
Effects of Environmental Changes on Gerromorpha (Heteroptera: Hemiptera) Communities from Amazonian Streams

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
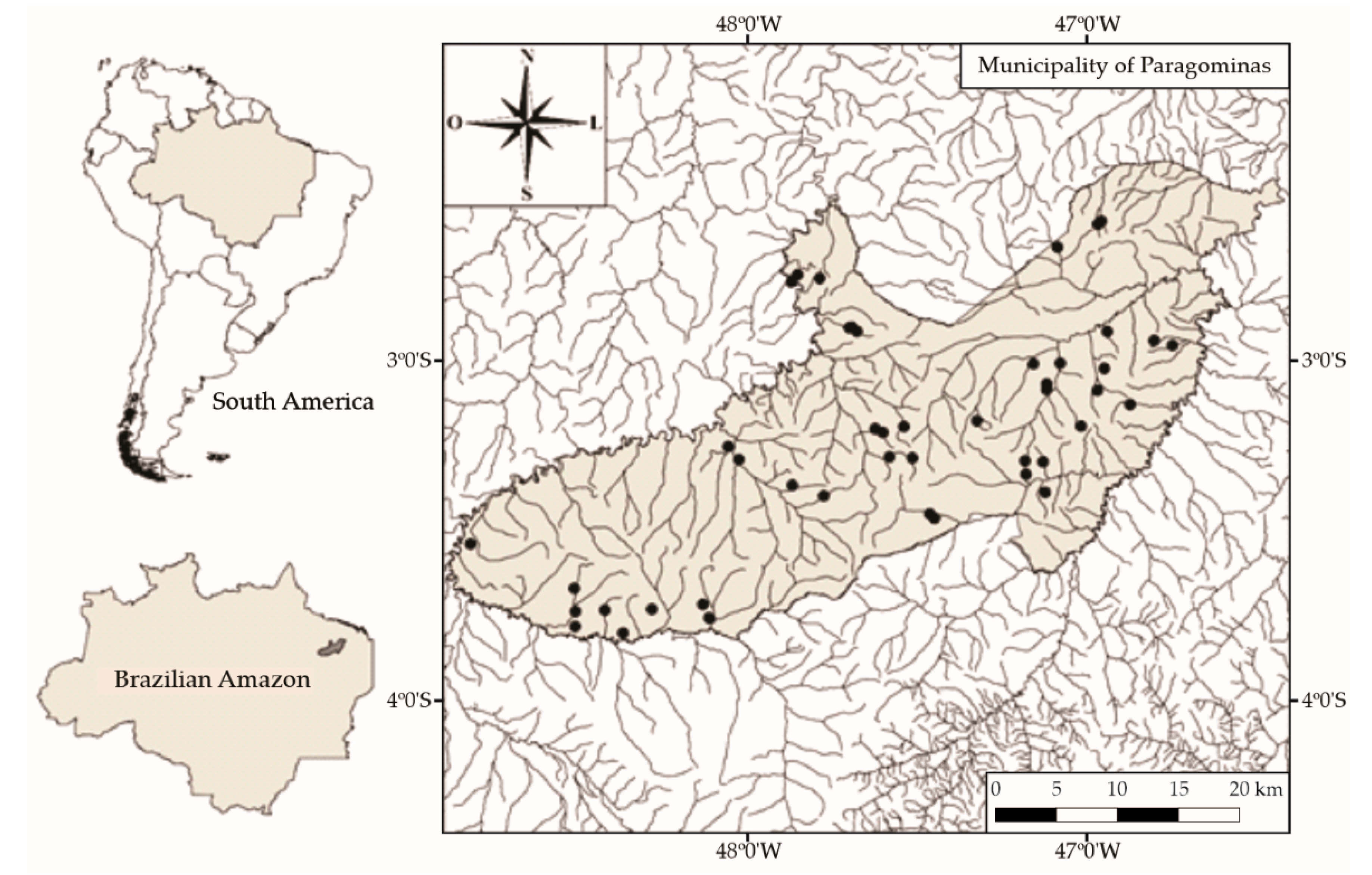
2.1. Study Area
2.2. Data Collection
2.2.1. Biological Sampling
2.2.2. Environmental Characteristics
2.2.3. Habitat Structure
2.3. Data Analysis
3. Results
3.1. Community Description
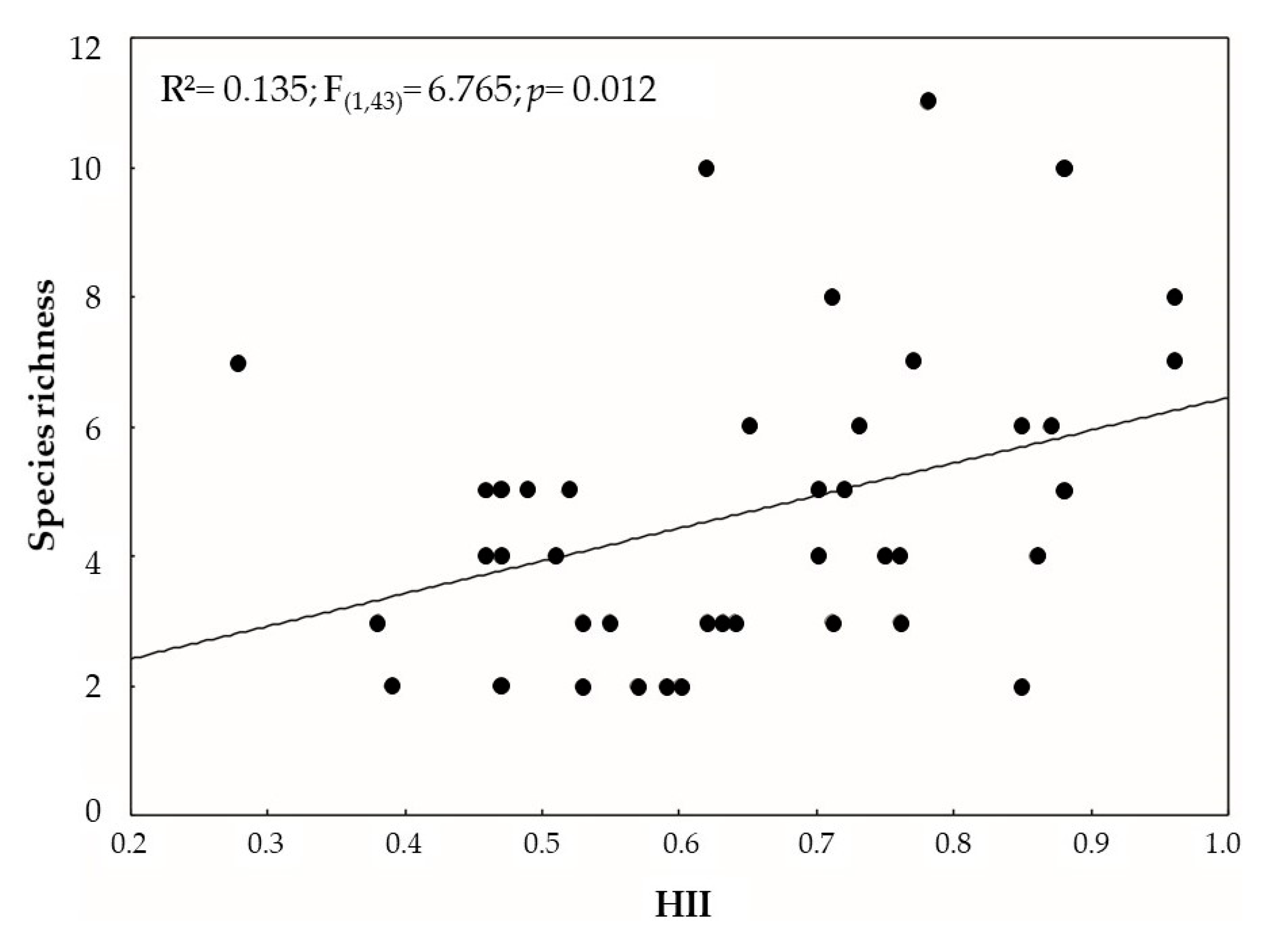
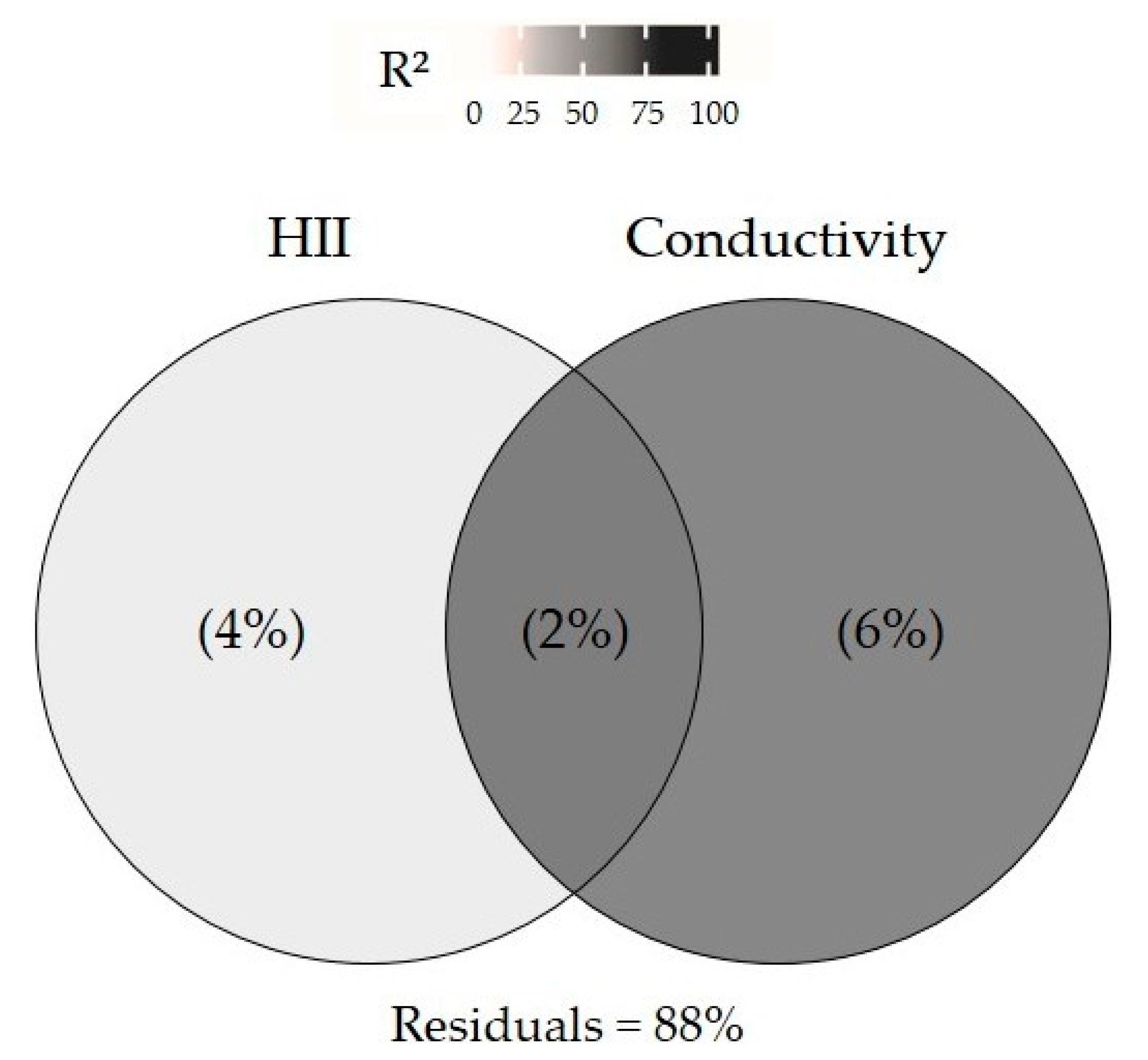
3.2. Effect of Environmental Variables on Community Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grasni, P.; Kent, M.E.; Kenneth, G.C. Distinguishing between yield advances and yield plateaus in historical crop production trend. Nat. Commun. 2013, 4, 2918. [Google Scholar] [CrossRef]
- Allan, J.D.; Castilho, M.M. Stream Ecology: Structure and Function of Running Waters; Kluwer Academic Publishers: Amsterdam, The Netherlands, 2007; p. 436. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Dala-Corte, R.B.; Melo, A.S.; Siqueira, T.; Bini, L.M.; Martins, R.T.; Cunico, A.M.; Pes, A.M.; Magalhães, A.L.B.; Godoy, B.S.; Leal, C.G.; et al. Thresholds of freshwater biodiversity in response to riparian vegetation loss in the Neotropical region. J. Appl. Ecol. 2020, 57, 1391–1402. [Google Scholar] [CrossRef]
- Latrubesse, E.M.; Arima, E.Y.; Dunne, T.; Park, E.; Baker, V.; Horta, F.M.; Wight, C.; Wittmann, F.; Zuanon, J.; Baker, P.A.; et al. Damming the rivers of the Amazon basin. Nature 2017, 546, 363–369. [Google Scholar] [CrossRef]
