
Estrogen Receptor β Isoforms Regulate Chemotherapy Resistance and the Cancer Stem Cell Population in Prostate Cancer Cells

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Cell Culture
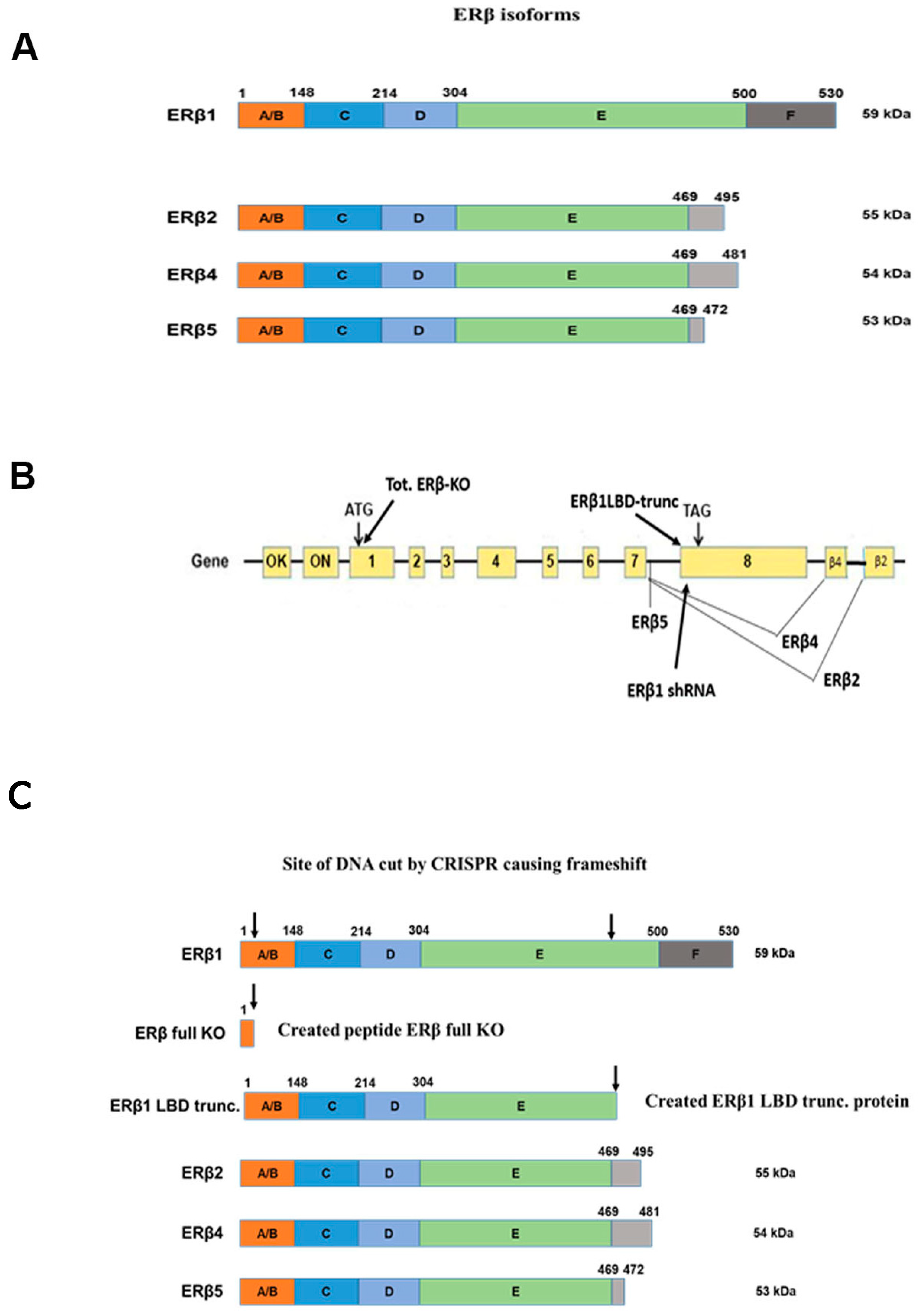
2.2. DU145, PC3, and LNCaP Cell Lines Manipulated Using CRISPR/Cas9 to Cause Either Full ERβ Knockout or LBD Truncation of Endogenous ERβ1
2.3. Chemotherapy Treatment
2.4. Protein Extract Preparation
2.5. Western Blotting
2.6. Immunocytochemistry
2.7. RNA Extraction and Real-Time PCR
2.8. CRISPR Manipulation of Cell Lines
2.9. Tumorosphere Assay
2.10. Statistics
3. Results
3.1. Prostate Cancer Cell Lines Express All ERβ Isoforms
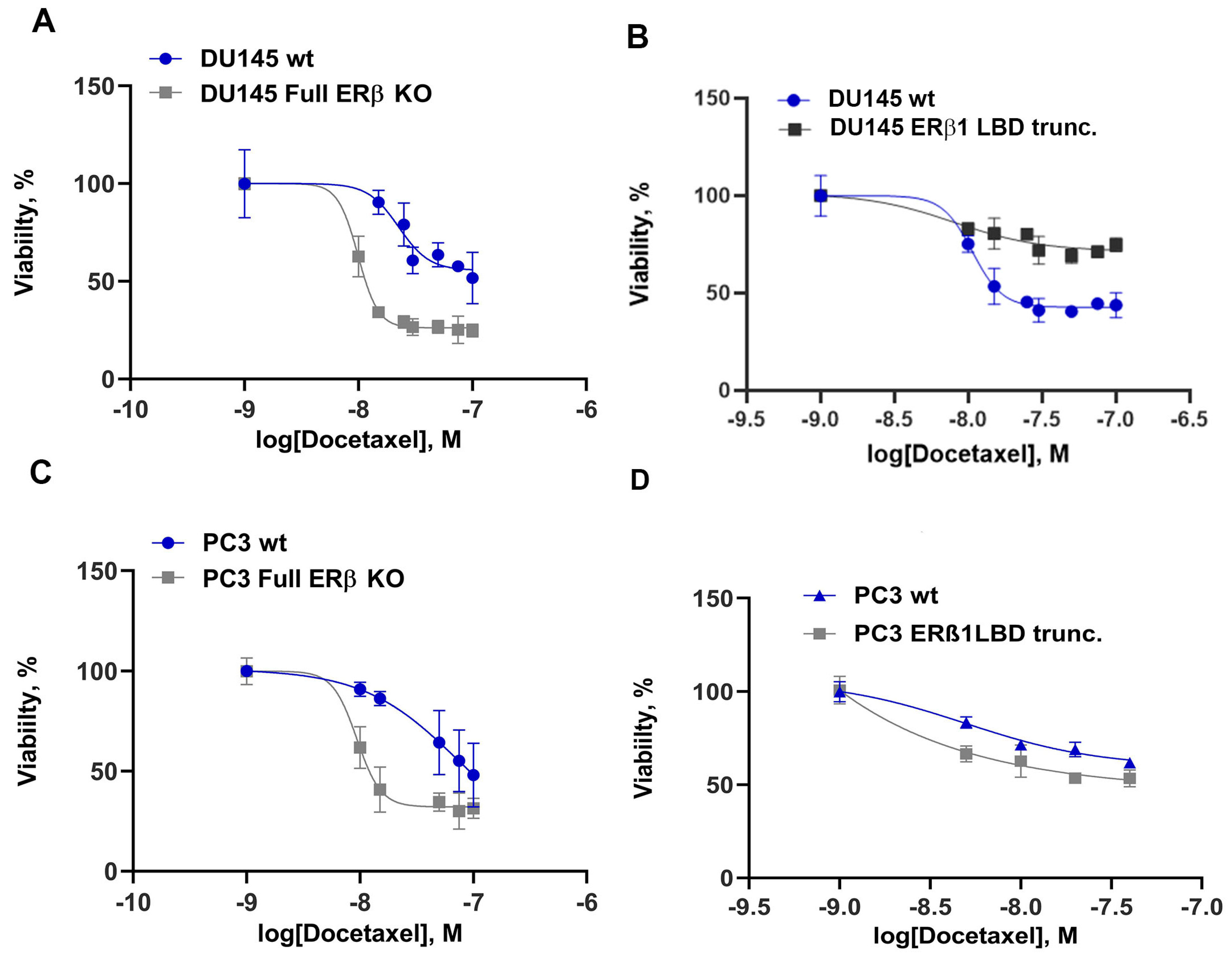
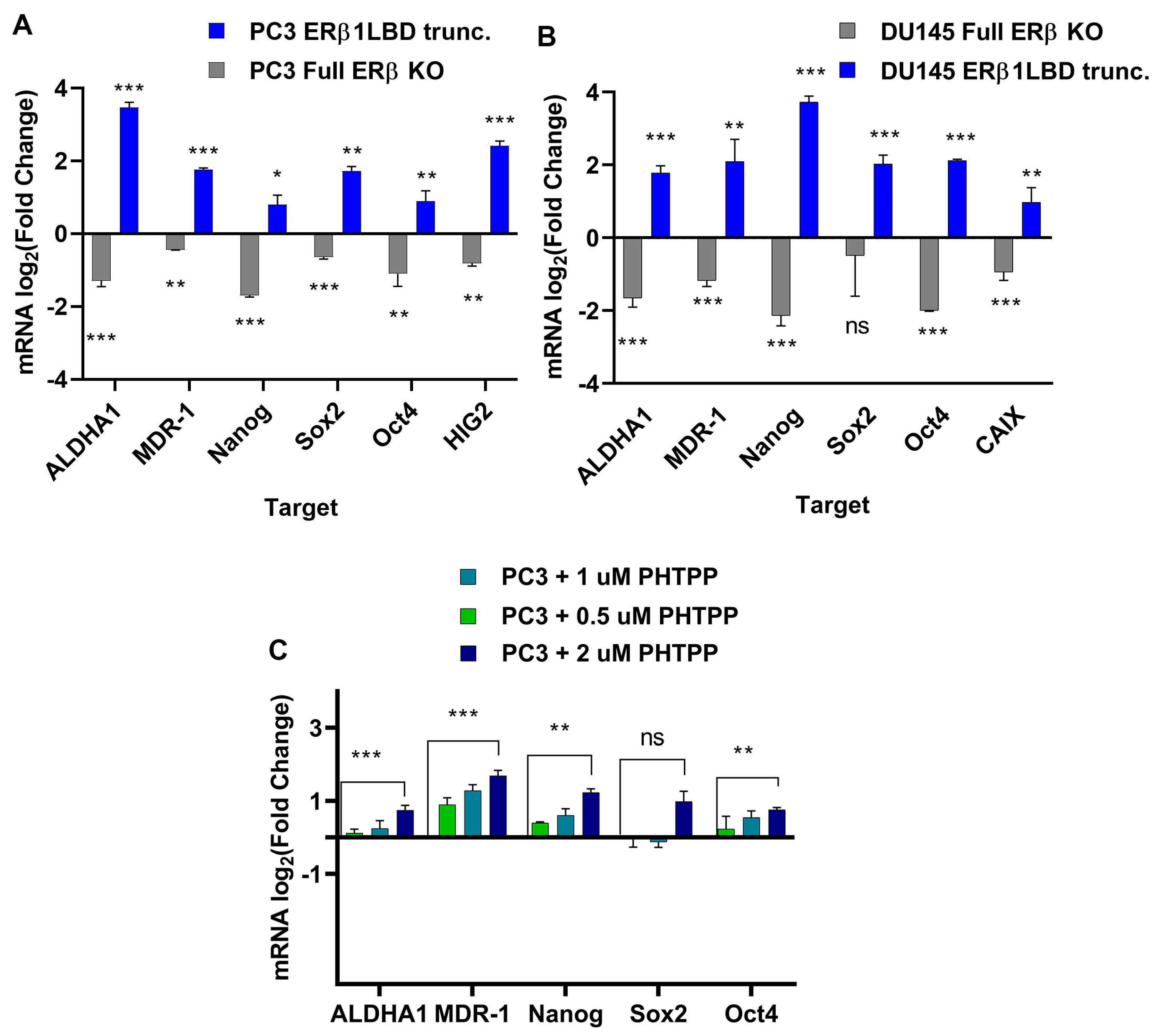
3.2. Endogenously Expressed ERβ Isoforms in Prostate Cancer Cell Lines Affect Chemotherapy Resistance and Stem Cell Factor Expression
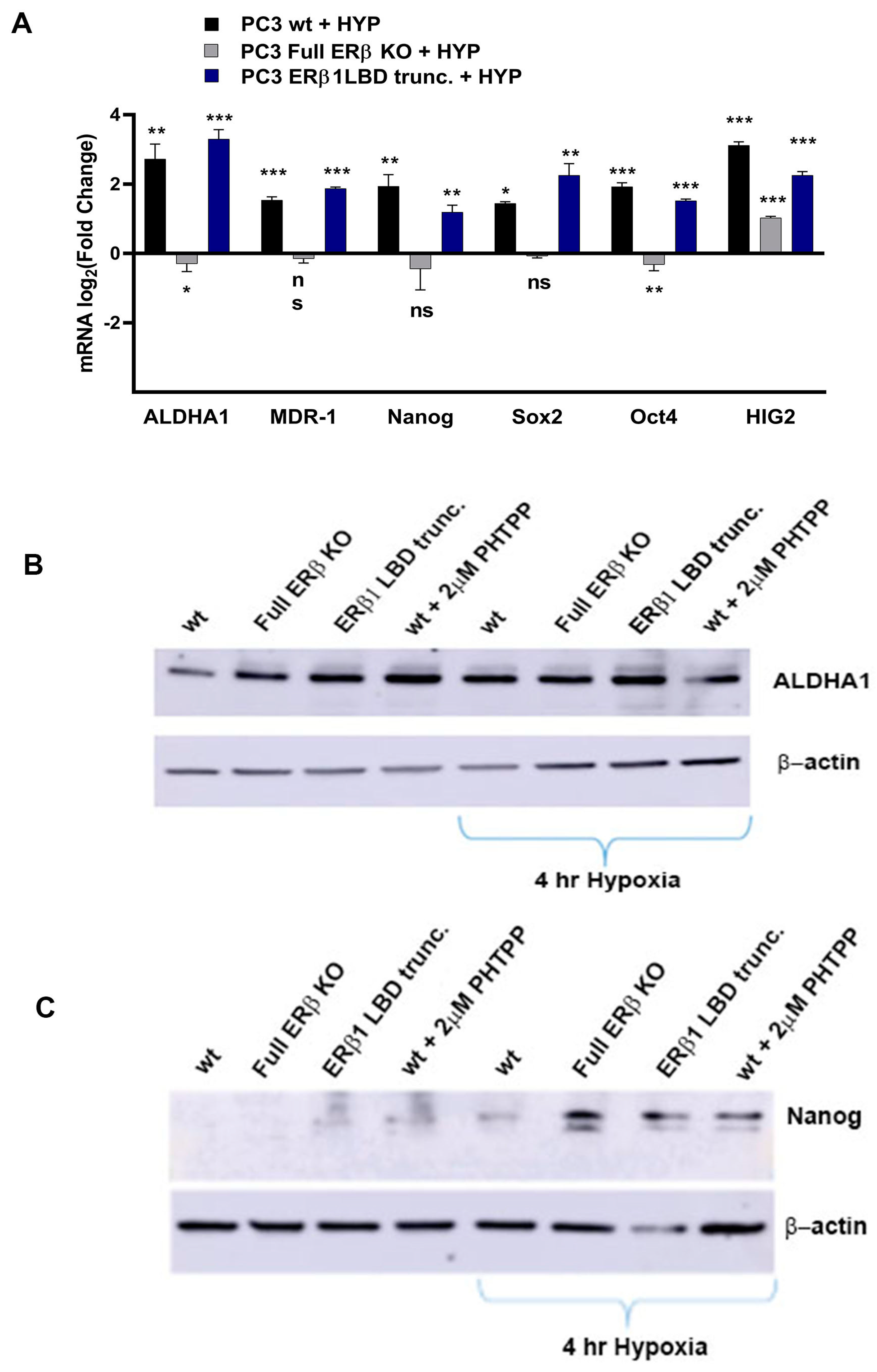
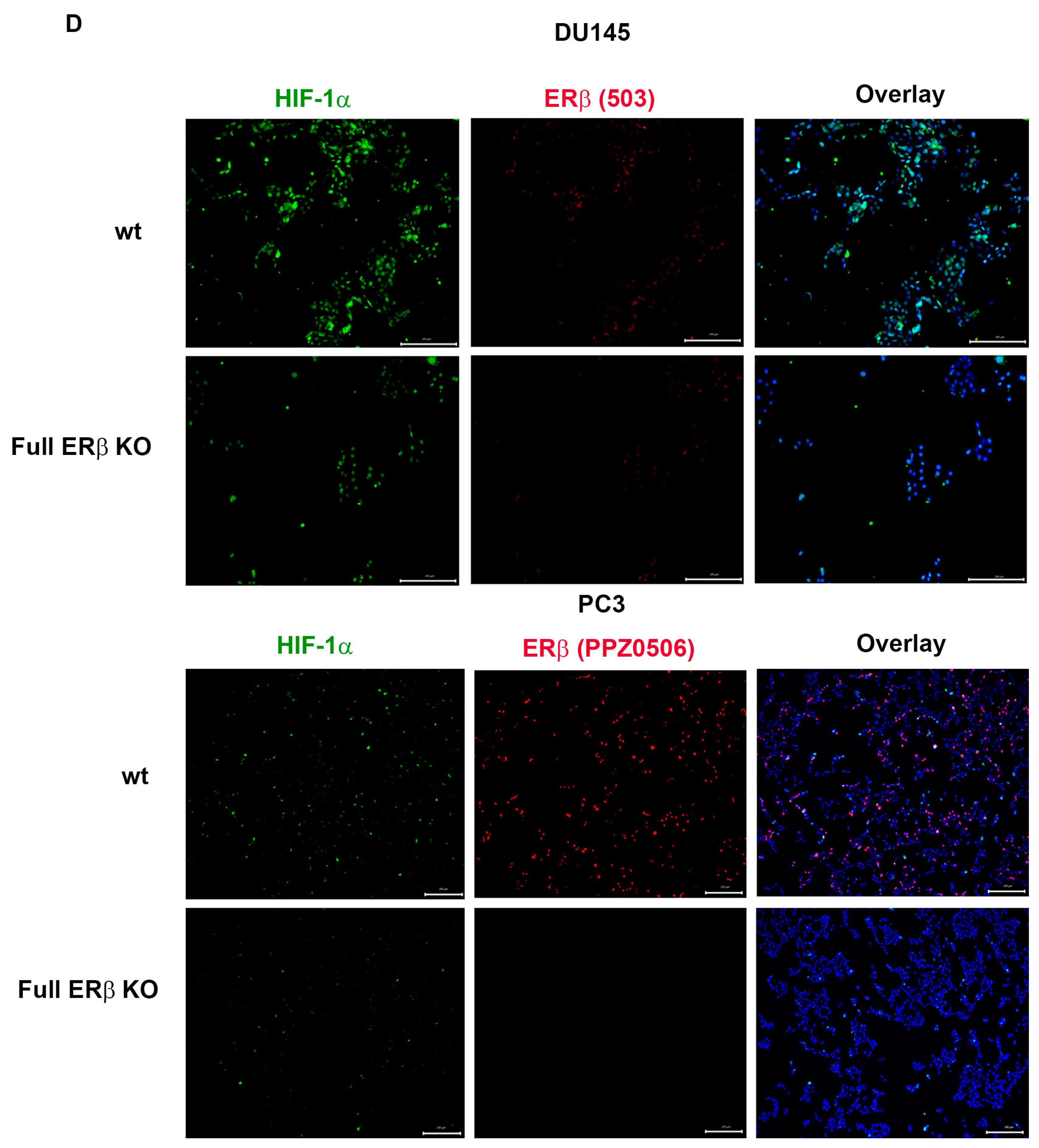
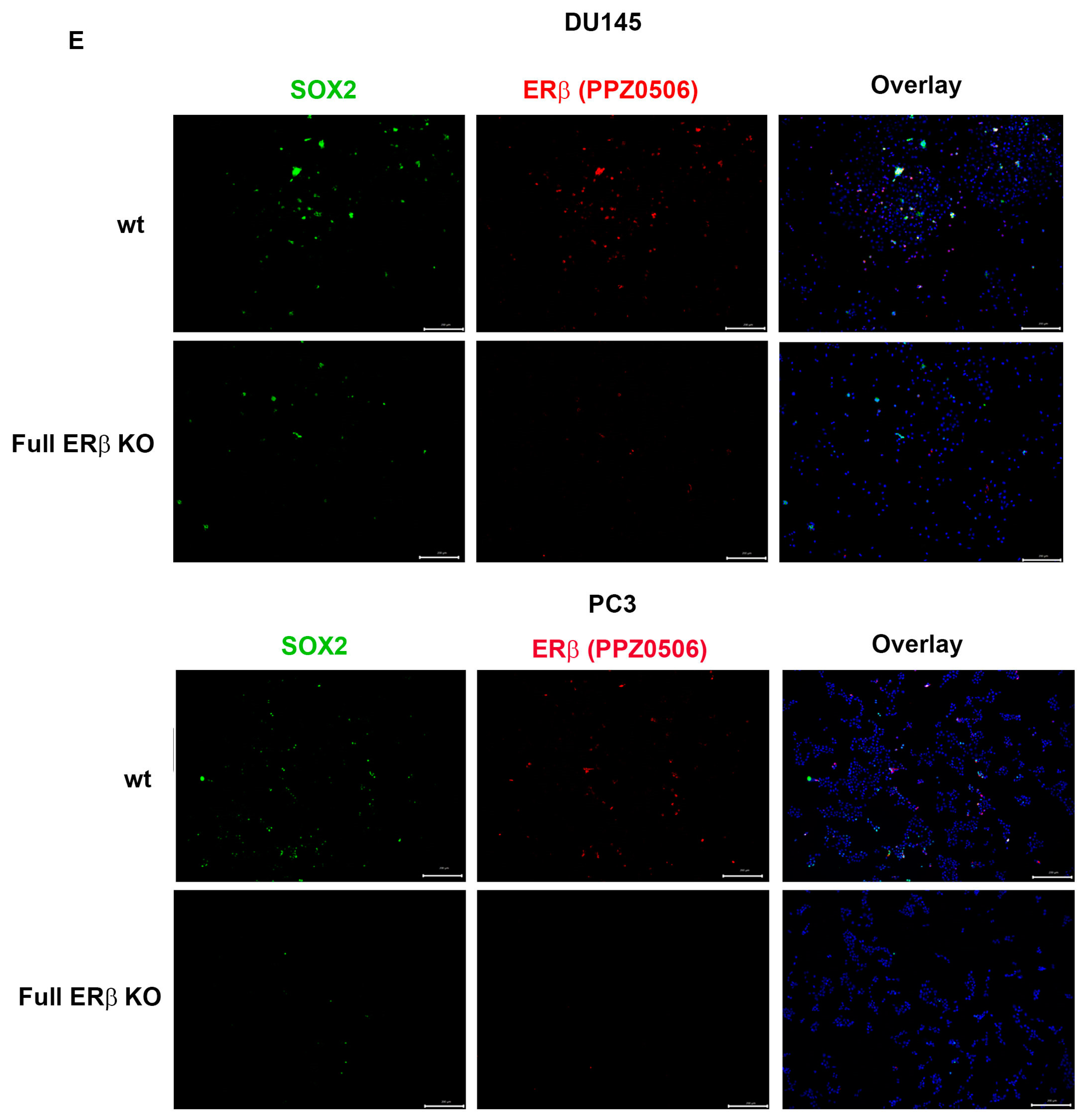
3.3. PC3 Cells Exposed to Hypoxia Show a Similar Gene Expression Profile to PC3 Cells with CRISPR/Cas9. Truncated ERβ1 LBD and the Full ERβ KO Attenuates the Increased Expression of CSC Markers in Hypoxia
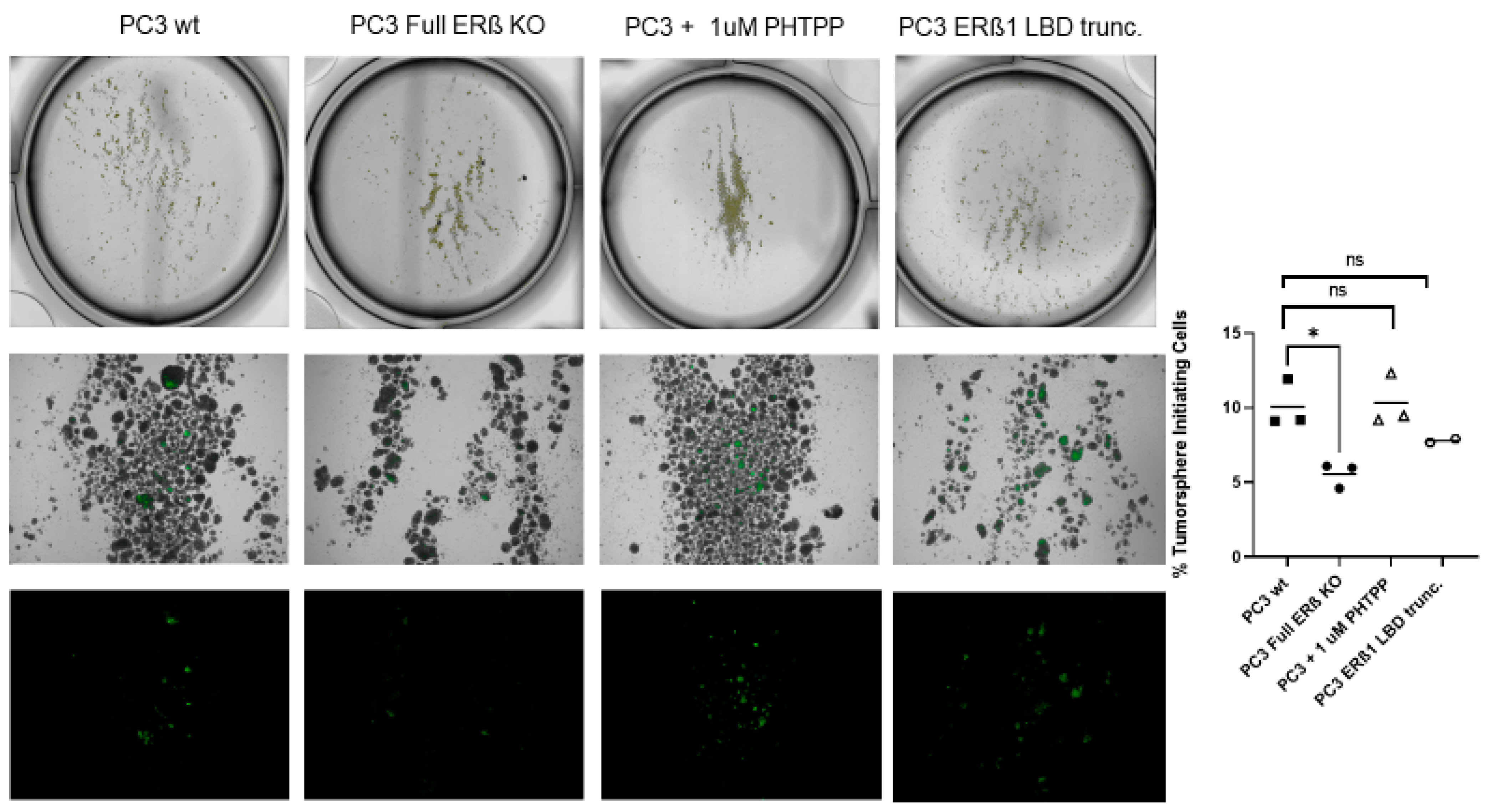
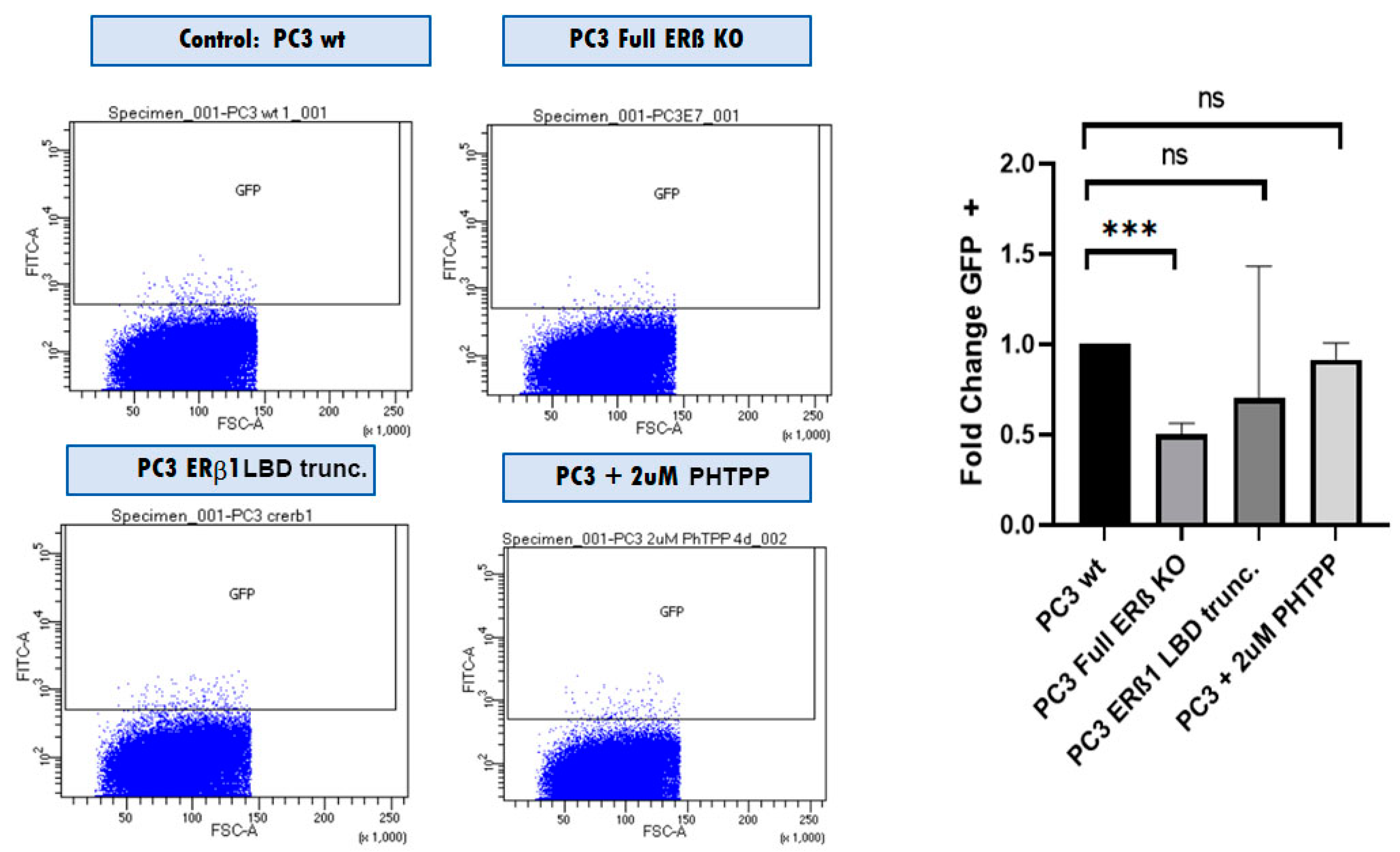
3.4. The Full ERβ Knockout Reduces the CSC Population
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuiper, G.G.; Enmark, E.; Pelto-Huikko, M.; Nilsson, S.; Gustafsson, J.A. Cloning of a novel receptor expressed in rat prostate and ovary. Proc. Natl. Acad. Sci. USA 1996, 93, 5925–5930. [Google Scholar] [CrossRef] [PubMed]
