
Evaluation of Germination under Different Storage Conditions of Four Endemic Plant Species from Ethiopia: Implications for Ex Situ Conservation in Seed Banks

Abstract
:1. Introduction
2. Material and Methods
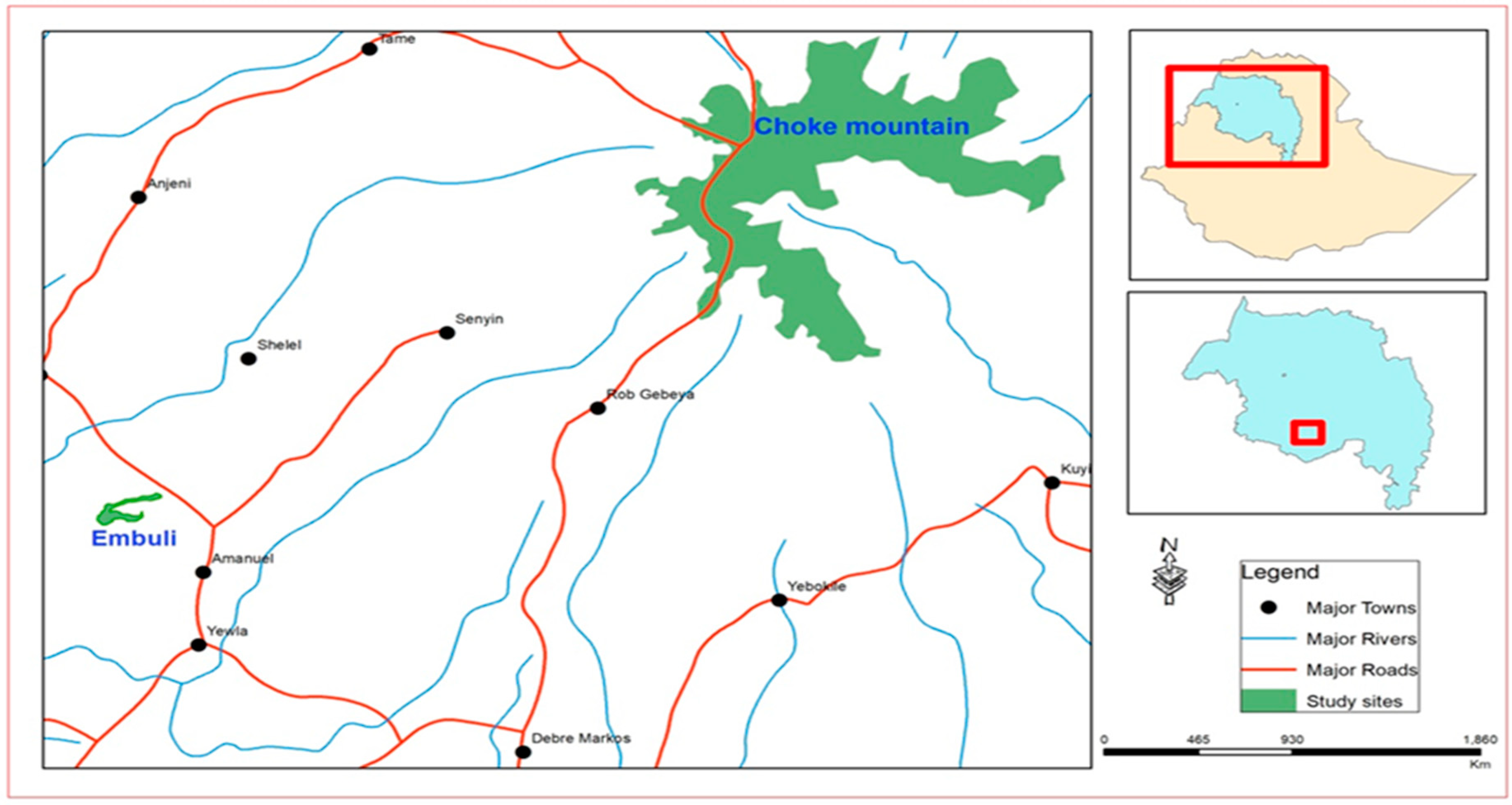
2.1. Study Site
2.2. Study Species Description and Seed Collection
2.3. Seed Storage
2.4. Germination Tests
2.5. Statistical Analyses
3. Results
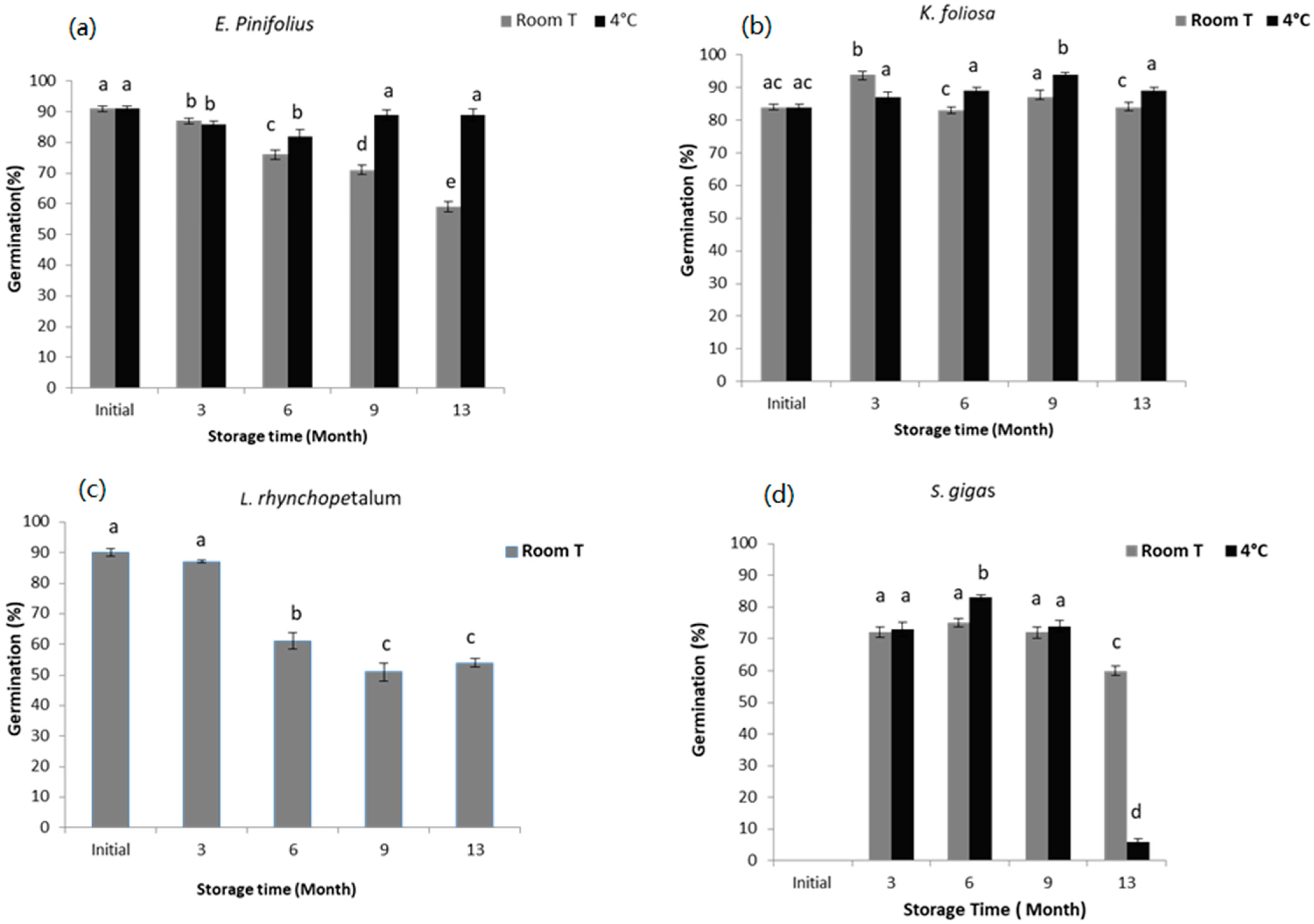
3.1. Germination Test before Drying
3.1.1. Euryops pinifolius
3.1.2. Kniphofia foliosa
3.1.3. Lobelia rhynchopetalum
3.1.4. Solanecio gigas
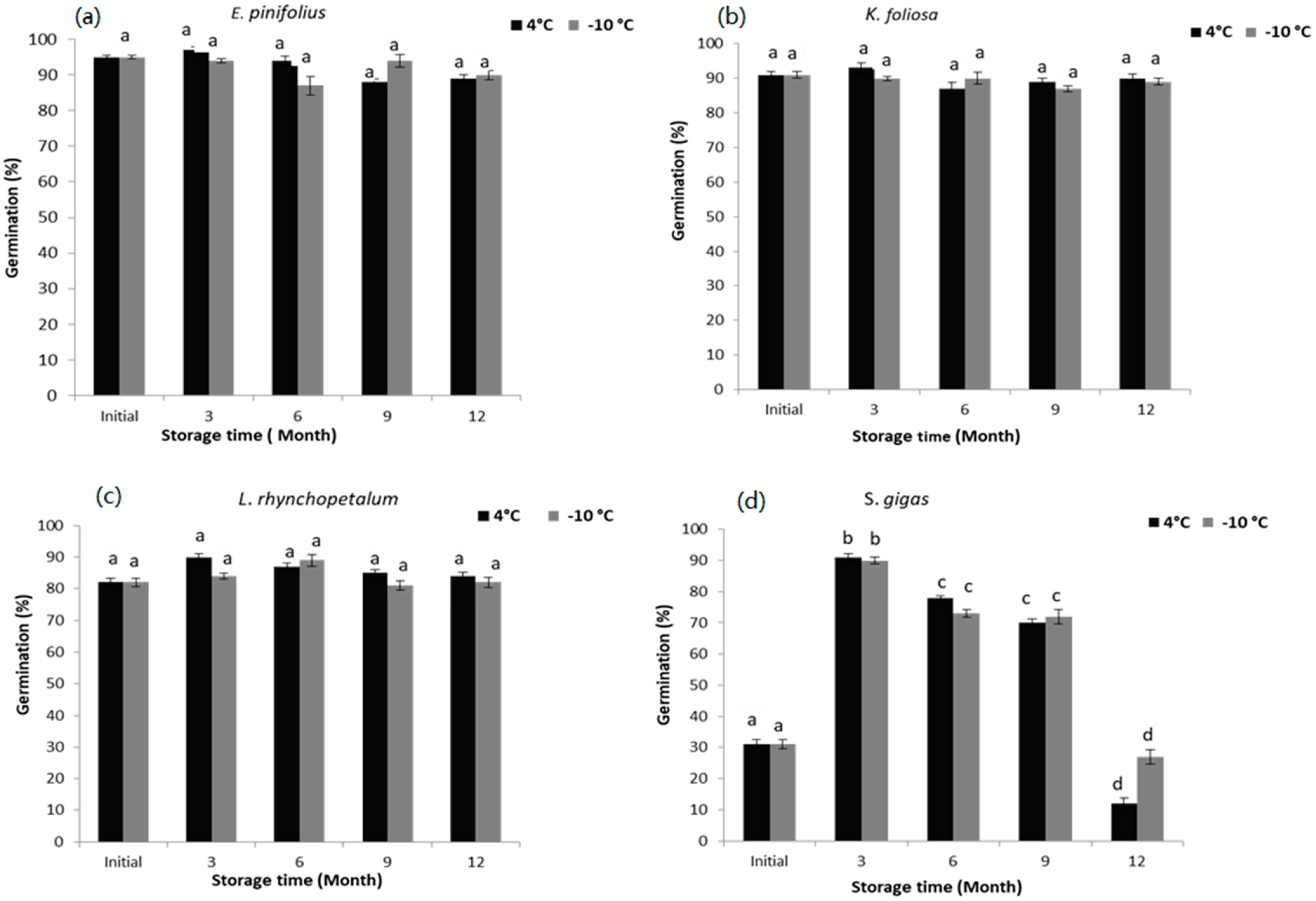
3.2. Germination Test after Drying
3.2.1. Euryops pinifolius
3.2.2. Kniphofia foliosa
3.2.3. Lobelia rhynchopetalum
3.2.4. Solanecio gigas
4. Discussion
4.1. Germination before Drying
4.2. Germination after Drying
5. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 19, 1024–1026. [Google Scholar] [CrossRef]
- Lenoir, J.; Gegout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H.A. Significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef] [PubMed]
- Diaz, H.F.; Grosjean, M.; Graumlich, L. Climate variability and change in high elevation regions: Past, present and future. Clim. Change 2003, 59, 1–4. [Google Scholar] [CrossRef]
- Chala, D.; Brochmann, C.; Psomas, A.; Ehrich, D.; Gizaw, A.; Masao, C.A.; Bakkestuen, V.; Zimmermann, N.E. Good-bye to tropical alpine plant giants under warmerclimates? Loss of range and genetic diversity in Lobelia rhynchopetalum. Ecol. Evol. 2016, 6, 8931–8941. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59. [Google Scholar] [CrossRef]
- Ruijven, J.V.; Berendse, F. Diversity enhances community recovery, but not resistance, after drought. J. Ecol. 2010, 98, 81–86. [Google Scholar] [CrossRef]
- Fetene, A.; Yeshitela, K.; Desta, H. Approaches to Conservation and Sustainable Use of Biodiversity—A Review. Nat. Sci. 2012, 10, 51–62. [Google Scholar]
- Bekele, T. Forest Genetic Resource conservation strategy of Ethiopia. In Proceedings of the National Conference on Forest Resources of Ethiopia: Status Challenges and Opportunities, Addis Ababa, Ethiopia, 27–29 November 2002. [Google Scholar]
