
Clinical and Therapeutic Implications of BCAAs Metabolism during Chronic Liver Disease in Humans: Crosstalk between Skeletal Muscle and Liver
 , , , and
, , , and
Abstract
:1. Introduction
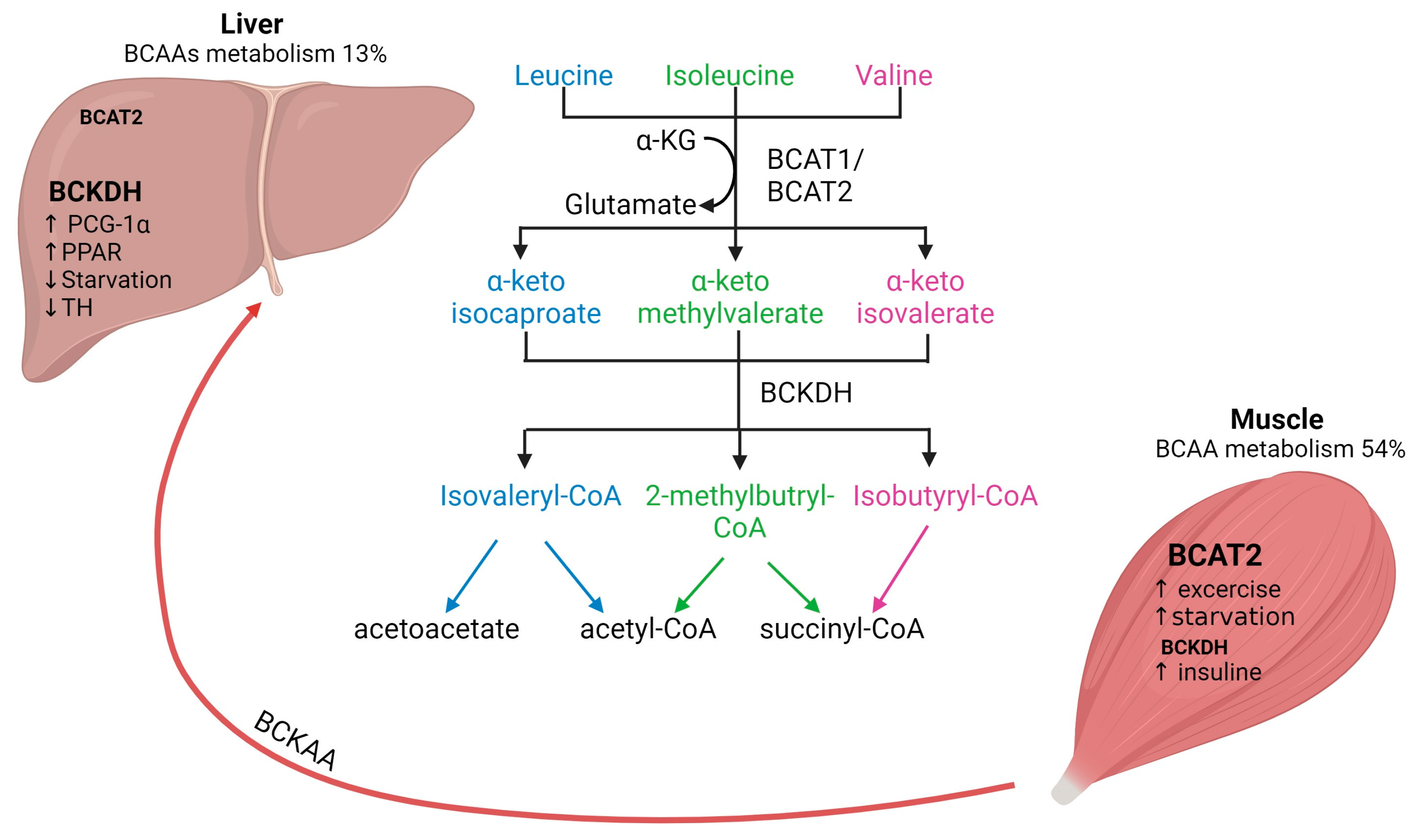
2. Metabolism of BCAAs: Interplay between Skeletal Muscle and Liver
2.1. BCAAs Transamination
2.2. BCAAs Oxidation
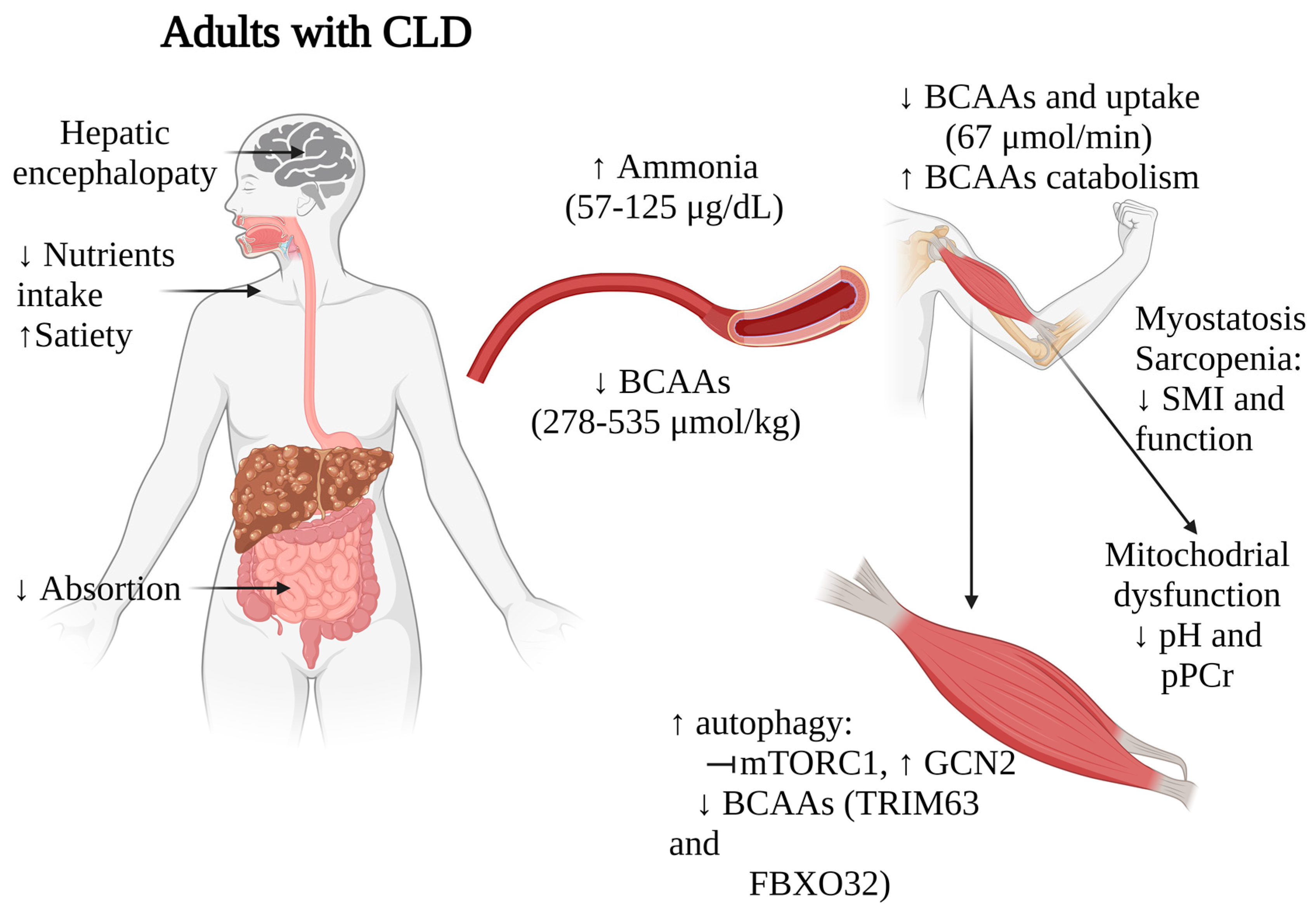
3. Clinical Insights into CLD: Role of BCAAs in Musculoskeletal Health
3.1. BCAAs Concentration in Patients with CLD
3.2. Correlation of Muscle Health and BCAAs in Patients with CLD
3.3. Interplay of BCAAs, mTOR Signaling, Ammonia, and Mitochondrial Dysfunction in the Skeletal Muscle during CLD
4. Interventions with BCAAs Supplementation in Patients with CLD: Implications for the Skeletal Muscle
4.1. Effects of BCAAs on Skeletal Muscle Mass and Skeletal Muscle Strength

{kind=link}
{kind=link}
| Reference | Type of Study | Intervention | Treatment Composition | Inclusion Criteria | Sample Size | Treatment Duration | Findings |
|---|---|---|---|---|---|---|---|
| [35] | Open-label prospective study. | Oral supplementation with BCAAs (4.5 g/day) as a late evening snack, compared to a combination of BCAAs supplementation with unsupervised walking exercise, or unsupervised walking exercise alone. | BCAAs 4.5 g, containing: Leu 3 g Ile 0.75 g Val 0.75 g | CLD with CP score A or B | 127 patients: BCAAs group, n = 42 Walking exercise, n = 43 Walking exercise plus BCAAs, n = 42 | 3 months. | -Walking exercise plus BCAAs significantly increased grip strength and SMI, reducing sarcopenia prevalence. -Albumin significantly increased in all groups. |
| [68] | RCT. | Oral supplementation with BCAAs (10.85 g/day) against regular diet. | BCAAs 5.425 g, containing: Leu 2.03 g Ile 1.76 g Val 1.635 g | CLD in frail patients with CP score A or B | 54 patients (27 in the BCAAs group). | 4 months. | -Increased SMI in the intervention group. -Improvement in all 4 domains of the physical component score of the SF-36 questionnaire in the BCAAs group. -Improvement in the LFI, BMI and serum albumin compared to controls. |
| [69] | RCT. | Oral supplementation with BCAAs (12 g/day) against placebo, in adition to a home-based exercise program (30 min/day), dietary counselling and standard medical therapy. | CLD with CP score A or B and sarcopenia | 60 patients (30 in the BCAAs group). | 6 months. | No changes in muscle mass with the addition of BCAAs to exercise, dietary counselling and standard medical therapy. | |
| [66] | RCT. | Oral supplementation with BCAAs (7.2 g/dose) against lactoalbumin (6.3 g/dose). | BCAAs 8.1 g, containing: Leu 1.2 g Ile 0.6 g Val 0.6 g | CLD, with portal hypertension and sarcopenia | 138 patients (69 in the BCAAs group) | 6 months. | -Improvement of the hand grip strength. -Improvement of the gait speed score from before treatment with BCAAs to after treatment. -Increased in the TAMA, TSF, and MAMC. -Lower progression of HE. |
| [70] | RCT. | Oral supplementation with BCAAs (5.24 g/day) and physical activity against nutritional and physical activity intervention. | Each 100-g sachet contained: Leu 2.61 g Ile 1.01 g Val 1.62 g | CLD and sarcopenia | 32 patients (15 in the BCAAs group) | 3 months. | -Improvement in muscle mass and serum albumin levels compared to controls. -Improvement in the LFI in the global cohort. |
| [36] | Cohort study. | Oral supplementation with BCAAs (4 g/dose) three times daily. | BCAAs 4 g, containing: Leu 1.9 g Ile 0.9 g Val 1.14 g | CLD with serum albumin levels of ≤3.5 g/dL | 21 patients. | 12 months. | -Decreased IMAC in patients with ameliorated hypoalbuminemia associated with BCAAs supplementation. -Higher liver-related event-free survival rates in patients with decreased IMAC. |
