
Antimicrobial Peptides and Their Assemblies

Biocolloids Laboratory, Biochemistry Department, Chemistry Institute, São Paulo State University, Avenida Professor Lineu Prestes, 748, Butantan, São Paulo 05508-000, Brazil
Future Pharmacol. 2023, 3(4), 763-788; https://doi.org/10.3390/futurepharmacol3040047
Submission received: 15 September 2023
/
Revised: 9 October 2023
/
Accepted: 16 October 2023
/
Published: 19 October 2023
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Antibiotic resistance requires alternatives to fight multi-drug resistant strains. Antimicrobial peptides (AMPs) act by disrupting or solubilizing microbial cell walls or membranes in accordance with mechanisms difficult to counteract from the microbe’s point of view. In this review, structure–activity relationships for AMPs and their assemblies are discussed, considering not only their self-assembly but also their interactions with their carriers for optimal delivery or their combinations with other complementary antimicrobials or moieties covalently bound to their chemical structure. The effect of the formulations on AMP activity is also evaluated, revealing a myriad of possibilities. Depending on the interaction forces between the AMP, the carrier, or the elements added to the formulations, AMP activity can be reduced, enhanced, or remain unaffected. Approaches protecting AMPs against proteolysis may also reduce their activity.
1. Introducing Antimicrobial Peptides in Pharmacology
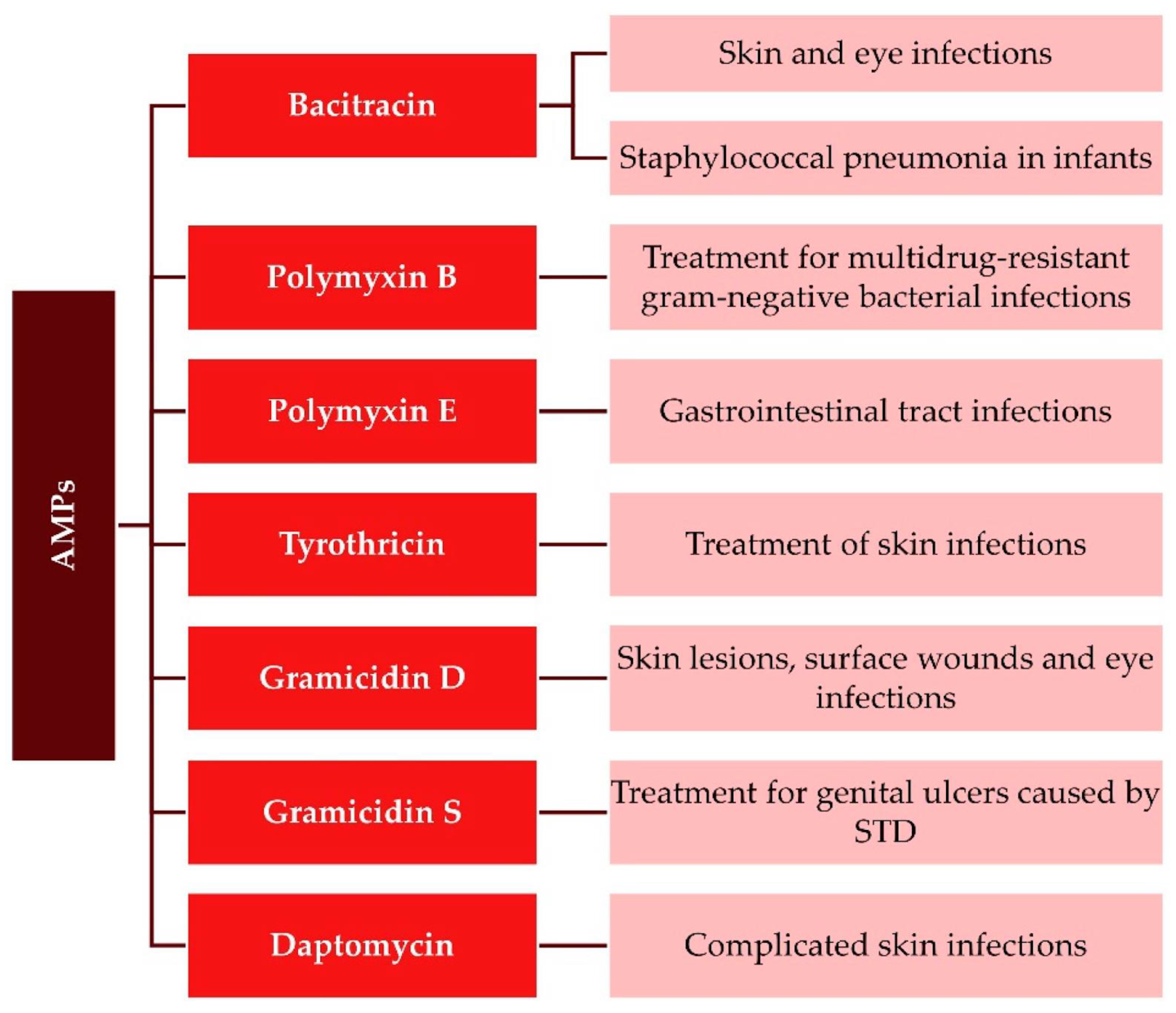
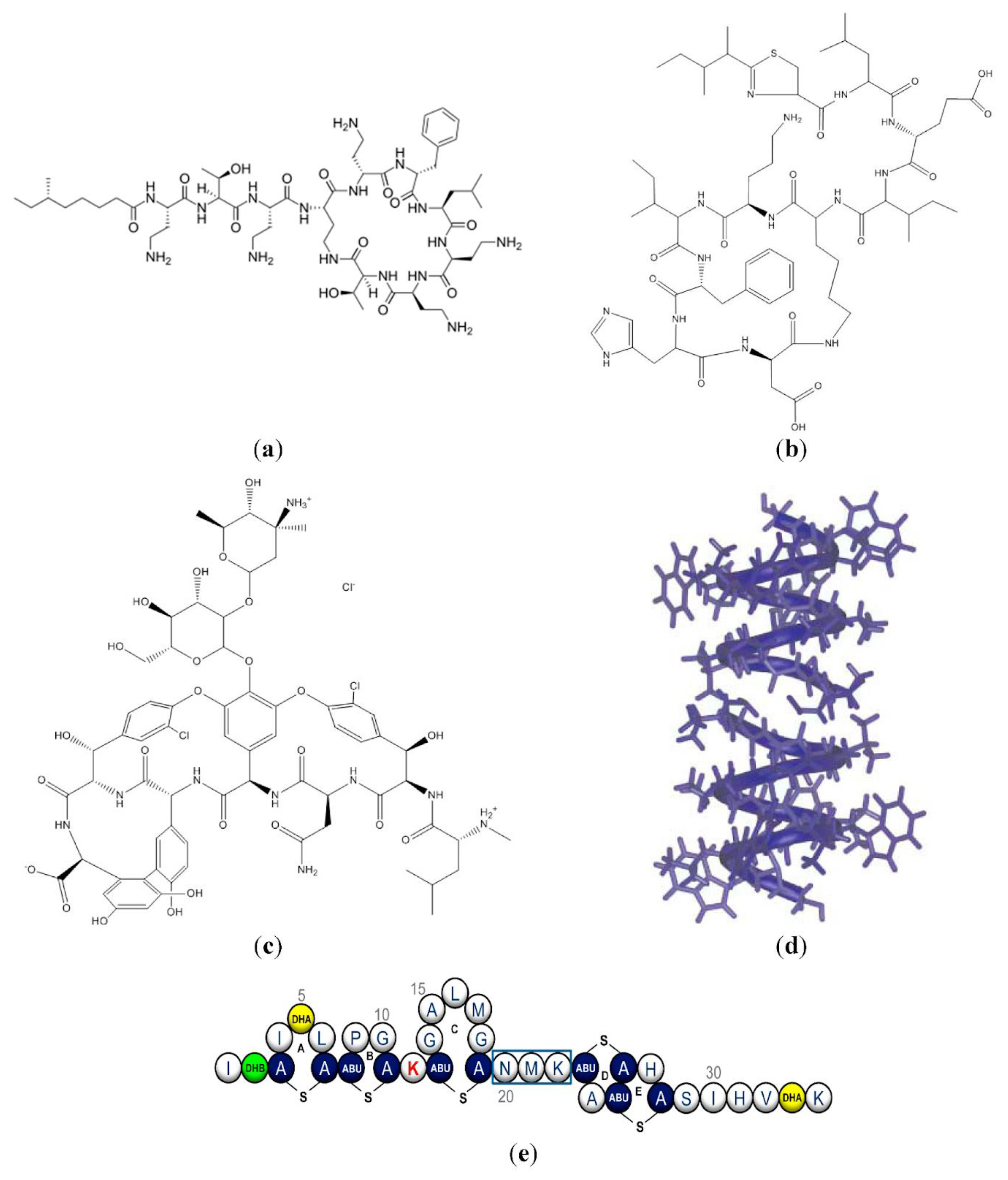
Antimicrobial peptides (AMPs) are ubiquitous in living organisms and are considered a potentially valuable tool against microbial resistance to antibiotics [1,2,3]. Although the number of natural or synthetic AMPs is ever increasing, with more than three thousand reported, only a few were approved by the Food and Drug Administration (FDA) for use in the clinic [4,5]; the approved ones are gramicidins [6], polymyxins [7], and nisins [8,9]. Highly modified AMPs such as daptomycin (lipopeptide) [10], bacitracin (cyclic peptide) [11], and gramicidin S (cyclic peptide) [12] are also in current clinical use [13]. Figure 1 illustrates approved AMPs and their clinical uses [4].
Gramicidins are valuable for dermal surface wounds and infections of the upper respiratory system; polymyxins can treat ocular and gastrointestinal infections and systemic infections by methicillin-resistant Gram-negative bacteria; daptomycin can treat complicated skin infections by Staphylococcus aureus and is often used in combination therapy to improve treatment efficacy. Nisin is the most investigated AMP; as a bacteriocin, it is produced by bacteria that kill other bacteria. Besides its applications in food preservation, other potential therapeutic applications have been described in dental care, stomach ulcer therapy, and the treatment of colonic infections in humans [8,9]. Although most of the currently reported AMPs are either antibacterial or antifungal peptides, some of them have been formulated against viruses, e.g., influenza A virus, severe acute respiratory syndrome coronavirus (SARS-CoV), HIV, HSV, West Nile Virus (WNV), and others; gramicidin S and melittin as antivirals can treat SARS-CoV-2 infections [14]. AMPs’ antiviral activities have also been recently reviewed [4,15,16].
AMPs generally are 10–40 amino acids long and bear a net positive charge; many also have hydrophobic moieties allowing helical peptide conformations upon binding to bacteria membranes so that they cause membrane disruption [17,18]. However, effective and safe therapeutics require not only lysis of the pathogen cell membrane but also the absence, or at least minimal rupture of the cellular membrane of the host; in addition, AMPs can interact with membranes as monomers or as oligomers; peptide oligomerization has been often reported to influence AMPs’ interaction with membranes and their antimicrobial activity [17,19,20,21].
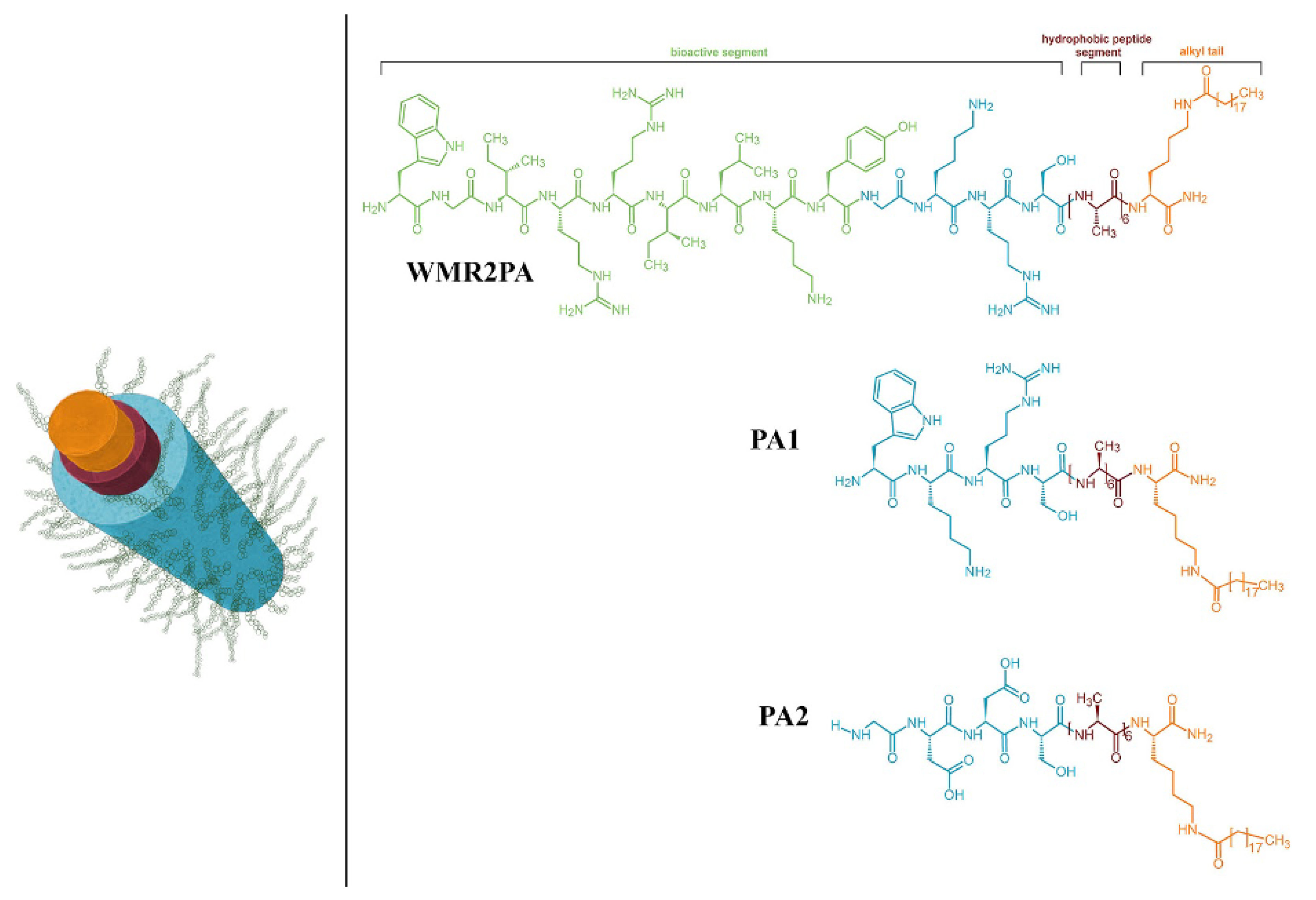
Numerous natural or synthetic AMPs self-assemble into supramolecular structures with intermolecular interactions driven by hydrophobic interactions between hydrophobic residues in adjacent side chains, intermolecular hydrogen bonding, aromatic moieties stacking, etc., forming a variety of nanostructures such as the nanoparticles (NPs) of gramicidin D [21] or nanofibers of synthetic WMR peptides engineered with moieties able to impart WMR self-assembly into stable nanofibers [20]. Figure 2 illustrates the engineered WMR peptide self-assembly, where the covalent linkage of aliphatic residues to the amino group of a terminal lysine generated the WMR peptide amphiphile (WMR PA). The self-assembly enhanced the potency of the AMP, allowing the eradication of biofilms of Pseudomonas aeruginosa and Candida albicans [20].
The cooperative action of hydrogen bonding, hydrophobic interaction, and electrostatic attraction drive the formation of self-assembled nanostructures of AMPs; properties such as hydrophobicity, aromaticity, geometry, charge, and isoelectric point of various amino acid residues determine AMPs’ secondary structures, allowing the formation of self-assembled nanostructures by carefully engineering amino acids sequences in synthetic AMPs [17,20,22,23].
The clinical applications for AMPs will possibly rely on overcoming the poor stability of natural AMPs and their toxic effects on the patient, e.g., hemolysis; therefore, developing synthetic and long-lasting AMP derivatives has been considered a powerful strategy for the future pharmacology of AMPs [24,25]. For example, antimicrobial peptide–polymer conjugates [24,26] not only preserve excellent antimicrobial activity but also show reduced toxicity, offering more functionalities as those represented by biomedical materials such as hydrogels [27], polymeric materials with applications in wound dressings [28], orthopedic implant coatings [29], antibiofilm materials [30,31], engineered tissues, etc. [32,33,34,35,36]. The covalent linkage of AMPs to polymers via labile bonds such as ester, amide, ether, or anhydride bonds imparts biodegradability to the conjugates [26].
