
Gene-Based Therapy: A New Approach to Feline Induced Sterilization?


Comprehensive Health Research Centre (CHRC), Department of Veterinary Medicine, Science and Technology School, University of Évora, Pole at Mitra, 7002-774 Évora, Portugal
Future Pharmacol. 2023, 3(4), 938-950; https://doi.org/10.3390/futurepharmacol3040057
Submission received: 2 October 2023
/
Revised: 13 November 2023
/
Accepted: 27 November 2023
/
Published: 4 December 2023
Abstract
:Feline population control remains a concern as to whether it is intended for the short- or long-term. Induced sterilization of felids is critical in the case of feral, free-roaming cats, or the management of wild populations in Zoos or sanctuaries. This narrative review explores the shifting paradigm in induced sterilization methods, driven by the development of gene editing approaches recently applied to control felid reproductive activity. Although gene therapy approaches have gained attention as alternatives to more traditional methods, their clinical applications remain in the realm of thought. The objective of this study was to provide a comprehensive overview of the current state and most recent advances in gene-based contraception options, consolidate current research and evidence, and share some considerations on its potential effectiveness, advantages or limitations, and implications for animal welfare and population control strategies. Gene-based contraception therapy tested in felines, targeting the AMH pathway, was unable to suppress the estrous cycle and follicular development. However, at an experimental level, preliminary results hint at the need to change towards different molecular targets. Moreover, their side effects remain largely unknown, and several questions remain unanswered, such as the regularity of treatment applications or cost.
1. Introduction
Because of their naturally high fertility, the control of cat reproduction is fundamental for managing free-roaming, feral populations. According to Vansandt et al. [1], four-fifths of the estimated 600 million domestic cats worldwide are free-roaming. Stray and homeless cats are critical problems worldwide [2,3,4]. Those who survive adversity adopt a free-roaming lifestyle and establish a community population of cats (colonies). Some of them cluster in groups and survive, supported by offers of food and shelter [4]. Others survive in urban and suburban areas, in smaller groups, or individually, accessing multiple scattered food sources [5].
Even though the proportion of owned neutered cats may vary across the globe, the contribution of homeless cats to population replenishment is substantially higher than that of owned cats [5]. Different strategies have been implemented to control the population of feral and homeless cats, focusing on permanent sterilization in association with other measures [5,6], such as relocation to shelters financed by governments or pet rescuing associations, the adoption of trap-neuter-return programs, or animal adoption by particulars [5]. In these situations, most animals undergo surgical or chemical sterilization (ovariohysterectomy in females and orchiectomy in males or intratesticular injection of sterilant to induce azoospermia, respectively), which requires financial investment, labor, and time, as it is followed by the intervention’ recovery period. In addition, some animals are unsuitable for adoption or suffer from stress when in captivity. In addition, it has been referred to as presenting important limitations, such as failing to reach the necessary scale to be effective [5,7], supporting the growing quest for more accessible and effective strategies to control cat reproductive function while guaranteeing the animals’ and environmental welfare.
Some wild felids kept in sanctuaries and Zoos may also be the object of induced sterilization because their natural fecundity would compromise the stability of their captive populations [8]. In threatened or endangered felids maintained in natural parks or Zoos, one main concern is to ensure that treated animals may recover the expected fertility of the species, thus contributing to their genetic diversity. Hence, reversible contraception is often implemented [8], and research on new and safe contraceptive methods remains ongoing. Moreover, fertility control in wild non-captive populations may also be necessary when mitigating human-wildlife conflicts. In the latter situation, permanent sterility is often required and considered a safe and humane approach to control overpopulation in prolific wildlife species [9].
Induced sterility is currently used to control domestic and wild felid populations by suppressing fertility. It can be used for transient, reversible suppression of the reproductive function [8], hence named contraception. Pedigree cat breeders favor reversible and safe contraception for managing the estrous cycle and assisted reproduction programs [10]. Contraception protocols may require multiple administrations of the elicited drug, mainly when used for long-term sterility. However, more frequently, induced sterility aims at a permanent reproductive incapacity and, consequently, the inability to evidence either the reproductive behavior or the ability to produce offspring. The distinction between these two assumptions is crucial when selecting therapeutic approaches.
Some aspects of the protocols, such as the route of administration or the interval to a new treatment cycle, as well as any existing deleterious effects when used in chronic treatments or the need to guarantee the animals’ fertility at the end of treatment, are crucial factors to ponder before implementing a specific protocol. At the end of the suppression period, the animal should be able to recover the predicted fertility potential at the beginning of the therapy. In contrast, if sterilization is foreseen, the animal’s fertility is abolished for good, but it could be essential to determine whether the animal’s reproductive or social behavior should be maintained. Sterilization or contraceptive treatments are currently used in felids, according to their intent: the reversible reproductive control is more often selected for genetically valuable felids, whereas the permanent reproductive control is reserved for generic wild felids, and stray or free-roaming cats’ control [8].
The cost of implementing fertility control measures for free-ranging populations depends on several factors, including the time, labor, and expenses required to achieve the desired population reduction [11]. Moreover, it requires a continuous and persistent effort [12], which adds to the real costs. When considering a surgical approach (e.g., spaying and neutering) the costs encompass anesthesia, pain control drugs, surgical materials, and staff work, as well as the need for adequately equipped facilities and post-surgical care. This method offers the advantage of being a definitive solution for the reproductive control of feral populations. Alternatively, the medical approach involves costs related to materials such as contraceptives, traps, or bait dispensers used for administering contraceptives, as well as personnel expenses for felid capture. These costs can also vary based on the number of administrations required to maintain infertility, according to the specific protocol employed in the targeted population [11,13].
This narrative review briefly overviews the hormonal and non-hormonal pharmacological methods for induced sterilization available for felids (domestic or wild species) before introducing and discussing the most recent advances in gene-based fertility control. This article also aims to share some considerations regarding gene-based sterilization and discuss its potential effectiveness, advantages or limitations, and implications for animal welfare and population control strategies as topics of interest for pet owners, authorities, and veterinarians. By consolidating current research and evidence, this article intends to open the discussion around the use of recent gene-based approaches for contraception or sterilization.
2. Hormonal vs. Non-Hormonal Pharmacological Methods for Feline Induced Sterilization
Progestogens are at the base of the most frequently used protocols to limit feline reproduction [14,15,16,17,18,19] and can be applied to domestic and wild felids. Despite being cheaper, they have several shortcomings. A significant limitation is that they present several side effects associated with prolonged progestogen administration, such as uterine diseases (e.g., pyometra or cystic endometrial hyperplasia) and mammary tumors [20]. Another drawback is that when progestogen-based methods are applied to animals not intended for reproduction, whether owned or feral cats or even wild felids, most protocols demand regular administration, and thus regular captures. These shortcomings have led to the search for new and less harmful treatments for contraception and sterilization.