- Franken, W.; Leopoldo, P.R. Hydrology of catchment areas of Central-Amazonian forest streams. In The Amazon; Springer: Dordrecht, The Netherlands, 1984; pp. 501–519. [Google Scholar]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The Food pulse concept in river-Floodplain system. Can. J. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Davidson, E.A.; Neill, C.; Krusche, A.V.; Ballester, M.V.R.; Markewitz, D.; Figueiredo, R.O. Loss of nutrients from terrestrial ecosystems to streams and the atmosphere following land use change in Amazonia. In Ecosystems and Land Use Change; Defries, R., Asner, G., Houghton, H.R., Eds.; American Geophysical Union: Washington, DC, USA, 2004; Volume 153, pp. 147–158. [Google Scholar]
- Teresa, F.B.; Casatti, L. Importância da vegetação ripária em região intensamente desmatada no sudeste do Brasil: Um estudo com peixes de riacho. Pan Am. J. Aquat. Sci. 2010, 5, 444–453. [Google Scholar]
- Höbinger, T.S.; Schindler, B.; Seaman, S.; Wrbka, T.; Weissenhofer, A. Impact of oil palm plantations on the structure of the agroforestry mosaic of La Gamba, southern Costa Rica: Potential implications for biodiversity. Agrofor. Syst. 2012, 85, 367–381. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juen, L.; Cunha, E.J.; Carvalho, F.G.; Ferreira, M.C.; Begot, T.O.; Andrade, A.L.; Montag, L.F.A. Effects of oil palm plantations on the habitat structure and biota of streams in Eastern Amazon. River Res. Appl. 2016, 32, 2081–2094. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding remarks. Cold Spring Harbor. Symp. Quant. Biol. 1957, 22, 415–427. [Google Scholar] [CrossRef]
- Vieira, T.B.; Dias-Silva, K.; Pacífico, E.S. Effects of riparian vegetation integrity on fish and heteroptera communities. Appl. Ecol. Environ. Res. 2015, 13, 53–65. [Google Scholar] [CrossRef]
- Cunha, E.J.; Montag, L.F.A.; Juen, L. Oil palm crops effects on environmental integrity of Amazonian streams and Heteropteran (Hemiptera) species diversity. Ecol. Indic. 2015, 52, 422–429. [Google Scholar] [CrossRef]