| [65] | RCT. | Oral supplementation with BCAAs (110 g/day) against high-fiber and high-protein diet. | BCAAs 110 g, containing: Leu 3.38 g Ile 2.75 g Val 2.5 g | CLD and CP score A or B | 72 patients (73 in the BCAAs group) | 6 months. | -Increased muscle mass and decreased fat mass compared to control. -No differences in ammonia and glucose levels. -No significant changes in the Psychometric HE Score or the Critical Flicker–Frequency score results. |
| [37] | RCT. | Single oral BCAAs mixture (15 g) enriched with Leu, and a primed constant (0.05 µmol.kg−1.min−1) infusion of L-[ring-2H5]-phenylalanine. | BCAAs 15 g, containing: Leu 7.5 g Ile 3.75 Val 3.75 g | CLD due to ALD, with CP score A, and alcohol abstinence for at least 6 months | 6 patients. 8 controls. | 7 h. | -BCAAs/Leu acutely reversed impaired mTOR1 signaling and increased autophagy in skeletal muscle of cirrhotic ALD patients. -Plasma BCAAs increased similarly in cirrhotic patients and controls after oral intake. -Muscle synthesis rate was slightly higher in controls. -BCAAs/Leu supplementation reduced elevated protein breakdown in cirrhosis patients. |
| [67] | RCT. | Oral supplementation with BCAAs (30 g) or maltodextrin twice a day. | BCAAs 30 g, containing: Leu 13.5 g Ile 9 g Val 7.5 g | CLD and a previous episode of HE | 116 patients (58 in the BCAAs group). | 14 months. | -Increased MAMC in the BCAAs group. -No difference in the risk of remaining free of HE among groups. |
4.2. Role of BCAAs in Frailty and Nutritional Status
4.3. Role of BCAAs on Skeletal Muscle Protein Metabolism
4.4. Combining BCAAs with Exercise
5. Clinical Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALD | alcohol-associated liver disease. |
| BCAAs | Branched-chain Amino Acids. |
| BCAT | branched-chain aminotransferase. |
| BCKAs | branched-chain α-ketoacids. |
| BCKDH | branched-chain α-keto acid dehydrogenase. |
| CFS | Clinical Frailty Scale. |
| CLD | Chronic Liver Disease. |
| HBV | hepatitis B virus. |
| HBV | hepatitis C virus. |
| HCC | hepatocellular carcinoma. |
| HPLC | high-performance liquid chromatography. |
| Ile | isoleucine. |
| IMAC | intramuscular adipose tissue content. |
| Leu | leucine. |
| LFI | liver frailty index. |
| MAMC | mid-arm muscle circumference. |
| MASLD | Metabolic Dysfunction-Associated Steatotic Liver Disease. |
| MELD | Model for End-Stage Liver Disease. |
| mTOR | mammalian target of rapamycin. |
| NMR | Proton (1H) nuclear magnetic resonance. |
| PT | prothrombin time. |
| RCT | randomized controlled trial. |
| SMI | skeletal muscle mass index. |
| TAMA | total abdominal muscle area. |
| TCA | citric acid cycle. |
| TSF | triceps skinfold. |
| Val | valine. |
References
- Neinast, M.; Murashige, D.; Arany, Z. Branched Chain Amino Acids. Annu. Rev. Physiol. 2019, 81, 139–164. [Google Scholar] [CrossRef]
- Mann, G.; Mora, S.; Madu, G.; Adegoke, O.A.J. Branched-chain Amino Acids: Catabolism in Skeletal Muscle and Implications for Muscle and Whole-Body Metabolism. Front. Physiol. 2021, 12, 702826. [Google Scholar] [CrossRef]
- Nie, C.; He, T.; Zhang, W.; Zhang, G.; Ma, X. Branched Chain Amino Acids: Beyond Nutrition Metabolism. Int. J. Mol. Sci. 2018, 19, 954. [Google Scholar] [CrossRef]
- Lynch, C.J.; Adams, S.H. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat. Rev. Endocrinol. 2014, 10, 723–736. [Google Scholar] [CrossRef]
- Freund, H.R.; Hanani, M. The metabolic role of branched-chain amino acids. Nutrition 2002, 18, 287–288. [Google Scholar] [CrossRef]
- Kimball, S.R.; Jefferson, L.S. Regulation of protein synthesis by branched-chain amino acids. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 39–43. [Google Scholar] [CrossRef]
- Holeček, M. Branched-chain amino acids in health and disease: Metabolism, alterations in blood plasma, and as supplements. Nutr. Metab. 2018, 15, 33. [Google Scholar] [CrossRef]