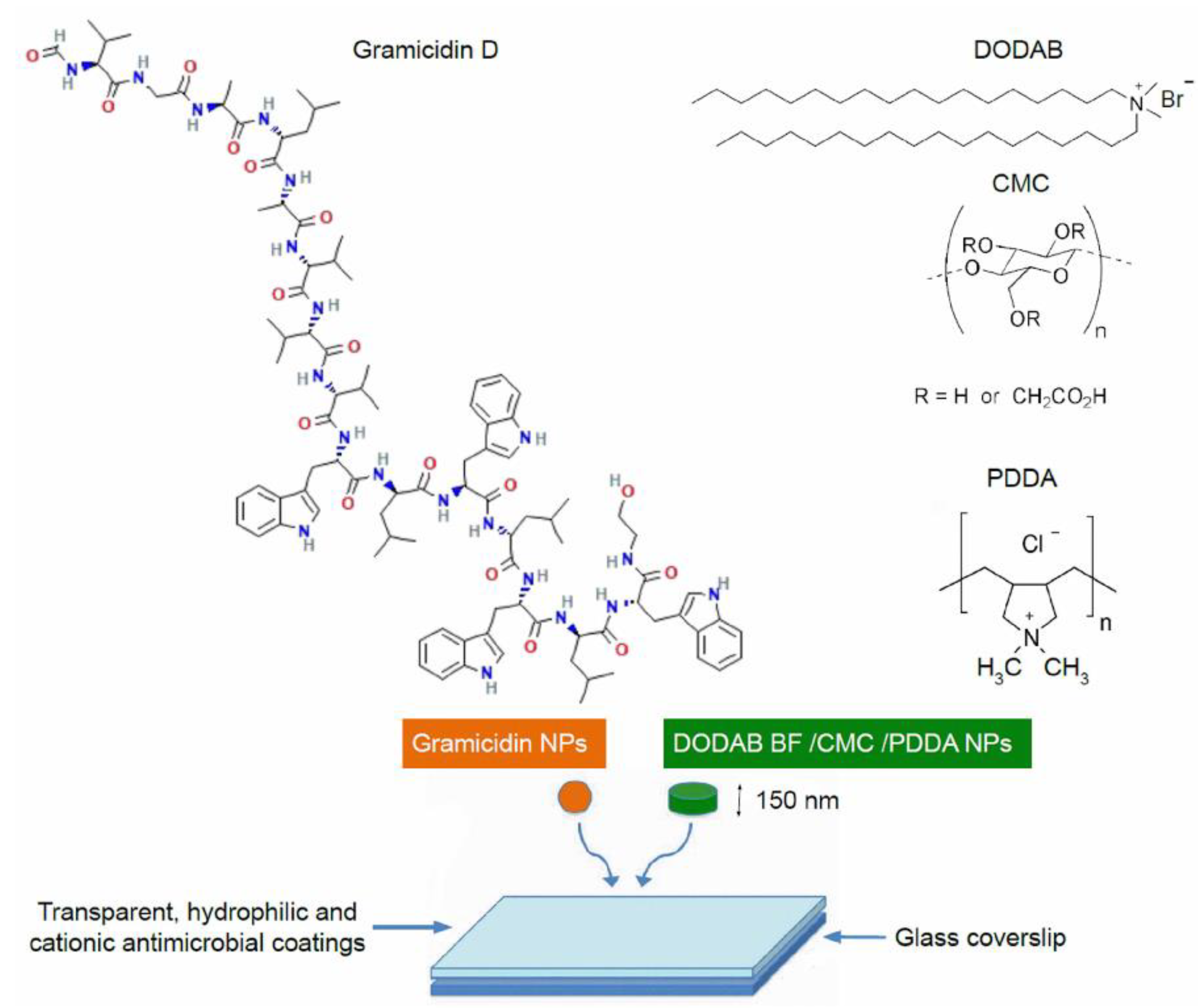
Nanotechnology has also been providing promising approaches for formulating and delivering AMPs [22,36,37,38,39]. AMPs can be protected by nanostructures, having their low stability circumvented and their release controlled by appropriate pharmacodynamics; thereby, antimicrobial activity, retention, and biosafety could be enhanced [40,41]. Approaches combining AMPs with antimicrobial nanomaterials may also lead to synergism and a broad spectrum of activity [21,42]. Figure 3 illustrates the preparation of a broad-spectrum and potent antimicrobial coating based on nanoparticles (NPs) [21,42]; NPs employed were obtained from the self-assembly of gramicidin D (Gr) in a water solution (spherical Gr NPs colored in orange) [21] and antimicrobial cationic NPs based on cationic bilayer fragments/carboxymethylcellulose/poly(diallyl dimethylammonium) chloride (colored in green) [43,44,45,46,47]. The combinations of gramicidin D and the antimicrobial polymer poly(diallyl dimethylammonium) chloride (PDDA) completely killed Gram-negative, Gram-positive, and fungus in vitro at reduced doses of both antimicrobial agents when dispersed as nanoparticles [21] or as coatings obtained by casting and drying the nanoparticles on surfaces [42].
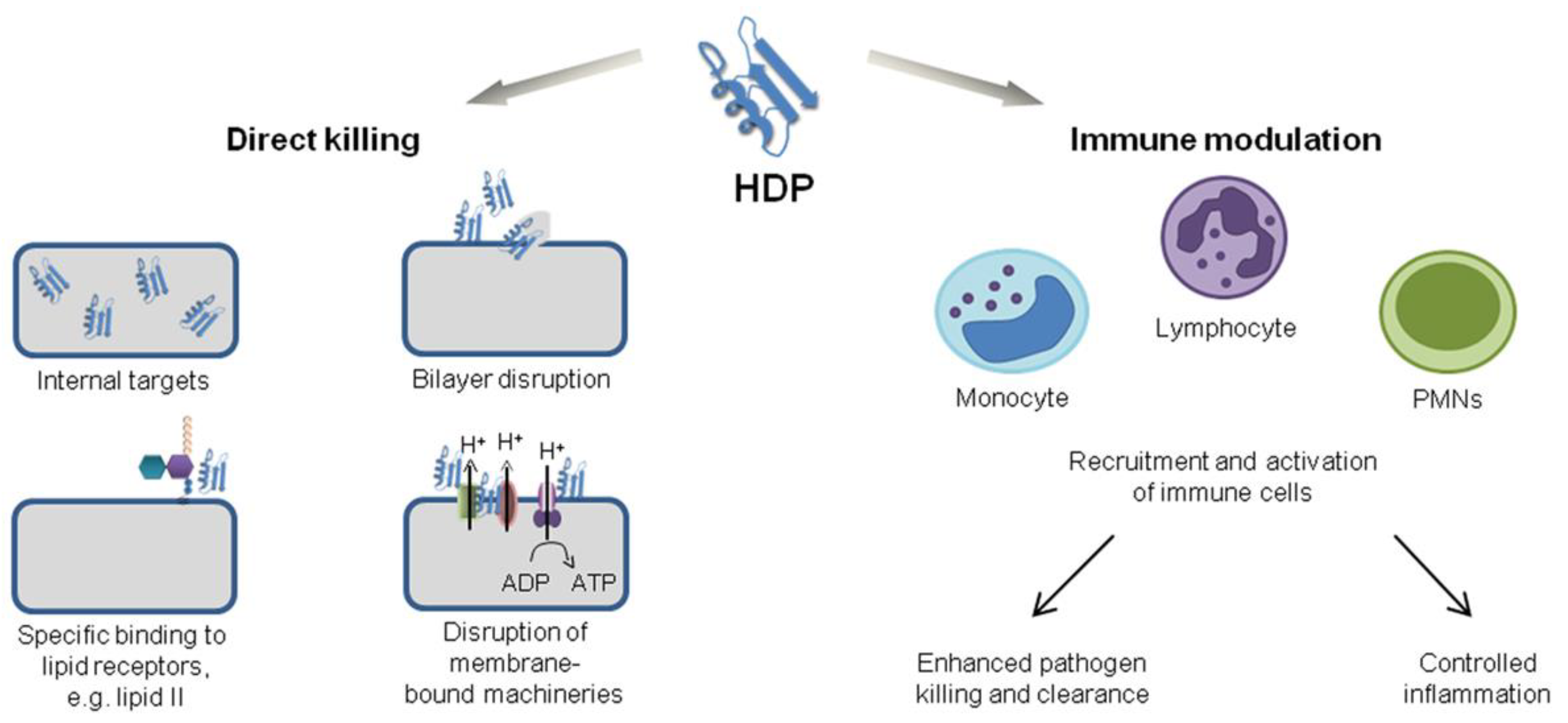
The clinical potential of cationic antimicrobial host defense peptides (CHDP) was recently reviewed; CHDP can control infections not only by their antimicrobial properties but also by modulating host immune response and limiting inflammation [48]. For example, one of the main undesirable side effects of corticosteroids currently used against inflammation is increasing the patient’s susceptibility to infections [49]; alternatively, CHDP can both kill pathogens and control harmful inflammation due to their direct microbicidal properties and/or ability to modulate the host immune responses. Since their first report in the 1960s, CDHP expression has been found across a broad diversity of species ranging from microorganisms, plants, and invertebrates to more complex amphibians and mammals [48]. Nowadays, it is accepted that CHDP has a critical role in immunity, from activation of innate immunity and enhancement in antigen presentation and phagocytosis to effects on adaptive immunity and memory functions, with additional and potent anti-inflammatory activity. Intense research in this field on developing CHDP as drugs may lead to clinical applications such as the control of antibiotic-resistant pathogens [50,51,52], chronic inflammation in colitis [53], asthma [54], arthritis [55], and cancer [56,57,58,59].
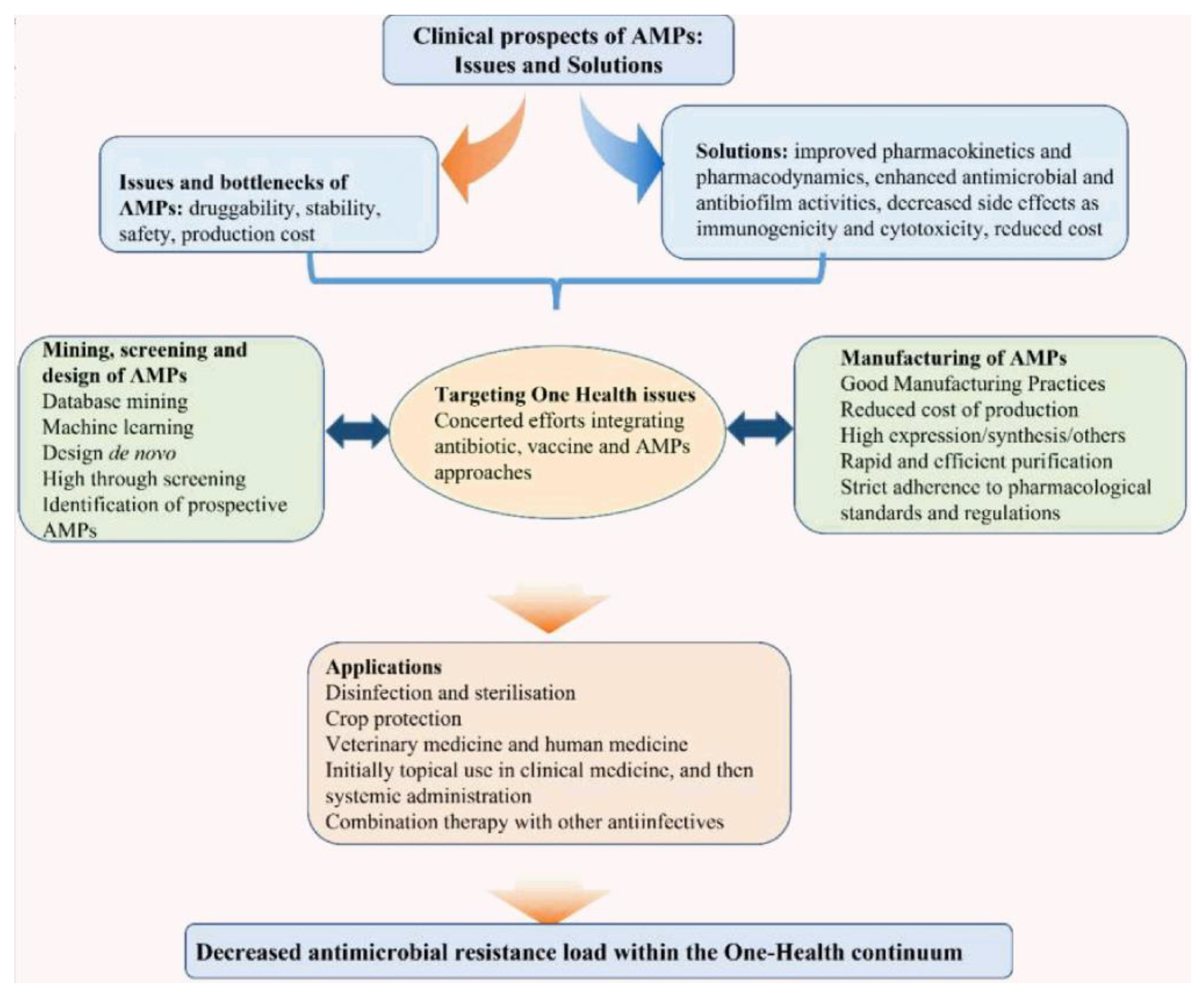
The major issues and possible solutions involved in bringing AMPs to the clinic have been recently reviewed, as schematically illustrated in Figure 4 [60]. AMPs’ drawbacks hampering their clinical use as drugs are related to their degradation by proteases, their cytotoxicity against the host cells, their immunogenicity, and finally, their poor bioavailability derived from their rapid metabolization by the liver and clearance by the kidneys [61,62].
2. Chemical Structure and Mode of Action for AMPs
The structures of AMPs can be found in an open access database named The Database of Antimicrobial Activity and Structure of Peptides (DBAASP, https://dbaasp.org/), which informs on the amino acid sequences, 3D structures, bioactivities, and toxicities of AMPs. Besides overviewing the structure–function of natural peptides here, we also review some interesting examples of synthetic peptoids [63]. A few natural AMPs, such as polymyxin, LL-37, and gramicidin D, were reported to self-assemble in a water solution; recently, this property was also ascribed to some synthetic peptoids mimicking natural peptides but designed for improved stability regarding proteolysis [21,63]. The chemical structure of some AMPs in clinical use is illustrated in Figure 5, taken from [37], with structures for polymyxin B [7], bacitracin A (cyclic peptide) [11], gramicidin A [64], and nisin [65]. Daptomycin (lipopeptide) [10] and gramicidin S (cyclic peptide) structures were not included in Figure 5 [12].
Polymyxins are cyclic AMPs synthesized by Paenibacillus polymyxa, which are not only active against MDR Gram-negative Pseudomonas aeruginosa and Escherichia coli but are also able to prevent endotoxin (lipopolysaccharides (LPS) from Gram-negative bacteria) toxicity in mice [66]. Hemoperfusion using polymyxin B-immobilized fiber column has been used to treat septic shock in humans [67]. In addition, this same treatment was also used with positive results for severe cases of COVID-19 progressing to organ dysfunction [67,68]; there was organ function recovery, hemodynamic improvement, and endotoxin level reduction [69]. At the core of these medical applications is the mechanism of action for the polymyxin AMPs. These AMPs display a high affinity for the endotoxins (LPS), including lipid A, due to multiple binding sites of the polycationic AMP to LPS phosphate moieties [70,71]. This binding facilitates the insertion of the AMP in the microbial membrane, increasing its permeability via a surfactant-like mode of action so that transient fluctuations of permeability in LPS/phospholipid model bilayers occur upon binding of polymyxin B [72]. Soon after their discovery, reversible nephrotoxicity was reported for different polymyxins; polymyxin B and colistin (polymyxin E) with similar antibacterial activity were the least nephrotoxic in studies using in vivo models [73]. In a recent review article on polymyxins, which is very worth reading, the authors conclude that the need to save lives from the deadly infections caused by Gram-negative “superbugs” justifies major efforts to improve the clinical utility of the last-line polymyxins, which are considered the last-resort option for treatment due to their adverse side effects [74].
Bacitracin A (Figure 5b) is the most well-known member of a family of structurally similar macrocyclic peptides produced by Bacillus lichenformis and Bacillus subtilis; it is active against Gram-positive bacteria, but its systemic toxicity has restricted its use to topical formulations [11]. Bacitracin’s antimicrobial properties derive from its ability to compromise the integrity of the bacterial envelope; it binds undecaprenyl pyrophosphate, a lipid carrier that shuttles cell wall biosynthetic intermediates from the cell’s cytoplasm to its exterior [75]. Thereby, the flow of peptidoglycan precursors to the site of cell wall biosynthesis is interrupted, ultimately leading to cell death [76,77]. Bacitracin can also bind a variety of metals, but the zinc form of bacitracin strongly supports the ternary complex of bacitracin–metal–lipid [78].
Daptomycin, gramicidin, and colistin (polymyxin E) are approved for clinical use and are AMPs acting on the microbe’s membrane [79].
Daptomycin is a last-line-of-defense AMP for treating Gram-positive infections; it possibly acts by forming transient pores in the target membranes, causing bacterial membrane depolarization through ion leakage [79]. However, its use has already created bacterial resistance. To search for its substitutes that might counteract the resistance, its molecular mechanism of action still requires further clarification. Daptomycin (Dap) forms a unique complex with calcium ions and phosphatidylglycerol (PG) molecules in the membrane at a specific stoichiometric ratio: Dap2Ca3PG2. The biophysics of peptide–membrane interaction was recently investigated using an interesting approach combining mass spectrometry (MS) and fast photochemical oxidation of peptides (FPOP). The daptomycin–membrane interactions with different lipid bilayer nanodiscs were studied; MS suggested that daptomycin incorporated randomly and did not prefer any specific oligomeric states when integrated into bilayers, whereas FPOP revealed significant protection in most bilayer environments; the stronger membrane interactions occurred with more rigid membranes, and pore formation took place in more fluid membranes exposing daptomycin to FPOP oxidation; membrane conductance supported the observation of polydisperse pore complexes depicted from the MS analysis [80].