- Dhillon, B.S.; Dua, R.P.; Brahmi, P.; Bisht, I.S. On-Farm conservation of plantgenetic resources for food and agriculture. Curr. Sci. 2004, 87, 557–559. [Google Scholar]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Panobianco, M.; Vieira, R.D.; Perecin, D. Electrical conductivity as an indicator of pea seed aging of stored at different temperatures. Sci. Agric. 2007, 64, 119–124. [Google Scholar] [CrossRef]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The evolution of seeds. New Phytologist. 2010, 186, 817–831. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-savage, W.E. The ecophysiology of seed persistence: A mechanistic view of the journey to germination or demise. Biol. Rev. Camb. Philos. Soc. 2014, 90, 31–59. [Google Scholar] [CrossRef]
- Donohue, K.; Rubio De Casas, R.; Burghardt, L.; Kovach, K.; Willis, C.G. Germination, postgermination adaptation, and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 293–319. [Google Scholar] [CrossRef]
- Kissmann, C.; Tozzia, H.H.; Martinsa, S.; Habermannb, G. Germination performance of congeneric Styrax species from the Cerrado sensu lato areas and their distribution pattern in different physiognomies. Flora 2012, 207, 673–681. [Google Scholar] [CrossRef]
- Roberts, E.H. Loss of viability: Ultrastuctural and physiological aspects. Seed Sci. Technol. 1973, 1, 529–545. [Google Scholar]
- Corner, K.F.; Sowa, S. Recalcitrant Behavior of Temperate Forest trees: Storage Biochemistry, and physiology. In Proceedings of the Eleventh Biennial Southern Silvicultural Research Conference, Knoxville, TN, USA, 20–22 March 2002; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2002; pp. 47–50. [Google Scholar]
- Hong, T.D.; Ellis, R.H. A Protocol to Determine Seed Storage Behavior. In International Plant Genetic Resources; Engels, J.M.M., Toll, J., Eds.; IPGRI Technical Bulletin. Institute: Rome, Italy, 1996. [Google Scholar]