- Fujimura, T.; Takahashi, S.; Urano, T.; Ogawa, S.; Ouchi, Y.; Kitamura, T.; Muramatsu, M.; Inoue, S. Differential expression of estrogen receptor beta (ERbeta) and its C-terminal truncated splice variant ERbetacx as prognostic predictors in human prostatic cancer. Biochem. Biophys. Res. Commun. 2001, 289, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Leung, Y.K.; Mak, P.; Hassan, S.; Ho, S.M. Estrogen receptor (ER)-beta isoforms: A key to understanding ER-beta signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 13162–13167. [Google Scholar] [CrossRef]
- Dey, P.; Jonsson, P.; Hartman, J.; Williams, C.; Strom, A.; Gustafsson, J.A. Estrogen receptors beta1 and beta2 have opposing roles in regulating proliferation and bone metastasis genes in the prostate cancer cell line PC3. Mol. Endocrinol. 2012, 26, 1991–2003. [Google Scholar] [CrossRef]
- Leung, Y.K.; Lam, H.M.; Wu, S.; Song, D.; Levin, L.; Cheng, L.; Wu, C.L.; Ho, S.M. Estrogen receptor beta2 and beta5 are associated with poor prognosis in prostate cancer, and promote cancer cell migration and invasion. Endocr. Relat. Cancer 2010, 17, 675–689. [Google Scholar] [CrossRef]
- Macedo-Silva, C.; Benedetti, R.; Ciardiello, F.; Cappabianca, S.; Jeronimo, C.; Altucci, L. Epigenetic mechanisms underlying prostate cancer radioresistance. Clin. Epigenetics 2021, 13, 125. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, N.; Dahut, W.L.; Steinberg, S.M.; Figg, W.D.; Tarassoff, C.; Arlen, P.; Gulley, J.L. A retrospective study of the time to clinical endpoints for advanced prostate cancer. BJU Int. 2005, 96, 985–989. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Davies, A.H.; Zoubeidi, A. The Androgen Receptor Bridges Stem Cell-Associated Signaling Nodes in Prostate Stem Cells. Stem Cells Int. 2016, 2016, 4829602. [Google Scholar] [CrossRef] [Green Version]
- Jeter, C.R.; Liu, B.; Liu, X.; Chen, X.; Liu, C.; Calhoun-Davis, T.; Repass, J.; Zaehres, H.; Shen, J.J.; Tang, D.G. NANOG promotes cancer stem cell characteristics and prostate cancer resistance to androgen deprivation. Oncogene 2011, 30, 3833–3845. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, B.G.; Bort, A.; Vara-Ciruelos, D.; Diaz-Laviada, I. Androgen Deprivation Induces Reprogramming of Prostate Cancer Cells to Stem-Like Cells. Cells 2020, 9, 1441. [Google Scholar] [CrossRef]
- Shen, M.M.; Abate-Shen, C. Molecular genetics of prostate cancer: New prospects for old challenges. Genes Dev. 2010, 24, 1967–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaurasiya, S.; Widmann, S.; Botero, C.; Lin, C.Y.; Gustafsson, J.A.; Strom, A.M. Estrogen receptor beta exerts tumor suppressive effects in prostate cancer through repression of androgen receptor activity. PLoS ONE 2020, 15, e0226057. [Google Scholar] [CrossRef]