- Holeček, M. The role of skeletal muscle in the pathogenesis of altered concentrations of branched-chain amino acids (valine, leucine, and isoleucine) in liver cirrhosis, diabetes, and other diseases. Physiol. Res. 2021, 70, 293–305. [Google Scholar] [CrossRef]
- Iwai, S.; Hasegawa, T.; Ikeda, H.O.; Tsujikawa, A. Branched Chain Amino Acids Promote ATP Production via Translocation of Glucose Transporters. Investig. Ophthalmol. Vis. Sci. 2022, 63, 7. [Google Scholar] [CrossRef]
- Wolfe, R.R. Branched-chain amino acids and muscle protein synthesis in humans: Myth or reality? J. Int. Soc. Sports Nutr. 2017, 14, 30. [Google Scholar] [CrossRef]
- Kimball, S.R.; Jefferson, L.S. Signaling pathways and molecular mechanisms through which branched-chain amino acids mediate translational control of protein synthesis. J. Nutr. 2006, 136 (Suppl. S1), 227S–231S. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A Branched-Chain Amino Acid-Related Metabolic Signature that Differentiates Obese and Lean Humans and Contributes to Insulin Resistance. Cell Metab. 2009, 9, 565–566. [Google Scholar] [CrossRef]
- Nishi, K.; Yoshii, A.; Abell, L.; Zhou, B.; Frausto, R.; Ritterhoff, J.; McMillen, T.S.; Sweet, I.; Wang, Y.; Gao, C.; et al. Branched-chain keto acids inhibit mitochondrial pyruvate carrier and suppress gluconeogenesis in hepatocytes. Cell Rep. 2023, 42, 112641. [Google Scholar] [CrossRef]
- Li, T.; Zhang, Z.; Kolwicz, S.C., Jr.; Abell, L.; Roe, N.D.; Kim, M.; Zhou, B.; Cao, Y.; Ritterhoff, J.; Gu, H.; et al. Defective Branched-Chain Amino Acid Catabolism Disrupts Glucose Metabolism and Sensitizes the Heart to Ischemia-Reperfusion Injury. Cell Metab. 2017, 25, 374–385. [Google Scholar] [CrossRef]
- Wu, T.; Wang, M.; Ning, F.; Zhou, S.; Hu, X.; Xin, H.; Reilly, S.; Zhang, X. Emerging role for branched-chain amino acids metabolism in fibrosis. Pharmacol. Res. 2023, 187, 106604. [Google Scholar] [CrossRef]
- Lo, E.K.K.; Felicianna Xu, J.H.; Zhan, Q.; Zeng, Z.; El-Nezami, H. The Emerging Role of Branched-Chain Amino Acids in Liver Diseases. Biomedicines 2022, 10, 1444. [Google Scholar] [CrossRef]
- Dimou, A.; Tsimihodimos, V.; Bairaktari, E. The Critical Role of the Branched Chain Amino Acids (BCAAs) Catabolism-Regulating Enzymes, Branched-Chain Aminotransferase (BCAT) and Branched-Chain α-Keto Acid Dehydrogenase (BCKD), in Human Pathophysiology. Int. J. Mol. Sci. 2022, 23, 4022. [Google Scholar] [CrossRef]
- Tajiri, K.; Shimizu, Y. Branched-chain amino acids in liver diseases. Transl. Gastroenterol. Hepatol. 2018, 3, 47. [Google Scholar] [CrossRef]
- Kinny-Köster, B.; Bartels, M.; Becker, S.; Scholz, M.; Thiery, J.; Ceglarek, U.; Kaiser, T. Plasma Amino Acid Concentrations Predict Mortality in Patients with End-Stage Liver Disease. PLoS ONE 2016, 11, e0159205. [Google Scholar] [CrossRef]
- Nong, X.; Zhang, C.; Wang, J.; Ding, P.; Ji, G.; Wu, T. The mechanism of branched-chain amino acid transferases in different diseases: Research progress and future prospects. Front. Oncol. 2022, 12, 988290. [Google Scholar] [CrossRef]
- She, P.; Reid, T.M.; Bronson, S.K.; Vary, T.C.; Hajnal, A.; Lynch, C.J.; Hutson, S.M. Disruption of BCATm in mice leads to increased energy expenditure associated with the activation of a futile protein turnover cycle. Cell Metab. 2007, 6, 181–194. [Google Scholar] [CrossRef]
- Roberson, P.A.; Haun, C.T.; Mobley, C.B.; Romero, M.A.; Mumford, P.W.; Martin, J.S.; Roberts, M.D. Skeletal muscle amino acid transporter and BCAT2 expression prior to and following interval running or resistance exercise in mode-specific trained males. Amino Acids 2018, 50, 961–965. [Google Scholar] [CrossRef]
- Paulusma, C.C.; Lamers, W.H.; Broer, S.; van de Graaf, S.F.J. Amino acid metabolism, transport and signalling in the liver revisited. Biochem. Pharmacol. 2022, 201, 115074. [Google Scholar] [CrossRef]