- Nessimian, J.L.; Venticinque, E.; Zuanon, J.; De Marco, P.; Gordo, M.; Fidelis, L.; Batista, J.D.; Juen, L. Land use, habitat integrity, and aquatic insect assemblages in Central Amazonian streams. Hydrobiologia 2008, 614, 117–131. [Google Scholar] [CrossRef]
- Brasil, L.S.; Lima, E.L.; Spigolonic, Z.A.; Ribeiro-Brasil, D.R.G.; Juen, L. The habitat integrity index and aquatic insect communities in tropical streams: A meta-analysis. Ecol. Indic. 2020, 116, 106495. [Google Scholar] [CrossRef]
- Veras, D.S.; Castro, E.R.; Lustosa, G.; Azevedo, C.A.S.; Juen, L. Evaluating the habitat integrity index as a potential surrogate for monitoring the water quality of streams in the cerrado-caatinga ecotone in northern Brazil. Environ. Monit. Assess. 2019, 191, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Dias-Silva, K.; Vieira, T.B.; de Matos, T.P.; Juen, L.; Simião-Ferreira, J.; Hughes, R.M.; De Marco, P., Jr. Measuring stream habitat conditions: Can remote sensing substitute for field data? Sci. Total Environ. 2021, 788, 147617. [Google Scholar] [CrossRef]
- Cunha, E.J.; Juen, L. Impacts of oil palm plantations on changes in environmental heterogeneity and Heteroptera (Gerromorpha and Nepomorpha) diversity. J. Insect Conserv. 2017, 21, 119. [Google Scholar] [CrossRef]
- Guterres, A.P.; Cunha, E.J.; Godoy, B.S.; Silva, R.R.; Juen, L. Co-occurrence patterns and morphological similarity of semiaquatic insects (Hemiptera: Gerromorpha) in streams of Eastern Amazonia. Ecol. Entomol. 2020, 45, 155–166. [Google Scholar] [CrossRef]
- Martins, R.T.; Brito, J.; Dias-Silva, K.; Leal, C.G.; Leitão, R.P.; Oliveira, V.C.; Oliveira-Junior, J.M.B.; Ferraz, S.B.; De Paula, F.; Roque, F.O.; et al. Low forest-loss thresholds threaten Amazonian fish and macroinvertebrate assemblage integrity. Ecol. Indic. 2021, 127, 107773. [Google Scholar] [CrossRef]
