

Methane Oxidation via Chemical and Biological Methods: Challenges and Solutions


Abstract
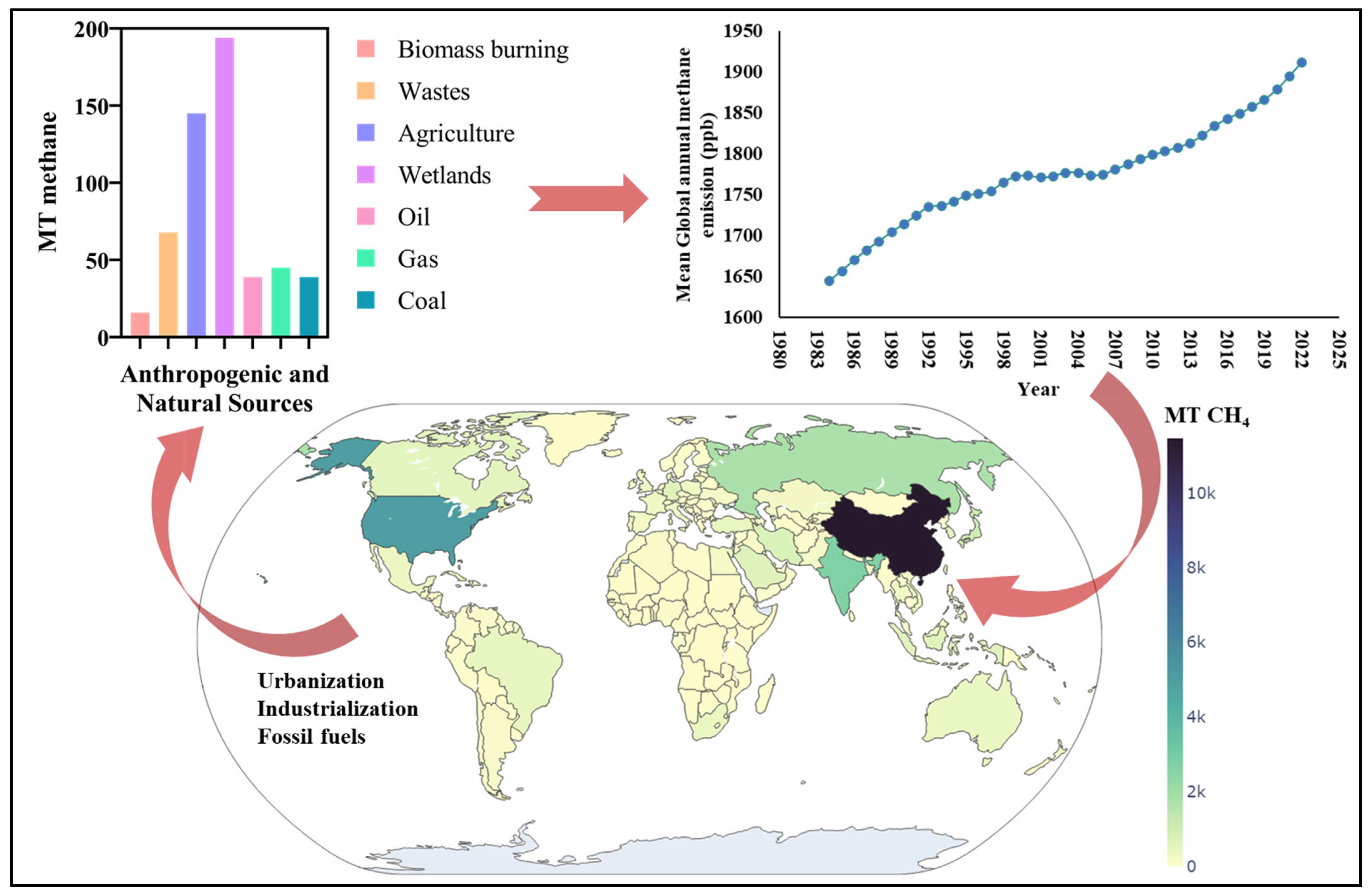
:1. Introduction

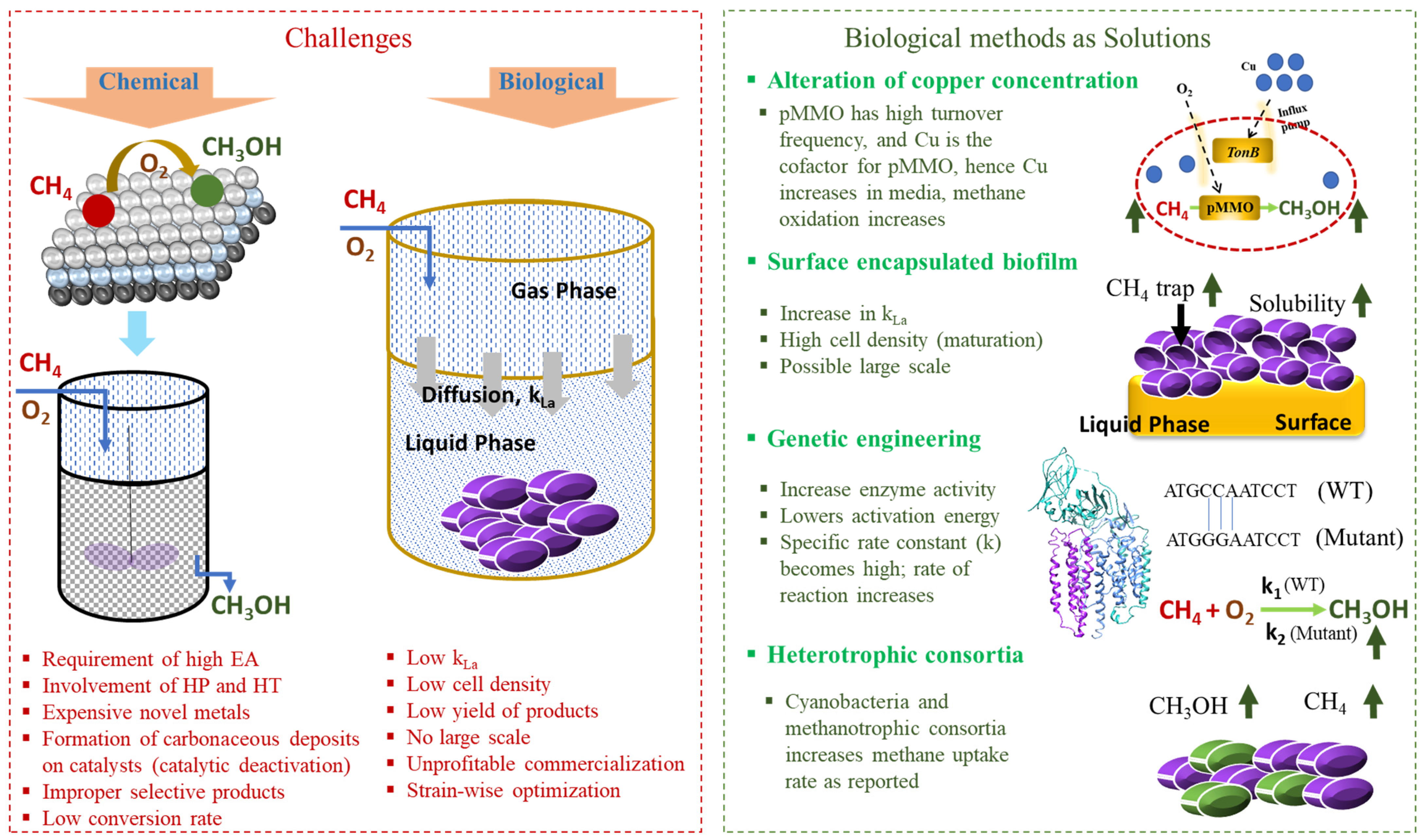
2. Challenges in Methane Oxidation
3. Solution to Overcome the Challenges
3.1. Chemical Methods
3.1.1. Traditional Methods for Direct Atmospheric Methane Removal
3.1.2. Chemical Methods and Reported Benchmark for Methane to Methanol Conversion
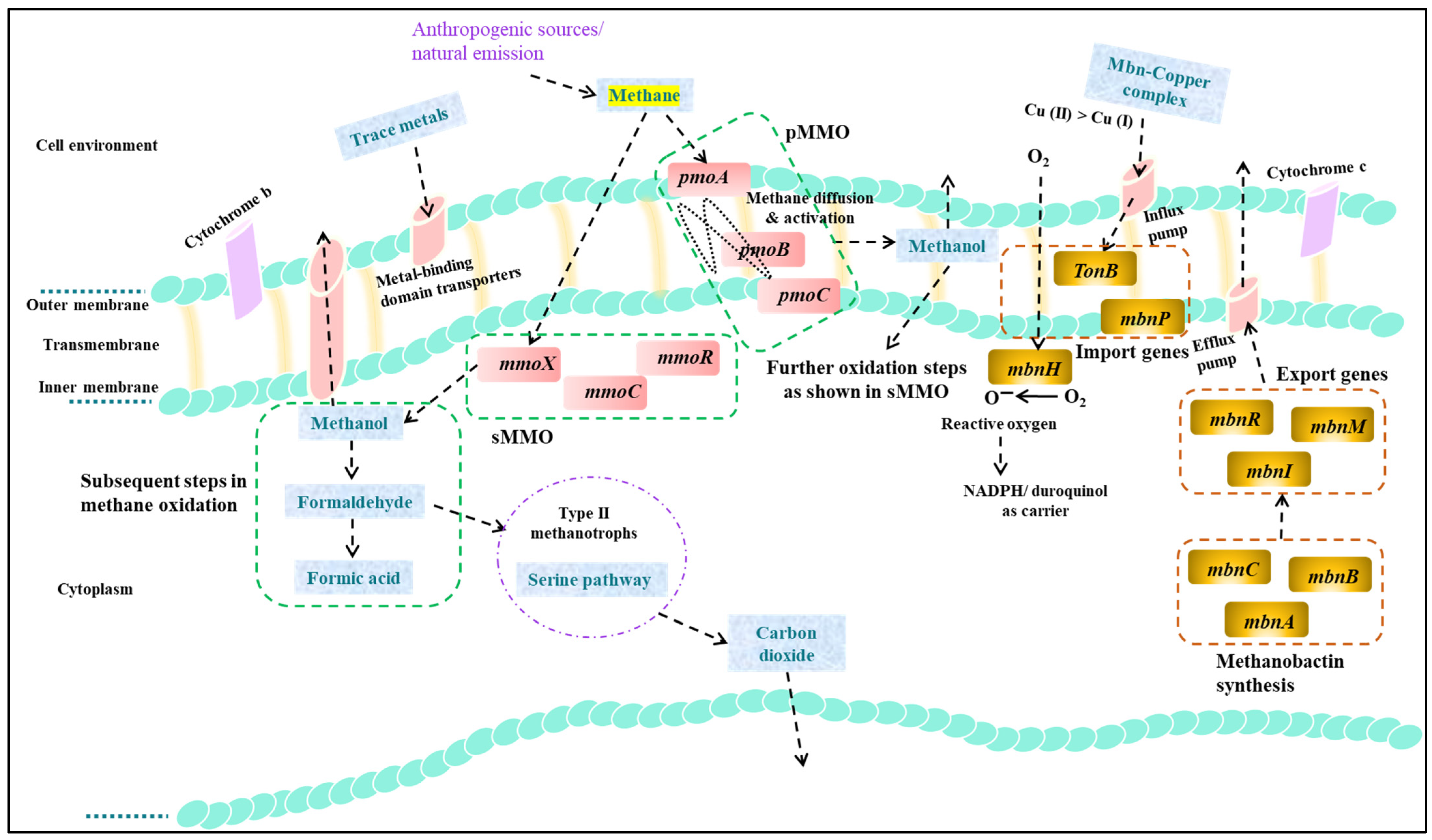
3.2. Biological Methods
3.2.1. Bioreactor and Biofilter Based Method
3.2.2. Alteration in Metal Concentrations
3.2.3. Synergetic Growth and Consortia
3.2.4. Genetic Engineering of MMOs
3.2.5. Enzyme and Whole Cell Immobilization
4. Challenges in Upscaling of Biological Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skea, J.; Shukla, P.; Kılkış, Ş. Climate Change 2022: Mitigation of Climate Change; Cambridge University Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Wuebbles, D.J.; Jain, A.K. Concerns About Climate Change and the Role of Fossil Fuel Use. Fuel Process. Technol. 2001, 71, 99–119. [Google Scholar] [CrossRef]
- Administration, National Oceanic and Atmospheric. Trends in Methane Emission. Available online: https://gml.noaa.gov/ccgg/trends_ch4/ (accessed on 18 February 2023).
- Agency, United States Environmental Protection. Global Methane Initiative. Environmental Protection Agency. Available online: https://www.epa.gov/gmi/importance-methane#:~:text=Methane%20is%20more%20than%2025,due%20to%20human%2Drelated%20activities (accessed on 1 May 2023).
- Halford, J.O.; Miller, G.A. Standard Heat Capacities of Gaseous Methanol, Ethanol, Methane and Ethane at 279 K. By Thermal Conductivity. J. Phys. Chem. 1957, 61, 1536–1539. [Google Scholar] [CrossRef]
- Karakurt, I.; Aydin, G.; Aydiner, K. Sources and mitigation of methane emissions by sectors: A critical review. Renew. Energy 2012, 39, 40–48. [Google Scholar] [CrossRef]
- Raihan, A.; Muhtasim, D.A.; Farhana, S.; Pavel, M.I.; Faruk, O.; Rahman, M.; Mahmood, A. Nexus between carbon emissions, economic growth, renewable energy use, urbanization, industrialization, technological innovation, and forest area towards achieving environmental sustainability in Bangladesh. Energy Clim. Chang. 2022, 3, 100080. [Google Scholar] [CrossRef]
- Chandra, R.; Takeuchi, H.; Hasegawa, T. Methane production from lignocellulosic agricultural crop wastes: A review in context to second generation of biofuel production. Renew. Sustain. Energy Rev. 2012, 16, 1462–1476. [Google Scholar] [CrossRef]
- Meylan, F.D.; Piguet, F.-P.; Erkman, S. Power-to-gas through CO2 methanation: Assessment of the carbon balance regarding EU directives. J. Energy Storage 2017, 11, 16–24. [Google Scholar] [CrossRef]
- Samanta, D.; Govil, T.; Salem, D.R.; Krumholz, L.R.; Gerlach, R.; Gadhamshetty, V.; Sani, R.K. Methane Monooxygenases: Their Regulations and Applications in Biofuel Production. In Microbes for Sustainable Development and Bioremediation; CRC Press: Boca Raton, FL, USA, 2019; pp. 187–206. [Google Scholar]
- Muh, E.; Tabet, F.; Amara, S. Biomass Conversion to Fuels and Value-Added Chemicals: A Comprehensive Review of the Thermochemical Processes. Curr. Altern. Energy 2021, 4, 3–25. [Google Scholar] [CrossRef]
- Soulivong, D.; Norsic, S.; Taoufik, M.; Coperet, C.; Thivolle-Cazat, J.; Chakka, S.; Basset, J.-M. Non-Oxidative Coupling Reaction of Methane to Ethane and Hydrogen Catalyzed by the Silica-Supported Tantalum Hydride: (SiO)2Ta−H. J. Am. Chem. Soc. 2008, 130, 5044–5045. [Google Scholar] [CrossRef]
- Ravi, M.; Sushkevich, V.L.; Knorpp, A.J.; Newton, M.A.; Palagin, D.; Pinar, A.B.; Ranocchiari, M.; van Bokhoven, J.A. Misconceptions and challenges in methane-to-methanol over transition-metal-exchanged zeolites. Nat. Catal. 2019, 2, 485–494. [Google Scholar] [CrossRef]
- Horn, R.; Schlögl, R. Methane activation by heterogeneous catalysis. Catal. Lett. 2015, 145, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Samanta, D.; Govil, T.; Salem, D.R.; Krumholz, L.R.; Gerlach, R.; Gadhamshetty, V.; Sani, R.K. 12 Methane Monooxygenases. In Microbes for Sustainable Development and Bioremediation, 1st ed.; CRC Press: Boca Raton, FL, USA, 2019; p. 187. [Google Scholar] [CrossRef]