Vancomycin (Figure 5c) is a glycopeptide that has been a life-saving treatment against multidrug-resistant Gram-positive infections [81,82,83,84,85]. For decades, the treatment of serious Gram-positive infections relied upon vancomycin as a last line of defense. However, the emergence of vancomycin resistance highlighted the importance of developing semisynthetic glycopeptides with enhanced antibacterial activities and improved safety profiles, as recently reviewed in [86]. Vancomycin acts by binding to L-aa-D-Ala-D-Ala groups in wall intermediates, inhibiting, by steric hindrance, the formation of the backbone glycan chains in a reaction catalyzed by peptidoglycan polymerase; the subsequent transpeptidation reaction that imparts rigidity to the cell wall also becomes inhibited; consequently, the active sites of two central enzymes in the construction of the cell wall cannot align themselves correctly, rendering the acquisition of resistance to the glycopeptide antibiotics more difficult [87,88].
Gramicidins (Gr) synthesized by Bacillus brevis are 15 amino acid residues long and come as a mixture of gramicidin A (80%), B (5%), and C (15%) known as gramicidin D; Gr A, B, and C differ in the 11th amino acid residue. Gr’s poor solubility in water is ascribed to its composition of hydrophobic amino acids and absence of charged side chains; this also imparts to Gr strong hydrophobic interaction with bilayer membranes. Gr approval by the FDA took place in 1955; it was first commercialized as Neosporin® for the control of ophthalmic Gram-positive infections. This pharmaceutical product also contains polymyxin B, a cyclic lipopeptide [89] (its chemical structure is shown in Figure 5a). Recently, we described the self-assembly of gramicidin D as spherical nanoparticles (NPs) in water solution; these nanostructures are finding interesting applications as water dispersions or coatings characterized by potent and broad activity against bacteria and fungus when combined with additional antimicrobials [21,42]. In these combinations, Gr action depends on insertion, dimeric channel formation, and an increase in ion permeabilization through the microbe’s membrane (see Figure 5d to visualize the Gr dimeric channel in the microbial cell membrane). Gr action was facilitated by a second very important component in the formulations, the potent antimicrobial and cationic polymer poly (diallyl dimethyl ammonium) bromide (PDDA) [44,45,47,90,91,92,93]. PDDA’s strong interaction with the microbial cell wall paved the way for cell wall disassembly and direct Gr insertion in the microbe’s cell membrane; as a consequence, synergic action in the combination Gof r/PDDA reduced required doses and broadened the spectra of action for both antimicrobials against bacteria and fungus [21,42].
Nisin’s molecular structure is shown in Figure 5e; it is produced by lactic acid bacteria and acts by interacting with lipid II, an essential precursor for the biosynthesis of cell walls. In addition, nisin forms pores responsible for cell lysis; thereby, nisin activity extends to both Gram-positive and -negative bacteria, finding applications in food preservation, veterinary, pharmaceutical, and healthcare devices [94]. Recently, the emerging role of nisin and its derivatives against MRSA was reviewed, reporting an increasing number of studies showing how microorganisms that produce nisin act as probiotics against MRSA [95]. In an interesting study in hospitalized humans, it was observed that microbiota-derived lantibiotics similar to nisin could reverse resistance in vancomycin-resistant Enterococcus faecium, a leading cause of hospital infection [96]. Consuming live microorganisms such as Lactobacillus lactis, able to produce nisin A, decreased infections and mortality in intensive care units [97,98]. The pore-forming ability of nisin was shown to depend on the lipid microenvironment, with the lipid cardiolipin facilitating pore formation in mitochondria, possibly leading to cell death and applications against cancer; small natural molecules, phloretin and capsaicin, were also found to potentiate the membrane activity of nisin [99].
In a recent review of clinical trials using AMPs, the structure–function relationship, activity, and toxicity were discussed and updated [100].
AMPs affect microorganisms by piercing cell walls and membranes; by inhibiting enzyme, DNA, or protein synthesis; and by modulating immune responses (Figure 6) [103]. It is broadly recognized that AMPs have great potential as antimicrobials, antibiofilm, and anti-inflammatory agents. Among all AMPs, those with anti-cancer potential derived from their pore-forming ability certainly deserve a special bibliographic search; as a case study, in the next section, we investigated the potential of gramicidin D.
3. Gramicidin D and Its Assemblies
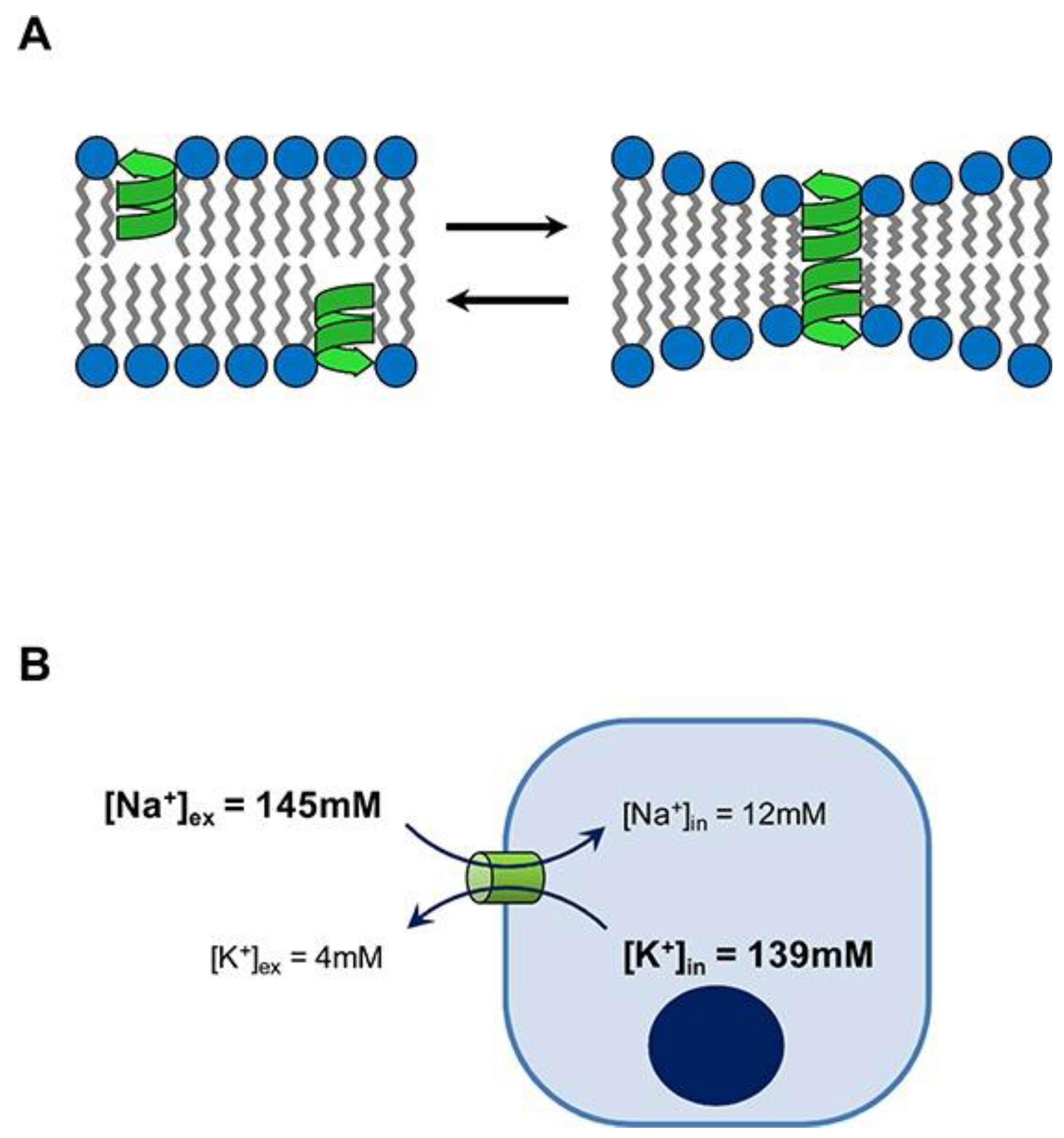
Linear gramicidins have been extensively used to study the organization, dynamics, and function of membrane-spanning channels due to their self-assembling as dimeric channels in bilayers, model membranes, and natural membranes [104,105]. They are pentadecapeptides (molecular weight of about 1900) consisting of alternating L- and D-amino acids [106]. Their cation-selective channels in model bilayer membranes exhibit conductances of about 107 ions per second [107]. Natural gramicidins (gramicidin D) consist of ∼85% gramicidin A, which has four tryptophan residues at positions 9, 11, 13, and 15; they are intrinsically fluorescent due to their tryptophan residues [108]. Their hydrophobic side chains impart to them not only low solubility in water but also their self-assembly as spherical nanoparticles in water solutions [21,42].
In our lab, we have been describing novel assemblies for gramicidins (Gr) and believe they may become important not only for improving Gr antimicrobial performance but also for developing novel anti-cancer assemblies. Dissipation of ion concentration gradients across cell membranes hamper cells from properly regulating their intracellular environment. The potential anticancer properties of channel formers have been largely overlooked. Thus, in this section, we overview Gr and its assemblies, hoping to reveal their potential for novel applications. Figure 7 illustrates Gr’s mode of action [109].
Several reports emphasize AMPs’ unexplored potential for novel applications, including against cancer [110,111,112]. Gramicidin A not only dissipates ion gradients across cancer cells but also becomes inserted in the inner mitochondrial membrane, diminishing the H+ gradient and hampering ATP synthesis; there was a cytostatic action at 1 nM Gr against the human breast cancer cell line MCF-7. Mitochondria malfunction induced mitophagy, and a lack of ATP caused cell cycle arrest [113]. In another study, similar results were obtained against gastric cancer cells; Gr inhibited the proliferation of human gastric cancer cells and the cell cycle and induced apoptosis [114].
Recently, glycoside–Gr A peptide conjugates were prepared; a galactose moiety was linked covalently to Gr A so that the conjugate became inserted in membranes of liver cancer cells due to recognition by the Galactose–asialoglycoprotein receptor; thereby, cancer cells died via apoptosis [110].
Cancer stem cells (CSCs) have unique properties, such as a lack of differentiation, self-renewal capability, pluripotency, resistance to chemo- and radiotherapy, and higher tumorigenicity in comparison with the general population of cancer cells; CSCs in pancreatic ductal adenocarcinoma contributes to an aggressive prognosis for this cancer and treatment resistance [115]. The effect of Gr A at 0.05 μM on pancreatic cancer stem cells was tumor sphere disintegration and a reduction in cell counting; there was CD47 down-regulation and modulation of macrophage/tumor cell interaction [116].
In another instance, Gr A reduced the proliferation potential of acute promyelocytic leukemia (APL) and chronic myeloid leukemia (CML) cell lines without any toxicity; in combination with other anticancer drugs, there was an additional anti-cancer effect related to down-regulation of oncogenes, such as c-Myc, Eya3, and Axin 2; furthermore, Gr A did not induce any hemolysis of red blood cells, which shows its safety with no toxicity [117].
The potential use of a variety of AMPs as novel therapeutic agents against gastrointestinal cancers was recently reviewed [118].
Recently, tyrosine kinase inhibitors, such as the natural product tannic acid and the AMP gramicidin A, were shown to bind strongly to four different tyrosine kinases (ALK, TRK, MET, and ABL), thereby reducing the viability of non-small lung cancer cells (A549 cells) in a concentration-dependent manner and improving the quality of life of cancer patients [119]. Gr A also inhibits cholangiocarcinoma cell growth and induces necrotic cell death, providing a treatment against bile duct solid carcinoma progression [120].
Although AMPs, in general, and Gr A, in particular, are promising anticancer agents, they still display disadvantages, such as low proteolytic and chemical stability, high cytotoxicity and hemolytic activity, and salt sensitivity, inhibiting their clinical application; these drawbacks need to be circumvented, and several strategies have been proposed. For example, structural modifications, e.g., alteration of amino acids, conjugation, or ligation, have been proposed; in addition, nanotechnology is an effective strategy to improve AMPs’ functionality and minimize or eliminate their side effects [121].
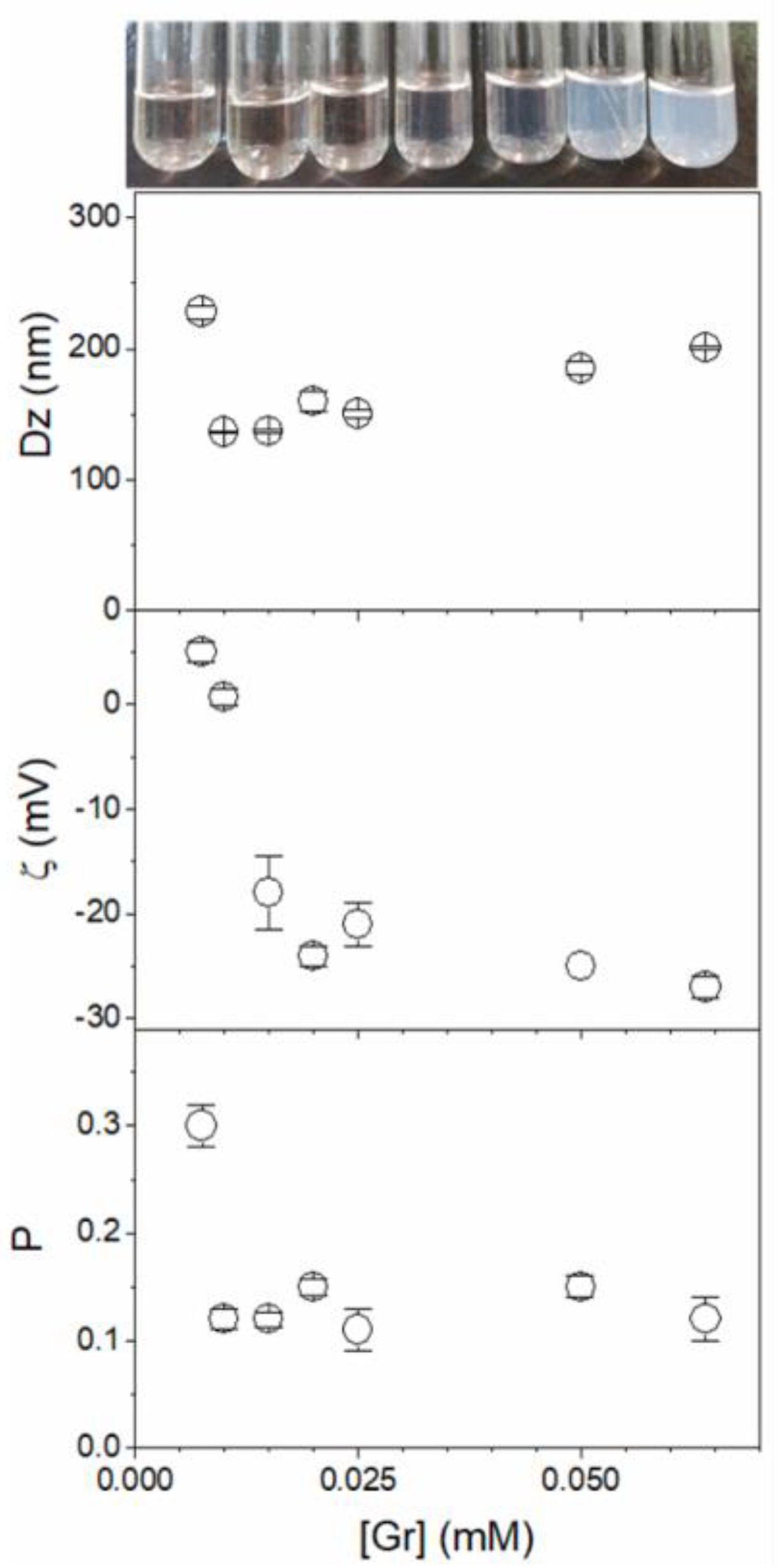
As an interesting example of assemblies or combinations involving Gr A, a recent development was the basic demonstration that Gr self-assembles in water dispersions as nanoparticles (NPs) over a range of Gr concentrations so that inner molecules in the NPs gain protection from the outer medium [21]. Figure 8 shows Gr NPs visualized by means of scanning electron microscopy (SEM); their photos in water dispersions; and their hydrodynamic diameters (Dz), zeta-potentials, and polydispersity (P) over a range of Gr concentrations, as determined via dynamic light scattering (DLS) [21].