- Dias-Silva, K.; Cabette, H.S.R.; Juen, L.; De Marco, P. The influence of habitat integrity and physical-chemical water variables on the structure of aquatic and semi-aquatic Heteroptera. Zoologia 2010, 27, 918–930. [Google Scholar] [CrossRef] [Green Version]
- Dias-Silva, K.; Brasil, L.S.; Juen, L.; Cabette, H.S.R.; Costa, C.C.; Freitas, P.V.; De Marco, P., Jr. Influence of local variables and landscape metrics on Gerromorpha (Insecta: Heteroptera) assemblages in savanna streams, Brasil. Neotrop. Entomol. 2020, 49, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Cunha, E.J.; Guterres, A.P.M.; Godoy, B.S.; Juen, L. Wing dimorphism in semiaquatic bugs (Hemiptera, Heteroptera, Gerromorpha) as a tool for monitoring streams altered by oil palm plantation in the Amazon. Ecol. Indic. 2020, 117, 106707. [Google Scholar] [CrossRef]
- Rosenberg, D.M.; Resh, V.H. Freshwater Biomonitoring and Benthic Macroinvertebrates; Chapman & Hall: New York, NY, USA, 1993; p. 487. [Google Scholar]
- Guterres, A.P.M.; Cunha, E.J.; Juen, L. Tolerant semiaquatic bugs species (Heteroptera: Gerromorpha) are associated to pasture and conventional logging in the Eastern Amazon. J. Insect Conserv. 2021, 25, 1–13. [Google Scholar] [CrossRef]
- Giehl, N.F.S.; Brasil, L.S.; Dias-Silva, K.; Nogueira, D.S.; Cabette, H.S.R. Environmental Thresholds of Nepomorpha in Cerrado Streams, Brazilian Savannah. Neotropic. Entomol. 2019, 48, 186–196. [Google Scholar] [CrossRef]
- Strahler, A.N. Quantitative analysis of watershed geomorphology. EOS Trans. Am. Geophys. Union 1957, 38, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Diniz, T.D.A. Caracterização Climática da Amazônia Oriental. Pesquisa Sobre Utilização e Conservação de Solo na Amazônia Oriental; Embrapa Cpatu: Belém, Portugal, 1986; p. 291. [Google Scholar]