- Lidstrom, M.E. Aerobic methylotrophic prokaryotes. Prokaryotes 2006, 2, 618. [Google Scholar]
- AlSayed, A.; Fergala, A.; Eldyasti, A. Sustainable biogas mitigation and value-added resources recovery using methanotrophs intergrated into wastewater treatment plants. Rev. Environ. Sci. Bio/Technol. 2018, 17, 351–393. [Google Scholar] [CrossRef]
- Murrell, J.C.; McDonald, I.R.; Bourne, D.G. Molecular methods for the study of methanotroph ecology. FEMS Microbiol. Ecol. 1998, 27, 103–114. [Google Scholar] [CrossRef]
- Khadem, A.F. Methanotrophy under Extreme Conditions: Biochemistry and Physiology of Methylacidiphilum Fumariolicum SolV. Ph.D Thesis, Radboud University, Nijmegen, The Netherlands, 2012. [Google Scholar]
- McKinsey Global Institute. The BioRevolution. In Innovations Transforming Economies, Societies, and Our Lives; McKinsey Global Institute: New York, NY, USA, 2020; pp. 1–200. [Google Scholar]
- Fei, Q.; Guarnieri, M.T.; Tao, L.; Laurens, L.M.; Dowe, N.; Pienkos, P.T. Bioconversion of natural gas to liquid fuel: Opportunities and challenges. Biotechnol. Adv. 2014, 32, 596–614. [Google Scholar] [CrossRef] [Green Version]
- Comesaña-Gándara, B.; García-Depraect, O.; Santos-Beneit, F.; Bordel, S.; Lebrero, R.; Muñoz, R. Recent trends and advances in biogas upgrading and methanotrophs-based valorization. Chem. Eng. J. Adv. 2022, 11, 100325. [Google Scholar] [CrossRef]
- Blanchette, C.D.; Knipe, J.M.; Stolaroff, J.K.; DeOtte, J.R.; Oakdale, J.S.; Maiti, A.; Lenhardt, J.M.; Sirajuddin, S.; Rosenzweig, A.C.; Baker, S.E. Printable enzyme-embedded materials for methane to methanol conversion. Nat. Commun. 2016, 7, 11900. [Google Scholar] [CrossRef] [Green Version]
- Sahoo, K.K.; Goswami, G.; Das, D. Biotransformation of methane and carbon dioxide into high-value products by methanotrophs: Current state of art and future prospects. Front. Microbiol. 2021, 12, 636486. [Google Scholar] [CrossRef]
- Menon, A.; Lyng, J.G. Circular bioeconomy solutions: Driving anaerobic digestion of waste streams towards production of high value medium chain fatty acids. Rev. Environ. Sci. Bio/Technol. 2021, 20, 189–208. [Google Scholar] [CrossRef]
- (IEA). International Energy Agency. Sources of Methane Emissions. Available online: https://www.iea.org/data-and-statistics/charts/sources-of-methane-emissions-2 (accessed on 1 May 2023).
- (NOAA). National Oceanic and Atmospheric Administration. Trends in Atmospheric Methane (Global CH4 Monthly Means). Available online: https://gml.noaa.gov/ccgg/trends_ch4/ (accessed on 10 May 2023).
- (IEA). International Energy Agency. Total Methane Emissions and Methane Intensity of Production in Selected Oil and Gas Producers. 2021. Available online: https://www.iea.org/data-and-statistics/charts/total-methane-emissions-and-methane-intensity-of-production-in-selected-oil-and-gas-producers-2021 (accessed on 1 May 2023).
- Saunois, M.; Stavert, A.R.; Poulter, B.; Bousquet, P.; Canadell, J.G.; Jackson, R.B.; Raymond, P.A.; Dlugokencky, E.J.; Houweling, S.; Patra, P.K. The global methane budget 2000–2017. Earth Syst. Sci. Data 2020, 12, 1561–1623. [Google Scholar] [CrossRef]
- Li, D.-z.; Chen, L.; Liu, G.; Yuan, Z.-y.; Li, B.-f.; Zhang, X.; Wei, J.-q. Porous metal–organic frameworks for methane storage and capture: Status and challenges. New Carbon Mater. 2021, 36, 468–496. [Google Scholar] [CrossRef]
- Peng, Y.; Krungleviciute, V.; Eryazici, I.; Hupp, J.T.; Farha, O.K.; Yildirim, T. Methane storage in metal–organic frameworks: Current records, surprise findings, and challenges. J. Am. Chem. Soc. 2013, 135, 11887–11894. [Google Scholar] [PubMed] [Green Version]
- Sadiq, M.M.; Rubio-Martinez, M.; Zadehahmadi, F.; Suzuki, K.; Hill, M.R. Magnetic framework composites for low concentration methane capture. Ind. Eng. Chem. Res. 2018, 57, 6040–6047. [Google Scholar] [CrossRef]
- Bakhtyari, A.; Mofarahi, M. Pure and binary adsorption equilibria of methane and nitrogen on zeolite 5A. J. Chem. Eng. Data 2014, 59, 626–639. [Google Scholar] [CrossRef]