- Vanweert, F.; Neinast, M.; Tapia, E.E.; van de Weijer, T.; Hoeks, J.; Schrauwen-Hinderling, V.B.; Blair, M.C.; Bornstein, M.R.; Hesselink, M.K.C.; Schrauwen, P.; et al. A randomized placebo-controlled clinical trial for pharmacological activation of BCAA catabolism in patients with type 2 diabetes. Nat. Commun. 2022, 13, 3508. [Google Scholar] [CrossRef]
- White, P.J.; Lapworth, A.L.; McGarrah, R.W.; Kwee, L.C.; Crown, S.B.; Ilkayeva, O.; An, J.; Carson, M.W.; Christopher, B.A.; Ball, J.R.; et al. Muscle-Liver Trafficking of BCAA-Derived Nitrogen Underlies Obesity-Related Glycine Depletion. Cell Rep. 2020, 33, 108375. [Google Scholar] [CrossRef]
- Monirujjaman, M.d.; Ferdouse, A. Metabolic and Physiological Roles of Branched-Chain Amino Acids. Adv. Mol. Biol. 2014, 2014, 1–6. [Google Scholar] [CrossRef]
- Lu, G.; Sun, H.; She, P.; Youn, J.Y.; Warburton, S.; Ping, P.; Vondriska, T.M.; Cai, H.; Lynch, C.J.; Wang, Y. Protein phosphatase 2Cm is a critical regulator of branched-chain amino acid catabolism in mice and cultured cells. J. Clin. Investig. 2009, 119, 1678–1687. [Google Scholar] [CrossRef]
- Zhou, M.; Shao, J.; Wu, C.Y.; Shu, L.; Dong, W.; Liu, Y.; Chen, M.; Wynn, R.M.; Wang, J.; Wang, J.; et al. Targeting BCAA catabolism to treat obesity-associated insulin resistance. Diabetes 2019, 68, 1730–1746. [Google Scholar] [CrossRef]
- Ye, Z.; Wang, S.; Zhang, C.; Zhao, Y. Coordinated Modulation of Energy Metabolism and Inflammation by Branched-Chain Amino Acids and Fatty Acids. Front. Endocrinol. 2020, 11, 617. [Google Scholar] [CrossRef]
- Huisman, E.J.; Trip, E.J.; Siersema, P.D.; Van Hoek, B.; Van Erpecum, K.J. Protein energy malnutrition predicts complications in liver cirrhosis. Eur. J. Gastroenterol. Hepatol. 2011, 23, 982–989. [Google Scholar] [CrossRef]
- Pentiuk, N.; Motsiuk, V. Evaluation of nutritional status in patients with liver cirrhosis. Validity and prognostic value of the Patient-Generated Subjective Global Assessment. Prz. Gastroenterol. 2023, 18, 327–333. [Google Scholar] [CrossRef]
- Montanari, A.; Simoni, I.; Vallisa, D.; Trifirò, A.; Colla, R.; Abbiati, R.; Borghi, L.; Novarini, A. Free amino acids in plasma and skeletal muscle of patients with liver cirrhosis. Hepatology 1988, 8, 1034–1039. [Google Scholar] [CrossRef]
- Dam, G.; Keiding, S.; Munk, O.L.; Ott, P.; Buhl, M.; Vilstrup, H.; Bak, L.K.; Waagepetersen, H.S.; Schousboe, A.; Møller, N.; et al. Branched-chain amino acids increase arterial blood ammonia in spite of enhanced intrinsic muscle ammonia metabolism in patients with cirrhosis and healthy subjects. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G269–G277. [Google Scholar] [CrossRef]
- Dam, G.; Sørensen, M.; Buhl, M.; Sandahl, T.D.; Møller, N.; Ott, P.; Vilstrup, H. Muscle metabolism and whole blood amino acid profile in patients with liver disease. Scand. J. Clin. Lab. Investig. 2015, 75, 674–680. [Google Scholar]
- Xiang, Q.; Xiong, J.; Zhao, Z.J.; Zhou, T.; Wu, J.; Chen, X. Walking exercise through smartphone application plus branched-chain amino acid supplementation benefits skeletal muscle mass and strength in liver cirrhosis: A prospective control trial. Z. Gastroenterol. 2023. [Google Scholar] [CrossRef]
- Kitajima, Y.; Takahashi, H.; Akiyama, T.; Murayama, K.; Iwane, S.; Kuwashiro, T.; Tanaka, K.; Kawazoe, S.; Ono, N.; Eguchi, T.; et al. Supplementation with branched-chain amino acids ameliorates hypoalbuminemia, prevents sarcopenia, and reduces fat accumulation in the skeletal muscles of patients with liver cirrhosis. J. Gastroenterol. 2018, 53, 427–437. [Google Scholar] [CrossRef]
- Tsien, C.; Davuluri, G.; Singh, D.; Allawy, A.; Ten Have, G.A.; Thapaliya, S.; Schulze, J.M.; Barnes, D.; McCullough, A.J.; Engelen, M.P.; et al. Metabolic and molecular responses to leucine-enriched branched chain amino acid supplementation in the skeletal muscle of alcoholic cirrhosis. Hepatology 2015, 61, 2018–2029. [Google Scholar] [CrossRef]