To mitigate the side effects associated with prolonged progestogen use, alternative strategies have been used to regulate the hypothalamus-pituitary axis via GnRH pulsatility control in cats. Three potential approaches involve the administration of melatonin to suppress gonadal activity, impairing GnRH secretion, and subsequently the gonadal function, or employing immunization against key reproductive molecules.
Using exogenous melatonin, the female cat enters an artificial anestrous period, suspending ovarian activity [21]. The treatment preferably uses a subcutaneous implant [22] to avoid the need for repetitive oral administration at a particular time of the day (~2 h before darkness) [10,21]. Still, the melatonin implants have a short duration of the ovarian suppressive effect (ranging from 2 to 4 months, depending on the estrous cycle stage at implantation; particularly, the follicular phase should be avoided) or around two months in male cats. Besides, the suppressive effect varies with the doses and the individual [23,24], making its clinical use in estrous cycle control difficult and predicting the resumption of ovarian activity. Other main limitations to the success of the treatment include the inability of melatonin to post-pone puberty and the sporadic descriptions of ovulations occurring early in the post-treatment period [23,25], and clinical pseudopregnancy conditions [23]. Melatonin treatments are not used in males, as the effects of photoperiod on male reproductive function are less evident [26,27]. Male cats usually show light to moderate seasonal changes in sperm quality [27], but libido is not largely affected, and they retain breeding ability [28] unless the negative photoperiod is extended [29]. The effects of melatonin treatment in male cats mimic season-associated changes in gonadal function [30].
The use of implants for a slow-release of GnRH agonists, such as deslorelin, is aimed at the desensitization of GnRH receptors, compromising the synthesis of gonadotropins and, consequently, the suppression of the gonadal function [31]. Considering the effects of deslorelin, treated animals revert to an artificial prepubertal-like stage. Despite its safety, a major constraint in deslorelin implants is the flare-up effect [32], similar to those observed in dogs [33]. Shortly after deslorelin implantation in females, stimulation of ovarian activity has been recorded in many cases, with cats displaying signs of estrus and, therefore, being able to conceive [32]. Because of this flare-up effect, deslorelin implants have been successfully used to induce estrous cycles in felines [24]. After the initial trigger effect, the deslorelin implant had a long suppressive effect on feline reproductive function. The reports respecting the length of treatment suggest it is unpredictable, ranging from 8 to 21 months for the 4.7 mg of deslorelin [24,32], which may be expanded to 25–28 months if the 9.4 mg deslorelin implant is used [34]. A breed effect has been suggested (for details see [24]), which should be considered when scheduling the administrations and defining the reproductive management of the treated animals. No side effects other than the flare-up were reported for the deslorelin implant (Suprelorin®, Virbac, Carros, France), whatever the implant dose (4.7 mg or 9.4 mg). Nonetheless, sporadic situations of feline mammary hyperplasia (fibroadenomatosis) or cystic endometrial hyperplasia have been reported [24], possibly associated with ovulation occurring due to a flare-up response, and should be monitored. After treatment, the animals recovered their fertility and were able to produce litters [24].
Immunocontraception targeting reproductive hormones (e.g., GnRH) or proteins (e.g., GnRH receptors, zona pellucida antigens) have been developed to control mammalian reproduction with controversial results [35]. Vaccines against the zona pellucida proteins present several side effects [36], including their transient effects on fertility, thus requesting frequent revaccinations, and also do not suppress the gonadal function [35]; for that, they are not adequate for feline-induced sterilization. Recently, new alternative protocols based on GnRH immunization have been tested in cats. These vaccines usually use a synthetic or recombinant GnRH molecule conjugated with a carrier protein (STF2, one of the flagellin proteins of Salmonella [37]) or a diphtheria toxoid [38] to enhance the response to the vaccine. Even though most studies consider this approach of potential interest, they only respect experimental situations (the observational period lasted 5 to 6 months), not allowing the forecast of success in free-ranging situations. The induced sterility is expected to occur 6 to 8 weeks after the second vaccination [37,38]. The main issues to overcome that hinder the application of the technique for prolonged sterilization include the dependency on booster vaccinations at still undetermined intervals and the existence of important individual variability that makes the identification of non-responders or poor responders’ cats before treatment difficult [37]. A positive aspect is the inexistence of side effects besides a slight decrease in some hematological parameters that remained within the reference range, or a deviated behavior associated with the vaccination (respectively [38] and [37]). A different approach to control GnRH activity targets vaccination against GnRH receptor [39]. Albeit the immunogenic product originated from cats, the vaccines were only tested in mice—thus they are not yet available for clinical use—although the authors intend to develop the product to be applied in the control feral feline populations.
Table 1 summarizes the most important constraints in the use of non-progestogen contraceptive methods mentioned above.
Limited options are available to induce reversible sterilization in feral or wild felids despite many of the aforementioned methods being routinely used in veterinary practice in owned cats. Three main issues remain to be addressed. These include limited options for long-term sterilization other than surgery. Furthermore, the application of available methods for wild felids remains poorly studied and largely relies on the translation of the information provided by domestic animals; however, species-specificities regarding the reproductive function may hinder the expected outcome, driven by the need to validate safe and effective tailored long-term contraception protocols in non-domestic species, to determine a species susceptibility to different products and test the most suitable and most extended interval between administrations in long-lasting protocols. Another essential but different concern respects the need for funding research to enhance the efficacy of the method and progress to occur, for instance, by investing in new delivery systems or devices facilitating repetitive administrations or even developing innovative methods.
3. Gene-Based Methods for Feline Induced Sterilization
Gene-based sterilization refers to the use of genetic manipulation to control reproductive processes and induce infertility or sterility. Gene therapy sterilization is taking its first steps in product development and in vivo testing, although no established method has yet been approved for widespread use.
Gene-based therapy concepts arose more than 50 years ago [40]. Multiple challenges have been overcome in recent decades, and tremendous progress has been made in this regard. Today, it represents the promise of durable clinical benefits to complex human diseases and brought significant progress in the medical field.
3.1. Mechanistic of Gene-Based Therapy in Brief
Gene therapy aims to manipulate or modify genes to treat diverse congenital or acquired disorders [40,41]. More recently, it has been extended to control reproductive function in males and females [42].