- Goodman, E.D.; Dai, S.; Yang, A.-C.; Wrasman, C.J.; Gallo, A.; Bare, S.R.; Hoffman, A.S.; Jaramillo, T.F.; Graham, G.W.; Pan, X. Uniform Pt/Pd bimetallic nanocrystals demonstrate platinum effect on palladium methane combustion activity and stability. ACS Catal. 2017, 7, 4372–4380. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, S.; Shan, J.-j.; Nguyen, L.; Zhan, S.; Gu, X.; Tao, F. In situ surface chemistries and catalytic performances of ceria doped with palladium, platinum, and rhodium in methane partial oxidation for the production of syngas. Acs Catal. 2013, 3, 2627–2639. [Google Scholar]
- Ross, M.O.; Rosenzweig, A.C. A tale of two methane monooxygenases. JBIC J. Biol. Inorg. Chem. 2017, 22, 307–319. [Google Scholar] [PubMed] [Green Version]
- Ross, M.O.; MacMillan, F.; Wang, J.; Nisthal, A.; Lawton, T.J.; Olafson, B.D.; Mayo, S.L.; Rosenzweig, A.C.; Hoffman, B.M. Particulate methane monooxygenase contains only mononuclear copper centers. Science 2019, 364, 566–570. [Google Scholar] [PubMed] [Green Version]
- Guerrero-Caballero, J.; Kane, T.; Haidar, N.; Jalowiecki-Duhamel, L.; Löfberg, A. Ni, Co, Fe supported on Ceria and Zr doped Ceria as oxygen carriers for chemical looping dry reforming of methane. Catal. Today 2019, 333, 251–258. [Google Scholar]
- Visvanathan, C.; Pokhrel, D.; Cheimchaisri, W.; Hettiaratchi, J.; Wu, J.S. Methanotrophic activities in tropical landfill cover soils: Effects of temperature, moisture content and methane concentration. Waste Manag. Res. 1999, 17, 313–323. [Google Scholar] [CrossRef]
- Nie, H.; Howe, J.Y.; Lachkov, P.T.; Chin, Y.-H.C. Chemical and structural dynamics of nanostructures in bimetallic Pt–Pd catalysts, their inhomogeneity, and their roles in methane oxidation. Acs Catal. 2019, 9, 5445–5461. [Google Scholar] [CrossRef]
- Bharadwaj, S.; Schmidt, L.D. Catalytic partial oxidation of natural gas to syngas. Fuel Process. Technol. 1995, 42, 109–127. [Google Scholar]
- Alrashed, W.; Chandra, R.; Abbott, T.; Lee, H.-S. Nitrite reduction using a membrane biofilm reactor (MBfR) in a hypoxic environment with dilute methane under low pressures. Sci. Total Environ. 2022, 841, 156757. [Google Scholar] [PubMed]
- Claridge, J.B.; York, A.P.; Brungs, A.J.; Marquez-Alvarez, C.; Sloan, J.; Tsang, S.C.; Green, M.L. New catalysts for the conversion of methane to synthesis gas: Molybdenum and tungsten carbide. J. Catal. 1998, 180, 85–100. [Google Scholar]
- Scheller, S.; Ermler, U.; Shima, S. Catabolic pathways and enzymes involved in anaerobic methane oxidation. Anaerob. Util. Hydrocarb. Oils Lipids 2020, 31–59. [Google Scholar] [CrossRef]
- Cai, X.; Hu, Y.H. Advances in catalytic conversion of methane and carbon dioxide to highly valuable products. Energy Sci. Eng. 2019, 7, 4–29. [Google Scholar]
- de Klerk, A. Engineering evaluation of direct methane to methanol conversion. Energy Sci. Eng. 2015, 3, 60–70. [Google Scholar] [CrossRef]
- Lawton, T.J.; Rosenzweig, A.C. Methane-oxidizing enzymes: An upstream problem in biological gas-to-liquids conversion. J. Am. Chem. Soc. 2016, 138, 9327–9340. [Google Scholar]
- Dummer, N.F.; Willock, D.J.; He, Q.; Howard, M.J.; Lewis, R.J.; Qi, G.; Taylor, S.H.; Xu, J.; Bethell, D.; Kiely, C.J.; et al. Methane Oxidation to Methanol. Chem. Rev. 2022, 123, 6359–6411. [Google Scholar] [CrossRef]
- Farrokhzadeh, H.; Hettiaratchi, J.P.A.; Jayasinghe, P.; Kumar, S. Aerated biofilters with multiple-level air injection configurations to enhance biological treatment of methane emissions. Bioresour. Technol. 2017, 239, 219–225. [Google Scholar] [CrossRef]
- Karthikeyan, O.P.; Saravanan, N.; Cirés, S.; Alvarez-Roa, C.; Razaghi, A.; Chidambarampadmavathy, K.; Velu, C.; Subashchandrabose, G.; Heimann, K. Culture scale-up and immobilisation of a mixed methanotrophic consortium for methane remediation in pilot-scale bio-filters. Environ. Technol. 2017, 38, 474–482. [Google Scholar] [CrossRef]
- Patricia, R.-R.; Gómez-Borraz, T.L.; Revah, S.; Morales, M. Methanotroph-Microalgae Co-Culture for Greenhouse Gas Mitigation: Effect of Initial Biomass Ratio and Methane Concentration. Chemosphere 2020, 259, 127418. [Google Scholar] [CrossRef]