- Trillos-Almanza, M.C.; Wessel, H.; Martínez-Aguilar, M.; van den Berg, E.H.; Douwes, R.M.; Moshage, H.; Connelly, M.A.; Bakker, S.J.L.; de Meijer, V.E.; Dullaart, R.P.F.; et al. Branched Chain Amino Acids Are Associated with Physical Performance in Patients with End-Stage Liver Disease. Biomolecules 2023, 13, 824. [Google Scholar] [CrossRef]
- Doi, J.; Shiraishi, K.; Haida, M.; Matsuzaki, S. Abnormality of energy metabolism in the skeletal muscle of patients with liver cirrhosis and changes under administration of glucose and branched-chain amino acids. Tokai J. Exp. Clin. Med. 2004, 29, 191–198. Available online: https://europepmc.org/article/MED/15717491 (accessed on 13 January 2024).
- Janota, B.; Krupowicz, A.; Noras, K.; Janczewska, E. Evaluation of the nutritional status of patients with liver cirrhosis. World J. Hepatol. 2023, 15, 914–924. [Google Scholar] [CrossRef]
- Sehgal, P.; Sharma, S.; Sood, A.; Dharni, K.; Kakkar, C.; Batta, S.; Sahotra, M. Assessment and prediction of malnutrition and sarcopenia in liver cirrhosis patients. Nutrire 2023, 48, 6. [Google Scholar] [CrossRef]
- Dasarathy, S.; Merli, M. Sarcopenia from mechanism to diagnosis and treatment in liver disease. J. Hepatol. 2016, 65, 1232–1244. [Google Scholar] [CrossRef]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A.; et al. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Kim, G.; Kang, S.H.; Kim, M.Y.; Baik, S.K. Prognostic value of sarcopenia in patients with liver cirrhosis: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0186990. [Google Scholar] [CrossRef]
- Yuan, S.; Larsson, S.C. Epidemiology of sarcopenia: Prevalence, risk factors, and consequences. Metab. Clin. Exp. 2023, 144, 155533. [Google Scholar] [CrossRef]
- Pahor, M.; Manini, T.; Cesari, M. Sarcopenia: Clinical evaluation, biological markers and other evaluation tools. J. Nutr. Health Aging 2009, 13, 724–728. [Google Scholar] [CrossRef]
- Gangitano, E.; Gnessi, L.; Merli, M. Protein Catabolism and the Dysregulation of Energy Intake-Related Hormones May Play a Major Role in the Worsening of Malnutrition in Hospitalized Cirrhotic Patients. Livers 2022, 2, 158–170. [Google Scholar] [CrossRef]
- Nardelli, S.; Lattanzi, B.; Merli, M.; Farcomeni, A.; Gioia, S.; Ridola, L.; Riggio, O. Muscle Alterations Are Associated with Minimal and Overt Hepatic Encephalopathy in Patients with Liver Cirrhosis. Hepatology 2019, 70, 1704–1713. [Google Scholar] [CrossRef]
- Kumar, A.; Welch, N.; Mishra, S.; Bellar, A.; Silva, R.N.; Li, L.; Singh, S.S.; Sharkoff, M.; Kerr, A.; Chelluboyina, A.K.; et al. Metabolic reprogramming during hyperammonemia targets mitochondrial function and postmitotic senescence. JCI Insight 2021, 6, e154089. [Google Scholar] [CrossRef]
- Mishra, S.; Welch, N.; Karthikeyan, M.; Bellar, A.; Musich, R.; Singh, S.S.; Zhang, D.; Sekar, J.; Attaway, A.H.; Chelluboyina, A.K.; et al. Dysregulated cellular redox status during hyperammonemia causes mitochondrial dysfunction and senescence by inhibiting sirtuin-mediated deacetylation. Aging Cell 2023, 22, e13852. [Google Scholar] [CrossRef]
- Davuluri, G.; Allawy, A.; Thapaliya, S.; Rennison, J.H.; Singh, D.; Kumar, A.; Sandlers, Y.; van Wagoner, D.R.; Flask, C.A.; Hoppel, C.; et al. Hyperammonaemia-induced skeletal muscle mitochondrial dysfunction results in cataplerosis and oxidative stress. J. Physiol. 2016, 594, 7341–7360. [Google Scholar] [CrossRef] [PubMed]
- Maurin, A.-C.; Parry, L.; B’chir, W.; Carraro, V.; Coudy-Gandilhon, C.; Chaouki, G.; Chaveroux, C.; Mordier, S.; Martinie, B.; Reinhardt, V.; et al. GCN2 upregulates autophagy in response to short-term deprivation of a single essential amino acid. Autophagy Rep. 2022, 1, 119–142. [Google Scholar] [CrossRef]
- Ezgu, F. Inborn Errors of Metabolism. Adv. Clin. Chem. 2016, 73, 195–250. [Google Scholar] [CrossRef]