- Peel, M.C.; Finlayson, B.L.; Mcmahon, T.A. Uptade world map of the Köppen-Geiger climate classification. Hydrol. Earth Syst. Sci. 2007, 11, 1633–1644. [Google Scholar] [CrossRef] [Green Version]
- Gardner, T.A.; Ferreira, J.; Barlow, J.; Lees, A.C.; Parry, L.; Zuanon, J. A social and ecological assessment of tropical land uses at multiple scales: The Sustainable Amazon Network. Philos. Trans. R. Soc. 2013, 368, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-Junior, J.M.B.; Teodósio, M.A.; Juen, L. Patterns of co-occurrence and body size in dragonflies and damselflies (Insecta: Odonata) in preserved and altered Amazonian streams. Austral. Entomol. 2021, 60, 436–450. [Google Scholar] [CrossRef]
- Baptista, D.F.; Dorvillé, L.F.M.; Buss, D.F.; Nessimian, J.L. Spatial and temporal organization of aquatic insects assemblages in the longitudinal gradient of a tropical river. Rev. Bras. Biol. 2001, 61, 295–394. [Google Scholar] [CrossRef] [Green Version]
- Heino, J. Taxonomic surrogacy; numerical resolution and responses of stream macroinvertebrate communities to ecological gradients: Are the inferences transferable among regions? Ecol. Indic. 2014, 36, 186–194. [Google Scholar] [CrossRef]
- Brasil, L.S.; Oliveira-Junior, J.M.B.; Calvão, L.B.; Carvalho, F.G.; Monteiro-Júnior, C.S.; Dias-Silva, K.; Juen, L. Spatial, biogeographic and environmental predictors of diversity in Amazonian Zygoptera. Insect Conserv. Divers. 2018, 11, 174–184. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Levine, P.; Robison, E.G.; Seeliger, C.; Peck, D.V. Quantifying Physical Habitat in Wadeable Streams; United States Environmental Protection Agency: Washington, DC, USA, 1999.