- Lee, S.G.; Goo, J.H.; Kim, H.G.; Oh, J.-I.; Kim, Y.M.; Kim, S.W. Optimization of methanol biosynthesis from methane using Methylosinus trichosporium OB3b. Biotechnol. Lett. 2004, 26, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.-y.; Cui, J.-r.; Niu, J.-z.; Hua, S.-f.; Xia, C.-g.; Li, S.-b.; Zhu, L.-m. Production of methanol from methane by methanotrophic bacteria. Biocatal. Biotransform. 2004, 22, 225–229. [Google Scholar] [CrossRef]
- Ravi, M.; Ranocchiari, M.; van Bokhoven, J.A. The direct catalytic oxidation of methane to methanol—A critical assessment. Angew. Chem. Int. Ed. 2017, 56, 16464–16483. [Google Scholar]
- Alvarez-Galvan, M.; Mota, N.; Ojeda, M.; Rojas, S.; Navarro, R.; Fierro, J. Direct methane conversion routes to chemicals and fuels. Catal. Today 2011, 171, 15–23. [Google Scholar] [CrossRef]
- Labinger, J.A.; Bercaw, J.E. Understanding and exploiting C–H bond activation. Nature 2002, 417, 507–514. [Google Scholar]
- Lunsford, J.H. The catalytic oxidative coupling of methane. Angew. Chem. Int. Ed. Engl. 1995, 34, 970–980. [Google Scholar] [CrossRef]
- Zaera, F. Selectivity in hydrocarbon catalytic reforming: A surface chemistry perspective. Appl. Catal. A Gen. 2002, 229, 75–91. [Google Scholar]
- Puliyalil, H.; Jurković, D.L.; Dasireddy, V.D.; Likozar, B. A review of plasma-assisted catalytic conversion of gaseous carbon dioxide and methane into value-added platform chemicals and fuels. RSC Adv. 2018, 8, 27481–27508. [Google Scholar]
- Viswanathan, M.B.; Raman, D.R.; Rosentrater, K.A.; Shanks, B.H. A technoeconomic platform for early-stage process design and cost estimation of Joint fermentative catalytic bioprocessing. Processes 2020, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Nair, L.G.; Agrawal, K.; Verma, P. An insight into the principles of lignocellulosic biomass-based zero-waste biorefineries: A green leap towards imperishable energy-based future. Biotechnol. Genet. Eng. Rev. 2022, 38, 288–338. [Google Scholar] [CrossRef] [PubMed]
- Koo, C.W.; Rosenzweig, A.C. Biochemistry of aerobic biological methane oxidation. Chem. Soc. Rev. 2021, 50, 3424–3436. [Google Scholar] [CrossRef] [PubMed]
- Glass, J.B.; Orphan, V.J. Trace metal requirements for microbial enzymes involved in the production and consumption of methane and nitrous oxide. Front. Microbiol. 2012, 3, 61. [Google Scholar] [CrossRef] [Green Version]
- Mancinelli, R.L. The regulation of methane oxidation in soil. Annu. Rev. Microbiol. 1995, 49, 581–605. [Google Scholar] [CrossRef]
- Kapoore, R.V.; Padmaperuma, G.; Maneein, S.; Vaidyanathan, S. Co-culturing microbial consortia: Approaches for applications in biomanufacturing and bioprocessing. Crit. Rev. Biotechnol. 2022, 42, 46–72. [Google Scholar] [CrossRef] [PubMed]
- Che, S.; Men, Y. Synthetic microbial consortia for biosynthesis and biodegradation: Promises and challenges. J. Ind. Microbiol. Biotechnol. 2019, 46, 1343–1358. [Google Scholar] [CrossRef] [PubMed]
- Reay, D.S.; Nedwell, D.B. Methane oxidation in temperate soils: Effects of inorganic N. Soil Biol. Biochem. 2004, 36, 2059–2065. [Google Scholar] [CrossRef]
- Smith, T.; Murrell, J. Methanotrophy/methane oxidation. Encycl. Microbiol. 2009, 3, 293–298. [Google Scholar]
- Tikhonova, E.N.; Kravchenko, I.K. Activity and diversity of aerobic methanotrophs in thermal springs of the russian far east. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1–30. [Google Scholar]
- Nazaries, L.; Murrell, J.C.; Millard, P.; Baggs, L.; Singh, B.K. Methane, microbes and models: Fundamental understanding of the soil methane cycle for future predictions. Environ. Microbiol. 2013, 15, 2395–2417. [Google Scholar] [CrossRef]
- Larkin, D.; Caldwell, T.; Lobban, L.; Mallinson, R.G. Oxygen pathways and carbon dioxide utilization in methane partial oxidation in ambient temperature electric discharges. Energy Fuels 1998, 12, 740–744. [Google Scholar] [CrossRef]
- Meraz, J.L.; Dubrawski, K.L.; El Abbadi, S.H.; Choo, K.-H.; Criddle, C.S. Membrane and fluid contactors for safe and efficient methane delivery in methanotrophic bioreactors. J. Environ. Eng. 2020, 146, 03120006. [Google Scholar] [CrossRef]
- Hwee, N. Methane Remediation Using Biocatalysts in Batch Gas-Solid Mass Transfer System Challenges and Prospects. 2022. Available online: https://www.osti.gov/servlets/purl/1860693 (accessed on 1 May 2023).