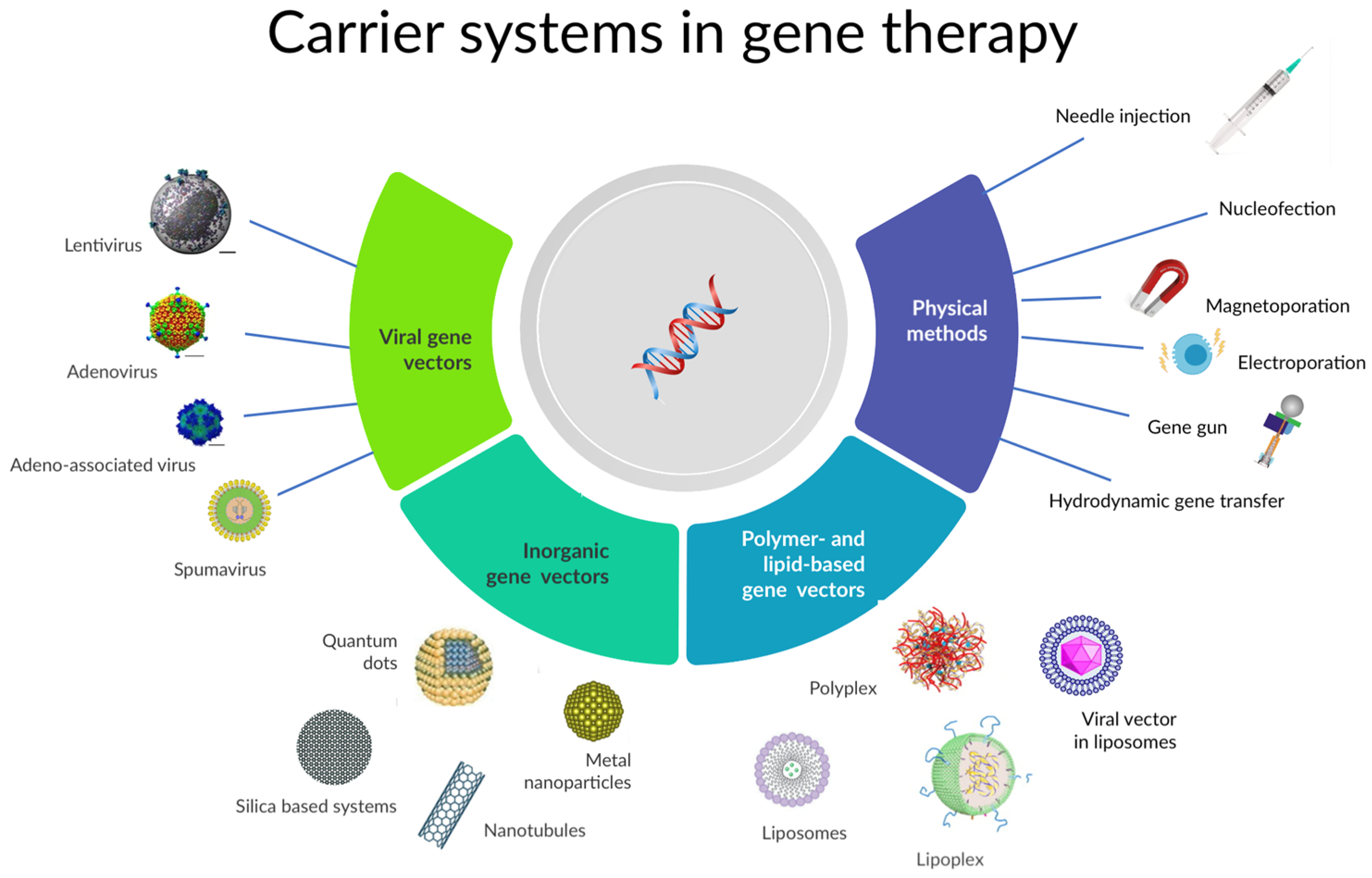
Gene-based therapy involves a complex process. In brief, for gene-based therapy, it is crucial to understand the pathophysiology of the disease or the biological process and the role of the gene and protein that is targeted; then it is needed to produce the modified gene (or transgene), according to the species-genome, and inserting the gene into a carrier system and finally, to inject the product. Usually, the gene-delivering system contains three components: a plasmid gene expression system (which regulates the function of the gene of interest), the transgene (i.e., a gene encoding for a target protein), and the delivery system—or vectors—(that will deliver the transgene into the body) [41]. These delivery systems can be viral (a non-pathogenic vector [43,44], whose main characteristics are listed in Table 2), non-viral (relying upon chemical or physical methods to introduce the protein in the cells, including the use of inorganic nanoparticles with a functionalized surface, such as liposomes and nanotubes, among others [44,45,46]), or hybrid (e.g., genetic combination of viral vectors [47]; combination of viral and chemical vectors [48]; hybrid viral nanoparticles [48] or non-viral nanovectors engineered with pH-sensitive materials [49,50]. Figure 1 depicts the variety of gene carrier systems that are available for gene therapy.
Viral vectors can rely on either RNA or DNA for carrying the transgene; the latter usually integrate their load into the genome and have long-lasting actions, while the former directly transcribe from the inoculated RNA transcripts, and therefore, their actions are not permanent [54]. RNA viral vectors are often used for vaccine development [53]. The most promising viral vectors used are retroviruses, particularly lentiviruses and spumaviruses, adenovirus, and adeno-associated adenoviruses (AAVs). These are usually selected because of their more efficient and non-toxic gene transfer [40]. In AVV vectors, viral coding sequences are replaced by transgenes. They usually carry smaller gene packages (up to 0.5 kb of DNA) [40], contrasting with lentiviruses and spumaviruses that can carry larger and more complex transgenes (Table 2); another difference between AVVs and the latter respect the ability to integrate the cell DNA [43,54]. When a lifelong effect is sought, particularly in gene therapies designed for the correction of congenital diseases, the vector must be able to integrate the carried information into the cell genome so that the gene will be transferred to the daughter cells during the mechanisms of cell replication, in the case of stem cells, or be stabilized in the cell, in the case of long-lived postmitotic cells [52], providing long-lasting expression.
Implementing gene therapy involves three steps [55]: administration of the compound, the transgene delivery into the target cells, and its expression to achieve the proposed outcome. The last but not the least important aspect to consider, is to survey the results of clinical trials, check the results, compare them to conventional therapies, and evaluate for potential side effects. Furthermore, gene therapy strategies can be integrated into two categories according to the administration pathways: in vivo, directly in the patient vs. ex vivo, and in cell cultures originating from a patient to be posteriorly transferred back [52].
3.2. Gene-Based Sterilization
One of the first references to the potential use of gene-based therapy in the reproductive field was presented by Stribley and colleagues [51] in 2002, who discussed the potential application of gene therapy in the reproductive medicine field. The potential areas of interest identified by the authors were obstetrics (for treating fetal pathology) and gynecologic oncology (for treating benign and malignant diseases, particularly ovarian and cervical cancer). The latter has been embraced and developed along with many other gene therapies in the oncological field and has entered the clinical trial stage [56,57].