- Bonner, F.T. Commercial seed supply of recalcitrant and intermediate seed: Present solution to the storage problem. In Proceeding of a Workshop on Improved Methods for Handling and Storage of Intermediate/Recalcitrant Tropical Forest Tree Seeds, Humleback, Denmark, 8–10 June 1995; Ouêdraogo, A.S., Poulsen, K., Stubsgaard, F., Eds.; IPGRI: Rome, Italy, 1996; pp. 27–33. [Google Scholar]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An intermediate category of seed storage behavior? I. Coffee. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Balcha, G.; Blyth, J. The use of Tetrazolium chloride test to determine viability of Afrocarpus gracilior (Pilger) C.N. Page (Podocarpaceae) seeds kept at different storage environments. Ethiop. J. Nat. Resour. 2000, 2, 79–92. [Google Scholar]
- Murthy, U.M.N.; Kumar, P.P.; Sun, W.Q. Mechanisms of seed ageing under different storage conditions for Vigna radiata (L.) Wilczek: Lipid peroxidation Sugar hydrolysis, Maillard reactions and their relationship to glass state transition. J. Exp. Bot. 2003, 54, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Kurek, K.; Plitta-Michalak, B.; Ratajczak, E. Reactive oxygen species as potential drivers of the seed aging process. Plants 2019, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, E.; Małecka, A.; Bagniewska-Zadworna, A.; Kalemba, E.M. The production, localization and spreading of reactive oxygen species contributes to the low vitality of long-term stored common beech (Fagus sylvatica L.) seeds. J. Plant Physiol. 2015, 174, 147–156. [Google Scholar] [CrossRef]
- Walsh, D.G.F.; Waldren, S.; Martin, J.R. Monitoring seed viability of fifteen species after storage in the Irish Threatened Plant Genebank. Biol. Environ. 2003, 103, 59–67. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Balcha, G. Conservation and Sustainable use of forest genetic resources. In Proceedings of the National Conference on Forest Resources of Ethiopia: Status Challenges and Opportunities, Addis Ababa, Ethiopia, 27–29 November 2002. [Google Scholar]
- Tenner, C. Establishing priorities for plant conservation Program. In Seed Conservation Turning Science into Practice; Smith, R.D., Dicke, J.B., Linington, S.H., Pritchard, H.W., Probert, R.J., Eds.; Royal Botanical Gardens: Kew, UK, 2003; pp. 27–35. [Google Scholar]