- Krishna, R.; Sie, S. Strategies for multiphase reactor selection. Chem. Eng. Sci. 1994, 49, 4029–4065. [Google Scholar] [CrossRef]
- Estrada, J.M.; Lebrero, R.; Quijano, G.; Pérez, R.; Figueroa-González, I.; García-Encina, P.A.; Muñoz, R. Methane abatement in a gas-recycling biotrickling filter: Evaluating innovative operational strategies to overcome mass transfer limitations. Chem. Eng. J. 2014, 253, 385–393. [Google Scholar] [CrossRef]
- Amaral, P.F.; Freire, M.G.; Rocha-Leão, M.H.M.; Marrucho, I.M.; Coutinho, J.A.; Coelho, M.A.Z. Optimization of oxygen mass transfer in a multiphase bioreactor with perfluorodecalin as a second liquid phase. Biotechnol. Bioeng. 2008, 99, 588–598. [Google Scholar] [CrossRef]
- Stone, K.A.; Hilliard, M.V.; He, Q.P.; Wang, J. A mini review on bioreactor configurations and gas transfer enhancements for biochemical methane conversion. Biochem. Eng. J. 2017, 128, 83–92. [Google Scholar] [CrossRef]
- Ramezani, M.; Legg, M.J.; Haghighat, A.; Li, Z.; Vigil, R.D.; Olsen, M.G. Experimental investigation of the effect of ethyl alcohol surfactant on oxygen mass transfer and bubble size distribution in an air-water multiphase Taylor-Couette vortex bioreactor. Chem. Eng. J. 2017, 319, 288–296. [Google Scholar] [CrossRef]
- Dong, T.; Knoshaug, E.P.; Pienkos, P.T.; Laurens, L.M. Lipid recovery from wet oleaginous microbial biomass for biofuel production: A critical review. Appl. Energy 2016, 177, 879–895. [Google Scholar] [CrossRef] [Green Version]
- Antoni, D.; Burckel, H.; Josset, E.; Noel, G. Three-dimensional cell culture: A breakthrough in vivo. Int. J. Mol. Sci. 2015, 16, 5517–5527. [Google Scholar] [CrossRef]
- Taylor, A.; Molzahn, P.; Bushnell, T.; Cheney, C.; LaJeunesse, M.; Azizian, M.; Semprini, L. Immobilization of Methylosinus trichosporium OB3b for methanol production. J. Ind. Microbiol. Biotechnol. 2018, 45, 201–211. [Google Scholar] [CrossRef]
- Tiwari, H.; Sonwani, R.K.; Singh, R.S. Bioremediation of dyes: A brief review of bioreactor performance. Environ. Technol. Rev. 2023, 12, 83–128. [Google Scholar] [CrossRef]
- Datsevich, L.B.; Datsevich, L.B. Analysis of Conventional Industrial Processes. Conv. Three-Phase Fixed-Bed Technol. Anal. Crit. 2012, 7, 33–62. [Google Scholar]
- Devarapalli, M.; Lewis, R.S.; Atiyeh, H.K. Continuous ethanol production from synthesis gas by Clostridium ragsdalei in a trickle-bed reactor. Fermentation 2017, 3, 23. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.; Atiyeh, H.K.; Stevenson, B.S.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Continuous syngas fermentation for the production of ethanol, n-propanol and n-butanol. Bioresour. Technol. 2014, 151, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Datar, R.P.; Shenkman, R.M.; Cateni, B.G.; Huhnke, R.L.; Lewis, R.S. Fermentation of biomass-generated producer gas to ethanol. Biotechnol. Bioeng. 2004, 86, 587–594. [Google Scholar] [CrossRef]
- Shen, Y.; Brown, R.; Wen, Z. Enhancing mass transfer and ethanol production in syngas fermentation of Clostridium carboxidivorans P7 through a monolithic biofilm reactor. Appl. Energy 2014, 136, 68–76. [Google Scholar] [CrossRef]
- Martin, M.E.; Richter, H.; Saha, S.; Angenent, L.T. Traits of selected Clostridium strains for syngas fermentation to ethanol. Biotechnol. Bioeng. 2016, 113, 531–539. [Google Scholar] [CrossRef]
- Orgill, J.J.; Atiyeh, H.K.; Devarapalli, M.; Phillips, J.R.; Lewis, R.S.; Huhnke, R.L. A comparison of mass transfer coefficients between trickle-bed, hollow fiber membrane and stirred tank reactors. Bioresour. Technol. 2013, 133, 340–346. [Google Scholar] [CrossRef]
- Gunes, B. A critical review on biofilm-based reactor systems for enhanced syngas fermentation processes. Renew. Sustain. Energy Rev. 2021, 143, 110950. [Google Scholar] [CrossRef]
- Havran, V.; Dudukovic, M.P.; Lo, C.S. Conversion of methane and carbon dioxide to higher value products. Ind. Eng. Chem. Res. 2011, 50, 7089–7100. [Google Scholar] [CrossRef]
- Liotta, L. Catalytic oxidation of volatile organic compounds on supported noble metals. Appl. Catal. B Environ. 2010, 100, 403–412. [Google Scholar] [CrossRef]
- Chen, Y.; Mu, X.; Luo, X.; Shi, K.; Yang, G.; Wu, T. Catalytic conversion of methane at low temperatures: A critical review. Energy Technol. 2020, 8, 1900750. [Google Scholar] [CrossRef]
- Hristov, A.N.; Oh, J.; Giallongo, F.; Frederick, T.W.; Harper, M.T.; Weeks, H.L.; Branco, A.F.; Moate, P.J.; Deighton, M.H.; Williams, S.R.O. An inhibitor persistently decreased enteric methane emission from dairy cows with no negative effect on milk production. Proc. Natl. Acad. Sci. USA 2015, 112, 10663–10668. [Google Scholar] [CrossRef] [PubMed]
- Boucher, O.; Folberth, G.A. New directions: Atmospheric methane removal as a way to mitigate climate change? Atmos. Environ. 2010, 44, 3343–3345. [Google Scholar] [CrossRef]
- Perot, G.; Guisnet, M. Advantages and disadvantages of zeolites as catalysts in organic chemistry. J. Mol. Catal. 1990, 61, 173–196. [Google Scholar] [CrossRef]
- Wang, P.; Sun, Q.; Zhang, Y.; Cao, J. Effective removal of methane using nano-sized zeolite 4A synthesized from kaolin. Inorg. Chem. Commun. 2020, 111, 107639. [Google Scholar] [CrossRef]
- Beznis, N.V.; van Laak, A.N.C.; Weckhuysen, B.M.; Bitter, J.H. Oxidation of methane to methanol and formaldehyde over Co–ZSM-5 molecular sieves: Tuning the reactivity and selectivity by alkaline and acid treatments of the zeolite ZSM-5 agglomerates. Microporous Mesoporous Mater. 2011, 138, 176–183. [Google Scholar] [CrossRef]
- Ming, T.; Li, W.; Yuan, Q.; Davies, P.; De Richter, R.; Peng, C.; Deng, Q.; Yuan, Y.; Caillol, S.; Zhou, N. Perspectives on removal of atmospheric methane. Adv. Appl. Energy 2022, 5, 100085. [Google Scholar] [CrossRef]
- Ma, C.; Dalby, F.R.; Feilberg, A.; Jacobsen, B.H.; Petersen, S.O. Low-Dose Acidification as a Methane Mitigation Strategy for Manure Management. ACS Agric. Sci. Technol. 2022, 2, 437–442. [Google Scholar] [CrossRef]
- Sturtz, T.M.; Jenkins, P.T.; de Richter, R. Environmental Impact Modeling for a Small-Scale Field Test of Methane Removal by Iron Salt Aerosols. Sustainability 2022, 14, 14060. [Google Scholar] [CrossRef]
- Oeste, F.D.; de Richter, R.; Ming, T.; Caillol, S. Climate engineering by mimicking natural dust climate control: The iron salt aerosol method. Earth Syst. Dyn. 2017, 8, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Ming, T.; de Richter, R.; Oeste, F.D.; Tulip, R.; Caillol, S. A nature-based negative emissions technology able to remove atmospheric methane and other greenhouse gases. Atmos. Pollut. Res. 2021, 12, 101035. [Google Scholar] [CrossRef]
- Stolaroff, J.K.; Bhattacharyya, S.; Smith, C.A.; Bourcier, W.L.; Cameron-Smith, P.J.; Aines, R.D. Review of methane mitigation technologies with application to rapid release of methane from the Arctic. Environ. Sci. Technol. 2012, 46, 6455–6469. [Google Scholar] [CrossRef]
- Lackner, K.S. Practical constraints on atmospheric methane removal. Nat. Sustain. 2020, 3, 357. [Google Scholar] [CrossRef] [Green Version]
- Post, M.F. Diffusion in zeolite molecular sieves. In Studies in Surface Science and Catalysis; Elsevier: Amsterdam, The Netherlands, 1991; Volume 58, pp. 391–443. [Google Scholar]
- Jiang, L.; Liu, W.; Wang, R.; Gonzalez-Diaz, A.; Rojas-Michaga, M.; Michailos, S.; Pourkashanian, M.; Zhang, X.; Font-Palma, C. Sorption direct air capture with CO2 utilization. Prog. Energy Combust. Sci. 2023, 95, 101069. [Google Scholar] [CrossRef]