- Qiu, J.; Thapaliya, S.; Runkana, A.; Yang, Y.; Tsien, C.; Mohan, M.L.; Narayanan, A.; Eghtesad, B.; Mozdziak, P.E.; McDonald, C.; et al. Hyperammonemia in cirrhosis induces transcriptional regulation of myostatin by an NF-κB-mediated mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 18162–18167. [Google Scholar] [CrossRef]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Cell Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef]
- Busquets, S.; Alvarez, B.; Llovera, M.; Agell, N.; López-Soriano, F.J.; Argilés, J.M. Branched-chain amino acids inhibit proteolysis in rat skeletal muscle: Mechanisms involved. J. Cell. Physiol. 2000, 184, 380–384. [Google Scholar] [CrossRef]
- Louard, R.J.; Barrett, E.J.; Gelfand, R.A. Overnight branched-chain amino acid infusion causes sustained suppression of muscle proteolysis. Metab. Clin. Exp. 1995, 44, 424–429. [Google Scholar] [CrossRef]
- Lysenko, E.A.; Vepkhvadze, T.F.; Lednev, E.M.; Vinogradova, O.L.; Popov, D.V. Branched-chain amino acids administration suppresses endurance exercise-related activation of ubiquitin proteasome signaling in trained human skeletal muscle. J. Physiol. Sci. JPS 2018, 68, 43–53. [Google Scholar] [CrossRef]
- Merli, M.; Berzigotti, A.; Zelber-Sagi, S.; Dasarathy, S.; Montagnese, S.; Genton, L.; Plauth, M.; Parés, A. EASL Clinical Practice Guidelines on nutrition in chronic liver disease. J. Hepatol. 2019, 70, 172–193. [Google Scholar] [CrossRef]
- Plauth, M.; Bernal, W.; Dasarathy, S.; Merli, M.; Plank, L.D.; Schütz, T.; Bischoff, S.C. ESPEN guideline on clinical nutrition in liver disease. Clin. Nutr. 2019, 38, 485–521. [Google Scholar] [CrossRef]
- Park, J.G.; Tak, W.Y.; Park, S.Y.; Kweon, Y.O.; Chung, W.J.; Jang, B.K.; Bae, S.H.; Lee, H.J.; Jang, J.Y.; Suk, K.T.; et al. Effects of Branched-Chain Amino Acid (BCAA) Supplementation on the Progression of Advanced Liver Disease: A Korean Nationwide, Multicenter, Prospective, Observational, Cohort Study. Nutrients 2020, 12, 1429. [Google Scholar] [CrossRef]
- Hanai, T.; Shiraki, M.; Imai, K.; Suetsugu, A.; Takai, K.; Shimizu, M. Late Evening Snack with Branched-Chain Amino Acids Supplementation Improves Survival in Patients with Cirrhosis. J. Clin. Med. 2020, 9, 1013. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Shiraishi, K.; Ito, T.; Suzuki, K.; Koreeda, C.; Ohtake, T.; Iwasa, M.; Tokumoto, Y.; Endo, R.; Kawamura, N.H.; et al. Branched-chain amino acids prevent hepatocarcinogenesis and prolong survival of patients with cirrhosis. Clin. Gastroenterol. Hepatol. 2014, 12, 1012–1018. [Google Scholar] [CrossRef]
- Hayaishi, S.; Chung, H.; Kudo, M.; Ishikawa, E.; Takita, M.; Ueda, T.; Kitai, S.; Inoue, T.; Yada, N.; Hagiwara, S.; et al. Oral branched-chain amino acid granules reduce the incidence of hepatocellular carcinoma and improve event-free survival in patients with liver cirrhosis. Dig. Dis. 2011, 29, 326–332. [Google Scholar] [CrossRef]
- Ruiz-Margáin, A.; Macías-Rodríguez, R.U.; Ríos-Torres, S.L.; Román-Calleja, B.M.; Méndez-Guerrero, O.; Rodríguez-Córdova, P.; Torre, A. Effect of a high-protein, high-fiber diet plus supplementation with branched-chain amino acids on the nutritional status of patients with cirrhosis. Rev. Gastroenterol. Mex. Engl. Ed. 2018, 83, 9–15. [Google Scholar] [CrossRef]
- Singh Tejavath, A.; Mathur, A.; Nathiya, D.; Singh, P.; Raj, P.; Suman, S.; Mundada, P.R.; Atif, S.; Rai, R.R.; Tomar, B.S. Impact of Branched Chain Amino Acid on Muscle Mass, Muscle Strength, Physical Performance, Combined Survival, and Maintenance of Liver Function Changes in Laboratory and Prognostic Markers on Sarcopenic Patients with Liver Cirrhosis (BCAAS Study): A Randomized Clinical Trial. Front. Nutr. 2021, 8, 619. [Google Scholar] [CrossRef]
- Les, I.; Doval, E.; García-Martínez, R.; Planas, M.; Cárdenas, G.; Gómez, P.; Flavià, M.; Jacas, C.; Mínguez, B.; Vergara, M.; et al. Effects of branched-chain amino acids supplementation in patients with cirrhosis and a previous episode of hepatic encephalopathy: A randomized study. Am. J. Gastroenterol. 2011, 106, 1081–1088. [Google Scholar] [CrossRef]