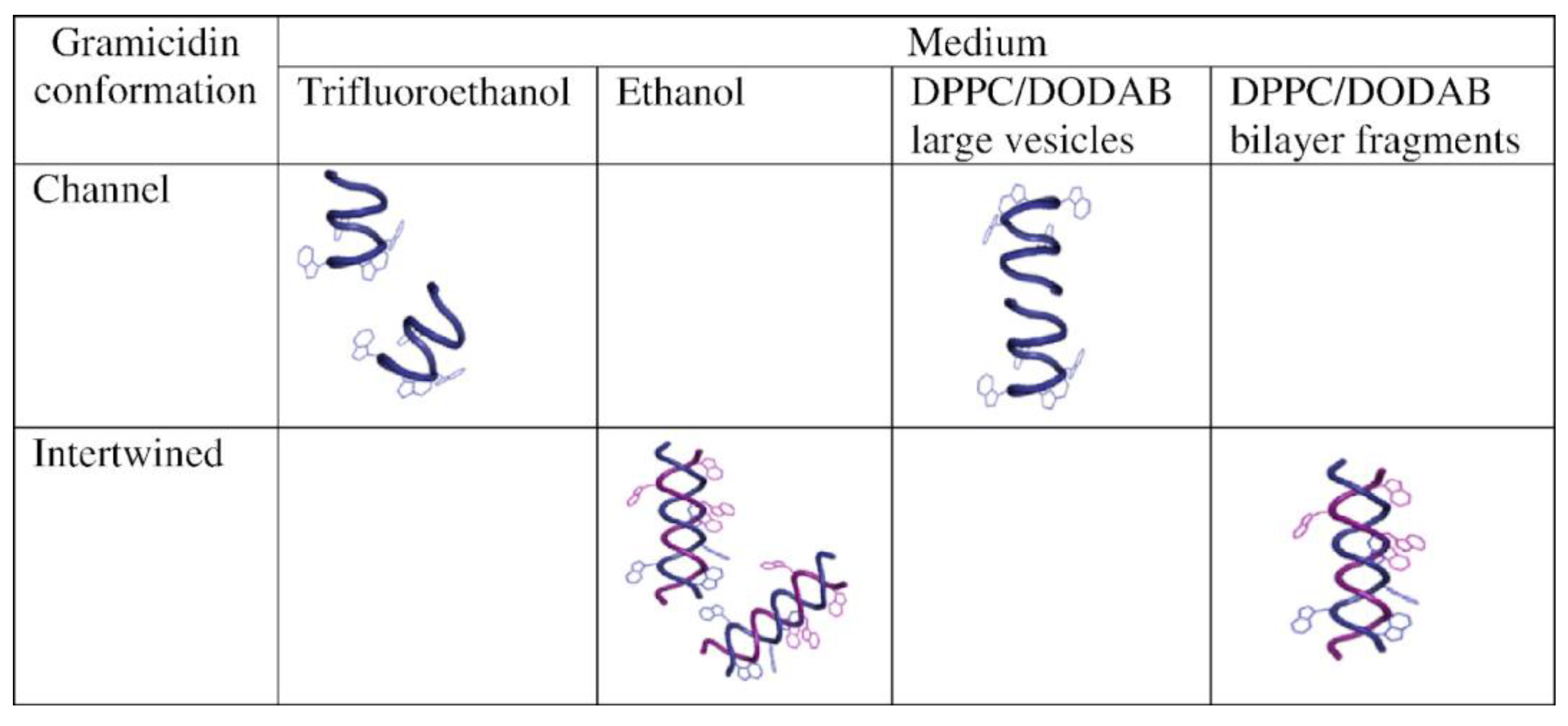
The interaction between Gr and dipalmitoylphosphatidylcholine (DPPC)/dioctadecyldimethylammonium bromide (DODAB) 1:1 large unilamellar vesicles (LVs) or bilayer fragments (BFs) was studied, revealing that the Gr dimeric channel occurred in LVs, whereas Gr in BFs displayed an intertwined dimeric, non-channel Gr conformation; this example illustrated Gr conformations assumed in different media, such as bilayer vesicles or bilayer fragments (see Figure 9) [122].
For the DPPC/DODAB 1:1 LV dispersions in water submitted to a 50 mM KCl or 100 mM glucose water solution, the turbidity at 400 nm increased as a function of time due to LV shrinkage caused by the water efflux from LV, in accordance with the solute gradient [122]; however, incorporation of the Gr channel in the LV bilayer dissipated the solute gradient, resulting in LV swelling and turbidity decrease kinetics due to solute entrance through the Gr channel accompanied by water [122].
The cationic lipid DODAB by itself assembles in water as large vesicles (LV) [123,124] or bilayer fragments (BF) [125,126,127,128,129,130] and displays antimicrobial activity against Gram-negative bacteria but poor activity against Candida albicans or S. aureus [131,132,133,134]; DODAB combinations with Gr proved to be advantageous [135]. The antimicrobial activity of DODAB, Gr, and DODAB/Gr assemblies against E. coli, S. aureus, and S. cerevisiae revealed the advantages of the combinations. There was indeed high activity of DODAB against E. coli [131,132] but poor activity against the yeast [134,136], which was complemented by the microbicidal activity of Gr against S. aureus in the absence of toxicity against the eukaryotic S. cerevisiae; therefore, the novelty of these combinations was the broadening of antimicrobial activity to encompass E. coli and S. aureus as representatives of Gram-negative and Gram-positive bacteria; in addition, the microbicidal activity occurred over a range of low Gr and DODAB concentrations that were not toxic to S. cerevisiae {135].
In another instance, Gr functional channels combined with DODAB-supported bilayers were characterized and tested against Escherichia coli and Staphylococcus aureus; firstly, nanoparticles of polystyrene sulfate (PSS) were covered by a cationic lipid bilayer of dioctadecyl dimethylammonium bromide (DODAB) incorporating Gr; the adsorption of the DODAB/Gr bilayer onto PSS nanoparticles (NPs) increased the zeta-average diameter by 8–10 nm, changed the zeta-potential of the NPs from negative to positive, and yielded narrow size distributions for the PSS/DODAB/Gr NPs, which displayed broad and maximal microbicidal activity at very small concentrations of the antimicrobials, namely, 0.057 and 0.0057 mM DODAB and Gr, respectively. In this case, the effective concentrations in the nanostructured assemblies not only protected Gr from the outer medium but also achieved a high organization level, minimizing the microbicidal dose of DODAB required [137]. Self-assembled antimicrobial nanomaterials, in general, and nanoparticles, in particular, with emphasis on biomimetic ones [138,139,140,141,142], will become very important in drug delivery against pathogens; peptide-based drugs, such as antimicrobial peptides, cyclopeptides, and glycopeptides, play important roles in the treatment of drug-resistant bacterial infections due to their unique lower resistance antibacterial mechanism [38,143]. In the next section, recent advances in formulations for AMPs are discussed.
4. Recent Formulations for Antimicrobial Peptides
Biomedical devices such as catheters, stents, pacemakers, prosthetic joints, implants, and orthopedic devices are foreign materials prone to microbial adhesion and infections, which may increase patient mortality; AMPs as alternatives to antibiotics are less prone to resistance development. Furthermore, peptide drugs are not limited to AMPs, and reviewing their formulations in general can contribute to our understanding of important requirements for formulating AMPs. A major review recently discussed successful formulations for peptides; therefore, formulations for AMPs could benefit enormously from a better understanding of the general essentials for peptide drugs [144]. As a good example, insulin therapy started in the 1920s, becoming a landmark for the development of more than 60 peptide drugs approved by the FDA [145]. As a therapeutic peptide, insulin requires administration routes other than the parenteral one; several novel formulations for this peptide hormone are under clinical trials aiming at more suitable administration routes. Oral insulin delivery has been attempted using lipid-based, synthetic polymer-based, and polysaccharide-based nano/microparticle formulations. Although insulin-transporting particles may preserve insulin in the acidic and enzymatic medium and decrease peptide degradation, in vivo results revealed a lower ability of formulations to reduce glucose in the blood than the subcutaneous form despite promising in vitro results [146].
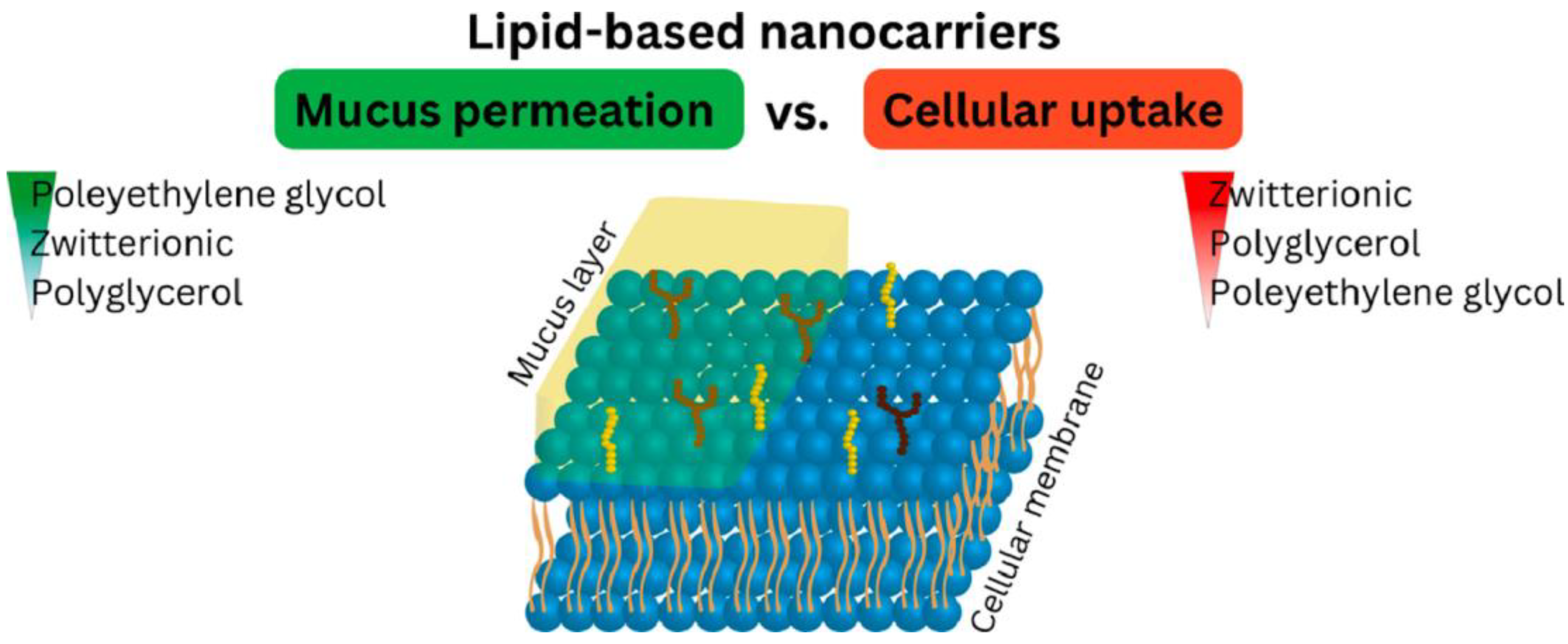
The use of advanced formulations involving micro/nanoparticles [135,137,147,148,149], hydrogels [150,151], particle/hydrogel combinations [152], and polymer-based scaffolds could protect peptides from proteolysis, maintaining their activity [153,154]. Oral peptide delivery using liposomes, polymeric nanoparticles, polysaccharides, hydrogels, and self-emulsifying systems as potential therapeutic approaches have been comprehensively reviewed [155,156]. Among the most promising formulations are lipid-based nanocarriers such as emulsions, lipid nanoparticles, liposomes, and micelles because gastrointestinal (GI) peptidases, as well as sulfhydryl compounds such as glutathione and dietary proteins, are too hydrophilic to enter the lipophilic phase of these carriers; the incorporated therapeutic peptide is thereby protected from enzymatic degradation as well as unintended thiol/disulfide exchange reactions. Nanocarriers (<200 nm in size) with PEG or zwitterionic surfaces can permeate the mucus layer, reach the absorption mucosal membrane, and be taken up by endocytosis or transcytosis or by simply fusing with the cell membrane, followed by releasing their cargo into the systemic circulation [157]. Recently, zwitterionic (ZW), polyglycerol (PG), and polyethylene glycol (PEG) surfactant oil-in-water nano-emulsions were shown to diffuse across the mucus barrier on intestinal mucosa and also be taken up by the epithelial cells of the intestinal mucosa, as illustrated by Figure 10; in this case, the lipid-based nanocarriers with PEG displayed a lower uptake by the epithelial cells than the ZW- and PG-based carriers [158].
Many peptides self-assemble into aggregates with defined structures in a concentration-dependent manner, such as the PTP-7b (FLGALFKALSHLL) [159], gramicidin D [21], and some cyclic lipopeptides from Bacillus [160,161]. In the case of PTP-7b, the self-assembly was concentration-dependent and attributed to the interactions between the peptide and cells with cell lysis due to peptide aggregates in the cell membrane; in the other two instances, the peptides self-assembled in water independently of their interaction with cells. The cyclic lipopeptides activity against susceptible cells depended on the preservation of the cyclic moiety, which conferred resistance to proteases; the aggregates were inactive per se at the pH of the culture medium, which was around 6 or below. Knock out of the sensitive cells only occurred when the aggregates were disassembled due to a high negative charge at pH above 6 causing electrostatic repulsion between adjacent peptide molecules in the aggregate [160]. The self-assembly of gramicidin D in water and its implications on activity and reduction of in vivo proteolysis by enzymes still deserves further investigation [21].
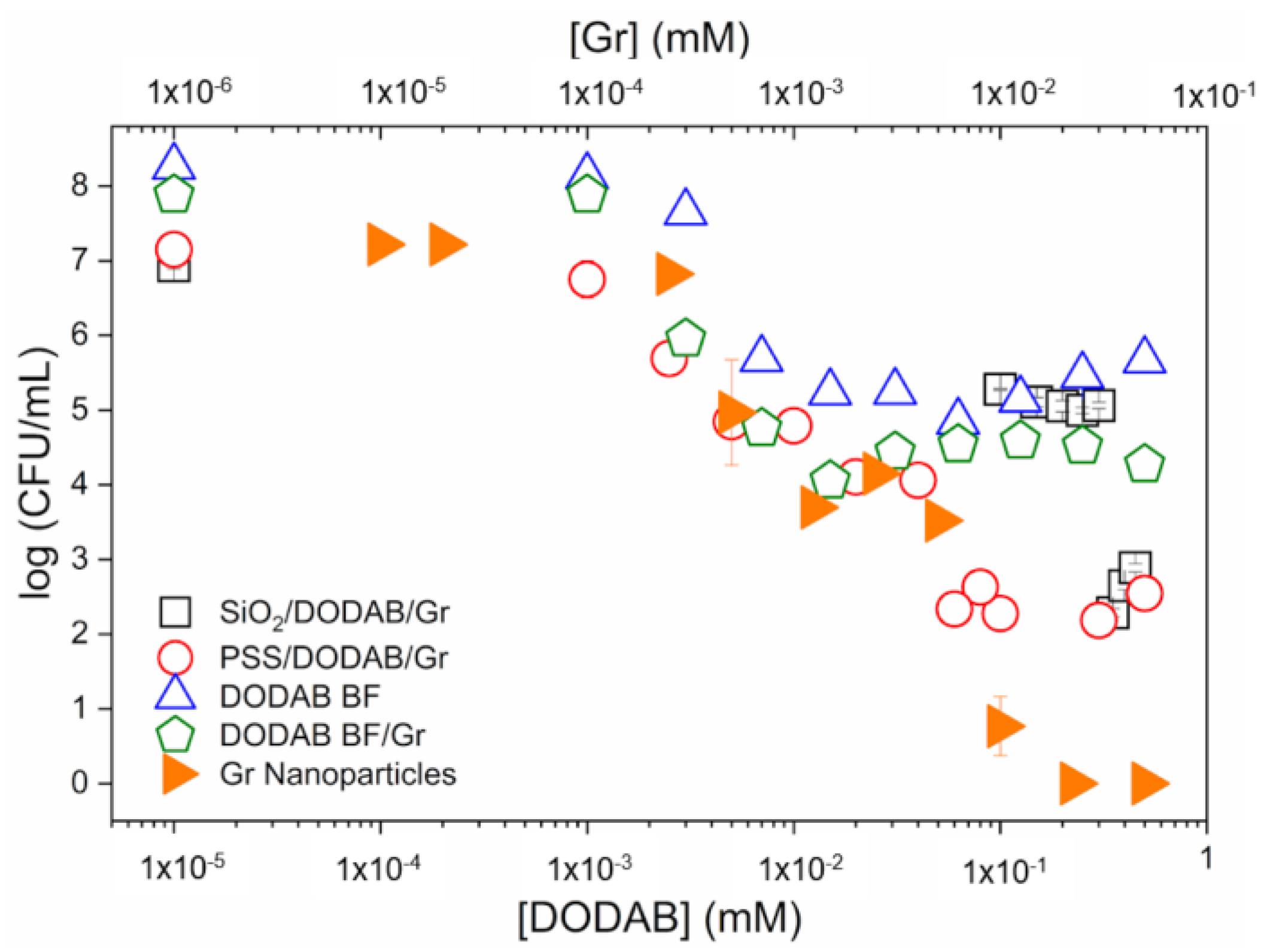
The cell viability of S. aureus was comparatively evaluated for a variety of gramicidin D formulations, including the Gr nanoparticles (Gr NPs) first reported in [21]. Figure 11, reproduced from [21], shows some Gr formulations with bilayers of the cationic antimicrobial lipid DODAB, supported or non-supported by nanoparticles. The DODAB bilayers were used to incorporate Gr and to determine antimicrobial activity against S. aureus. The cationic lipid bilayer of DODAB bears quaternary ammonium moieties well known for their activity against Gram-negative bacteria but yielding a poor performance against Gram-positive ones such as S. aureus. Gr dimeric channels found a very appropriate microenvironment in the DODAB bilayer prepared as large DODAB vesicles or bilayer fragments [135]. Furthermore, strong interaction with the DODAB bilayer hampered, to a certain extent, Gr’s interaction with the coccus [135,137]. With Gr insertion in supported DODAB bilayers on polymeric nanoparticles of polystyrene sulfate (PSS NPs) [137,162] or on silica [163], despite some improvements in the activity against S. aureus as compared to the free DODAB bilayers [164], the complete killing of the pathogen did not take place; the highest activity against S. aureus was achieved by the Gr NPs themselves, reaching a 7-log reduction in cell viability; therefore, the insertion of Gr in supported or non-supported DODAB bilayers diminished Gr activity against S. aureus as compared to the Gr NPs (Figure 11) [21]. Recent advances in self-assembled peptide nanostructures have been reviewed [143,165].