- Wu, W.F.; Maneix, L.; Insunza, J.; Nalvarte, I.; Antonson, P.; Kere, J.; Yu, N.Y.; Tohonen, V.; Katayama, S.; Einarsdottir, E.; et al. Estrogen receptor beta, a regulator of androgen receptor signaling in the mouse ventral prostate. Proc. Natl. Acad. Sci. USA 2017, 114, E3816–E3822. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.F.; Wang, L.; Spetsieris, N.; Boukovala, M.; Efstathiou, E.; Brossner, C.; Warner, M.; Gustafsson, J.A. Estrogen receptor beta and treatment with a phytoestrogen are associated with inhibition of nuclear translocation of EGFR in the prostate. Proc. Natl. Acad. Sci. USA 2021, 118, e2011269118. [Google Scholar] [CrossRef]
- Dey, P.; Velazquez-Villegas, L.A.; Faria, M.; Turner, A.; Jonsson, P.; Webb, P.; Williams, C.; Gustafsson, J.A.; Strom, A.M. Estrogen Receptor beta2 Induces Hypoxia Signature of Gene Expression by Stabilizing HIF-1alpha in Prostate Cancer. PLoS ONE 2015, 10, e0128239. [Google Scholar] [CrossRef]
- Faria, M.; Shepherd, P.; Pan, Y.; Chatterjee, S.S.; Navone, N.; Gustafsson, J.A.; Strom, A. The estrogen receptor variants beta2 and beta5 induce stem cell characteristics and chemotherapy resistance in prostate cancer through activation of hypoxic signaling. Oncotarget 2018, 9, 36273–36288. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Huang, Z.; Chen, C.Z.; Liu, C.; Evans, C.P.; Gao, A.C.; Zhou, F.; Chen, H.W. Therapeutic Targeting of MDR1 Expression by RORgamma Antagonists Resensitizes Cross-Resistant CRPC to Taxane via Coordinated Induction of Cell Death Programs. Mol. Cancer Ther. 2020, 19, 364–374. [Google Scholar] [CrossRef]
- Comerford, K.M.; Wallace, T.J.; Karhausen, J.; Louis, N.A.; Montalto, M.C.; Colgan, S.P. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 2002, 62, 3387–3394. [Google Scholar] [PubMed]