- Miranto, M. Living Collections of Botanic Gardens as a Means of Ex-Situ Conservation. Case Study of African Violets (Saintpaulia) in Europe. Master’s Thesis, Faculty of Biological and Environmental Sciences, University of Helsinki, Helsinki, UK, 2005. [Google Scholar]
- Demissew, S.; Friis, I.; Weber, O. Diversity and endemism of the flora of Ethiopia and Eritrea: State of knowledge and future perspectives. Rend. Lincei 2021, 32, 675–697. [Google Scholar] [CrossRef]
- Teketay, D. Deforestation, wood famine, and environmental degradation in Ethiopia’s highland ecosystems: Urgent need for action. N. Afr. Stud. 2001, 8, 53–76. [Google Scholar] [CrossRef]
- Vivero, J.L.; Kelbessa, E.; Demissew, S. The Red List of Endemic Trees & Shrubs of Ethiopia and Eritrea; Fauna & Flora International: Cambridge, UK, 2005; p. 14. [Google Scholar]
- Erenso, F.; Maryo, M. Endemic plant species composition and their status in Boda Dry evergreen Montane forest, West Shewa, Ethiopia. Int. J. Biodivers. Conserv. 2014, 6, 563–569. [Google Scholar] [CrossRef]
- Teferi, E.; Uhlenbrook, S.; Bewket, W.; Wenninger, J.; Simane, B. The use of remote sensing to quantify wetland loss in Choke Mountain range, upper Blue Nile basin, Ethiopia. Hydrol. Earth Syst. Sci. 2010, 7, 6243–6284. [Google Scholar] [CrossRef]
- Simane, B.; Zaitchik, B.F.; Ozdogan, M. Agro ecosystem analysis of the Choke Mountain watersheds, Ethiopia. Sustainability 2013, 5, 592–616. [Google Scholar] [CrossRef]
- Tadesse, M. The Family Asteraceae (Compositae). In Flora of Ethiopia and Eritrea; Hedberg, I., Friis, I., Edwards, S., Eds.; Uppsala University: Uppsala, Sweden, 2004; Volume 4, pp. 249–267. [Google Scholar]
- Thulin, M. The family Lobeliaceae. In Flora of Ethiopia and Eritrea; Hedberg, I., Kelbessa, E., Edwards, E., Demissew, S., Persson, E., Eds.; Uppsala University: Uppsala, Sweden, 2006; Volume 5, p. 40. [Google Scholar]
- Demissew, S.; Nordal, I. The family Asphodelaceae. In Flora of Ethiopia and Eritrea; Edwards, S., Demissew, S., Hedberg, I., Eds.; Uppsala University: Uppsala, Sweden, 1997; Volume 6, p. 107. [Google Scholar]