Hydrogels are a good example of versatile carriers that can deliver AMPs while displaying convenient properties such as photopolymerization, self-assembly, and controlled release of AMPs [27]. Polymers employed for constructing hydrogels can be synthetic, such as PEG and PLGA, or natural, such as collagen, hyaluronic acid, and gelatin, being highly biocompatible and displaying tunable properties. Hydrogels prepared from hydrophilic polymers can hold large amounts of water in their three-dimensional networks [166]. Natural polysaccharides such as hemicellulose [167] and alginate are also ideal materials for the preparation of biomimetic hydrogels because of their good biocompatibility and biodegradability [168]. In a very interesting study, severe inflammation and dysbiosis in ulcerative colitis were addressed using epigallocatechin-3-gallate (EGCG)-loaded silk fibroin-based nanoparticle functionalized with the AMP cathelicidin-BF (CBF); the construct could be internalized by Colon-26 cells and RAW 264.7 macrophages, restoring colonic epithelial barriers, relieving oxidative stress, promoting epithelium migration, diminishing pro-inflammatory factors, and upregulating anti-inflammatory ones. In addition, oral administration of the chitosan/alginate hydrogel embedding CBF-EGCG-NPs not only treated the ulcerative colitis but also increased the diversity and richness of beneficial bacteria such as Firmicutes and Lactobacillaceae [169].
In another instance, AMPs were grafted onto chitosan using thiol–norbornene photopolymerization, creating antibacterial hydrogels able to hamper biofilm formation [170]. For wound healing, a combination of reactive oxygen species–scavenger represented by cerium oxide nanoparticles and one AMP were formulated as a sprayable hydrogel prepared by chemically conjugating gelatin with dopamine motifs imparting improved affinity to wet skin surfaces; the rapid release of AMP from the hydrogel provided the desired release and contact ablation against four representative bacterial strains; sprayability, adhesiveness, antimicrobial activity, as well as ROS-scavenging and skin-remodeling ability, highlighted the potential of this hydrogel for wound management [171]. The lipopeptide battacin, with high activity against P. aeruginosa and S. aureus, was covalently linked from its N-terminal cysteine onto polyethylene glycol using a thiol-ene reaction; thereafter, a crosslinker, a diacrylate spacer, PEG, a photo-initiator, and UV irradiation promoted gelation. At 0.5 wt% peptide relative to the total polymer content, there was effective inhibition of bacterial growth and dispersion of established biofilms of P. aeruginosa and S. aureus, plus the absence of hemolysis [172,173].
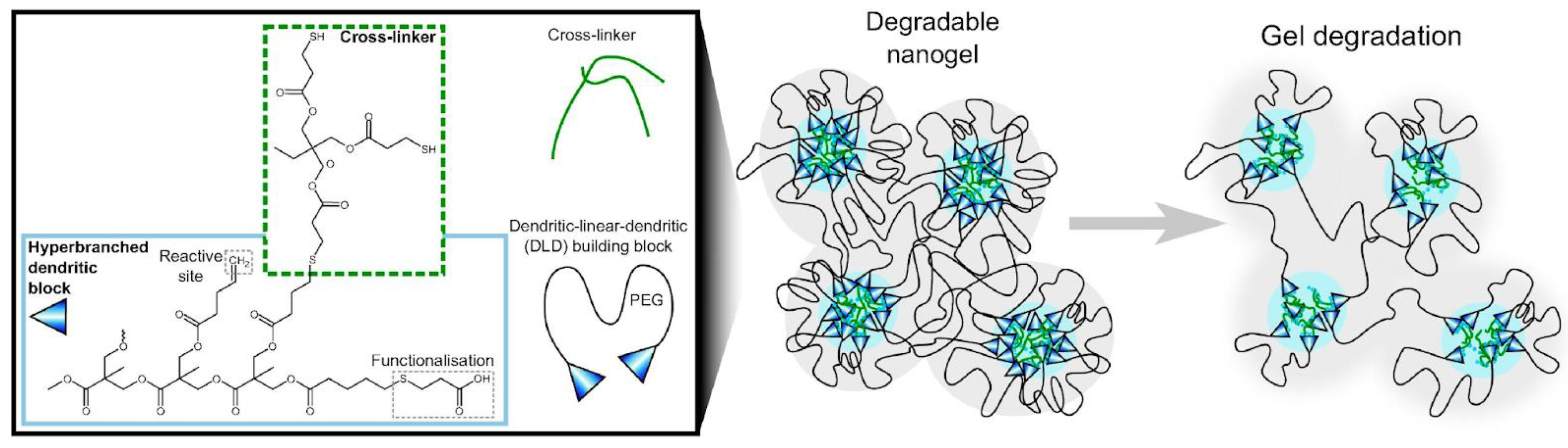
In dentistry, implants require protection against infection; several adhesive formulations containing AMPs have been developed. Streptococcus mutans is an early colonizer from dental composites used in restorations that can lead to restoration failure due to lactic acid demineralizing the adjoining tooth, subsequent growth of other bacteria, and biofilm formation; spacer-integrated AMPs conjugated to methacrylate (MA) yielding the MA–AMP monomers were next copolymerized into dental adhesives as AMP–polymer conjugates, which displayed a higher compressive modulus than hydrogels and acted against Streptococcus mutans, promoting efficient restoration. The AMP used was chosen among the most active against oral pathogenic bacteria [174,175]. Anionic poly(ethyl acrylate-co-methacrylic acid) microgels were found to incorporate considerable amounts of the cationic antimicrobial peptides LL-37, protecting them from proteolysis; however, their activity depended on LL-37 release, which was achieved by decreasing peptide length and microgel charge density [176]. Peptide loading to the microgels was driven by their opposite charges, and peptide release took place at physiological ionic strength; for peptide-loaded microgels, contact killing provided the primary antimicrobial effect at low ionic strength, whereas at high ionic strength, released peptides accounted for the antimicrobial effects [177]. Degradable anionic dendritic nanogels bearing anionic carboxylic acid binding sites for cationic AMPs, surrounded by PEG chains, were able to load cationic AMPs such as LL-37, but peptide release was much faster than nanogel degradation, and bacterial membrane destabilization was largely independent of the nanogel degradation. Figure 12 illustrates the degradable nanogels and their design, consisting of polyester chains functionalized with crosslinker and carboxyl moieties [178].
Electrospinning of polymers combined with AMPs is a key technique for next-generation polymer-based wound dressings, devices for tissue regeneration, and coatings for prostheses [35,36,179]. Examples of polymers that can form fibers by electrospinning are silk fibroin, gelatin, chitosan, PLGA, PEO, poly (acrylic acid), and others. For the treatment of osteomyelitis caused by several pathogens, fluconazole, vancomycin, and ceftazidime were incorporated into biodegradable poly lactic-co-glycolic acid (PLGA) nanofibers sustainably releasing high levels of the three agents for 30 and 56 days in vitro and in vivo, respectively [180]. Delivery systems for AMPs were recently reviewed [177,181,182]. The special role and therapeutic potential of AMPs for the treatment of infected wounds were also very recently updated [28].
In recent years, an exponential growth in research focusing on antimicrobial peptides and their assemblies can be observed in the literature (AMPs) [183,184,185,186,187,188,189,190,191,192,193,194,195,196,197,198,199,200,201,202,203,204,205,206,207,208,209,210,211,212,213,214,215,216,217,218,219,220,221,222,223,224,225,226,227,228,229,230,231,232,233,234]. Natural AMPs are important immunological defenses against microorganisms [48,49,54,55,56,103,183,218,234]. They have been extracted from several organisms, such as bacteria, plants, insects, amphibians, and humans. AMP activity is generally associated with their interaction with microbial cell membranes or with the inhibition of microbe cell wall biosynthesis [17,18,21,42,87,88]. AMPs as antibiotics exhibit some advantages: they are relatively small peptides and easy to synthesize, and they display a broad spectrum of antimicrobial activity and low susceptibility to the development of drug resistance. However, AMPs as antibiotics for clinical use are still associated with a few obstacles, including susceptibility towards proteolytic degradation, moderate activity, toxicity against eukaryotic cells, and potential immunogenicity. Consequently, only a few peptides (vancomycin, teicoplanin gramicidins, and daptomycin) have been approved as antimicrobials. Before their therapeutic use, their quoted disadvantages have to be eliminated. In this work, the methods of modifying or formulating AMPs described so far were discussed, focusing on structure–activity relationships for AMPs and their assemblies, considering not only their self-assembly but also their interactions with their carriers for optimal delivery or their combinations with other complementary antimicrobials or moieties covalently bound to their chemical structure. The effervescent research that occurs nowadays dealing with AMPs and their assemblies gives us hope for obtaining compounds with strong antimicrobial properties and controllable drawbacks.
5. Conclusions
This overview of the recent present for AMPs allows some insight into their future; their limitations regarding toxicity to the patient, biodegradable character, rapid clearance, and poor targeting to their site of action have been circumvented by multidisciplinary approaches intimately related to formulation design. In this vast field, there are several players, and strategic design involves deep knowledge of the AMP structure–function relationship and AMP’s interaction with its carrier. The interaction cannot be too strong; otherwise, the AMP will not leave the carrier to reach its microbial target. The interaction should be preferentially weak so that AMP–carrier disassembly allows AMP interaction with the microbe; this is more so when the carrier is not just a carrier and can also actively contribute to the microbicidal activity, preferentially in a complementary way, sometimes triggering a most desirable synergistic action and lowering the effective doses required for activity.
Funding
This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico, grant number 302758/2019-4.
Conflicts of Interest
The author declares no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bechinger, B. Rationalizing the Membrane Interactions of Cationic Amphipathic Antimicrobial Peptides by Their Molecular Shape. Curr. Opin. Colloid Interface Sci. 2009, 14, 349–355. [Google Scholar] [CrossRef]
- Bechinger, B.; Lohner, K. Detergent-like Actions of Linear Amphipathic Cationic Antimicrobial Peptides. Biochim. Biophys. Acta (BBA)—Biomembr. 2006, 1758, 1529–1539. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, M. Antimicrobial Peptides: From Design to Clinical Application. Antibiotics 2022, 11, 349. [Google Scholar] [CrossRef]
- Talapko, J.; Meštrović, T.; Juzbašić, M.; Tomas, M.; Erić, S.; Horvat Aleksijević, L.; Bekić, S.; Schwarz, D.; Matić, S.; Neuberg, M.; et al. Antimicrobial Peptides—Mechanisms of Action, Antimicrobial Effects and Clinical Applications. Antibiotics 2022, 11, 1417. [Google Scholar] [CrossRef]
- Browne, K.; Chakraborty, S.; Chen, R.; Willcox, M.D.; Black, D.S.; Walsh, W.R.; Kumar, N. A New Era of Antibiotics: The Clinical Potential of Antimicrobial Peptides. Int. J. Mol. Sci. 2020, 21, 7047. [Google Scholar] [CrossRef] [PubMed]
- Pavithrra, G.; Rajasekaran, R. Gramicidin Peptide to Combat Antibiotic Resistance: A Review. Int. J. Pept. Res. Ther. 2020, 26, 191–199. [Google Scholar] [CrossRef]
- Avedissian, S.N.; Liu, J.; Rhodes, N.J.; Lee, A.; Pais, G.M.; Hauser, A.R.; Scheetz, M.H. A Review of the Clinical Pharmacokinetics of Polymyxin B. Antibiotics 2019, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Field, D.; Fernandez de Ullivarri, M.; Ross, R.P.; Hill, C. After a Century of Nisin Research—Where Are We Now? FEMS Microbiol. Rev. 2023, 47, fuad023. [Google Scholar] [CrossRef]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical Applications of Nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef]
- Vilhena, C.; Bettencourt, A. Daptomycin: A Review of Properties, Clinical Use, Drug Delivery and Resistance. Mini-Rev. Med. Chem. 2012, 12, 202–209. [Google Scholar] [CrossRef]
- Sharma, C.K.; Sharma, M. Up Scaling Strategies to Improve the Industrial Production of Bacitracin at Largescale. Mini-Rev. Med. Chem. 2017, 17, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q.; Huang, S.; Jin, Y.; Campagne, R.; Alezra, V.; Wan, Y. Recent Advances in the Exploration of Therapeutic Analogues of Gramicidin S, an Old but Still Potent Antimicrobial Peptide. J. Med. Chem. 2019, 62, 7603–7617. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, N.; Mishra, B.; Felix, L.; Mylonakis, E. Antimicrobial Peptides and Small Molecules Targeting the Cell Membrane of Staphylococcus Aureus. Microbiol. Mol. Biol. Rev. 2023, 87, e00037222023. [Google Scholar] [CrossRef] [PubMed]
- Enayathullah, M.G.; Parekh, Y.; Banu, S.; Ram, S.; Nagaraj, R.; Kumar, B.K.; Idris, M.M. Gramicidin S and Melittin: Potential Anti-Viral Therapeutic Peptides to Treat SARS-CoV-2 Infection. Sci. Rep. 2022, 12, 3446. [Google Scholar] [CrossRef]
- Mousavi Maleki, M.S.; Sardari, S.; Ghandehari Alavijeh, A.; Madanchi, H. Recent Patents and FDA-Approved Drugs Based on Antiviral Peptides and Other Peptide-Related Antivirals. Int. J. Pept. Res. Ther. 2023, 29, 5. [Google Scholar] [CrossRef]
- Elnagdy, S.; AlKhazindar, M. The Potential of Antimicrobial Peptides as an Antiviral Therapy against COVID-19. ACS Pharmacol. Transl. Sci. 2020, 3, 780–782. [Google Scholar] [CrossRef]
- Malekkhaiat Häffner, S.; Malmsten, M. Influence of Self-Assembly on the Performance of Antimicrobial Peptides. Curr. Opin. Colloid Interface Sci. 2018, 38, 56–79. [Google Scholar] [CrossRef]
- Juhl, D.W.; Glattard, E.; Aisenbrey, C.; Bechinger, B. Antimicrobial Peptides: Mechanism of Action and Lipid-Mediated Synergistic Interactions within Membranes. Faraday Discuss. 2021, 232, 419–434. [Google Scholar] [CrossRef]
- Bechinger, B.; Salnikov, E.S. The Membrane Interactions of Antimicrobial Peptides Revealed by Solid-State NMR Spectroscopy. Chem. Phys. Lipids 2012, 165, 282–301. [Google Scholar] [CrossRef]
- Lombardi, L.; Shi, Y.; Falanga, A.; Galdiero, E.; de Alteriis, E.; Franci, G.; Chourpa, I.; Azevedo, H.S.; Galdiero, S. Enhancing the Potency of Antimicrobial Peptides through Molecular Engineering and Self-Assembly. Biomacromolecules 2019, 20, 1362–1374. [Google Scholar] [CrossRef]
- Pérez-Betancourt, Y.; Zaia, R.; Evangelista, M.F.; Ribeiro, R.T.; Roncoleta, B.M.; Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer. Pharmaceutics 2022, 14, 2053. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.; Zhu, G.; Song, M.; Zhao, X.; Ma, G.; Zhang, M. Study on the Self-Assembly of Aromatic Antimicrobial Peptides Based on Different PAF26 Peptide Sequences. e-Polymers 2022, 22, 276–284. [Google Scholar] [CrossRef]
- Hu, X.; Liao, M.; Gong, H.; Zhang, L.; Cox, H.; Waigh, T.A.; Lu, J.R. Recent Advances in Short Peptide Self-Assembly: From Rational Design to Novel Applications. Curr. Opin. Colloid Interface Sci. 2020, 45, 1–13. [Google Scholar] [CrossRef]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The Antimicrobial Peptides and Their Potential Clinical Applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar]
- Jiang, Y.; Chen, Y.; Song, Z.; Tan, Z.; Cheng, J. Recent Advances in Design of Antimicrobial Peptides and Polypeptides toward Clinical Translation. Adv. Drug Deliv. Rev. 2021, 170, 261–280. [Google Scholar] [CrossRef]
