |
|---|---|---|---|---|---|
| Acetic acid | 3.81 ± 0.73 d | 6.00 ± 0.47 b | 5.57 ± 0.22 bc | 5.92 ± 0.55 b | 9.24 ± 1.79 a |
| Propionic acid | 0.48 ± 0.06 b | 1.12 ± 0.18 a | 1.18 ± 0.16 a | 1.07 ± 0.15 a | 0.90 ± 0.15 a |
| Butyric acid | 0.81 ± 0.07 d | 1.37 ± 0.05 c | 1.56 ± 0.17 c | 1.80 ± 0.05 b | 2.36 ± 0.10 a |
| Total SCFAs | 5.10 ± 0.68 c | 8.49 ± 0.32 b | 8.31 ± 0.20 b | 8.79 ± 0.44 b | 12.50 ± 1.76 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, H.; Sun, Y.; Zeng, Y.; Zheng, T.; Jia, F.; Xu, P.; Xu, Y.; Cao, Y.; He, K.; Yang, Y. Effects of Four Extraction Methods on Structure and In Vitro Fermentation Characteristics of Soluble Dietary Fiber from Rape Bee Pollen. Molecules 2023, 28, 4800. https://doi.org/10.3390/molecules28124800
Zheng H, Sun Y, Zeng Y, Zheng T, Jia F, Xu P, Xu Y, Cao Y, He K, Yang Y. Effects of Four Extraction Methods on Structure and In Vitro Fermentation Characteristics of Soluble Dietary Fiber from Rape Bee Pollen. Molecules. 2023; 28(12):4800. https://doi.org/10.3390/molecules28124800
Chicago/Turabian StyleZheng, Hui, Yan Sun, Yiqiong Zeng, Tao Zheng, Fan Jia, Pan Xu, Yao Xu, Yuxin Cao, Kai He, and Yong Yang. 2023. "Effects of Four Extraction Methods on Structure and In Vitro Fermentation Characteristics of Soluble Dietary Fiber from Rape Bee Pollen" Molecules 28, no. 12: 4800. https://doi.org/10.3390/molecules28124800