- Peck, D.V.; Herlihy, A.T.; Hill, B.H.; Hughes, R.M.; Kaufmann, P.R.; Klemm, J.; Lazorchak, J.M.; Mccormick, F.H.; Peterson, S.A.; Ringold, P.L.; et al. Environmental Monitoring and Assessment Program-Surface Waters Western Pilot Study: Field Operations Manual for Wadeable Streams; Environmental Protection Agency, Office of Research and Development: Washington, DC, USA, 2006.
- Cabette, H.R.S.; Giehl, N.F.D.S.; Dias-Silva, K.; Juen, L.; Bastista, J.D. Distribuição de Nepomorpha e Gerromorpha (Insecta: Heteroptera) da Bacia do Rio Suiá-Miçú, MT: Riqueza relacionada à qualidade de água e de habitat. In Gestão e Educação Ambiental: Água, Biodiversidade e Cultura; Santos, J.E., Galbiati, C., Moschini, L.E., Eds.; RiMa: São Carlos, Brazil, 2010; pp. 113–137. [Google Scholar]
- Kenaga, E.E. The genus Telmatometra Bergroth (Hemiptera-Gerridae). Univ. Kans. 1941, 17, 169–183. [Google Scholar]
- Kenaga, E.E. A new genus in the Holobatinae (Gerridae-Hemiptera). J. Kans. Entomol. Soc. 1942, 15, 136–141. [Google Scholar]
- Drake, C.J.; Hussey, R.F. Concerning some American Microvelia (Hemiptera: Veliidae). Fla. Entomol. 1951, 34, 137–145. [Google Scholar] [CrossRef]
- Nieser, N.; Melo, A.L. Os Heterópteros Aquáticos de Minas Gerais: Guia Introdutório com Chave de Identificação para as Espécies de Nepomorpha e Gerromorpha; Universidade Federal de Minas Gerais: Belo Horizonte, Brazil, 1997. [Google Scholar]
- Moreira, F.F.F.; Alecrim, V.; Ribeiro, J.R.I.; Nessimian, J.L. Identification key to the Gerridae (Insecta: Heteroptera: Gerromorpha) from the Amazon River floodplain, Brazil, with new records for the Brazilian Amazon. Zoologia 2011, 28, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Moreira, F.F.F.; Barbosa, J.F. A new Rhagovelia (Hemiptera: Heteroptera: Veliidae) from the Brazilian Amazon, with a key to species of the robusta group known from the country. Zootaxa 2014, 3790, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Aristizábal-García, H.; Floriano, C.F.B.; Moreira, F.F.F.; Bispo, P.C. New species and notes on Microveliinae from northern South America (Hemiptera: Heteroptera: Veliidae). Zootaxa 2015, 3980, 591–596. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, O.M.; Moreira, F.F.F.; Galvão, C.A. New species of Rhagovelia Mayr, 1865 (Hemiptera: Heteroptera: Veliidae) from Pará state, with an updated key to Brazilian species of the robusta group. Zootaxa 2016, 4171, 586–594. [Google Scholar] [CrossRef]
- Floriano, C.F.B.; Paladini, A.; Cavichioli, R.R. Systematics of the South American species of Cylindrostethus Mayr, 1865 (Hemiptera: Heteroptera: Gerridae), with a new species from Amazonian Brazil and Peru. Invertebr. Syst. 2016, 30, 431–462. [Google Scholar] [CrossRef]
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Wagner, H.H. Package ‘adespatial’. R Package 2018, 2018, 3–8. Available online: http://cran.nexr.com/web/packages/adespatial/adespatial.pdf (accessed on 20 April 2020).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Wagner, H. R Package; Version 1.17–4; Vegan: Community Ecology Package; CRAM. 2010. Available online: http://CRAN.R-project.org/package=vegan (accessed on 20 April 2020).