- Zhou, J.J.; Deng, X.G.; He, X.Y.; Zhou, Y.; Yu, M.; Gao, W.C.; Zeng, B.; Zhou, Q.B.; Li, Z.H.; Chen, R.F. Knockdown of NANOG enhances chemosensitivity of liver cancer cells to doxorubicin by reducing MDR1 expression. Int. J. Oncol. 2014, 44, 2034–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butterworth, K.T.; McCarthy, H.O.; Devlin, A.; Ming, L.; Robson, T.; McKeown, S.R.; Worthington, J. Hypoxia selects for androgen independent LNCaP cells with a more malignant geno- and phenotype. Int. J. Cancer 2008, 123, 760–768. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kim, H.; Pollack, S. ERbeta Isoforms Have Differential Clinical Significance in Breast Cancer Subtypes and Subgroups. Curr. Issues Mol. Biol. 2022, 44, 1564–1586. [Google Scholar] [CrossRef]
- Fraga, A.; Ribeiro, R.; Principe, P.; Lopes, C.; Medeiros, R. Hypoxia and Prostate Cancer Aggressiveness: A Tale With Many Endings. Clin. Genitourin. Cancer 2015, 13, 295–301. [Google Scholar] [CrossRef]
- Keith, B.; Simon, M.C. Hypoxia-inducible factors, stem cells, and cancer. Cell 2007, 129, 465–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitani, T.; Yamaji, R.; Higashimura, Y.; Harada, N.; Nakano, Y.; Inui, H. Hypoxia enhances transcriptional activity of androgen receptor through hypoxia-inducible factor-1alpha in a low androgen environment. J. Steroid Biochem. Mol. Biol. 2011, 123, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Ragnum, H.B.; Roe, K.; Holm, R.; Vlatkovic, L.; Nesland, J.M.; Aarnes, E.K.; Ree, A.H.; Flatmark, K.; Seierstad, T.; Lilleby, W.; et al. Hypoxia-independent downregulation of hypoxia-inducible factor 1 targets by androgen deprivation therapy in prostate cancer. Int. J. Radiat. Oncol. Biol. Phys. 2013, 87, 753–760. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.; Takahashi, K.; Okita, K.; Ichisaka, T.; Yamanaka, S. Hypoxia enhances the generation of induced pluripotent stem cells. Cell Stem Cell 2009, 5, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Christoforou, P.; Christopoulos, P.F.; Koutsilieris, M. The role of estrogen receptor beta in prostate cancer. Mol. Med. 2014, 20, 427–434. [Google Scholar] [CrossRef]
- Mak, P.; Chang, C.; Pursell, B.; Mercurio, A.M. Estrogen receptor beta sustains epithelial differentiation by regulating prolyl hydroxylase 2 transcription. Proc. Natl. Acad. Sci. USA 2013, 110, 4708–4713. [Google Scholar] [CrossRef]
- Mak, P.; Leav, I.; Pursell, B.; Bae, D.; Yang, X.; Taglienti, C.A.; Gouvin, L.M.; Sharma, V.M.; Mercurio, A.M. ERbeta impedes prostate cancer EMT by destabilizing HIF-1alpha and inhibiting VEGF-mediated snail nuclear localization: Implications for Gleason grading. Cancer Cell 2010, 17, 319–332. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Karagounis, I.V.; Srivastava, R.K.; Vrettos, N.; Nikolos, F.; Francois, N.; Huang, M.; Gong, S.; Long, Q.; Kumar, S.; et al. Estrogen Receptor beta-Mediated Inhibition of Actin-Based Cell Migration Suppresses Metastasis of Inflammatory Breast Cancer. Cancer Res. 2021, 81, 2399–2414. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Cheng, L.; Gupta, K.; Chiang, H.C.; Gupta, H.B.; Sareddy, G.R.; Wang, D.; Lathrop, K.; Elledge, R.; Wang, P.; et al. Tyrosine phosphorylation regulates ERbeta ubiquitination, protein turnover, and inhibition of breast cancer. Oncotarget 2016, 7, 42585–42597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stossi, F.; Barnett, D.H.; Frasor, J.; Komm, B.; Lyttle, C.R.; Katzenellenbogen, B.S. Transcriptional profiling of estrogen-regulated gene expression via estrogen receptor (ER) alpha or ERbeta in human osteosarcoma cells: Distinct and common target genes for these receptors. Endocrinology 2004, 145, 3473–3486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bano, A.; Stevens, J.H.; Modi, P.S.; Gustafsson, J.A.; Strom, A.M. Estrogen Receptor beta4 Regulates Chemotherapy Resistance and Induces Cancer Stem Cells in Triple Negative Breast Cancer. Int. J. Mol. Sci. 2023, 24, 5867. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Ajani, J.A.; Song, S. Drug resistance and Cancer stem cells. Cell Commun. Signal. 2021, 19, 19. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | FWD | REV |
|---|---|---|
| GAPDH | 5′-TGACAACTTTGGTATCGTGGAGG-3′ | 5′-AGGCAGGGATGTTCTGGAGAG-3′ |
| 36B4 | 5′-GTGTTCGACAATGGCAGCAT-3′ | 5′-GACACCCTCCAGGAAGCGA-3′ |
| ALDHA1 | 5′-GCTGGCGACAATGG AGTCAA-3′ | 5′-ACGGCCCTGGATCTTGTCAG-3′ |
| OCT4 | 5′-GGATGCTGTGA GCCAAGG-3′ | 5′-GAACAATGATGAGTGACAGACAG-3′ |