Over time, the Anti-Müllerian hormone (AMH) ‘s role in follicular development and sustaining ovarian activity in different species was researched. It has been shown that in females, but not males, AMH is a determinant player in follicle recruitment and growth [58]. Indirectly, AMH also determines the estrous cycle exhibition by controlling follicular development and maturation. Also, it has been shown that high concentrations of AMH in adult male rats inhibit adult Leydig cell steroidogenesis, thereby decreasing the testosterone secretion by Leydig cells [59]. Evidence collected from AMH research led to the hypothesis that a vectored-gene delivery approach might also successfully induce long-term infertility in males.
This hypothesis has been developed and tested in the Dr. Pépin laboratory, linked to gene therapy research in oncology. In 2015, Pépin et al. [60] reported the use of adeno-associated virus-delivered gene therapy to block primordial follicle activation by up-regulating human AMH synthesis. Later, another study from the same team showed that the arrest of primordial follicle development achieved with this gene therapy was reversible once the standard hormone levels were re-established, e.g., by transplanting the ovaries of treated mice. The authors defend that this technique would also protect the ovarian follicular pool from the deleterious effects of chemotherapy in young animals [61]. Moreover, they also analyzed the genital tract development in pups born from transplanted ovaries to demonstrate that no vertical transmission of the edited gene occurred in litters born from treated mothers with high levels of MIS.
At the same moment, revisiting the immunocontraception ideas, Li et al. [42] tested the use of a therapeutic antibody gene transfer approach to induce long-term sterilization in mice. In this approach, the gene controlling the production of a specific antibody is introduced in the body, driving the production of endogenous antibodies against that specific molecule. The authors report the results for two different antibody genes [42]. In one of the experiments, a vectored anti-GnRH antibody gene was delivered by a recombinant AAV, administered in a single intramuscular dose, to male and female mice, originating a long-term suppression of the reproductive function in a dose-dependent manner (seven unevenly dosed-groups of 3 to 11 mice each). Despite that females developing titers above 200 g/mL were mainly infertile, four animals (out of 42) reversed the situation after an initial period of 8 weeks. Fertility in females was tested by the production of pups after breeding with a fertile, untreated male at predefined moments of the experiment (at 8-, 28-, 36-, and 44-weeks post-administration). Females treated with higher titers evidenced a complete suspension of follicular development in the ovaries, whereas the counterpart-treated males showed a reduction of the testicular size, along with a decrease in testosterone production and the arrest of meiosis in seminiferous tubules, associated with a lack of spermatozoa in the epididymis [42]. In the second experiment, the authors claimed to achieve long-term sterilization in female mice treated with a vectored anti-zona pellucida_2 (ZP2) antibody gene [42]. Even though, in this experiment, around 42% (5 in 12) of the females were able to produce pups (although with reduced litter sizes) after a first breeding trial five weeks after the product administration, all but one of the treated females failed to produce progeny in subsequent allowed mating periods. The histological evaluation of the gonads of treated females evidenced developing follicles and corpora lutea, like those of control mice, although the zona pellucida around oocytes showed disturbed morphology. As a conclusion for this second experiment, the authors defend that the vectored expression of anti-ZP antibodies was able to induce long-term infertility [42]. Nonetheless, sterility was not obtained, and the evidence hints at the persistence of ovarian cyclic activity as the treatment failed to disrupt the normal follicular maturation.
Some disadvantages have been experienced or expected when cells are transfected with immunoglobulin genes, limiting the technique’s efficiency, namely: the rise in the antibody titers registered following the treatment response was variable with the individual, originating unpredictable “non-responders”; their persistence in plasma tended to decrease with time, driving a temporary effect requesting repeated administrations; reproductive behavior and fertility were not completely abolished, but only transitory infertility was obtained.
Despite the reported challenges, the experiments in mice [42,61] contributed to supporting the hypothesis that using vectored transgene delivery upon a single product administration was a feasible approach for lifetime, non-surgical induced contraception in small animals [62], which the authors deem particularly useful to control community or feral animals’ populations. According to the authors, administering molecules with contraceptive potential (such as an antibody against a specific molecule or hormone transgene) would allow the body to synthesize that molecule for long periods, bypassing the natural reactivity and time-limited response of the body’s immune system, and consequently leading to the annulment of crucial reproductive pathways, for which they named the technique vectored contraception. Nonetheless, the developed vectored gonadotropin releasing hormone vaccine’s limited contraceptive effect, particularly when using homologous GnRH [63,64].
Embedding this line of thought, a very recent publication reported the development and experimental use of an adeno-associated viral vector carrying an AMH transgene that, used in a single dose, was able to suppress the ovarian function in intact young and mature female cats throughout a period of 42 months without evidence of deleterious side-effects [1]. The treatment led to a sharp increase in serum AMH concentrations at the end of the first week, the change depending on the product dose. This increase was followed by a gradual decrease, starting five months after the treatment, which was more pronounced at the end of the first year. AMH values stabilized at lower values at the beginning of the second year after the injection, even though they remained higher than the physiological threshold of 0.25 µg/mL [1]. One-third of the treated females demonstrated unwanted estrous behavior in the scheduled testing mounting periods. The breeding activity failed to induce ovulation and the formation of corpora lutea in only one female in the high-dose group [1]. The authors reported the absence of antibodies against the vector complexes when using the AMH transgene developed exclusively from the cat genome [1]. The authors recognize that their experiment shows that an ectopic expression of anti-Müllerian hormone failed to hinder sex steroid secretion or the expression of regular estrous cycles, even though it prevented breeding-induced ovulation [1]. The major inconvenience inferred from this study could be the inability to suppress feline estrous cycles and follicular development, a crucial aspect when the expected outcome will be to control feline populations.
3.3. Risks and Limitations in Gene Therapy
It is important to note that gene therapy is generally still largely experimental and faces numerous scientific, ethical, and regulatory challenges. Besides, these methods may not be readily accepted without reservations by the community, as they are yet to be proven effective, long-lasting, and safe. Consequently, an existing reluctance to accept gene therapy techniques may compromise the willingness to participate in study trials, delaying the gathering of the necessary volume of data to determine the efficiency and efficacy of these techniques.
Screening the available information in gene-based therapy, mainly when applied to induced contraception or sterilization raises two main concerns: the safety of the therapy and its efficacy. An additional issue can be the cost, which will limit the broad use of this technique in different veterinary medicine contexts.