- Zar, J.H. Biostatistical Analysis; Pearson Prentice-Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 20 April 2020).
- Shimano, Y.; Juen, L.; Salles, F.F.; Nogueira, D.S.; Cabette, H.S.R. Environmental and spatial processes determining Ephemeroptera (Insecta) structures in tropical streams. Ann. Limnol. Int. J. Lim. 2013, 49, 31–41. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Juen, L. Structuring of Dragonfly Communities (Insecta: Odonata) in Eastern Amazon: Effects of Environmental and Spatial Factors in Preserved and Altered Streams. Insects 2019, 10, 322. [Google Scholar] [CrossRef] [Green Version]
- Poff, N.L.; DeCino, R.D.; Ward, J.V. Size-dependent drift responses of mayflies to experimental hydrologic variation: Active predator avoidance of passive hydrodynamic displacement? Oecologia 1991, 88, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, B.A.; Diniz-Filho, J.A.F.; Jaramillo, C.A.; Soeller, S.A. Climate, niche conservatism, and the global bird diversity gradient. Am. Nat. 2007, 170, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Chase, J.M. Community assembly: When should history matter? Oecologia 2003, 136, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; p. 302. [Google Scholar]
- Pereira, D.F.G.; De Oliveira, J.M.B.; Juen, L. Environmental changes promote larger species of Odonata (Insecta) in Amazonian streams. Ecol. Indic. 2018, 98, 179–192. [Google Scholar] [CrossRef]
- Mendes, T.P.; Oliveira-Junior, J.M.B.; Cabette, H.S.R.; Batista, J.D.; Juen, L. Congruence and the Biomonitoring of Aquatic Ecosystems: Are Odonate Larvae or Adults the Most Effective for the Evaluation of Impacts. Neotrop. Entomol. 2017, 46, 631–641. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Dias-Silva, K.; Teodósio, M.A.; Juen, L. The response of neotropical Dragonflies (Insecta: Odonata) to local and regional abiotic factors in small streams of the Amazon. Insects 2019, 10, 446. [Google Scholar] [CrossRef] [Green Version]
- Luiza-Andrade, A.; Brasil, L.S.; Benone, N.L.; Shimano, Y.; Farias, A.P.J.; Montag, L.F.; Dolédec, S.; Juen, L. Influence of oil palm monoculture on the taxonomic and functional composition of aquatic insect communities in eastern Brazilian Amazonia. Ecol. Indic. 2017, 82, 478–483. [Google Scholar] [CrossRef]
- Savage, A. The distribution of Corixidae in relation to water quality of British lakes: A monitoring model. Freshw. Forum 2010, 4, 32–61. [Google Scholar]
- Couceiro, S.R.M.; Hamada, N.; Luz, S.L.B.; Forsberg, B.R.; Pimentel, T.P. Deforestation and sewage effects on aquatic macroinvertebrates in urban streams in Manaus, Amazonas, Brazil. Hydrobiologia 2007, 575, 271–284. [Google Scholar] [CrossRef]
- Karaouzas, L.; Gritzalis, K.C. Local and regional factors determining aquatic and semi-aquatic bug (Heteroptera) assemblages in rivers and streams of Greece. Hydrobiologia 2006, 573, 199–212. [Google Scholar] [CrossRef]
- Taybi, A.F.; Mabrouki, Y.; Bozdoğan, H.; Millan, A. Are aquatic Hemiptera good indicators of environmental river conditions? Aquat. Ecol. 2021, 55, 791–806. [Google Scholar] [CrossRef]