- Rao, N.K.; Hanson, J.; Dulloo, M.E.; Ghosh, K.; Nowell, D.; Larine, M. Seed moisture content determination and drying. In Manuals of Seed Handling in Genebanks; Handbooks for Genebanks No. 8; Bioversity International: Rome, Italy, 2006; pp. 28–34. [Google Scholar]
- Yenesew, A.; Dagne, E.; Muller, M.; Steglkh, W. An anthrone, an anthraquinone and two oxanthrones from Kniphofia foliosa. Photochemistry 1994, 37, 525–528. [Google Scholar] [CrossRef]
- Amsalu, N.; Bezie, Y.; Fentahun, M.; Alemayehu, A.; Amsalu, G. Use and Conservation of Medicinal Plants by Indigenous People of Gozamin Wereda, East Gojjam Zone of Amhara Region, Ethiopia: An Ethnobotanical Approach. Evid.-Based Complement. Altern. Med. 2018, 2018, 2973513. [Google Scholar] [CrossRef]
- Simegniew Birhan, Y.; Leshe Kitaw, S.; Abebe Alemayehu, Y.; Minuye Mengesha, N. Ethnobotanical Study of Medicinal Plants Used to treat Human Diseases in Enarj Enawga District, East Gojjam Zone, Amhara Region, Ethiopia. SM J. Med. Plant Stud. 2017, 1, 1006. [Google Scholar] [CrossRef]
- Dagnachew, S.; Teketay, D.; Demissew, S.; Hawas, T.; Kindu, M. Herbarium-based study of phenology of twelve indigenous and endemic species from Ethiopia. South Afr. J. Bot. 2022, 150, 260–274. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall: Hoboken, NJ, USA, 2010; pp. 778–779. [Google Scholar]
- Sivakumar, V.; Anandalakshmi, R.; Warrier, R.R.; Tigabu, M.; Odén, P.C.; Vijayachandran, S.N.; Geetha, S.; Singh, B.G. Effects of presowing treatments, desiccation and storage conditions on germination of Strychnos nux-vomica seeds, a valuable medicinal plant. New For. 2006, 32, 121–131. [Google Scholar] [CrossRef]
- Chauhan, R.S.; Nautiyal, M.C. Seed germination and seed storage behavior of Nardostachys jatamansi D.C., an endangered medicinal herb of high-altitude Himalaya. Curr. Sci. 2007, 92, 1620–1624. [Google Scholar]