- Su, S.; Beath, A.; Guo, H.; Mallett, C. An assessment of mine methane mitigation and utilisation technologies. Prog. Energy Combust. Sci. 2005, 31, 123–170. [Google Scholar] [CrossRef]
- Shih, C.F.; Zhang, T.; Li, J.; Bai, C. Powering the future with liquid sunshine. Joule 2018, 2, 1925–1949. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, M.; Vanshika; Kumar, A.; Chauhan, D.; Jain, A.K. Renewable Feedstocks for Biofuels. Biofuel Extr. Technol. 2023, 151–176. [Google Scholar] [CrossRef]
- Srivastava, R.K. Biological Approaches for Advanced Fuels Development from Biological/Plant Residues or Wastes. In Biotechnology for Waste Biomass Utilization; Apple Academic Press: Cambridge, UK, 2022; pp. 129–148. [Google Scholar]
- Bicer, Y.; Khalid, F. Life cycle environmental impact comparison of solid oxide fuel cells fueled by natural gas, hydrogen, ammonia and methanol for combined heat and power generation. Int. J. Hydrogen Energy 2020, 45, 3670–3685. [Google Scholar] [CrossRef]
- Mohammadi, R.; Harandi, M.H. Methanol to Olefin (MTO) Value Chain Management. New Appl. Stud. Manag. Econ. Account. 2023, 6, 7–17. [Google Scholar]
- Galusnyak, S.C.; Petrescu, L.; Chisalita, D.A.; Cormos, C.-C. Life cycle assessment of methanol production and conversion into various chemical intermediates and products. Energy 2022, 259, 124784. [Google Scholar] [CrossRef]
- Sarangi, P.K.; Nanda, S.; Vo, D.-V.N. Technological advancements in the production and application of biomethanol. Biorefinery Altern. Resour. Target. Green Fuels Platf. Chem. 2020, 127–139. [Google Scholar] [CrossRef]
- Oklu, N.K.; Matsinha, L.C.; Makhubela, B.C. Bio-solvents: Synthesis, industrial production and applications. Solvents Ion. Liq. Solvent Eff. 2019, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Karmakar, T.; Bjelobrk, Z.; Gong, J.; Parrinello, M. Solvent-mediated morphology selection of the active pharmaceutical ingredient isoniazid: Experimental and simulation studies. Chem. Eng. Sci. 2019, 204, 320–328. [Google Scholar] [CrossRef]
- Liu, K.; Wang, X.; Yan, Y. Network analysis of industrial symbiosis in chemical industrial parks: A case study of Nanjing Jiangbei new materials high-tech park. Sustainability 2022, 14, 1381. [Google Scholar] [CrossRef]
- Markets and Markets. Methanol Market. In Methanol Market by Feedstock; Markets and Markets Publisher: Maharashtra, India, 2023. [Google Scholar]
- Newswire, C.P.R. Global Methanol Industry. In Global Methanol Market to Reach 99.8 Million Metric Tons by 2030; Global Industry Analysts Publisher: San Jose, CA, USA, 2023; p. 523. [Google Scholar]
- Agarwal, N.; Freakley, S.J.; McVicker, R.U.; Althahban, S.M.; Dimitratos, N.; He, Q.; Morgan, D.J.; Jenkins, R.L.; Willock, D.J.; Taylor, S.H. Aqueous Au-Pd colloids catalyze selective CH4 oxidation to CH3OH with O2 under mild conditions. Science 2017, 358, 223–227. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Wang, L.; Zuidema, E.; Mondal, K.; Zhang, M.; Zhang, J.; Wang, C.; Meng, X.; Yang, H.; Mesters, C. Hydrophobic zeolite modification for in situ peroxide formation in methane oxidation to methanol. Science 2020, 367, 193–197. [Google Scholar] [CrossRef]
- Grundner, S.; Markovits, M.A.; Li, G.; Tromp, M.; Pidko, E.A.; Hensen, E.J.; Jentys, A.; Sanchez-Sanchez, M.; Lercher, J.A. Single-site trinuclear copper oxygen clusters in mordenite for selective conversion of methane to methanol. Nat. Commun. 2015, 6, 7546. [Google Scholar] [CrossRef] [Green Version]
- Narsimhan, K.; Iyoki, K.; Dinh, K.; Román-Leshkov, Y. Catalytic oxidation of methane into methanol over copper-exchanged zeolites with oxygen at low temperature. ACS Cent. Sci. 2016, 2, 424–429. [Google Scholar] [CrossRef] [Green Version]
- Bi, W.; Tang, Y.; Li, X.; Dai, C.; Song, C.; Guo, X.; Ma, X. One-step direct conversion of methane to methanol with water in non-thermal plasma. Commun. Chem. 2022, 5, 124. [Google Scholar] [CrossRef]
- Koo, C.W. Structural Studies of Particulate Methane Monooxygenase in a Native Lipid Bilayer. Ph.D. Thesis, Northwestern University, Evanston, IL, USA, 2022. [Google Scholar]
- Murrell, J.C.; Gilbert, B.; McDonald, I.R. Molecular biology and regulation of methane monooxygenase. Arch. Microbiol. 2000, 173, 325–332. [Google Scholar] [CrossRef]
- Semrau, J.D.; DiSpirito, A.A.; Gu, W.; Yoon, S. Metals and methanotrophy. Appl. Environ. Microbiol. 2018, 84, e02289-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begonja, A.; Hrsak, D. Effect of growth conditions on the expression of soluble methane monooxygenase. Food Technol. Biotechnol. 2001, 39, 29–36. [Google Scholar]
- Chan, S.I.; Chen, K.H.-C.; Yu, S.S.-F.; Chen, C.-L.; Kuo, S.S.-J. Toward delineating the structure and function of the particulate methane monooxygenase from methanotrophic bacteria. Biochemistry 2004, 43, 4421–4430. [Google Scholar] [CrossRef]
- Nielsen, A.K.; Gerdes, K.; Murrell, J.C. Copper-dependent reciprocal transcriptional regulation of methane monooxygenase genes in Methylococcus capsulatus and Methylosinus trichosporium. Mol. Microbiol. 1997, 25, 399–409. [Google Scholar] [CrossRef]
- Srinivas, V.; Banerjee, R.; Lebrette, H.; Jones, J.C.; Aurelius, O.; Kim, I.-S.; Pham, C.C.; Gul, S.; Sutherlin, K.D.; Bhowmick, A. High-resolution XFEL structure of the soluble methane monooxygenase hydroxylase complex with its regulatory component at ambient temperature in two oxidation states. J. Am. Chem. Soc. 2020, 142, 14249–14266. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Carey, J.N.; Semrau, J.D. Feasibility of atmospheric methane removal using methanotrophic biotrickling filters. Appl. Microbiol. Biotechnol. 2009, 83, 949–956. [Google Scholar] [CrossRef]
- Sun, M.-T.; Zhao, Y.-Z.; Yang, Z.-M.; Shi, X.-S.; Wang, L.; Dai, M.; Wang, F.; Guo, R.-B. Methane Elimination Using Biofiltration Packed With Fly Ash Ceramsite as Support Material. Front. Bioeng. Biotechnol. 2020, 8, 351. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Smith, A.L. Methane-driven microbial fuel cells recover energy and mitigate dissolved methane emissions from anaerobic effluents. Environ. Sci. Water Res. Technol. 2018, 4, 67–79. [Google Scholar] [CrossRef]
- Jawaharraj, K.; Dhiman, S.S.; Bedwell, S.; Vemuri, B.; Islam, J.; Sani, R.K.; Gadhamshetty, V. Electricity from methane by Methylococcus capsulatus (Bath) and Methylosinus trichosporium OB3b. Bioresour. Technol. 2021, 321, 124398. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Kim, T. Development of a novel methanotrophic process with the helper micro-organism Hyphomicrobium sp. NM3. J. Appl. Microbiol. 2019, 126, 534–544. [Google Scholar] [CrossRef]
- Chidambarampadmavathy, K.; Karthikeyan, O.P.; Huerlimann, R.; Maes, G.E.; Heimann, K. Response of mixed methanotrophic consortia to different methane to oxygen ratios. Waste Manag. 2017, 61, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Cruz, S.; Vaksmaa, A.; Horn, M.A.; Niemann, H.; Pijuan, M.; Ho, A. Methanotrophs: Discoveries, environmental relevance, and a perspective on current and future applications. Front. Microbiol. 2021, 12, 678057. [Google Scholar] [CrossRef] [PubMed]
- Mardina, P.; Li, J.; Patel, S.K.; Kim, I.-W.; Lee, J.-K.; Selvaraj, C. Potential of immobilized whole-cell Methylocella tundrae as a biocatalyst for methanol production from methane. J. Microbiol. Biotechnol. 2016, 26, 1234–1241. [Google Scholar] [CrossRef] [Green Version]