- Godoy, B.S.; Queiroz, L.L.; Lodi, S.; Oliveira, L.G. Environment and spatial influences on aquatic insect communities in cerrado streams: The relative importance of conductivity, altitude, and conservation areas. Neotrop. Entomol. 2017, 46, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.G.; Pompeu, P.S.; Gardner, T.A.; Leitão, R.P.; Hughes, R.M.; Kaufmann, P.R.; Zuanon, J.; De Paula, F.R.; Ferraz, S.F.B.; Thomson, J.R.; et al. Multi-scale assessment of human-induced changes to Amazonian instream habitats. Landsc. Ecol. 2016, 31, 1725–1745. [Google Scholar] [CrossRef]
- Hu, D.L.; Chan, B.; Bush, J.W.M. The hydrodynamics of water strider locomotion. Nature 2003, 424, 663–666. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Family | Species | N. Individuals |
|---|---|---|
| Gerridae | Brachymetra lata Shaw, 1933 | 56 |
| Cylindrostethus palmaris Drake and Harris, 1934 | 535 | |
| Limnogonus aduncus Drake and Harris, 1933 | 10 | |
| Limnogonus cf. ignotus Drake and Harris, 1934 | 2 | |
| Limnogonus recurvus Drake and Harris, 1930 | 12 | |
| Neogerris lotus (White, 1879) | 28 | |
| Tachygerris adamnsoni (Drake, 1942) | 3 | |
| Tachygerris celosis (Drake and Harris, 1931) | 6 | |
| Hydrometridae | Hydrometra comata (Torre-Bueno, 1926) | 4 |
| Hydrometra guianana (Hungerford and Evans, 1934) | 2 | |
| Veliidae | Euvelia concava Polhemus and Polhemus, 1984 | 18 |
| Microvelia longipes Uhler, 1893 | 1 | |
| Microvelia pulchella Westwood, 1834 | 2 | |
| Paravelia bullialata Polhemus and Polhemus, 1984 | 9 | |
| Rhagovelia aff. scitula (Bacon, 1956) | 1 | |
| Rhagovelia elegans Uhler, 1894 | 317 | |
| Rhagovelia evidis Bacon, 1948 | 683 | |
| Rhagovelia hambletoni Drake and Harris, 1933 | 104 | |
| Rhagovelia jubata Bacon, 1948 | 58 | |
| Rhagovelia robusta Gould, 1931 | 1 | |
| Rhagovelia sp. 1 | 15 | |
| Rhagovelia sp. 2 | 24 | |
| Rhagovelia sp. 3 | 1 | |
| Rhagovelia sp. 4 | 13 | |
| Rhagovelia sp. 5 | 55 | |
| Rhagovelia sp. 6 (angustipes complex) | 103 | |
| Stridulivelia alia (Drake, 1957) | 1 | |
| Stridulivelia astralis (Drake and Harris, 1938) | 12 | |
| Stridulivelia cf. ayacucho Polhemus and Spangler, 1995 | 3 | |
| Stridulivelia quadrispinosa (Hungerford, 1929) | 1 | |
| Stridulivelia raspa (Hungerford, 1929) | 5 | |
| Stridulivelia sp. 1 | 2 | |
| Stridulivelia stridulata (Hungerford, 1929) | 2 | |
| Stridulivelia strigosa (Hungerford, 1929) | 16 | |
| Stridulivelia tersa (Drake and Harris, 1941) | 102 | |
| Stridulivelia transversa (Hungerford, 1929) | 5 | |
| Total | 2212 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moy, K.M.; Brasil, L.S.; Oliveira-Junior, J.M.B.; Juen, L.; Vieira, T.B.; Dias-Silva, K. Effects of Environmental Changes on Gerromorpha (Heteroptera: Hemiptera) Communities from Amazonian Streams. Hydrobiology 2022, 1, 111-121. https://doi.org/10.3390/hydrobiology1010008
Moy KM, Brasil LS, Oliveira-Junior JMB, Juen L, Vieira TB, Dias-Silva K. Effects of Environmental Changes on Gerromorpha (Heteroptera: Hemiptera) Communities from Amazonian Streams. Hydrobiology. 2022; 1(1):111-121. https://doi.org/10.3390/hydrobiology1010008
Chicago/Turabian StyleMoy, Karen Monteiro, Leandro Schlemmer Brasil, José Max Barbosa Oliveira-Junior, Leandro Juen, Thiago Bernardi Vieira, and Karina Dias-Silva. 2022. "Effects of Environmental Changes on Gerromorpha (Heteroptera: Hemiptera) Communities from Amazonian Streams" Hydrobiology 1, no. 1: 111-121. https://doi.org/10.3390/hydrobiology1010008