In general, there are some safety concerns about viral usage, i.e., the ability of the viral genome to be incorporated into the receiver species [65] and the possibility that viral shedding occurs in bodily fluids [66] and poses a public health risk. Using non-pathogenic viral vectors with stable genomes or the engineering of nonviral carriers allows for circumventing this issue and mitigating potential safety risks. Nonetheless, further studies are needed to ensure safety during clinical trials and therapy applications [66,67]. Another safety concern relates to the interaction with the receiver organism, both by the accidental activation or inhibition of endogenous gene expression (e.g., driving oncogene expression [44]), the non-specific uptake by non-targeted organs, which could originate unwanted side effects or loss of efficacy [67], or the ability to induce unwanted inflammatory and immune reactions [44]. Besides any discomfort and temporary disease-like conditions, the reactions against the delivered complex may foster the development of antibodies against the vectored transgene, thus reducing its therapeutic action and originating unpredictable “non-responders”. To avoid this risk, researchers have explored the manipulation of immune tolerance, the choice of delivery vector and dosage, the design of new vectors, and the study of alternative routes of inoculation, among others [44,67].
The main efficacy-related concerns expand from the previously mentioned to include the gene carrier systems, the length of the successful therapy (or the intervals between administrations), and the specificity of the response [68,69]. Most advances in this respect result from the increasing number of multiple-stage clinical trials (cells, animal models, and humans) and the progress in genetic engineering. The success of some clinical trials opened space to offer innovative treatments to selected patients and the route to the production of new laws and regimentations. However, this field is still expanding, and much work is still foreseen.
When extending the abovementioned concerns to the use of gene therapy to induce long-term sterilization and control feline feral and free-roaming populations, according to data reported in some studies [1,42,61], the proposed approaches still fail to achieve the promised long-lasting sterilization that enables the control of community or feral populations of cats. Contrasting to the conceptualization of the term contraception used in human medicine, where the main goal is to prevent pregnancy while retaining the sexual drive, the paradigm of contraception in veterinary medicine includes annulling all the reproductive activity from the animal, including the expression of the estrous cycle and breeding behavior that might reflect in the group dynamics (by enhancing animals roaming and fighting). Until this moment, the reported methods have failed to suppress gonadal activity completely. To achieve such desired control, a logical approach would be to target the GnRH hormone, as the central master controlling gonadal activity. So far, virus-vectored immunocontraceptive vaccines tested in other species have failed to ensure contraception. Despite the vector effect on the production of anti-GnRH antibodies, the contraceptive effect is limited and highly variable between individuals. This raises a different question: are these results related to the species’ particularities, and should the rhythm of administration be tailored to the species? Or is the brain-blood barrier, preventing the passive diffusion of antibodies [70], contributing to impairing an immune-modulated suppression of gonadotrophin secretion from the pituitary? Anyway, considering that for the control of feral and colony populations, a long-term suspension of reproductive activity is desired, it may be possible that a completely different approach must be faced, such as the gene silencing of a main pathway controlling reproduction.
The costs of gene therapy are possibly the most expensive treatment in human medicine because of the costs (including the R&D costs for cell and gene therapies [71], the intellectual property costs, and the production and delivery costs [72]), limiting the access of many to the new gene therapies. The cost brings a new concern regarding the translation of gene therapy technologies to the veterinary medicine field, particularly for much-needed population control.
4. Concluding Remarks
Gene therapy holds great promise for treating multiple diseases. It has been proposed that its benefits could also extend to induced sterilization, a problematic/almost intangible outcome in feral or free-roaming and wild feline population control.
Nevertheless, the inability of the method to suppress the estrous behavior or mating behavior, as well as the need for a two-month adaptation period to reach the full contraceptive effect, questions the gene therapy efficacy for induced sterilization, since we can still not achieve better than a durable infertility condition. Also, the veritable length of the effect remains to be determined, as in cats, treated animals were followed for a limited period of time. Another concern still unanswered is the existence of long-term side effects associated with gene-vectored treatment. Moreover, because of the prevalence of progesterone-associated uterine and mammary conditions, the possibility of treated animals achieving ovulation and experiencing a luteal phase remains a strong negative aspect of gene-based therapy for contraception in cats. The costs of gene therapy are very high even in human medicine, which brings a new concern to the technique’s application in the veterinary medicine context, particularly for population control. Therefore, gene-based long-term sterilization in felids remains in the realm of aspirations.
Funding