- Razumovsky, S.; Efremenko, E.; Makhlis, T.; Senko, O.; Bikhovsky, M.Y.; Podmaster’Ev, V.; Varfolomeev, S. Effect of immobilization on the main dynamic characteristics of the enzymatic oxidation of methane to methanol by bacteria Methylosinus sporium B-2121. Russ. Chem. Bull. 2008, 57, 1633–1636. [Google Scholar] [CrossRef]
- Chistoserdova, L. Modularity of methylotrophy, revisited. Environ. Microbiol. 2011, 13, 2603–2622. [Google Scholar] [CrossRef]
- Smith, T.; Dalton, H. Biocatalysis by methane monooxygenase and its implications for the petroleum industry. Stud. Surf. Sci. Catal. 2004, 151, 177–192. [Google Scholar]
- Smith, S.M.; Balasubramanian, R.; Rosenzweig, A.C. Metal reconstitution of particulate methane monooxygenase and heterologous expression of the pmoB subunit. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 495, pp. 195–210. [Google Scholar]
- Lloyd, J.S.; De Marco, P.; Dalton, H.; Murrell, J.C. Heterologous expression of soluble methane monooxygenase genes in methanotrophs containing only particulate methane monooxygenase. Arch. Microbiol. 1999, 171, 364–370. [Google Scholar] [CrossRef]
- La, H.; Hettiaratchi, J.P.A.; Achari, G.; Dunfield, P.F. Biofiltration of methane. Bioresour. Technol. 2018, 268, 759–772. [Google Scholar] [CrossRef]
- Dalcin Martins, P.; de Jong, A.; Lenstra, W.K.; van Helmond, N.A.; Slomp, C.P.; Jetten, M.S.; Welte, C.U.; Rasigraf, O. Enrichment of novel Verrucomicrobia, Bacteroidetes, and Krumholzibacteria in an oxygen-limited methane-and iron-fed bioreactor inoculated with Bothnian Sea sediments. MicrobiologyOpen 2021, 10, e1175. [Google Scholar] [CrossRef]
- Streese, J.; Stegmann, R. Potentials and limitations of biofilters for methane oxidation. In Proceedings of the Sardinia ‘05, International Waste Management and Landfill Symposium, Cagliari, Italy, 3–7 October 2005. [Google Scholar]
- Jawad, J.; Khalil, M.J.; Sengar, A.K.; Zaidi, S.J. Experimental analysis and modeling of the methane degradation in a three stage biofilter using composted sawdust as packing media. J. Environ. Manag. 2021, 286, 112214. [Google Scholar] [CrossRef]
- Streese, J.; Stegmann, R. Design of biofilters for methane oxidation. In Proceedings of the Sardinia 2003, Ninth International Waste Management and Landfill Symposium, Cagliari, Italy, 6–10 October 2003; pp. 6–10. [Google Scholar]
- Cáceres, M.; Dorado, A.D.; Gentina, J.C.; Aroca, G. Oxidation of methane in biotrickling filters inoculated with methanotrophic bacteria. Environ. Sci. Pollut. Res. 2017, 24, 25702–25712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, E.M.F.; Duarte, F.V.; Vieira, J.P.R.; Melo, V.M.; Souza, C.L.; Araújo, J.C.; Chernicharo, C.A.L. The use of novel packing material for improving methane oxidation in biofilters. J. Environ. Manag. 2016, 182, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Veillette, M.; Viens, P.; Ramirez, A.A.; Brzezinski, R.; Heitz, M. Effect of ammonium concentration on microbial population and performance of a biofilter treating air polluted with methane. Chem. Eng. J. 2011, 171, 1114–1123. [Google Scholar] [CrossRef]
- Ren, Z.J. Microbial fuel cells: Running on gas. Nat. Energy 2017, 2, 17093. [Google Scholar] [CrossRef]
- Song, C. Fuel processing for low-temperature and high-temperature fuel cells: Challenges, and opportunities for sustainable development in the 21st century. Catal. Today 2002, 77, 17–49. [Google Scholar] [CrossRef]
- Pandit, S.; Savla, N.; Sonawane, J.M.; Sani, A.M.d.; Gupta, P.K.; Mathuriya, A.S.; Rai, A.K.; Jadhav, D.A.; Jung, S.P.; Prasad, R. Agricultural waste and wastewater as feedstock for bioelectricity generation using microbial fuel cells: Recent advances. Fermentation 2021, 7, 169. [Google Scholar] [CrossRef]
- Li, W.-W.; Yu, H.-Q.; He, Z. Towards sustainable wastewater treatment by using microbial fuel cells-centered technologies. Energy Environ. Sci. 2014, 7, 911–924. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Rabiee, H.; Frank, J.; Cai, C.; Stark, T.; Virdis, B.; Yuan, Z.; Hu, S. Enhancing methane oxidation in a bioelectrochemical membrane reactor using a soluble electron mediator. Biotechnol. Biofuels 2020, 13, 173. [Google Scholar] [CrossRef]
- Myung, J.; Saikaly, P.E.; Logan, B.E. A two-staged system to generate electricity in microbial fuel cells using methane. Chem. Eng. J. 2018, 352, 262–267. [Google Scholar] [CrossRef]
- Xing, Z.; Zhao, T.; Zhang, L.; Gao, Y.; Liu, S.; Yang, X. Effects of copper on expression of methane monooxygenases, trichloroethylene degradation, and community structure in methanotrophic consortia. Eng. Life Sci. 2018, 18, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Wang, X.; Zhou, J.; Zhang, Y. Enrichments of methanotrophic–heterotrophic cultures with high poly-β-hydroxybutyrate (PHB) accumulation capacities. J. Environ. Sci. 2018, 65, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-W.; Keeney, D.R.; Lim, D.-H.; Dispirito, A.A.; Semrau, J.D. Mixed pollutant degradation by Methylosinus trichosporium OB3b expressing either soluble or particulate methane monooxygenase: Can the tortoise beat the hare? Appl. Environ. Microbiol. 2006, 72, 7503–7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lontoh, S.; Semrau, J.D. Methane and trichloroethylene degradation by Methylosinus trichosporium OB3b expressing particulate methane monooxygenase. Appl. Environ. Microbiol. 1998, 64, 1106–1114. [Google Scholar] [CrossRef] [PubMed]
- Samanta, D.; Govil, T.; Saxena, P.; Gadhamshetty, V.; Krumholz, L.R.; Salem, D.R.; Sani, R.K. Enhancement of Methane Catalysis Rates in Methylosinus trichosporium OB3b. Biomolecules 2022, 12, 560. [Google Scholar] [CrossRef]
- Yu, S.S.-F.; Chen, K.H.-C.; Tseng, M.Y.-H.; Wang, Y.-S.; Tseng, C.-F.; Chen, Y.-J.; Huang, D.-S.; Chan, S.I. Production of high-quality particulate methane monooxygenase in high yields from Methylococcus capsulatus (Bath) with a hollow-fiber membrane bioreactor. J. Bacteriol. 2003, 185, 5915–5924. [Google Scholar] [CrossRef] [Green Version]
- Jahng, D.; Wood, T.K. Trichloroethylene and chloroform degradation by a recombinant pseudomonad expressing soluble methane monooxygenase from Methylosinus trichosporium OB3b. Appl. Environ. Microbiol. 1994, 60, 2473–2482. [Google Scholar] [CrossRef]
- Jahng, D.; Kim, C.S.; Hanson, R.S.; Wood, T.K. Optimization of trichloroethylene degradation using soluble methane monooxygenase of Methylosinus trichosporium OB3b expressed in recombinant bacteria. Biotechnol. Bioeng. 1996, 51, 349–359. [Google Scholar] [CrossRef]
- Gou, Z.; Xing, X.-H.; Luo, M.; Jiang, H.; Han, B.; Wu, H.; Wang, L.; Zhang, F. Functional expression of the particulate methane mono-oxygenase gene in recombinant Rhodococcus erythropolis. FEMS Microbiol. Lett. 2006, 263, 136–141. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.K.; Jeong, J.-H.; Mehariya, S.; Otari, S.V.; Madan, B.; Haw, J.R.; Lee, J.-K.; Zhang, L.; Kim, I.-W. Production of methanol from methane by encapsulated Methylosinus sporium. J. Microbiol. Biotechnol. 2016, 26, 2098–2105. [Google Scholar] [CrossRef] [Green Version]
- Van der Ha, D.; Bundervoet, B.; Verstraete, W.; Boon, N. A sustainable, carbon neutral methane oxidation by a partnership of methane oxidizing communities and microalgae. Water Res. 2011, 45, 2845–2854. [Google Scholar] [CrossRef]