- Sobczak, M.; Dębek, C.; Olędzka, E.; Kozłowski, R. Polymeric Systems of Antimicrobial Peptides—Strategies and Potential Applications. Molecules 2013, 18, 14122–14137. [Google Scholar] [CrossRef]
- Copling, A.; Akantibila, M.; Kumaresan, R.; Fleischer, G.; Cortes, D.; Tripathi, R.S.; Carabetta, V.J.; Vega, S.L. Recent Advances in Antimicrobial Peptide Hydrogels. Int. J. Mol. Sci. 2023, 24, 7563. [Google Scholar] [CrossRef]
- Haidari, H.; Melguizo-Rodríguez, L.; Cowin, A.J.; Kopecki, Z. Therapeutic Potential of Antimicrobial Peptides for Treatment of Wound Infection. Am. J. Physiol. Cell Physiol. 2023, 324, C29–C38. [Google Scholar] [CrossRef]
- Costa, B.; Martínez-de-Tejada, G.; Gomes, P.A.C.; Martins, M.C.L.; Costa, F. Antimicrobial Peptides in the Battle against Orthopedic Implant-Related Infections: A Review. Pharmaceutics 2021, 13, 1918. [Google Scholar] [CrossRef]
- Luo, Y.; Song, Y. Mechanism of Antimicrobial Peptides: Antimicrobial, Anti-Inflammatory and Antibiofilm Activities. Int. J. Mol. Sci. 2021, 22, 11401. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Alford, M.A.; Haney, E.F. Antibiofilm Activity of Host Defence Peptides: Complexity Provides Opportunities. Nat. Rev. Microbiol. 2021, 19, 786–797. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Huang, J.; Zhang, W.; Zheng, X.; Wang, H.; Liu, J.; Leng, H.; Yuan, W.; Song, C. Simvastatin-Hydroxyapatite Coatings Prevent Biofilm Formation and Improve Bone Formation in Implant-Associated Infections. Bioact. Mater. 2023, 21, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Ma, L.; Sun, X.; Sloan, A.J.; O’Brien-Simpson, N.M.; Li, W. The Overview of Antimicrobial Peptide-Coated Implants against Oral Bacterial Infections. Aggregate 2023, 4, e309. [Google Scholar] [CrossRef]
- Sun, H.; Hong, Y.; Xi, Y.; Zou, Y.; Gao, J.; Du, J. Synthesis, Self-Assembly, and Biomedical Applications of Antimicrobial Peptide-Polymer Conjugates. Biomacromolecules 2018, 19, 1701–1720. [Google Scholar] [CrossRef]
- Patrulea, V.; Borchard, G.; Jordan, O. An Update on Antimicrobial Peptides (AMPs) and Their Delivery Strategies for Wound Infections. Pharmaceutics 2020, 12, 840. [Google Scholar] [CrossRef] [PubMed]
- Riool, M.; Patrulea, V.; Monteiro, C. Antimicrobial Peptide–Polymer Conjugates. Pharmaceutics 2022, 14, 2171. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; De Melo Carrasco, L.D. Novel Formulations for Antimicrobial Peptides. Int. J. Mol. Sci. 2014, 15, 18040–18083. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Self-Assembled Antimicrobial Nanomaterials. Int. J. Environ. Res. Public Health 2018, 15, 1408. [Google Scholar] [CrossRef]
- Leite, M.L.; da Cunha, N.B.; Costa, F.F. Antimicrobial Peptides, Nanotechnology, and Natural Metabolites as Novel Approaches for Cancer Treatment. Pharmacol. Ther. 2018, 183, 160–176. [Google Scholar] [CrossRef]
- Carratalá, J.V.; Serna, N.; Villaverde, A.; Vázquez, E.; Ferrer-Miralles, N. Nanostructured Antimicrobial Peptides: The Last Push towards Clinics. Biotechnol. Adv. 2020, 44, 107603. [Google Scholar] [CrossRef]
- Radaic, A.; de Jesus, M.B.; Kapila, Y.L. Bacterial Anti-Microbial Peptides and Nano-Sized Drug Delivery Systems: The State of the Art toward Improved Bacteriocins. J. Control. Release 2020, 321, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Zaia, R.; Quinto, G.M.; Camargo, L.C.S.; Ribeiro, R.T.; Carmona-Ribeiro, A.M. Transient Coatings from Nanoparticles Achieving Broad-Spectrum and High Antimicrobial Performance. Pharmaceuticals 2023, 16, 816. [Google Scholar] [CrossRef] [PubMed]
- Melo, L.D.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Antimicrobial Particles from Cationic Lipid and Polyelectrolytes. Langmuir 2010, 26, 12300–12306. [Google Scholar] [CrossRef]
- de Melo Carrasco, L.D.; Sampaio, J.L.M.; Carmona-Ribeiro, A.M. Supramolecular Cationic Assemblies against Multidrug-Resistant Microorganisms: Activity and Mechanism of Action. Int. J. Mol. Sci. 2015, 16, 6337–6352. [Google Scholar] [CrossRef] [PubMed]
- Vieira, D.B.; Carmona-Ribeiro, A.M. Cationic Nanoparticles for Delivery of Amphotericin B: Preparation, Characterization and Activity in Vitro. J. Nanobiotechnology 2008, 6, 6. [Google Scholar] [CrossRef]
- Carrasco, L.D.d.M.; Bertolucci, R.J.; Ribeiro, R.T.; Sampaio, J.L.M.; Carmona-Ribeiro, A.M. Cationic Nanostructures against Foodborne Pathogens. Front. Microbiol. 2016, 7, 1804. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; Araújo, P.M. Antimicrobial Polymer−Based Assemblies: A Review. Int. J. Mol. Sci. 2021, 22, 5424. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial Host Defence Peptides: Functions and Clinical Potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Iannella, H.; Luna, C.; Waterer, G. Inhaled Corticosteroids and the Increased Risk of Pneumonia: What’s New? A 2015 Updated Review. Ther. Adv. Respir. Dis. 2016, 10, 235–255. [Google Scholar] [CrossRef]
- Mant, C.T.; Jiang, Z.; Gera, L.; Davis, T.; Nelson, K.L.; Bevers, S.; Hodges, R.S. De Novo Designed Amphipathic α-Helical Antimicrobial Peptides Incorporating Dab and Dap Residues on the Polar Face To Treat the Gram-Negative Pathogen, Acinetobacter Baumannii. J. Med. Chem. 2019, 62, 3354–3366. [Google Scholar] [CrossRef]
- Mishra, B.; Reiling, S.; Zarena, D.; Wang, G. Host Defense Antimicrobial Peptides as Antibiotics: Design and Application Strategies. Curr. Opin. Chem. Biol. 2017, 38, 87–96. [Google Scholar] [CrossRef]
- Jiang, S.; Deslouches, B.; Chen, C.; Di, M.E.; Di, Y.P. Antibacterial Properties and Efficacy of a Novel SPLUNC1-Derived Antimicrobial Peptide, A4-Short, in a Murine Model of Respiratory Infection. mBio 2019, 10, e00226-19. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.; Pothoulakis, C.; Koon, H.W. Antimicrobial Peptides and Colitis. Curr. Pharm. Des. 2013, 19, 40–47. [Google Scholar] [CrossRef]
- Piyadasa, H.; Hemshekhar, M.; Altieri, A.; Basu, S.; van der Does, A.M.; Halayko, A.J.; Hiemstra, P.S.; Mookherjee, N. Immunomodulatory Innate Defence Regulator (IDR) Peptide Alleviates Airway Inflammation and Hyper-Responsiveness. Thorax 2018, 73, 908–917. [Google Scholar] [CrossRef]
- Chow, L.N.Y.; Choi, K.-Y.G.; Piyadasa, H.; Bossert, M.; Uzonna, J.; Klonisch, T.; Mookherjee, N. Human Cathelicidin LL-37-Derived Peptide IG-19 Confers Protection in a Murine Model of Collagen-Induced Arthritis. Mol. Immunol. 2014, 57, 86–92. [Google Scholar] [CrossRef]
- Li, D.; Wang, W.; Shi, H.; Fu, Y.; Chen, X.; Chen, X.; Liu, Y.; Kan, B.; Wang, Y. Gene Therapy with Beta-Defensin 2 Induces Antitumor Immunity and Enhances Local Antitumor Effects. Hum. Gene Ther. 2014, 25, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Felício, M.R.; Silva, O.N.; Gonçalves, S.; Santos, N.C.; Franco, O.L. Peptides with Dual Antimicrobial and Anticancer Activities. Front. Chem. 2017, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Tornesello, A.L.; Borrelli, A.; Buonaguro, L.; Buonaguro, F.M.; Tornesello, M.L. Antimicrobial Peptides as Anticancer Agents: Functional Properties and Biological Activities. Molecules 2020, 25, 2850. [Google Scholar] [CrossRef] [PubMed]
- Wodlej, C.; Riedl, S.; Rinner, B.; Leber, R.; Drechsler, C.; Voelker, D.R.; Choi, J.-Y.; Lohner, K.; Zweytick, D. Interaction of Two Antitumor Peptides with Membrane Lipids—Influence of Phosphatidylserine and Cholesterol on Specificity for Melanoma Cells. PLoS ONE 2019, 14, e0211187. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Aminov, R.; Franco, O.L.; de la Fuente-Nunez, C.; Wang, J. Editorial: Community Series in Antimicrobial Peptides: Molecular Design, Structure Function Relationship and Biosynthesis Optimization. Front. Microbiol. 2023, 14, 1125426. [Google Scholar] [CrossRef]
- Yang, Z.; He, S.; Wu, H.; Yin, T.; Wang, L.; Shan, A. Nanostructured Antimicrobial Peptides: Crucial Steps of Overcoming the Bottleneck for Clinics. Front. Microbiol. 2021, 12, 710199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Y.; Yan, Z.-B.; Meng, Y.-M.; Hong, X.-Y.; Shao, G.; Ma, J.-J.; Cheng, X.-R.; Liu, J.; Kang, J.; Fu, C.-Y. Antimicrobial Peptides: Mechanism of Action, Activity and Clinical Potential. Mil. Med. Res. 2021, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.E.; Alford, M.A.; Yung, D.B.Y.; Molchanova, N.; Fortkort, J.A.; Lin, J.S.; Diamond, G.; Hancock, R.E.W.; Jenssen, H.; Pletzer, D.; et al. Self-Assembly of Antimicrobial Peptoids Impacts Their Biological Effects on ESKAPE Bacterial Pathogens. ACS Infect. Dis. 2022, 8, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Jordan, J.B.; Easton, P.L.; Hinton, J.F. Effects of Phenylalanine Substitutions in Gramicidin A on the Kinetics of Channel Formation in Vesicles and Channel Structure in SDS Micelles. Biophys. J. 2005, 88, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Molloy, E.M.; Field, D.; O’ Connor, P.M.; Cotter, P.D.; Hill, C.; Ross, R.P. Saturation Mutagenesis of Lysine 12 Leads to the Identification of Derivatives of Nisin A with Enhanced Antimicrobial Activity. PLoS ONE 2013, 8, e58530. [Google Scholar] [CrossRef]
- Rifkind, D. Prevention by Polymyxin B of Endotoxin Lethality in Mice. J. Bacteriol. 1967, 93, 1463–1464. [Google Scholar] [CrossRef]
- Shoji, H.; Ferrer, R. Potential Survival Benefit and Early Recovery from Organ Dysfunction with Polymyxin B Hemoperfusion: Perspectives from a Real-World Big Data Analysis and the Supporting Mechanisms of Action. J. Anesth. Analg. Crit. Care 2022, 2, 27. [Google Scholar] [CrossRef]
- Katagiri, D.; Ishikane, M.; Asai, Y.; Izumi, S.; Takasaki, J.; Katsuoka, H.; Kondo, I.; Ide, S.; Nakamura, K.; Nakamoto, T.; et al. Direct Hemoperfusion Using a Polymyxin B-Immobilized Polystyrene Column for COVID-19. J. Clin. Apher. 2021, 36, 313–321. [Google Scholar] [CrossRef]
- De Rosa, S.; Cutuli, S.L.; Ferrer, R.; Antonelli, M.; Ronco, C. COVID-19 EUPHAS2 Collaborative Group. Polymyxin B Hemoperfusion in Coronavirus Disease 2019 Patients with Endotoxic Shock: Case Series from EUPHAS2 Registry. Artif. Organs 2021, 45, E187–E194. [Google Scholar] [CrossRef]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025288. [Google Scholar] [CrossRef]
- Moore, R.A.; Bates, N.C.; Hancock, R.E. Interaction of Polycationic Antibiotics with Pseudomonas Aeruginosa Lipopolysaccharide and Lipid A Studied by Using Dansyl-Polymyxin. Antimicrob. Agents Chemother. 1986, 29, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Schröder, G.; Brandenburg, K.; Seydel, U. Polymyxin B Induces Transient Permeability Fluctuations in Asymmetric Planar Lipopolysaccharide/Phospholipid Bilayers. Biochemistry 1992, 31, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Storm, D.R.; Rosenthal, K.S.; Swanson, P.E. Polymyxin and Related Peptide Antibiotics. Annu. Rev. Biochem. 1977, 46, 723–763. [Google Scholar] [CrossRef] [PubMed]
- Nang, S.C.; Azad, M.A.K.; Velkov, T.; Zhou, Q.T.; Li, J. Rescuing the Last-Line Polymyxins: Achievements and Challenges. Pharmacol. Rev. 2021, 73, 679–728. [Google Scholar] [CrossRef]
- Economou, N.J.; Cocklin, S.; Loll, P.J. High-Resolution Crystal Structure Reveals Molecular Details of Target Recognition by Bacitracin. Proc. Natl. Acad. Sci. USA 2013, 110, 14207–14212. [Google Scholar] [CrossRef]
- Storm, D.R. Mechanism of Bacitracin Action: A Specific Lipid-Peptide Interaction. Ann. N. Y. Acad. Sci. 1974, 235, 387–398. [Google Scholar] [CrossRef]
- Toscano, W.A.; Storm, D.R. Bacitracin. Pharmocol. Ther. 1982, 16, 199–210. [Google Scholar] [CrossRef]
- Stone, K.J.; Strominger, J.L. Mechanism of Action of Bacitracin: Complexation with Metal Ion and C55-Isoprenyl Pyrophosphate. Proc. Natl. Acad. Sci. USA 1971, 68, 3223–3227. [Google Scholar] [CrossRef]
- Huang, H.W. DAPTOMYCIN, Its Membrane-Active Mechanism vs. That of Other Antimicrobial Peptides. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183395. [Google Scholar] [CrossRef]
- Reid, D.J.; Dash, T.; Wang, Z.; Aspinwall, C.A.; Marty, M.T. Investigating Daptomycin-Membrane Interactions Using Native MS and Fast Photochemical Oxidation of Peptides in Nanodiscs. Anal. Chem. 2023, 95, 4984–4991. [Google Scholar] [CrossRef]
- Acharya, Y.; Dhanda, G.; Sarkar, P.; Haldar, J. Pursuit of Next-Generation Glycopeptides: A Journey with Vancomycin. Chem. Commun. 2022, 58, 1881–1897. [Google Scholar] [CrossRef] [PubMed]
- Hassan, D.; Omolo, C.A.; Gannimani, R.; Waddad, A.Y.; Mocktar, C.; Rambharose, S.; Agrawal, N.; Govender, T. Delivery of Novel Vancomycin Nanoplexes for Combating Methicillin Resistant Staphylococcus Aureus (MRSA) Infections. Int. J. Pharm. 2019, 558, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Jang, C.H.; Cho, Y.B.; Jang, Y.S.; Kim, M.S.; Kim, G.H. Antibacterial Effect of Electrospun Polycaprolactone/Polyethylene Oxide/Vancomycin Nanofiber Mat for Prevention of Periprosthetic Infection and Biofilm Formation. Int. J. Pediatr. Otorhinolaryngol. 2015, 79, 1299–1305. [Google Scholar] [CrossRef]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus Aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Acaroğlu Degitz, İ.; Hakkı Gazioğlu, B.; Burak Aksu, M.; Malta, S.; Demir Sezer, A.; Eren, T. Antibacterial and Hemolytic Activity of Cationic Polymer-Vancomycin Conjugates. Eur. Polym. J. 2020, 141, 110084. [Google Scholar] [CrossRef]
- Courvalin, P. Vancomycin Resistance in Gram-Positive Cocci. Clin. Infect. Dis. 2006, 42 (Suppl. S1), S25–S34. [Google Scholar] [CrossRef]
- Hammes, W.P.; Neuhaus, F.C. On the Mechanism of Action of Vancomycin: Inhibition of Peptidoglycan Synthesis in Gaffkya Homari. Antimicrob. Agents Chemother. 1974, 6, 722–728. [Google Scholar] [CrossRef]
- Reynolds, P.E. Structure, Biochemistry and Mechanism of Action of Glycopeptide Antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef]
- Hallett, J.W.; Wolkowicz, M.I.; Leopold, I.H. Ophthalmic Use of Neosporin. Am. J. Ophthalmol. 1956, 41, 850–853. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; de Melo Carrasco, L.D. Cationic Antimicrobial Polymers and Their Assemblies. Int. J. Mol. Sci. 2013, 14, 9906–9946. [Google Scholar] [CrossRef]