- Pradhan, B.K.; Badola, H.K. Effect of storage conditions and storage periods on seed germination in eleven populations of Swertia chirayita: A critically endangered medicinal herb in Himalaya. Sci. World J. 2012, 2012, 128105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.H.; Baskin, C.C.; Chen, W.; Du, G.Z. Variation in seed germination between populations of five sub-alpine woody species from eastern Qinghai Tibet Plateau following dry storage at low temperatures. Ecol. Res. 2010, 25, 195–203. [Google Scholar] [CrossRef]
- Ruiz, M.; Martin, I.; de la Cuadra, C. Cereal seed viability after 10 years of storage in active and base germplasm collections. Field Crops Res. 1999, 64, 229–236. [Google Scholar] [CrossRef]
- Nagel, M.; Börner, A. The longevity of crop seeds stored under ambient conditions. Seed Sci. Res. 2010, 20, 1–12. [Google Scholar] [CrossRef]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. 2008, 101, 213–228. [Google Scholar] [CrossRef]
- Pradhan, B.K.; Badola, H.K. Seed germination response of populations of Swertia chirayita following periodical storage. Seed Technol. 2008, 30, 63–69. [Google Scholar]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: New York, NY, USA, 2005; p. 97. [Google Scholar]
- Bewley, J.D.; Black, M. Physiology and Biochemistry of Seeds in Relation to Germination. In Physiology and Biochemistry of Seeds in Relation to Germination; Springer: Berlin/Heidelberg, Germany, 1978; p. 13. [Google Scholar] [CrossRef]
- Nyo, H.T.; Htwe, N.N.; Win, K.K. Effect of Different Packaging Materials and Storage Environments on Seed Quality of Sesame (Sesamum indicum, L.). J. Biol. Life Sci. 2019, 11, 1. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S No | Species | Local Name | Family | Altitudinal Range and Brief Botanical Description | Geographical Distribution throughout the Country | Place, Month, Year, and Altitude of Collection (Geographical Coordinates) | Seed Moisture Content (%) during Storage | Local Uses | Conservation Status * |
|---|---|---|---|---|---|---|---|---|---|
| 1 | E. pinifolius A. Rich. | Gimiy | Asteraceae | Alt 3200–3700 m.Shrub or shrublet, 30–100 cm high. Branches leafy in upper parts, leafless below with prominent leaf scars. | GJ, WU, and SU | Choke; January–February 2019; 3920 m (10°41′47.8′′ N × 37°50′14.5′′ E) | 2.8 | Used to treat stomachache. (kurba) | VU |
