
Peptide Co-Agonists for Combined Activation of the APJ and GLP-1 Receptors with Insulinotropic and Satiety Actions Show Potential for Alleviation of Metabolic Dysfunction in Type 2 Diabetes †


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
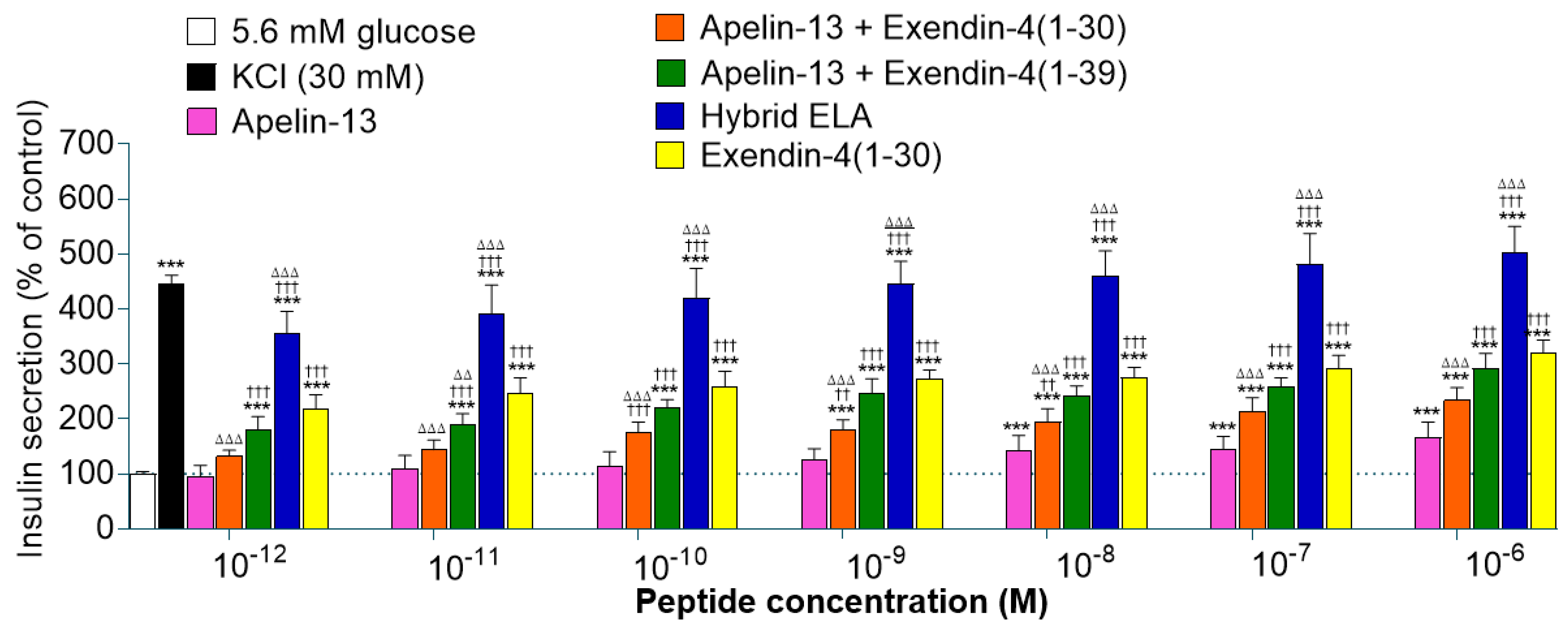
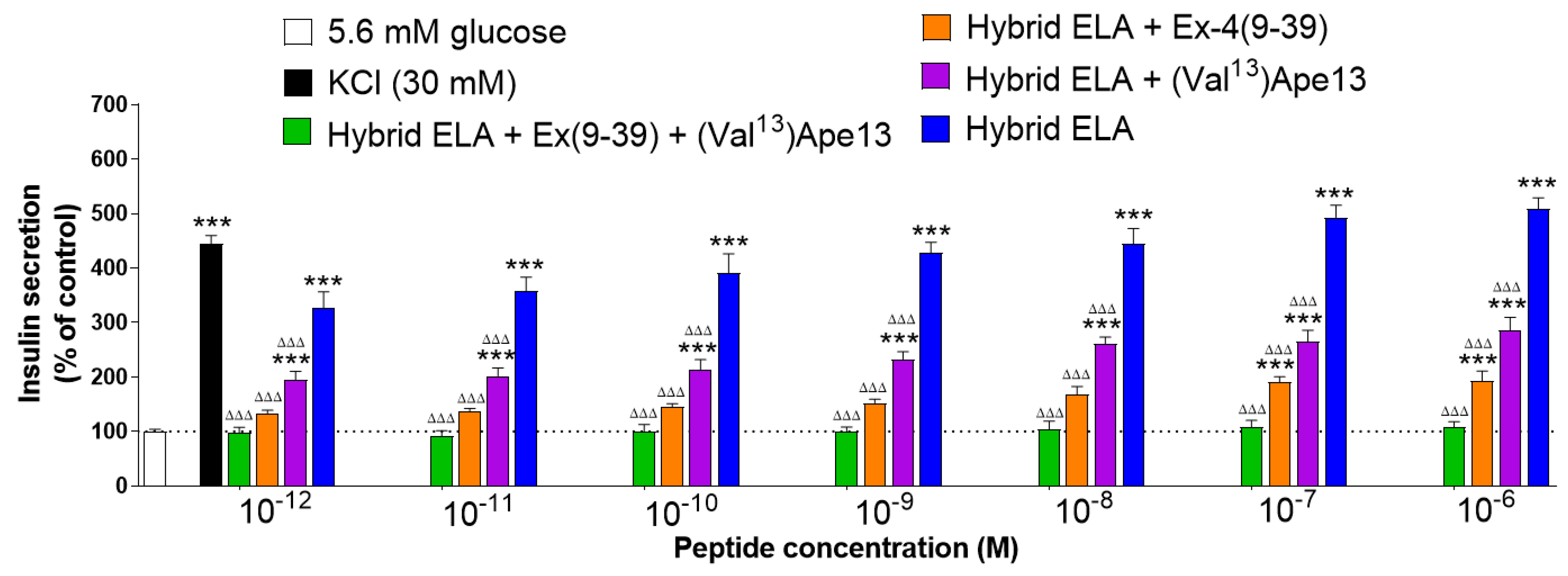
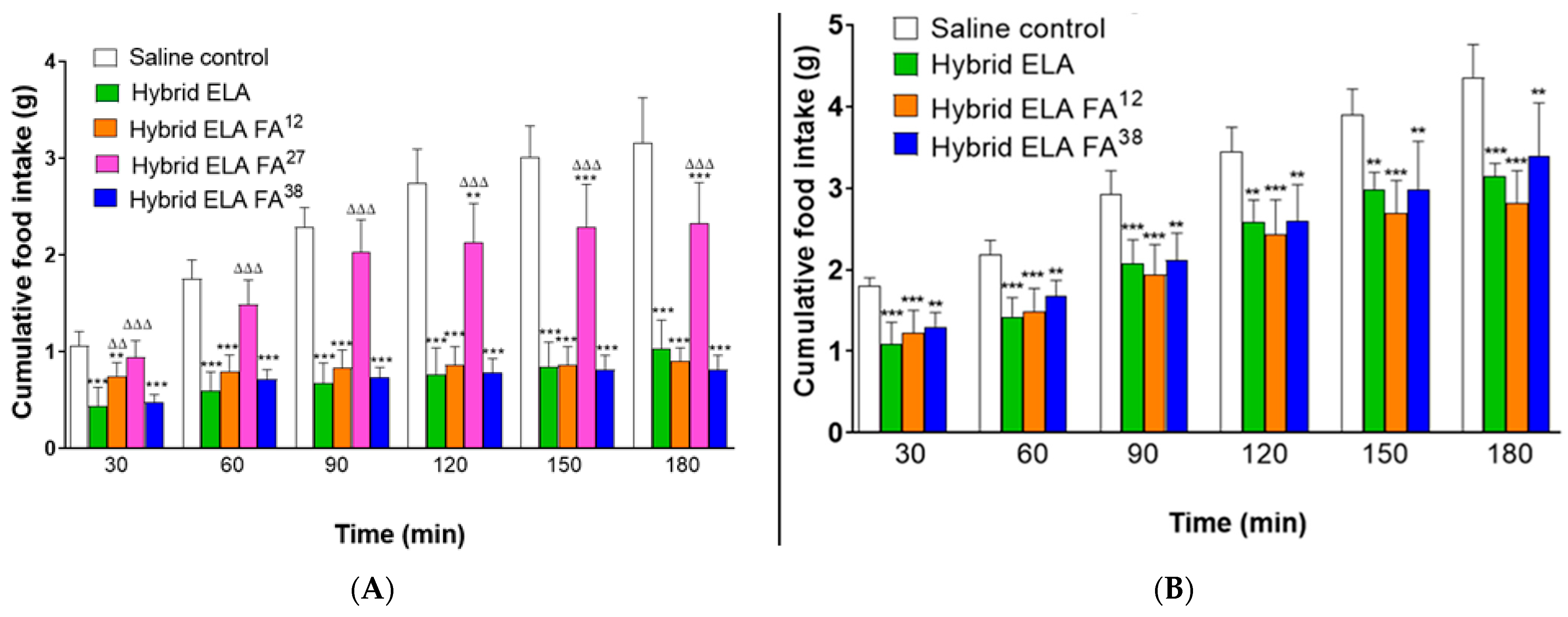
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Patents
References
- Boucher, J.; Masri, B.; Daviaud, D.V.; Gesta, S.; Guigné, C.; Mazzucotelli, A.; Castan-Laurell, I.; Tack, I.; Knibiehler, B.; Carpéné, C.; et al. Apelin, a newly identified adipokine up-regulated by insulin and obesity. Endocrinology 2005, 146, 1764–1771. [Google Scholar] [CrossRef] [PubMed]
- Castan-Laurell, I.; Dray, C.; Knauf, C.; Kunduzova, O.; Valet, P. Apelin, a promising target for type 2 diabetes treatment? Trends Endocrinol. Metab. 2012, 23, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Englander, E.W.; Gomez, G.A.; Rastellini, C.; Quertermous, T.; Kundu, R.K.; Greeley, G.H., Jr. Pancreatic islet APJ deletion reduces islet density and glucose tolerance in mice. Endocrinology 2015, 156, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- Ringström, C.; Nitert, M.D.; Bennet, H.; Fex, M.; Valet, P.; Rehfeld, J.F.; Wierup, N. Apelin is a novel islet peptide. Regul. Pept. 2010, 162, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Palmer, E.S.; Irwin, N.; O’Harte, F.P.M. Potential Therapeutic Role for Apelin and Related Peptides in Diabetes: An Update. Clin. Med. Insights Endo Diabetes 2022, 15, 11795514221074679. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Gheblawi, M.; Wang, K.; Vu, J.; Kondaiah, P.; Oudit, G.Y. Interaction between the apelinergic system and ACE2 in the cardiovascular system: Therapeutic implications. Clin. Sci. 2020, 134, 2319–2336. [Google Scholar] [CrossRef] [PubMed]
- Teague, M.; Martinez, A.; Walker, E.; El-Rifai, M.; Carris, N.W. Use and Interchange of Incretin Mimetics in the Treatment of Metabolic Diseases: A Narrative Review. Clin. Ther. 2023, 45, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes state-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef] [PubMed]
- Lafferty, R.A.; Tanday, N.; Flatt, P.R.; Irwin, N. Development and characterisation of a peptidergic N-and C-terminally stabilised mammalian NPY1R agonist which protects against diabetes induction. Biochim. Biophys. Acta Gen. Subj. 2020, 1864, 129543. [Google Scholar] [CrossRef] [PubMed]
- McClenaghan, N.H.; Barnett, C.R.; Ah-Sing, E.; Abdel-Wahab, Y.H.A.; O’Harte, F.P.M.; Yoon, T.W.; Swanston-Flatt, S.K.; Flatt, P.R. Characterization of a novel glucose-responsive insulin-secreting cell line, BRIN-BD11, produced by electrofusion. Diabetes 1996, 45, 1132–1140. [Google Scholar] [CrossRef]
- Flatt, P.R.; Bailey, C.J. Abnormal plasma glucose and insulin responses in heterozygous lean (ob/+) mice. Diabetologia 1981, 20, 573–577. [Google Scholar] [CrossRef]
- O’Harte, F.P.M.; Parthsarathy, V.; Hogg, C.; Flatt, P.R. Apelin-13 analogues show potent in vitro and in vivo insulinotropic and glucose lowering actions. Peptides 2018, 100, 219–228. [Google Scholar] [CrossRef] [PubMed]