This research was funded in whole or in part by the Fundação para a Ciência e a Tecnologia, I.P. (FCT, Funder ID = 50110000187) under Grant (UIDP/04923/2020).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The author kindly appreciate the language editing of the manuscript by Margarida Simões.
Conflicts of Interest
The author declares no conflict of interest.
References
- Vansandt, L.M.; Meinsohn, M.C.; Godin, P.; Nagykery, N.; Sicher, N.; Kano, M.; Kashiwagi, A.; Chauvin, M.; Saatcioglu, H.D.; Barnes, J.L.; et al. Durable contraception in the female domestic cat using viral-vectored delivery of a feline anti-Müllerian hormone transgene. Nat. Commun. 2023, 14, 3140. [Google Scholar] [CrossRef]
- Hamilton, F. Implementing Nonlethal Solutions for Free-Roaming Cat Management in a County in the Southeastern United States. Front. Vet. Sci. 2019, 6, 259. [Google Scholar] [CrossRef]
- Kennedy, B.P.A.; Cumming, B.; Brown, W.Y. Global Strategies for Population Management of Domestic Cats. Animals 2020, 10, 663. [Google Scholar] [CrossRef]
- Luzardo, O.P.; Zaldívar-Laguía, J.E.; Zumbado, M.; Travieso-Aja, M.D.M. The Role of Veterinarians in Managing Community Cats: A Contextualized, Comprehensive Approach for Biodiversity, Public Health, and Animal Welfare. Animals 2023, 13, 1586. [Google Scholar] [CrossRef]
- Hurley, K.F.; Levy, J.K. Rethinking the Animal Shelter’s Role in Free-Roaming Cat Management. Front. Vet. Sci. 2022, 9, 847081. [Google Scholar] [CrossRef]
- Crawford, H.M.; Calver, M.C.; Fleming, P.A. A Case of Letting the Cat out of The Bag-Why Trap-Neuter-Return Is Not an Ethical Solution for Stray Cat. Animals 2019, 9, 171. [Google Scholar] [CrossRef]
- Calver, M.C.; Crawford, H.M.; Scarff, F.R.; Bradley, J.S.; Dormon, P.; Boston, S.; Fleming, P.A. Intensive Adoption as a Management Strategy for Unowned, Urban Cats: A Case Study of 25 Years of Trap–Assess–Resolve (TAR) in Auckland, New Zealand. Animals 2022, 12, 2301. [Google Scholar] [CrossRef]
- Munson, L. Contraception in felids. Theriogenology 2006, 66, 126–134. [Google Scholar] [CrossRef]
- Massei, G.; Cowan, D. Fertility control to mitigate human–wildlife conflicts: A review. Wildl. Res. 2014, 41, 1–21. [Google Scholar] [CrossRef]
- Kutzler, M.A. Alternative methods for feline fertility control: Use of melatonin to suppress reproduction. J. Feline Med. Surg. 2015, 17, 753–757. [Google Scholar] [CrossRef]
- Massei, G. Fertility Control for Wildlife: A European Perspective. Animals 2023, 13, 428. [Google Scholar] [CrossRef]
- Gunther, I.; Hawlena, H.; Azriel, L.; Gibor, D.; Berke, O.; Klement, E. Reduction of free-roaming cat population requires high-intensity neutering in spatial contiguity to mitigate compensatory effects. Proc. Natl. Acad. Sci. USA 2022, 119, e2119000119. [Google Scholar] [CrossRef]
- Massei, G.; Miller, L.A. Nonsurgical fertility control for managing free-roaming dog populations: A review of products and criteria for field applications. Theriogenology 2013, 80, 829–838. [Google Scholar] [CrossRef]
- Kutzler, M.; Wood, A. Non-surgical methods of contraception and sterilization. Theriogenology 2006, 66, 514–525. [Google Scholar] [CrossRef]
- Rhodes, L. New approaches to non-surgical sterilization for dogs and cats: Opportunities and challenges. Reprod. Domest. Anim. 2017, 52 (Suppl. 2), 327–331. [Google Scholar] [CrossRef]
- Goericke-Pesch, S.; Wehrend, A.; Georgiev, P. Suppression of fertility in adult cats. Reprod. Domest. Anim. 2014, 49 (Suppl. 2), 33–40. [Google Scholar] [CrossRef]
- Goericke-Pesch, S. Reproduction control in cats: New developments in non-surgical methods. J. Feline Med. Surg. 2010, 12, 539–546. [Google Scholar] [CrossRef]
- Romagnoli, S.; Ferre-Dolcet, L. Reversible Control of Reproduction In Queens: Mastering the use of reproductive drugs to manipulate cyclicity. J. Feline Med. Surg. 2022, 24, 853–870. [Google Scholar] [CrossRef]
- Romagnoli, S. Progestins to control feline reproduction: Historical abuse of high doses and potentially safe use of low doses. J. Feline Med. Surg. 2015, 17, 743–752. [Google Scholar] [CrossRef]
- Johnston, S.D.; Root Kustritz, M.V.; Olson, P.S. Prevention and termination of feline pregnancy. In Canine and Feline Theriogenology, 1st ed.; Johnston, S.D., Root Kustritz, M.V., Olson, P.S., Eds.; Saunders: Philadelphia, PA, USA, 2001; pp. 447–452. [Google Scholar]
- Graham, L.H.; Swanson, W.F.; Wildt, D.E.; Brown, J.L. Influence of oral melatonin on natural and gonadotropin-induced ovarian function in the domestic cat. Theriogenology 2004, 61, 1061–1076. [Google Scholar] [CrossRef]
- Gimenez, F.; Stornelli, M.C.; Tittarelli, C.M.; Savignone, C.A.; Dorna, I.V.; de la Sota, R.L.; Stornelli, M.A. Suppression of estrus in cats with melatonin implants. Theriogenology 2009, 72, 493–499. [Google Scholar] [CrossRef]
- Schäfer-Somi, S. Effect of melatonin on the reproductive cycle in female cats: A review of clinical experiences and previous studies. J. Feline Med. Surg. 2017, 19, 5–12. [Google Scholar] [CrossRef]
- Furthner, E.; Roos, J.; Niewiadomska, Z.; Maenhoudt, C.; Fontbonne, A. Contraceptive implants used by cat breeders in France: A study of 140 purebred cats. J. Feline Med. Surg. 2020, 22, 984–992. [Google Scholar] [CrossRef]
- Griffin, B.; Heath, A.M.; Young, D.W.; Wright, J.; Rolsma, M.D.; Baker, H.; Ritchey, S. Effects of melatonin implants on ovarian function in the domestic cat. In Proceedings of the 19th American College of Veterinary Internal Medicine, Denver, CO, USA; 2001; p. 843. [Google Scholar]
- Blottner, S.; Jewgenow, K. Moderate seasonality in testis function of domestic cat. Reprod. Domest. Anim. 2007, 42, 536–540. [Google Scholar] [CrossRef]
- Nuñez-Favre, R.; Bonaura, M.C.; Tittarelli, C.M.; Mansilla-Hermann, D.; de la Sota, R.L.; Stornelli, M.A. Effect of natural photoperiod on epididymal sperm quality and testosterone serum concentration in domestic cat (Felis silvestris catus). Reprod. Domest. Anim. 2012, 47 (Suppl. 6), 232–234. [Google Scholar] [CrossRef]