- Sanches, L.M.; Petri, D.F.S.; de Melo Carrasco, L.D.; Carmona-Ribeiro, A.M. The Antimicrobial Activity of Free and Immobilized Poly (Diallyldimethylammonium) Chloride in Nanoparticles of Poly (Methylmethacrylate). J. Nanobiotechnol. 2015, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Galvão, C.N.; Sanches, L.M.; Mathiazzi, B.I.; Ribeiro, R.T.; Petri, D.F.S.; Carmona-Ribeiro, A.M. Antimicrobial Coatings from Hybrid Nanoparticles of Biocompatible and Antimicrobial Polymers. Int. J. Mol. Sci. 2018, 19, 2965. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.T.; Galvão, C.N.; Betancourt, Y.P.; Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Microbicidal Dispersions and Coatings from Hybrid Nanoparticles of Poly (Methyl Methacrylate), Poly (Diallyl Dimethyl Ammonium) Chloride, Lipids, and Surfactants. Int. J. Mol. Sci. 2019, 20, 6150. [Google Scholar] [CrossRef]
- de Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Vessoni Penna, T.C. Nisin Biotechnological Production and Application: A Review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Jalalifar, S.; Mirzaei, R.; Motallebirad, T.; Razavi, S.; Talebi, M. The Emerging Role of Probiotics and Their Derivatives against Biofilm-Producing MRSA: A Scoping Review. Biomed. Res. Int. 2022, 2022, 4959487. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.G.; Becattini, S.; Moody, T.U.; Shliaha, P.V.; Littmann, E.R.; Seok, R.; Gjonbalaj, M.; Eaton, V.; Fontana, E.; Amoretti, L.; et al. Microbiota-Derived Lantibiotic Restores Resistance against Vancomycin-Resistant Enterococcus. Nature 2019, 572, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, S.A.; Mahmoud Ahmed, S.; Elmenshawy, A.M.; Sultan, G.M.; Asser, S.L. Study of the Gut Microbiome as a Novel Target for Prevention of Hospital-Associated Infections in Intensive Care Unit Patients. Acute Crit. Care 2023, 38, 76–85. [Google Scholar] [CrossRef]
- Petrof, E.O.; Dhaliwal, R.; Manzanares, W.; Johnstone, J.; Cook, D.; Heyland, D.K. Probiotics in the Critically Ill: A Systematic Review of the Randomized Trial Evidence. Crit. Care Med. 2012, 40, 3290–3302. [Google Scholar] [CrossRef]
- Chernyshova, D.N.; Tyulin, A.A.; Ostroumova, O.S.; Efimova, S.S. Discovery of the Potentiator of the Pore-Forming Ability of Lantibiotic Nisin: Perspectives for Anticancer Therapy. Membranes 2022, 12, 1166. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B.K.H.L. Review: Lessons Learned From Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef]
- Nayab, S.; Aslam, M.A.; Rahman, S.U.; Sindhu, Z.U.D.; Sajid, S.; Zafar, N.; Razaq, M.; Kanwar, R.; Amanullah. A Review of Antimicrobial Peptides: Its Function, Mode of Action and Therapeutic Potential. Int. J. Pept. Res. Ther. 2022, 28, 46. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Bharath Prasad, A.S.; Mehta, C.H.; Nayak, U.Y. Antimicrobial Peptide Polymers: No Escape to ESKAPE Pathogens—A Review. World J. Microbiol. Biotechnol. 2020, 36, 131. [Google Scholar] [CrossRef]
- Ulm, H.; Wilmes, M.; Shai, Y.; Sahl, H.-G. Antimicrobial Host Defensins—Specific Antibiotic Activities and Innate Defense Modulation. Front. Immunol. 2012, 3, 249. [Google Scholar] [CrossRef] [PubMed]
- Kelkar, D.A.; Chattopadhyay, A. The Gramicidin Ion Channel: A Model Membrane Protein. Biochim. Biophys. Acta 2007, 1768, 2011–2025. [Google Scholar] [CrossRef] [PubMed]
- Koeppe, R.E.; Anderson, O.S. Engineering the Gramicidin Channel. Annu. Rev. Biophys. Biomol. Struct. 1996, 25, 231–258. [Google Scholar] [CrossRef]
- Sarges, R.; Witkop, B. Gramicidin A. V. The Structure of Valine- and Isoleucine-gramicidin A. J. Am. Chem. Soc. 1965, 87, 2011–2020. [Google Scholar] [CrossRef]
- Urban, B.W.; Hladky, S.B.; Haydon, D.A. Ion Movements in Gramicidin Pores. An Example of Single-File Transport. Biochim. Biophys. Acta (BBA)—Biomembr. 1980, 602, 331–354. [Google Scholar] [CrossRef]
- Mukherjee, S.; Chattopadhyay, A. Motionally Restricted Tryptophan Environments at the Peptide-Lipid Interface of Gramicidin Channels. Biochemistry 1994, 33, 5089–5097. [Google Scholar] [CrossRef]
- David, J.M.; Rajasekaran, A.K. Gramicidin A: A New Mission for an Old Antibiotic. J. Kidney Cancer VHL 2015, 2, 15–24. [Google Scholar] [CrossRef]
- Haoyang, W.-W.; Xiao, Q.; Ye, Z.; Fu, Y.; Zhang, D.-W.; Li, J.; Xiao, L.; Li, Z.-T.; Hou, J.-L. Gramicidin A-Based Unimolecular Channel: Cancer Cell-Targeting Behavior and Ion Transport-Induced Apoptosis. Chem. Commun. 2021, 57, 1097–1100. [Google Scholar] [CrossRef]
- David, J.M.; Owens, T.A.; Barwe, S.P.; Rajasekaran, A.K. Gramicidin A Induces Metabolic Dysfunction and Energy Depletion Leading to Cell Death in Renal Cell Carcinoma Cells. Mol. Cancer Ther. 2013, 12, 2296–2307. [Google Scholar] [CrossRef] [PubMed]
- Raileanu, M.; Popescu, A.; Bacalum, M. Antimicrobial Peptides as New Combination Agents in Cancer Therapeutics: A Promising Protocol against HT-29 Tumoral Spheroids. Int. J. Mol. Sci. 2020, 21, 6964. [Google Scholar] [CrossRef]
- Xue, Y.-W.; Itoh, H.; Dan, S.; Inoue, M. Gramicidin A Accumulates in Mitochondria, Reduces ATP Levels, Induces Mitophagy, and Inhibits Cancer Cell Growth. Chem. Sci. 2022, 13, 7482–7491. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, Y.; Yang, Y.; Yu, K.; Cao, X.; Su, F.; Xu, H.; Peng, Y.; Hu, Y.; Qian, F.; et al. Gramicidin Inhibits Human Gastric Cancer Cell Proliferation, Cell Cycle and Induced Apoptosis. Biol. Res. 2019, 52, 57. [Google Scholar] [CrossRef] [PubMed]
- Bubin, R.; Uljanovs, R.; Strumfa, I. Cancer Stem Cells in Pancreatic Ductal Adenocarcinoma. Int. J. Mol. Sci. 2023, 24, 7030. [Google Scholar] [CrossRef]
- Wang, R.-Q.; Geng, J.; Sheng, W.-J.; Liu, X.-J.; Jiang, M.; Zhen, Y.-S. The Ionophore Antibiotic Gramicidin A Inhibits Pancreatic Cancer Stem Cells Associated with CD47 Down-Regulation. Cancer Cell Int. 2019, 19, 145. [Google Scholar] [CrossRef]
- Maham, S.; Awan, S.N.; Adnan, F.; Kakar, S.J.; Mian, A.; Khan, D. PB1837: Antibiotics; a Possible Alternate Treatment Option for Myeloid Leukemia. Hemasphere 2023, 7 (Suppl. S3), e1313111. [Google Scholar] [CrossRef]
- Yang, X.; Hua, C.; Lin, L.; Ganting, Z. Antimicrobial Peptides as Potential Therapy for Gastrointestinal Cancers. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 396, 2831–2841. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ruan, P.; Jiang, G.; Zhang, W. Screening and in Vitro Biological Evaluation of Novel Multiple Tyrosine Kinases Inhibitors as Promising Anticancer Agents. Anticancer Agents Med. Chem. 2023. [Google Scholar] [CrossRef]
- Gong, X.; Zou, L.; Wang, M.; Zhang, Y.; Peng, S.; Zhong, M.; Zhou, J.; Li, X.; Ma, X. Gramicidin Inhibits Cholangiocarcinoma Cell Growth by Suppressing EGR4. Artif. Cells Nanomed. Biotechnol. 2020, 48, 53–59. [Google Scholar] [CrossRef]
- Yang, M.; Liu, S.; Zhang, C. Antimicrobial Peptides with Antiviral and Anticancer Properties and Their Modification and Nanodelivery Systems. Curr. Res. Biotechnol. 2023, 5, 100121. [Google Scholar] [CrossRef]
- Carvalho, C.A.; Olivares-Ortega, C.; Soto-Arriaza, M.A.; Carmona-Ribeiro, A.M. Interaction of Gramicidin with DPPC/DODAB Bilayer Fragments. Biochim. Biophys. Acta 2012, 1818, 3064–3071. [Google Scholar] [CrossRef] [PubMed]
- Carmona Ribeiro, A.M.; Chaimovich, H. Preparation and Characterization of Large Dioctadecyldimethylammonium Chloride Liposomes and Comparison with Small Sonicated Vesicles. Biochim. Biophys. Acta (BBA)—Biomembr. 1983, 733, 172–179. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Bilayer-Forming Synthetic Lipids: Drugs or Carriers? Curr. Med. Chem. 2003, 10, 2425–2446. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Castuma, C.E.; Sesso, A.; Schreier, S. Bilayer Structure and Stability in Dihexadecyl Phosphate Dispersions. J. Phys. Chem. 1991, 95, 5361–5366. [Google Scholar] [CrossRef]
- Andersson, M.; Hammarstroem, L.; Edwards, K. Effect of Bilayer Phase Transitions on Vesicle Structure, and Its Influence on the Kinetics of Viologen Reduction. J. Phys. Chem. 1995, 99, 14531–14538. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Lipid Bilayer Fragments and Disks in Drug Delivery. Curr. Med. Chem. 2006, 13, 1359–1370. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Synthetic Amphiphile Vesicles. Chem. Soc. Rev. 1992, 21, 209–214. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; Midmore, B.R. Surface Potential in Charged Synthetic Amphiphile Vesicles. J. Phys. Chem. 1992, 96, 3542–3547. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Bilayer Vesicles and Liposomes as Interface Agents. Chem. Soc. Rev. 2001, 30, 241–247. [Google Scholar] [CrossRef]
- Campanhã, M.T.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Interactions between Cationic Liposomes and Bacteria: The Physical-Chemistry of the Bactericidal Action. J. Lipid Res. 1999, 40, 1495–1500. [Google Scholar] [CrossRef] [PubMed]
- Martins, L.M.S.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Cationic Vesicles as Bactericides. Langmuir 1997, 13, 5583–5587. [Google Scholar] [CrossRef]
- Sicchierolli, S.M.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Bacteria Flocculation and Death by Cationic Vesicles. Langmuir 1995, 11, 2991–2995. [Google Scholar] [CrossRef]
- Campanhã, M.T.N.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Interactions between Cationic Vesicles and Candida Albicans. J. Phys. Chem. B 2001, 105, 8230–8236. [Google Scholar] [CrossRef]
- Ragioto, D.A.; Carrasco, L.D.; Carmona-Ribeiro, A.M. Novel Gramicidin Formulations in Cationic Lipid as Broad-Spectrum Microbicidal Agents. Int. J. Nanomed. 2014, 9, 3183–3192. [Google Scholar] [CrossRef]
- Carmonaribeiro, A.; Carrasco, L. Fungicidal Assemblies and Their Mode of Action. OA Biotechnol. 2013, 2, 25. [Google Scholar] [CrossRef]
- Xavier, G.R.S.; Carmona-Ribeiro, A.M. Cationic Biomimetic Particles of Polystyrene/Cationic Bilayer/Gramicidin for Optimal Bactericidal Activity. Nanomaterials 2017, 7, 422. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Systems in Nanomedicine. In Handbook of Nanobiomedical Research: Fundamentals, Applications and Recent Developments; World Scientific: Singapore, 2014; pp. 401–456. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Nanoparticles: Preparation, Characterization and Biomedical Applications. Int. J. Nanomed. 2010, 5, 249–259. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Lipid Polymer Nanoparticles for Drug Delivery. In Nanoparticles in Biology and Medicine: Methods and Protocols; Ferrari, E., Soloviev, M., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2020; pp. 45–60. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Lipid-Based Biomimetics in Drug and Vaccine Delivery. In Biomimetics Learning from Nature; IntechOpen: London, UK, 2010. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Nanomaterials from the Assembly of Polymers, Lipids, and Surfactants. In Surfactants and Detergents; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Zou, P.; Chen, W.-T.; Sun, T.; Gao, Y.; Li, L.-L.; Wang, H. Recent Advances: Peptides and Self-Assembled Peptide-Nanosystems for Antimicrobial Therapy and Diagnosis. Biomater. Sci. 2020, 8, 4975–4996. [Google Scholar] [CrossRef]
- Sahandi Zangabad, P.; Abousalman Rezvani, Z.; Tong, Z.; Esser, L.; Vasani, R.B.; Voelcker, N.H. Recent Advances in Formulations for Long-Acting Delivery of Therapeutic Peptides. ACS Appl. Bio Mater. 2023, 6, 3532–3554. [Google Scholar] [CrossRef]
- Lau, J.L.; Dunn, M.K. Therapeutic Peptides: Historical Perspectives, Current Development Trends, and Future Directions. Bioorg. Med. Chem. 2018, 26, 2700–2707. [Google Scholar] [CrossRef] [PubMed]
- Barfar, A.; Javadzadeh, Y.; Alizadeh, H.; Masoomzadeh, S. Oral Insulin Delivery: A Review On Recent Advancements and Novel Strategies. Curr. Drug Deliv. 2023. [Google Scholar] [CrossRef]
- Uhl, P.; Grundmann, C.; Sauter, M.; Storck, P.; Tursch, A.; Özbek, S.; Leotta, K.; Roth, R.; Witzigmann, D.; Kulkarni, J.A.; et al. Coating of PLA-Nanoparticles with Cyclic, Arginine-Rich Cell Penetrating Peptides Enables Oral Delivery of Liraglutide. Nanomedicine 2020, 24, 102132. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Xu, W.; Wu, Y.; Niu, B.; Zhang, X. Macroporous Organosilicon Nanocomposites Co-Deliver Bcl2-Converting Peptide and Chemotherapeutic Agent for Synergistic Treatment against Multidrug Resistant Cancer. Cancer Lett. 2020, 469, 340–354. [Google Scholar] [CrossRef]
- Yu, Q.; Deng, T.; Lin, F.-C.; Zhang, B.; Zink, J.I. Supramolecular Assemblies of Heterogeneous Mesoporous Silica Nanoparticles to Co-Deliver Antimicrobial Peptides and Antibiotics for Synergistic Eradication of Pathogenic Biofilms. ACS Nano 2020, 14, 5926–5937. [Google Scholar] [CrossRef]
- Moorcroft, S.C.T.; Roach, L.; Jayne, D.G.; Ong, Z.Y.; Evans, S.D. Nanoparticle-Loaded Hydrogel for the Light-Activated Release and Photothermal Enhancement of Antimicrobial Peptides. ACS Appl. Mater. Interfaces 2020, 12, 24544–24554. [Google Scholar] [CrossRef]
- Dutta, K.; Das, R.; Ling, J.; Monibas, R.M.; Carballo-Jane, E.; Kekec, A.; Feng, D.D.; Lin, S.; Mu, J.; Saklatvala, R.; et al. In Situ Forming Injectable Thermoresponsive Hydrogels for Controlled Delivery of Biomacromolecules. ACS Omega 2020, 5, 17531–17542. [Google Scholar] [CrossRef]
- Wang, P.; Zhuo, X.; Chu, W.; Tang, X. Exenatide-Loaded Microsphere/Thermosensitive Hydrogel Long-Acting Delivery System with High Drug Bioactivity. Int. J. Pharm. 2017, 528, 62–75. [Google Scholar] [CrossRef]
- Xiong, Z.; Cui, W.; Sun, T.; Teng, Y.; Qu, Y.; Yang, L.; Zhou, J.; Chen, K.; Yao, S.; Shao, Z.; et al. Sustained Delivery of PlGF-2123-144*-Fused BMP2-Related Peptide P28 from Small Intestinal Submucosa/Polylactic Acid Scaffold Material for Bone Tissue Regeneration. RSC Adv. 2020, 10, 7289–7300. [Google Scholar] [CrossRef]
- Luo, J.; Zhang, H.; Zhu, J.; Cui, X.; Gao, J.; Wang, X.; Xiong, J. 3-D Mineralized Silk Fibroin/Polycaprolactone Composite Scaffold Modified with Polyglutamate Conjugated with BMP-2 Peptide for Bone Tissue Engineering. Colloids Surf. B Biointerfaces 2018, 163, 369–378. [Google Scholar] [CrossRef]