- Siramolpiwat, S.; Limthanetkul, N.; Pornthisarn, B.; Vilaichone, R.K.; Chonprasertsuk, S.; Bhanthumkomol, P.; Nunanan, P.; Issariyakulkarn, N. Branched-chain amino acids supplementation improves liver frailty index in frail compensated cirrhotic patients: A randomized controlled trial. BMC Gastroenterol. 2023, 23, 154. [Google Scholar] [CrossRef]
- Mohta, S.; Anand, A.; Sharma, S.; Qamar, S.; Agarwal, S.; Gunjan, D.; Singh, N.; Madhusudhan, K.S.; Pandey, R.M.; Saraya, A. Randomised clinical trial: Effect of adding branched chain amino acids to exercise and standard-of-care on muscle mass in cirrhotic patients with sarcopenia. Hepatol. Int. 2022, 16, 680–690. [Google Scholar] [CrossRef]
- Hernández-Conde, M.; Llop, E.; Gómez-Pimpollo, L.; Fernández Carrillo, C.; Rodríguez, L.; Van Den Brule, E.; Perelló, C.; López-Gómez, M.; Abad, J.; Martínez-Porras, J.L.; et al. Adding Branched-Chain Amino Acids to an Enhanced Standard-of-Care Treatment Improves Muscle Mass of Cirrhotic Patients with Sarcopenia: A Placebo-Controlled Trial. Am. J. Gastroenterol. 2021, 116, 2241–2249. [Google Scholar] [CrossRef]
- Yoo, S.Z.; No, M.H.; Heo, J.W.; Park, D.H.; Kang, J.H.; Kim, S.H.; Kwak, H.B. Role of exercise in age-related sarcopenia. J. Exerc. Rehabil. 2018, 14, 551–558. [Google Scholar] [CrossRef]
- Konopka, A.R.; Harber, M.P. Skeletal muscle hypertrophy after aerobic exercise training. Exerc. Sport Sci. Rev. 2014, 42, 53–61. [Google Scholar] [CrossRef]
- Meyer, F.; Bannert, K.; Wiese, M.; Esau, S.; Sautter, L.F.; Ehlers, L.; Aghdassi, A.A.; Metges, C.C.; Garbe, L.A.; Jaster, R.; et al. Molecular Mechanism Contributing to Malnutrition and Sarcopenia in Patients with Liver Cirrhosis. Int. J. Mol. Sci. 2020, 21, 5357. [Google Scholar] [CrossRef]


| Reference | Type of Study | Population | Etiology | BCAAs Analysis | Liver Function | Skeletal Muscle Function |
|---|---|---|---|---|---|---|
| [32] | Comparative study | 14 adults with CLD (12 males, 2 females) and 9 healthy controls. | 11-ALD 3-cryptogenetic | Spectrophotometry ↑ BCAAs (μM/kg) controls: Leu 132, Val 215, T. BCAAs 423. ↓ BCAAs (μM/kg) patients: Leu 84, Val 54, T. BCAAs 278. | Patients profile: -Albumin (gm/L): 28.6 ± 5.1 -PT (%): 67 ± 14 -Ammonia (µg/dL): 125 ± 50 -Ascites: 7 patients -Malnutrition: 7 patients | Intracellular muscle Val: Patients: 222 μM/kg (↓) Control: 368 μM/kg. |
| [33] | Cohort design | 14 patients with CLD (13 males, 1 female) and 7 healthy subjects (5 males, 2 females). | Non specify | HPLC with fluorescence detection. ↑ BCAAs (μM/kg) controls: Leu 132, Val 215, T. BCAAs 423. ↓ BCAAs (μM/kg) patients: Leu 84, Val 54, T. BCAAs 278. | Patients profile: -Albumin (μmol/L): 520 ± 21 -Total bilirubin (μmol/L): 21.8 ± 3.2-PT index: 0.58 ± 0.03 -CP score: A: 6; B: 8; C: 0 patients. Healthy profile: -Albumin (μmol/L): 639 ± 27 -Total bilirubin (μmol/L): 7.14 ± 1.1 -PT index: 0.94 ± 0.05 | Uptake of BCAAs across muscle leg (femoral artery): Control: 196 ± 67 μmol/min Patients: −84.7 ± 110 μmol/min). |
| [34] | Cohort design, retrospective | 13 patients with CLD and 6 patients with an episode of alcoholic hepatitis (16 males, 4 females) and 6 healthy controls (2 females, 5 males) | 6-ALD 13-Other | ↑ BCAAs (μmol/L) controls: Leu 244 ± 19, Val 256 ± 19, T. BCAAs 646 ± 44. ↓ BCAAs (μmol/L) patients: Leu 185 ± 18, Val 201 ± 15, T. BCAAs 535 ± 42. | ALD patients profile: -Albumin (μmol/L): 469 ± 42 -Total bilirubin (μmol/L): 380 ± 42 -PT index: 0.24 ± 0.04 -CP score: A: 0; B: 2; C: 4 patients. CLD patients profile: -Albumin (μmol/L): 520 ± 21 -Total bilirubin (μmol/L): 21.8 ± 3 -PT index: 0.58 ± 0.03 -CP score: A: 6; B: 7; C: 0 patients | Leg muscle BCAAs uptake: Patients with alcoholic hepatitis: Total BCAAs 0.48 ± 23 μmol/L Stable cirrhosis: Total BCAAs 32 ± 22 μmol/L Controls: Total BCAAs −11.9 ± 22 μmol/L |