| CAIX | 5′-AGATGAGAAGGCAGCACAGAA-3′ | 5′-GAAGTGGCATAATGAGCAGGA-3′ |
| SOX2 | 5′-TTGCTGCCTCTTTAAGACTAGGA-3′ | 5′-CTGGGGCTCAAAC TTCTCTC-3′ |
| Nanog | 5′-ACCCAGCTGTGTGTACTCAA-3′ | 5′-GGAAGAGTAAAGGCTGGGGT-3′ |
| HIG2 | 5′-CCGACTTTCCTCCGGACT-3′ | 5′-CCTTCTGAAAGGCC TCTGG-3′ |
| MDR1 | 5′-AGGAAGCCAATGCCTATGACTTTA-3′ | 5′-CAACTGGGCCCCT CTCTCTC-3′ |
| ERβ isoform specific primers | ||
| ERβ1 | 5′-GTCAGGCATGCGAGTAACAA-3′ | 5′-GGGAGCCCTCTTTGCTTTTA-3′ |
| ERβ2/4 | 5′-ACTTGCTGAACGCCGTGACC-3′ | |
| ERβ2 | 5′-AGAGGCCTTTTCTG CCCTCGC-3′ | |
| ERβ4 | 5′-TTTTCTCCCCATCTCGCATGC-3′ | |
| ERβ5 | 5′-GACCACAGGTCCTGTGTATACAAGTAA-3′ | 5′-GGCACTAAGATAACTTCAAATGAATGAATT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stevens, J.H.; Bano, A.; Bensaoula, L.; Strom, A.M.; Gustafsson, J.-Å. Estrogen Receptor β Isoforms Regulate Chemotherapy Resistance and the Cancer Stem Cell Population in Prostate Cancer Cells. Receptors 2023, 2, 176-190. https://doi.org/10.3390/receptors2030012
Stevens JH, Bano A, Bensaoula L, Strom AM, Gustafsson J-Å. Estrogen Receptor β Isoforms Regulate Chemotherapy Resistance and the Cancer Stem Cell Population in Prostate Cancer Cells. Receptors. 2023; 2(3):176-190. https://doi.org/10.3390/receptors2030012
Chicago/Turabian StyleStevens, Jessica H., Ayesha Bano, Lamia Bensaoula, Anders M. Strom, and Jan-Åke Gustafsson. 2023. "Estrogen Receptor β Isoforms Regulate Chemotherapy Resistance and the Cancer Stem Cell Population in Prostate Cancer Cells" Receptors 2, no. 3: 176-190. https://doi.org/10.3390/receptors2030012