- O’Harte, F.P.M.; Parthsarathy, V.; Hogg, C.; Flatt, P.R. Acylated apelin-13 amide analogues exhibit enzyme resistance and prolonged insulin releasing, glucose lowering and anorexic properties. Biochem. Pharmacol. 2017, 146, 165–173. [Google Scholar] [CrossRef] [PubMed]
- O’Harte, F.P.M.; Parthsarathy, V.; Hogg, C.; Flatt, P.R. Long-term treatment with acylated analogues of apelin-13 amide ameliorates diabetes and improves lipid profile of high-fat fed mice. PLoS ONE 2018, 13, e0202350. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Harte, F.; Parthsarathy, V.; Craig, S.; Palmer, E.; Irwin, N. Peptide Co-Agonists for Combined Activation of the APJ and GLP-1 Receptors with Insulinotropic and Satiety Actions Show Potential for Alleviation of Metabolic Dysfunction in Type 2 Diabetes. Med. Sci. Forum 2023, 23, 1. https://doi.org/10.3390/msf2023023001
O’Harte F, Parthsarathy V, Craig S, Palmer E, Irwin N. Peptide Co-Agonists for Combined Activation of the APJ and GLP-1 Receptors with Insulinotropic and Satiety Actions Show Potential for Alleviation of Metabolic Dysfunction in Type 2 Diabetes. Medical Sciences Forum. 2023; 23(1):1. https://doi.org/10.3390/msf2023023001
Chicago/Turabian StyleO’Harte, Finbarr, Vadivel Parthsarathy, Sarah Craig, Ethan Palmer, and Nigel Irwin. 2023. "Peptide Co-Agonists for Combined Activation of the APJ and GLP-1 Receptors with Insulinotropic and Satiety Actions Show Potential for Alleviation of Metabolic Dysfunction in Type 2 Diabetes" Medical Sciences Forum 23, no. 1: 1. https://doi.org/10.3390/msf2023023001