- Ferré-Dolcet, L.; Romagnoli, S. Reversible control of reproduction in tom cats: Medical options for manipulating libido and fertility. J. Feline Med. Surg. 2023, 25, 1098612X231171406. [Google Scholar] [CrossRef]
- Nuñez Favre, R.; Bonaura, M.C.; Tittarelli, C.M.; Stornelli, M.C.; de la Sota, R.L.; Stornelli, M.A. Effect of refractoriness to long photoperiod on sperm production and quality in tomcats. Reprod. Domest. Anim. 2012, 47 (Suppl. 6), 235–237. [Google Scholar] [CrossRef]
- Favre, R.N.; Bonaura, M.C.; Praderio, R.; Stornelli, M.C.; de la Sota, R.L.; Stornelli, M.A. Effect of melatonin implants on spermatogenesis in the domestic cat (Felis silvestris catus). Theriogenology 2014, 82, 851–856. [Google Scholar] [CrossRef]
- Padula, A.M. GnRH analogues—Agonists and antagonists. Anim. Reprod. Sci. 2005, 88, 115–126. [Google Scholar] [CrossRef]
- Fontaine, C. Long-term contraception in a small implant: A review of Suprelorin (deslorelin) studies in cats. J. Feline Med. Surg. 2015, 17, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Borges, P.; Fontaine, E.; Maenhoudt, C.; Payan-Carreira, R.; Santos, N.; Leblond, E.; Fontaine, C.; Fontbonne, A. Fertility in Adult Bitches Previously Treated with a 4.7 mg Subcutaneous Deslorelin Implant. Reprod. Domest. Anim. 2015, 50, 965–971. [Google Scholar] [CrossRef]
- Romagnoli, S.; Baldan, A.; Ferro, S.; Righetti, C.; Scenna, L.; Gabai, G.; Badon, T.; Fontaine, C.; Mollo, A.; Stelletta, C.; et al. Length of efficacy and effect of implant location in adult tom cats treated with a 9.4 mg deslorelin subcutaneous implant. J. Feline Med. Surg. 2019, 21, 507–519. [Google Scholar] [CrossRef]
- Levy, J.K. Contraceptive vaccines for the humane control of community cat populations. Am. J. Reprod. Immunol. 2011, 66, 63–70. [Google Scholar] [CrossRef]
- Munson, L.; Harrenstien, L.A.; Acton, A.E.; Graham, P.A.; Chassy, L.M.; Kirkpatrick, J.F. Immunologic responses and adverse reactions to Freund’s-adjuvanted porcine zona pellucida immuno-contraceptives in domestic cats. Vaccine 2005, 23, 5646–5654. [Google Scholar] [CrossRef]
- Lee, Y.J.; Jo, E.J.; Lee, H.W.; Hwang, B.R.; Kim, Y.H.; Park, B.J.; Cho, Y.J.; Lee, Y.A.; Choi, I.S.; Han, J.S. Evaluation of infertility efficacy of the E. coli expressed STF2-GnRH vaccine in male cats. J. Vet. Sci. 2019, 20, e30. [Google Scholar] [CrossRef]
- Ochoa, J.S.; Favre, R.N.; García, M.F.; Stornelli, M.C.; Sangache, W.C.; Rearte, R.; de la Sota, L.; Stornelli, M.A. Immunocontraception of male domestic cats using GnRH vaccine Improvac. Theriogenology 2023, 198, 211–216. [Google Scholar] [CrossRef]
- Samoylov, A.; Napier, I.; Morrison, N.; Cochran, A.; Schemera, B.; Wright, J.; Cattley, R.; Samoylova, T. DNA Vaccine Targeting Gonadotropin-Releasing Hormone Receptor and Its Application in Animal Contraception. Mol. Biotechnol. 2019, 61, 73–83. [Google Scholar] [CrossRef]
- Dunbar, C.E.; High, K.A.; Joung, J.K.; Kohn, D.B.; Ozawa, K.; Sadelain, M. Gene therapy comes of age. Science 2018, 359, eaan4672. [Google Scholar] [CrossRef]
- Alnasser, S.M. Review on mechanistic strategy of gene therapy in the treatment of disease. Gene 2021, 769, 145246. [Google Scholar] [CrossRef]
- Li, J.; Olvera, A.I.; Akbari, O.S.; Moradian, A.; Sweredoski, M.J.; Hess, S.; Hay, B.A. Vectored antibody gene delivery mediates long-term contraception. Curr. Biol. 2015, 25, R820–R822. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, K.; Suda, T.; Zhang, G.; Liu, D. Advances in Gene Delivery Systems. Pharm. Med. 2011, 25, 293–306. [Google Scholar] [CrossRef]
- Butt, M.H.; Zaman, M.; Ahmad, A.; Khan, R.; Mallhi, T.H.; Hasan, M.M.; Khan, Y.H.; Hafeez, S.; Massoud, E.E.S.; Rahman, M.H.; et al. Appraisal for the Potential of Viral and Nonviral Vectors in Gene Therapy: A Review. Genes 2022, 13, 1370. [Google Scholar] [CrossRef]
- Guo, X.; Huang, L. Recent advances in nonviral vectors for gene delivery. Acc. Chem. Res. 2012, 45, 971–979. [Google Scholar] [CrossRef]
- Wang, C.; Pan, C.; Yong, H.; Wang, F.; Bo, T.; Zhao, Y.; Ma, B.; He, W.; Li, M. Emerging non-viral vectors for gene delivery. J. Nanobiotechnol. 2023, 21, 272. [Google Scholar] [CrossRef]
- Huang, S.; Kamihira, M. Development of hybrid viral vectors for gene therapy. Biotechnol. Adv. 2013, 31, 208–223. [Google Scholar] [CrossRef]
- Mahato, M.; Jayandharan, G.R.; Vemula, P.K. Viral- and Non-viral-Based Hybrid Vectors for Gene Therapy. In Gene and Cell Therapy: Biology and Applications; Jayandharan, G.R., Ed.; Springer: Singapore, 2018; pp. 111–130. [Google Scholar]
- Zhang, J.; Jia, Y.; Li, X.; Hu, Y. Facile engineering of biocompatible materials with pH-modulated degradability. Adv. Mater. 2011, 23, 3035–3040. [Google Scholar] [CrossRef]
- Chen, H.; Liu, X.; Dou, Y.; He, B.; Liu, L.; Wei, Z.; Li, J.; Wang, C.; Mao, C.; Zhang, J.; et al. A pH-responsive cyclodextrin-based hybrid nanosystem as a nonviral vector for gene delivery. Biomaterials 2013, 34, 4159–4172. [Google Scholar] [CrossRef]
- Stribley, J.M.; Rehman, K.S.; Niu, H.; Christman, G.M. Gene therapy and reproductive medicine. Fertil. Steril. 2002, 77, 645–657. [Google Scholar] [CrossRef]
- Anguela, X.M.; High, K.A. Entering the Modern Era of Gene Therapy. Annu. Rev. Med. 2019, 70, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. RNA Viruses as Tools in Gene Therapy and Vaccine Development. Genes 2019, 10, 189. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.K.; Kim, S.W. Recent advances in the development of gene delivery systems. Biomater. Res. 2019, 23, 8. [Google Scholar] [CrossRef]
- Ledley, F.D. Nonviral gene therapy: The promise of genes as pharmaceutical products. Hum. Gene Ther. 1995, 6, 1129–1144. [Google Scholar] [CrossRef]
- Áyen, Á.; Jiménez Martínez, Y.; Marchal, J.A.; Boulaiz, H. Recent Progress in Gene Therapy for Ovarian Cancer. Int. J. Mol. Sci. 2018, 19, 1930. [Google Scholar] [CrossRef]
- Drakopoulou, E.; Anagnou, N.P.; Pappa, K.I. Gene Therapy for Malignant and Benign Gynaecological Disorders: A Systematic Review of an Emerging Success Story. Cancers 2022, 14, 3238. [Google Scholar] [CrossRef]