| 2 | K. foliosa Hochst. | Ashengdiy | Asphodelaceae | Alt 2500–4000 m.Robust plants forming dense clumps, with thick erect rhizomes, sometimes with a stem up to 40 cm high. | TU, GD GJ, WU, SU, AR, BA, and HA | Choke; March 2019; 3757 m (10°44′01.2′′ N × 37°47′05.5′′ E) | 2.8 | Used for the treatment of abdominal cramps and for wound healing, inhibit the growth of malaria parasite. | unknown |
| 3 | L. rhynchopetalum Hemsl. | Gibira | Lobeliaceae | Alt 3000–4350 m.Plant up to 7 m high when in flower; stem erect, more or less woody towards the base. | GD, GJ, SU, AR, BA, and HA | Choke; March 2019; 3906 m (10°39′47.8′′ N × 037°69′34.2′′ E) | 3.5 | The hollow woody stems serve as long trumpets for shepherd boys, used to treat evil eye. | High riskof extinction |
| 4 | S. gigas (Vatke) C. Jeffrey | Boz | Asteraceae | Alt 1750–3350 m.Shrub or small tree, 1.75–6 m high. Stems and branches leafless below, but with a leafy crown near apex. | GD, GJ, WU, SU, AR, SD, IL, KF, BA, and HA | Embuli; January 2019; 2302 m(10°29′19.7′′ N × 037°31′17.5′′ E) | 2.9 | Used to treat dysentery and bloating, juice of leaves used as internal medicine for cattle and hedge plant around small home gardens. | LC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dagnachew, S.; Teketay, D.; Demissew, S.; Awas, T. Evaluation of Germination under Different Storage Conditions of Four Endemic Plant Species from Ethiopia: Implications for Ex Situ Conservation in Seed Banks. Seeds 2023, 2, 45-59. https://doi.org/10.3390/seeds2010005
Dagnachew S, Teketay D, Demissew S, Awas T. Evaluation of Germination under Different Storage Conditions of Four Endemic Plant Species from Ethiopia: Implications for Ex Situ Conservation in Seed Banks. Seeds. 2023; 2(1):45-59. https://doi.org/10.3390/seeds2010005
Chicago/Turabian StyleDagnachew, Sinework, Demel Teketay, Sebsebe Demissew, and Tesfaye Awas. 2023. "Evaluation of Germination under Different Storage Conditions of Four Endemic Plant Species from Ethiopia: Implications for Ex Situ Conservation in Seed Banks" Seeds 2, no. 1: 45-59. https://doi.org/10.3390/seeds2010005