- Tays, C. Growth Characterization and Transcriptomics of Methanotrophic Bacteria as Effected by Carbon and Nitrogen Sources. Ph.D. Thesis, University of Alberta, Edmonton, AB, Canada, 2019. [Google Scholar]
- Akberdin, I.R.; Thompson, M.; Hamilton, R.; Desai, N.; Alexander, D.; Henard, C.A.; Guarnieri, M.T.; Kalyuzhnaya, M.G. Methane utilization in Methylomicrobium alcaliphilum 20ZR: A systems approach. Sci. Rep. 2018, 8, 2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Parameters | Chemical Methane Oxidation | Biological Methane Oxidation |
|---|---|---|
| Catalyst | The most common catalysts are palladium (Pd) and platinum (Pt) and are associated with high catalytic activity and selectivity towards methane oxidation [34,35]. | The catalysts are the enzymes methane monooxygenases (MMOs) that consist of soluble MMOs (sMMOs) and particulate MMOs (pMMOs) [36,37]. |
| Temperature | Requires high temperatures (between 500 °C and 800 °C) for effective conversion [38]. | Requires comparatively lower temperatures (20 °C and 30 °C) [39]. |
| Pressure | The operating pressure depends on the catalyst used. For Pd catalysts, the optimal pressure range is between 0.1 and 10 atm, while for Pt catalysts, between 1 and 10 atm is used [40,41] | The operating pressure depends on the organism used. Some methanotrophic bacteria can grow at atmospheric pressure, while others can grow at pressures as high as 20 atm [42]. |
| Oxidant | The oxidants used are typically oxygen (O2) or air. The amount of oxidant used depends on the catalyst used and the desired methane conversion percentage [43]. | The oxidants used are typically molecular oxygen (O2) or nitrate (NO3-). The amount of oxidant used depends on the organism used and the desired methane conversion percentage [44]. |
| Methane conversion (%) | The conversion depends on the catalyst used, temperature, pressure, and oxidant. The highest conversion percentage for Pd catalysts is 80% at 700 °C and 5 atm pressure, while for Pt catalysts, is 60% at 750 °C and 1 atm pressure [41,45]. The carbon efficiency for direct methane to methanol conversion is around 35%, roughly half that of indirect conversion methods [46]. | The conversion percentage depends on the organism used, temperature, pressure, and oxidant. The highest methane uptake for sMMOs is 22 mmol gDCW−1 h−1, while for pMMOs, it is 9.0 mmol gDCW−1 h−1 [47]. The turnover frequency of sMMO is reported to be 95 molmethanol molFe–1 h–1 with a 100% selectivity of methane over methanol [48]. The methane conversion efficiencies are reported up to 85–97% [49,50]. |
| Selectivity | Further oxidation of methane oxidation product, methanol, leads to the formation of unwanted products such as formaldehyde, lowering the selectivity of desired to undesired products [51]. | Further methanol oxidation within the cell system can be limited by inhibiting the subsequent enzyme, methanol dehydrogenase, thus improved selectivity can be achieved [52,53]. |
| Methods/Strategies | Description | Advantages | Disadvantages |
|---|---|---|---|
| Acidification [100] | Treatment of slurry wastes with certain concentration of acids (e.g., concentrated sulfuric acid). | Reduction in CH4 emission is 46–96%. Below pH 6, methanogens get inhibited. | Time and cost inefficient. Only low dose acidification is cost-effective. |
| Iron-salt aerosols [99,101,102,103] | Chemical mimicry of natural reactions associated with mineral dust particles (contains iron) in the atmosphere. Uses Fenton and photo-Fenton reactions. | Releases iron salt aerosols in the lower troposphere, creating a sink for chlorine and hydroxyl ions. No negative impact on stratospheric ozone. | Cannot be deployed in urban areas due to activation of chlorine ions leads to tropospheric ozone layer depletion. |
| Zeolite minerals [96,97,105] | Zeolite sorbents followed by catalytic destruction. | High thermostability, less corrosion, easy set up continuous process, and no environmental waste. | Deactivation by irreversible adsorption and steric blockage due to secondary chemicals. |
| Molecular sieves [106,107] | The mechanisms are governed by Langmuir and Freundlich adsorption and diffusion. | Structural modifications can improve efficiency. | Involves a sizable quantity that is not feasible from an economic standpoint. Lower CH4 concentrations in the air increases the energy requirements for capturing and circulating the air through adsorbers. |
| Cryogenic separation [108] | Involves compression of methane to increase the pressure until it liquefies. | Converts coal mine methane to cold refrigerant stream. | Blockage of piping and cost inefficient. |
| Methods/Strategies | Description | Advantages | Disadvantages |
|---|---|---|---|
| Bio-trickling filter 1 [133] | Air stream containing pollutants is passed through the filter, and as it comes into contact with the microorganisms in the filter, they consume or oxidize the pollutants. | A total of 5 to 40 tons of CH4 could be mitigated per biofilter per year near landfills and concentrated animal feeding operations. | Operates at a higher airflow rate and thus requires enough CH4 concentration (>500 ppmv) for cell viability. |
| Biofiltration Packed with fly ash [134] | Methanotrophic cultures are inoculated on biofilters packed with supportive materials such as carbon, perlite, etc. | Methane elimination capacity of biofilters with fly ash ceramic packing is 4.628 g h−1 m−3), which is higher than other biofilters. | Membrane fouling and poor surface biocompatibility of support material |
| Microbial fuel cells 1 [135,136] | Methane-oxidizers donate electrons directly to anode during the oxidation of methane to organic products. | Produces energy (e.g., electricity) from methane, which is sourced from anaerobic membrane bioreactors. | Occasional irregularities involve biofouling at cathode that leads to decrease in voltage. |
| Co-cultivation and symbiotic growth 1 [137,138,139] | Heterotrophic interactions mainly observed with cyanobacteria | Enhanced methanotrophic growth and high cell density. Potential for simultaneous production of valuable products. | Challenging to maintain stability and balance between microorganisms. |
| Encapsulated biofilm technology [140,141] | Adherence of methanotrophic pure culture or consortia on suitable attaching surfaces | Increased surface area for methanotrophic growth, resulting in enhanced gas to liquid mass transfer | Limited diffusion of methane into biofilm. Potential for biofilm detachment and clogging of carriers |
| Genetic engineering [142,143,144,145] | Engineering the catalytic domain of MMOs | Targeted modification and has potential use for substrate and product selectivity | Technical challenges with regard to availability of genetic tools. May be time-consuming and expensive. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samanta, D.; Sani, R.K. Methane Oxidation via Chemical and Biological Methods: Challenges and Solutions. Methane 2023, 2, 279-303. https://doi.org/10.3390/methane2030019
Samanta D, Sani RK. Methane Oxidation via Chemical and Biological Methods: Challenges and Solutions. Methane. 2023; 2(3):279-303. https://doi.org/10.3390/methane2030019
Chicago/Turabian StyleSamanta, Dipayan, and Rajesh K. Sani. 2023. "Methane Oxidation via Chemical and Biological Methods: Challenges and Solutions" Methane 2, no. 3: 279-303. https://doi.org/10.3390/methane2030019