- Durlinger, A.L.; Visser, J.A.; Themmen, A.P. Regulation of ovarian function: The role of anti-Müllerian hormone. Reproduction 2002, 124, 601–609. [Google Scholar] [CrossRef]
- Sriraman, V.; Niu, E.; Matias, J.R.; Donahoe, P.K.; Maclaughlin, D.T.; Hardy, M.P.; Lee, M.M. Müllerian Inhibiting Substance Inhibits Testosterone Synthesis in Adult Rats. J. Androl. 2001, 22, 750–758. [Google Scholar] [CrossRef]
- Pépin, D.; Sosulski, A.; Zhang, L.; Wang, D.; Vathipadiekal, V.; Hendren, K.; Coletti, C.M.; Yu, A.; Castro, C.M.; Birrer, M.J.; et al. AAV9 delivering a modified human Mullerian inhibiting substance as a gene therapy in patient-derived xenografts of ovarian cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E4418–E4427. [Google Scholar] [CrossRef]
- Kano, M.; Sosulski, A.E.; Zhang, L.; Saatcioglu, H.D.; Wang, D.; Nagykery, N.; Sabatini, M.E.; Gao, G.; Donahoe, P.K.; Pépin, D. AMH/MIS as a contraceptive that protects the ovarian reserve during chemotherapy. Proc. Natl. Acad. Sci. USA 2017, 114, E1688–E1697. [Google Scholar] [CrossRef] [PubMed]
- Hay, B.A.; Li, J.; Guo, M. Vectored gene delivery for lifetime animal contraception: Overview and hurdles to implementation. Theriogenology 2018, 112, 63–74. [Google Scholar] [CrossRef] [PubMed]
- Jensen, R.; Johnson, A.; Wilborn, R.; Braden, T.; Kutzler, M.; Roberts, S.; Van Kampen, K.; Trumble, J.; Baker, H. Adenoviral-vectored gonadotropin releasing hormone vaccine for estrus suppression in mares. Clin. Theriogenol. 2020, 12, 124–133. [Google Scholar]
- Wu, X.; Yang, Y.; Kling, C.; Seigler, L.; Gallardo-Romero, N.F.; Martin, B.E.; Smith, T.G.; Olson, V.A. Inactivated Rabies Virus-Vectored Immunocontraceptive Vaccine in a Thermo-Responsive Hydrogel Induces High and Persistent Antibodies against Rabies, but Insufficient Antibodies against Gonadotropin-Releasing Hormone for Contraception. Vaccines 2019, 7, 73. [Google Scholar] [CrossRef]
- Shahryari, A.; Burtscher, I.; Nazari, Z.; Lickert, H. Engineering Gene Therapy: Advances and Barriers. Adv. Ther. 2021, 4, 2100040. [Google Scholar] [CrossRef]
- Brown, A.M.; Blind, J.; Campbell, K.; Ghosh, S. Safeguards for Using Viral Vector Systems in Human Gene Therapy: A Resource for Biosafety Professionals Mitigating Risks in Health Care Settings. Appl. Biosaf. 2020, 25, 184–193. [Google Scholar] [CrossRef]
- Ghosh, S.; Brown, A.M.; Jenkins, C.; Campbell, K. Viral Vector Systems for Gene Therapy: A Comprehensive Literature Review of Progress and Biosafety Challenges. Appl. Biosaf. 2020, 25, 7–18. [Google Scholar] [CrossRef]
- Bakker, J.M.; Bleeker, W.K.; Parren, P.W. Therapeutic antibody gene transfer: An active approach to passive immunity. Mol. Ther. 2004, 10, 411–416. [Google Scholar] [CrossRef]
- Alton, E.W.F.W.; Boyd, A.C.; Davies, J.C.; Gill, D.R.; Griesenbach, U.; Harman, T.E.; Hyde, S.; McLachlan, G. Gene Therapy for Respiratory Diseases: Progress and a Changing Context. Hum. Gene Ther. 2020, 31, 911–916. [Google Scholar] [CrossRef]
- Iwasaki, A. Immune Regulation of Antibody Access to Neuronal Tissues. Trends Mol. Med. 2017, 23, 227–245. [Google Scholar] [CrossRef]
- Sabatini, M.T.; Chalmers, M. The Cost of Biotech Innovation: Exploring Research and Development Costs of Cell and Gene Therapies. Pharm. Med. 2023, 37, 365–375. [Google Scholar] [CrossRef]
- Editorial, N. The gene-therapy revolution risks stalling if we don’t talk about drug pricing. Nature 2023, 616, 629–630. [Google Scholar] [CrossRef]
Figure 1.
Different types of vectors available in gene therapy.


{kind=link}
Table 1.
Summary of the main disadvantages and side effects associated with non-progestogen based contraceptive therapies.
Table 1.
Summary of the main disadvantages and side effects associated with non-progestogen based contraceptive therapies.
| Treatment | Disadvantages | Reported Side-Effects | Comments |
|---|---|---|---|
| Melatonin implants (18 mg) |
|
|
|
| Slow-release GnRH implants (4.7 mg or 9.4 mg) |
|
|
|
| Immunocontraception |
|
|
|
|
|
|
Table 2.
Summary of the main properties of the most frequently used virus-based gene delivery systems (compiled from [43,51,52,53]).
| Features | Retroviruses | Lentiviruses | Spumaviruses | Adenovirus | Adeno-Associated Adenoviruses |
|---|---|---|---|---|---|
| Viral genome | Single-stranded RNA | Single-stranded RNA | Single-stranded RNA | DNA | DNA |
| Cell division requirements in target cells | Dividing cells | G1 phase | No preferences | No | No |
| Gene loading limitation | 8 kb | 8 kb | 9.2 kb | 8–37 kb | 4–5 kb |
| Immune responses to the vector | Low | Low | Low | Extensiveat the inoculation local | Low |
| Genome integration | Yes | Yes | No preferences | No | Some integration ability |
| Main disadvantages | Random integration Low titers | Random integration Potential for pathogenic vector mutation | Random integration | Transient expression Requests repeated administration | Later onset of expression |
| Main advantages | Persistent gene transfer in dividing cells Long term expression | Broad host range High transduction efficiency Persistent gene transfer in transduced tissues | Highly effective in dividing cells No expression of viral proteins | Highly effective in transducing various tissues Large gene loading capacity | Elicits few inflammatory responses Sustained gene expression Non-pathogenic |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Payan-Carreira, R. Gene-Based Therapy: A New Approach to Feline Induced Sterilization? Future Pharmacol. 2023, 3, 938-950. https://doi.org/10.3390/futurepharmacol3040057
AMA Style
Payan-Carreira R. Gene-Based Therapy: A New Approach to Feline Induced Sterilization? Future Pharmacology. 2023; 3(4):938-950. https://doi.org/10.3390/futurepharmacol3040057
Chicago/Turabian StylePayan-Carreira, Rita. 2023. "Gene-Based Therapy: A New Approach to Feline Induced Sterilization?" Future Pharmacology 3, no. 4: 938-950. https://doi.org/10.3390/futurepharmacol3040057