| [35] | Prospective control trial | 127 patients with CLD (81 males, 43 females) | 68-HBV 23-ALD 26-MASLD 7-Other | Colorimetric method Average of BCAAs 453.73 +/− 40.17 (μmol/L) | Patients profile: -Albumin (g/dL): 32.48 ± 4.53 -Total bilirubin (μmol/L): 36.65 ± 13.16 (μmol/L) -PT (%) 62.48 ± 10.45 -CP score: A: 71; B: 53 patients. | Patients had decreased handgrip strength (kg): 25.47 ± 5.84 and SMI (cm2/m2): 49.61 ± 8.84. 37 patients with sarcopenia. |
| [36] | Case control | 21 patients with CLD (9 males, 12 females) | 18-HCV 2-HBV 1-ALD | Patients with Albumin improvement (11): ↓ Total BCAAs 377.4 ± 102.2 μmol/L Patients with no Albumin improvement (10): ↑ Total BCAAs 413.4 ± 89.2 μmol/L. | Patients profile: -Albumin (g/dL): 3.2 ± 0.4 -Total bilirubin (mg/dL): 1.2 ± 0.5 -Ammonia (μg/dL) 57.7 ± 30.1 -CP score: A: 10; B: 11; C: 12 patients. | SMI area: 12.4 ± 2.7 cm2/m2. IMAC: −0.11 ± 0.16. |
| [37] | Control trial | 8 healthy controls (4 males, 4 females). 6 patients with CLD (5 males, 1 female). | ALD cirrhosis | Ion exchange chromatography ↑ BCAAs (μmol/L) controls: Leu 119.13 ± 9.35, Ile 77.02 ± 5.78, Val 245.70 ± 18.69. ↓ BCAAs (μmol/L) patients: Leu 81.33 ± 7.75, Ile 55.15 ± 4.12, Val 166.99 ± 12.44. | Patients profile: -Albumin (μmol/L): 4.20 ± 0.12 -CP score: 5.17 ± 0.15 -MELD score: 6.21 ± 0.23 -Ammonia (μg/dL) 98.24 ± 6.75 | Patients muscle tissue: -Lower activation of mTOR-related proteins (p70S6K, S6, 4EBP1) -↑ Intracellular amino acid sensor (GCN2) and myostatin protein. |
| [38] | Cross-sectional, single-center analysis. | 92 patients with CLD (60 males, 32 females). | 7-Viral hepatitis 24-Autoimmune 28-MASLD 20-ALD 2-Storage disorder 3-Malignancy 8-Other | NMR spectroscopy platform Control group: ↑ T. BCAAs (μmol/L): -Men: 412.7 -Women: 339.7 Patients group: ↓ T. BCAAs (μmol/L): -Men: 307.0 -Women: 213.5 | Patients profile: -Albumin (g/L): 32.7 ± 6.5 -Total bilirubin (µmol/L): 41 -Ammonia (µmol/L) 68.8 ± 33.4 -MELD score 15 ± 6. | -Lowest BCAAs tertile had higher CP scores (7 ± 2) -The 4 m walking test, standing balance test and CFS were not significantly different between BCAAs tertiles -Inverse correlations of the Timed up and go performance test with total BCAAs, Val and Ile. -Total BCAAs, Val and Leu were inversely related in the sit-to-stand test |
| [39] | Comparative study | 8 healthy subjects (6 males, 2 females) and 8 patients with CLD (4 males, 4 females) | 6-HCV 1-HBV 1-Other | Patients group: Fischer’s ratio 0.68–2.31 | Patients profile: -Albumin (g/dL): 2.8–3.4 -Total bilirubin (mg/dL): 0.6–2.1 -Ammonia (µg/dL): 46–122 -PT (%): 58–77 | After exercise loading intramuscular ⊿pH: -Controls 0.20 ± 0.18 -Patients 0.49 ± 0.16 Creatine phosphate: -Controls 0.35 ± 0.19 -Patients 0.55 ± 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trillos-Almanza, M.C.; Martinez-Aguilar, M.; Arroyave-Ospina, J.C.; van Vilsteren, F.; Blokzijl, H.; Moshage, H. Clinical and Therapeutic Implications of BCAAs Metabolism during Chronic Liver Disease in Humans: Crosstalk between Skeletal Muscle and Liver. Muscles 2024, 3, 71-87. https://doi.org/10.3390/muscles3010008
Trillos-Almanza MC, Martinez-Aguilar M, Arroyave-Ospina JC, van Vilsteren F, Blokzijl H, Moshage H. Clinical and Therapeutic Implications of BCAAs Metabolism during Chronic Liver Disease in Humans: Crosstalk between Skeletal Muscle and Liver. Muscles. 2024; 3(1):71-87. https://doi.org/10.3390/muscles3010008
Chicago/Turabian StyleTrillos-Almanza, Maria Camila, Magnolia Martinez-Aguilar, Johanna C. Arroyave-Ospina, Frederike van Vilsteren, Hans Blokzijl, and Han Moshage. 2024. "Clinical and Therapeutic Implications of BCAAs Metabolism during Chronic Liver Disease in Humans: Crosstalk between Skeletal Muscle and Liver" Muscles 3, no. 1: 71-87. https://doi.org/10.3390/muscles3010008