- Tran, H.; ElSayed, M.E.H. Progress and Limitations of Oral Peptide Delivery as a Potentially Transformative Therapy. Expert. Opin. Drug Deliv. 2022, 19, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, X.; Zhao, Y.; Zheng, Q.; Wu, Q.; Yu, Y. Nanocarrier System: An Emerging Strategy for Bioactive Peptide Delivery. Front. Nutr. 2022, 9, 1050647. [Google Scholar] [CrossRef] [PubMed]
- Haddadzadegan, S.; Dorkoosh, F.; Bernkop-Schnürch, A. Oral Delivery of Therapeutic Peptides and Proteins: Technology Landscape of Lipid-Based Nanocarriers. Adv. Drug Deliv. Rev. 2022, 182, 114097. [Google Scholar] [CrossRef] [PubMed]
- Veider, F.; Knoll, P.; Jörgensen, A.M.; Stengel, D.; Bernkop-Schnürch, A. Oral Drug Delivery: Influence of Mucus on Cellular Interactions and Uptake of Lipid-Based Nanocarriers in Caco-2 Cells. Acta Biomater. 2023, 167, 416–424. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Patrone, N.; Liang, J.F. Peptide Self-Assembly on Cell Membranes to Induce Cell Lysis. Biomacromolecules 2012, 13, 3327–3333. [Google Scholar] [CrossRef]
- Pueyo, M.T.; Mutafci, B.A.; Soto-Arriaza, M.A.; Di Mascio, P.; Carmona-Ribeiro, A.M. The Self-Assembly of a Cyclic Lipopeptides Mixture Secreted by a B. Megaterium Strain and Its Implications on Activity against a Sensitive Bacillus Species. PLoS ONE 2014, 9, e97261. [Google Scholar] [CrossRef]
- Pueyo, M.T.; Bloch, C.; Carmona-Ribeiro, A.M.; di Mascio, P. Lipopeptides Produced by a Soil Bacillus Megaterium Strain. Microb. Ecol. 2009, 57, 367–378. [Google Scholar] [CrossRef]
- Lincopan, N.; Espíndola, N.; Vaz, A.; Carmonaribeiro, A. Cationic Supported Lipid Bilayers for Antigen Presentation. Int. J. Pharm. 2007, 340, 216–222. [Google Scholar] [CrossRef]
- Ribeiro, R.T.; Braga, V.H.A.; Carmona-Ribeiro, A.M. Biomimetic Cationic Nanoparticles Based on Silica: Optimizing Bilayer Deposition from Lipid Films. Biomimetics 2017, 2, 20. [Google Scholar] [CrossRef]
- Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Hybrid Nanoparticles of Poly (Methyl Methacrylate) and Antimicrobial Quaternary Ammonium Surfactants. Pharmaceutics 2020, 12, 340. [Google Scholar] [CrossRef]
- Simonson, A.W.; Aronson, M.R.; Medina, S.H. Supramolecular Peptide Assemblies as Antimicrobial Scaffolds. Molecules 2020, 25, 2751. [Google Scholar] [CrossRef]
- Ahmed, E.M. Hydrogel: Preparation, Characterization, and Applications: A Review. J. Adv. Res. 2015, 6, 105–121. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Chen, T.; Dong, C.; Pan, X. Biomedical Applications of Hemicellulose-Based Hydrogels. Curr. Med. Chem. 2020, 27, 4647–4659. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, L.; Wang, J.; Meng, Q.; Zhong, S.; Gao, Y.; Cui, X. Recent Advances in Polysaccharide-Based Self-Healing Hydrogels for Biomedical Applications. Carbohydr. Polym. 2022, 283, 119161. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Cao, Y.; Ma, L.; Sun, J.; Ramos-Mucci, L.; Ma, Y.; Yang, X.; Zhu, Z.; Zhang, J.; Xiao, B. Oral Antimicrobial Peptide-EGCG Nanomedicines for Synergistic Treatment of Ulcerative Colitis. J. Control. Release 2022, 347, 544–560. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.M.; Pereira, R.F.; Costa, B.; Tassi, N.; Teixeira, C.; Leiro, V.; Monteiro, C.; Gomes, P.; Costa, F.; Martins, M.C.L. Thiol–Norbornene Photoclick Chemistry for Grafting Antimicrobial Peptides onto Chitosan to Create Antibacterial Biomaterials. ACS Appl. Polym. Mater. 2022, 4, 5012–5026. [Google Scholar] [CrossRef]
- Cheng, H.; Shi, Z.; Yue, K.; Huang, X.; Xu, Y.; Gao, C.; Yao, Z.; Zhang, Y.S.; Wang, J. Sprayable Hydrogel Dressing Accelerates Wound Healing with Combined Reactive Oxygen Species-Scavenging and Antibacterial Abilities. Acta Biomater. 2021, 124, 219–232. [Google Scholar] [CrossRef] [PubMed]
- De Zoysa, G.H.; Wang, K.; Lu, J.; Hemar, Y.; Sarojini, V. Covalently Immobilized Battacin Lipopeptide Gels with Activity against Bacterial Biofilms. Molecules 2020, 25, 5945. [Google Scholar] [CrossRef]
- Qian, C.-D.; Wu, X.-C.; Teng, Y.; Zhao, W.-P.; Li, O.; Fang, S.-G.; Huang, Z.-H.; Gao, H.-C. Battacin (Octapeptin B5), a New Cyclic Lipopeptide Antibiotic from Paenibacillus Tianmuensis Active against Multidrug-Resistant Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2012, 56, 1458–1465. [Google Scholar] [CrossRef]
- Xie, S.-X.; Song, L.; Yuca, E.; Boone, K.; Sarikaya, R.; VanOosten, S.K.; Misra, A.; Ye, Q.; Spencer, P.; Tamerler, C. Antimicrobial Peptide–Polymer Conjugates for Dentistry. ACS Appl. Polym. Mater. 2020, 2, 1134–1144. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, X.; Jiang, W.; Wang, K.; Luo, J.; Li, W.; Zhou, X.; Zhang, L. Antimicrobial Peptide GH12 Suppresses Cariogenic Virulence Factors of Streptococcus mutans. J. Oral Microbiol. 2018, 10, 1442089. [Google Scholar] [CrossRef] [PubMed]
- Nordström, R.; Nyström, L.; Andrén, O.C.J.; Malkoch, M.; Umerska, A.; Davoudi, M.; Schmidtchen, A.; Malmsten, M. Membrane Interactions of Microgels as Carriers of Antimicrobial Peptides. J. Colloid Interface Sci. 2018, 513, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Caselli, L.; Malmsten, M. Skin and Wound Delivery Systems for Antimicrobial Peptides. Curr. Opin. Colloid Interface Sci. 2023, 65, 101701. [Google Scholar] [CrossRef]
- Nordström, R.; Andrén, O.C.J.; Singh, S.; Malkoch, M.; Davoudi, M.; Schmidtchen, A.; Malmsten, M. Degradable Dendritic Nanogels as Carriers for Antimicrobial Peptides. J. Colloid Interface Sci. 2019, 554, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Dart, A.; Bhave, M.; Kingshott, P. Antimicrobial Peptide-Based Electrospun Fibers for Wound Healing Applications. Macromol. Biosci. 2019, 19, e1800488. [Google Scholar] [CrossRef]
- Hsu, Y.-H.; Yu, Y.-H.; Chou, Y.-C.; Lu, C.-J.; Lin, Y.-T.; Ueng, S.W.-N.; Liu, S.-J. Sustained Release of Antifungal and Antibacterial Agents from Novel Hybrid Degradable Nanofibers for the Treatment of Polymicrobial Osteomyelitis. Int. J. Mol. Sci. 2023, 24, 3254. [Google Scholar] [CrossRef]
- Li, G.; Lai, Z.; Shan, A. Advances of Antimicrobial Peptide-Based Biomaterials for the Treatment of Bacterial Infections. Adv. Sci. 2023, 10, 2206602. [Google Scholar] [CrossRef]
- Cesaro, A.; Lin, S.; Pardi, N.; de la Fuente-Nunez, C. Advanced Delivery Systems for Peptide Antibiotics. Adv. Drug Deliv. Rev. 2023, 196, 114733. [Google Scholar] [CrossRef]
- Giordano, N.P.; Cian, M.B.; Dalebroux, Z.D. Outer Membrane Lipid Secretion and the Innate Immune Response to Gram-Negative Bacteria. Infect. Immun. 2020, 88, e00920-19. [Google Scholar] [CrossRef]
- Upert, G.; Luther, A.; Obrecht, D.; Ermert, P. Emerging peptide antibiotics with therapeutic potential. Med. Drug Discov. 2021, 9, 100078. [Google Scholar] [CrossRef]
- Cardoso, P.; Glossop, H.; Meikle, T.G.; Aburto-Medina, A.; Conn, C.E.; Sarojini, V.; Valery, C. Molecular engineering of antimicrobial peptides: Microbial targets, peptide motifs and translation opportunities. Biophys. Rev. 2021, 13, 35–69. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Chatterjee, R.; Chakravortty, D. Evolving and assembling to pierce through: Evolutionary and structural aspects of antimicrobial peptides. Comput. Struct. Biotechnol. J. 2022, 20, 2247–2258. [Google Scholar] [CrossRef] [PubMed]
- Juhász, T.; Quemé-Peña, M.; Kővágó, B.; Mihály, J.; Ricci, M.; Horváti, K.; Bősze, S.; Zsila, F.; Beke-Somfai, T. Interplay between membrane active host defense peptides and heme modulates their assemblies and in vitro activity. Sci. Rep. 2021, 11, 18328. [Google Scholar] [CrossRef] [PubMed]
- Glossop, H.D.; De Zoysa, G.H.; Hemar, Y.; Cardoso, P.; Wang, K.; Lu, J.; Valéry, C.; Sarojini, V. Battacin-Inspired Ultrashort Peptides: Nanostructure Analysis and Antimicrobial Activity. Biomacromolecules 2019, 20, 2515–2529. [Google Scholar] [CrossRef]
- Claro, B.; Peón, A.; González-Freire, E.; Goormaghtigh, E.; Amorín, M.; Granja, J.R.; Garcia-Fandiño, R.; Bastos, M. Macromolecular assembly and membrane activity of antimicrobial D,L-α-Cyclic peptides. Colloids Surf. B Biointerfaces 2021, 208, 112086. [Google Scholar] [CrossRef]
- Guo, Z.; Wang, Y.; Tan, T.; Ji, Y.; Hu, J.; Zhang, Y. Antimicrobial d-Peptide Hydrogels. ACS Biomater. Sci. Eng. 2021, 7, 1703–1712. [Google Scholar] [CrossRef]
- Dahan, S.; Aibinder, P.; Khalfin, B.; Moran-Gilad, J.; Rapaport, H. Hybrid Hydrogels of FKF-Peptide Assemblies and Gelatin for Sustained Antimicrobial Activity. ACS Biomater. Sci. Eng. 2023, 9, 352–362. [Google Scholar] [CrossRef]
- Tan, A.; Xu, F.; Yokoyama, C.; Yano, S.; Konno, H. Design, synthesis, and evaluation of the self-assembled antimicrobial peptides based on the ovalbumin-derived peptide TK913. J. Pept. Sci. 2022, 28, e3375. [Google Scholar] [CrossRef]
- Baltutis, V.; O’Leary, P.D.; Martin, L.L. Self-Assembly of Linear, Natural Antimicrobial Peptides: An Evolutionary Perspective. ChemPlusChem 2022, 87, e202200240. [Google Scholar] [CrossRef]
- Matthyssen, T.; Li, W.; Holden, J.A.; Lenzo, J.C.; Hadjigol, S.; O’Brien-Simpson, N.M. The Potential of Modified and Multimeric Antimicrobial Peptide Materials as Superbug Killers. Front. Chem. 2022, 9, 795433. [Google Scholar] [CrossRef]
- Fa, K.; Liu, H.; Gong, H.; Zhang, L.; Liao, M.; Hu, X.; Ciumac, D.; Li, P.; Webster, J.; Petkov, J.; et al. In-Membrane Nanostructuring of Cationic Amphiphiles Affects Their Antimicrobial Efficacy and Cytotoxicity: A Comparison Study between a De Novo Antimicrobial Lipopeptide and Traditional Biocides. Langmuir 2022, 38, 6623–6637. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, X.; Nie, J.; Zhao, H.; Li, W. Nano-Antimicrobial Peptides Based on Constitutional Isomerism-Dictated Self-Assembly. Biomacromolecules 2022, 23, 1302–1313. [Google Scholar] [CrossRef] [PubMed]
- Hurtado-Rios, J.J.; Carrasco-Navarro, U.; Almanza-Pérez, J.C.; Ponce-Alquicira, E. Ribosomes: The New Role of Ribosomal Proteins as Natural Antimicrobials. Int. J. Mol. Sci. 2022, 23, 9123. [Google Scholar] [CrossRef] [PubMed]
- Caselli, L.; Rodrigues, G.R.; Franco, O.L.; Malmsten, M. Pulmonary delivery systems for antimicrobial peptides. Crit. Rev. Biotechnol. 2023, 1–18. [Google Scholar] [CrossRef]
- Townsend, J.A.; Marty, M.T. What’s the defect? Using mass defects to study oligomerization of membrane proteins and peptides in nanodiscs with native mass spectrometry. Methods 2023, 218, 1–13. [Google Scholar] [CrossRef]
- Lan, X.; Zhong, J.; Huang, R.; Liu, Y.; Ma, X.; Li, X.; Zhao, D.; Qing, G.; Zhang, Y.; Liu, L.; et al. Conformation Dependent Architectures of Assembled Antimicrobial Peptides with Enhanced Antimicrobial Ability. Adv. Healthc. Mater. 2023, 4, e2301688. [Google Scholar] [CrossRef]
- Ramos-Martín, F.; D’Amelio, N. Biomembrane lipids: When physics and chemistry join to shape biological activity. Biochimie 2022, 203, 118–138. [Google Scholar] [CrossRef]
- Akshaya, S.; Rowlo, P.K.; Dukle, A.; Nathanael, A.J. Antibacterial Coatings for Titanium Implants: Recent Trends and Future Perspectives. Antibiotics 2022, 11, 1719. [Google Scholar] [CrossRef]
- Yang, W.; Seo, J.; Kim, J.H. Protein-mimetic peptoid nanoarchitectures for pathogen recognition and neutralization. Nanoscale 2023, 15, 975–986. [Google Scholar] [CrossRef]
- Rosetti, B.; Scarel, E.; Colomina-Alfaro, L.; Adorinni, S.; Pierri, G.; Bellotto, O.; Mamprin, K.; Polentarutti, M.; Bandiera, A.; Tedesco, C.; et al. Self-Assembly of Homo- and Hetero-Chiral Cyclodipeptides into Supramolecular Polymers towards Antimicrobial Gels. Polymers 2022, 14, 4554. [Google Scholar] [CrossRef]
- Serrano-Aroca, Á.; Cano-Vicent, A.; Sabater I Serra, R.; El-Tanani, M.; Aljabali, A.; Tambuwala, M.M.; Mishra, Y.K. Scaffolds in the microbial resistant era: Fabrication, materials, properties and tissue engineering applications. Mater. Today Bio 2022, 16, 100412. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Gong, H.; Quan, X.; Wang, Z.; Hu, X.; Chen, Z.; Li, Z.; Liu, H.; Zhang, L.; McBain, A.J.; et al. Intramembrane Nanoaggregates of Antimicrobial Peptides Play a Vital Role in Bacterial Killing. Small 2023, 19, e2204428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wu, Z.; Zhou, J.; Wang, Z.; Yang, C.; Wang, P.; Fareed, M.S.; He, Y.; Su, J.; Cha, R.; et al. The Antimicrobial, Hemostatic, and Anti-Adhesion Effects of a Peptide Hydrogel Constructed by the All-d-Enantiomer of Antimicrobial Peptide Jelleine-1. Adv. Healthc. Mater. 2023, 8, e2301612. [Google Scholar] [CrossRef]
- Fuertes Llanos, M.; Gómara, M.J.; Haro, I.; Sánchez López, E. Peptide Amphiphiles for Pharmaceutical Applications. Curr. Med. Chem. 2023. [Google Scholar] [CrossRef]
- Cardoso, P.; Appiah Danso, S.; Hung, A.; Dekiwadia, C.; Pradhan, N.; Strachan, J.; McDonald, B.; Firipis, K.; White, J.F.; Aburto-Medina, A.; et al. Rational design of potent ultrashort antimicrobial peptides with programmable assembly into nanostructured hydrogels. Front. Chem. 2023, 10, 1009468. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, E.; Rajasekharan, A.K.; Atefyekta, S.; Andersson, M. Cross-linked lyotropic liquid crystal particles functionalized with antimicrobial peptides. Int. J. Pharm. 2022, 627, 122215. [Google Scholar] [CrossRef]
- Sharma, R.; Lalhall, A.; Puri, S.; Wangoo, N. Design of Fmoc-Phenylalanine Nanofibrillar Hydrogel and Mechanistic Studies of Its Antimicrobial Action against Both Gram-Positive and Gram-Negative Bacteria. ACS Appl. Bio Mater. 2023, 6, 494–506. [Google Scholar] [CrossRef]
- Hamley, I.W. Self-Assembly, Bioactivity, and Nanomaterials Applications of Peptide Conjugates with Bulky Aromatic Terminal Groups. ACS Appl. Bio Mater. 2023, 6, 384–409. [Google Scholar] [CrossRef]
- Dissanayake, S.; He, J.; Yang, S.H.; Brimble, M.A.; Harris, P.W.R.; Cameron, A.J. Flow-Based Fmoc-SPPS Preparation and SAR Study of Cathelicidin-PY Reveals Selective Antimicrobial Activity. Molecules 2023, 28, 1993. [Google Scholar] [CrossRef]
- Cole, T.J.; Parker, J.K.; Feller, A.L.; Wilke, C.O.; Davies, B.W. Evidence for Widespread Class II Microcins in Enterobacterales Genomes. Appl. Environ. Microbiol. 2022, 88, e0148622. [Google Scholar] [CrossRef]
- George, N.L.; Orlando, B.J. Architecture of a complete Bce-type antimicrobial peptide resistance module. Nat. Commun. 2023, 14, 3896. [Google Scholar] [CrossRef]
- Prasad, A.K.; Martin, L.L.; Panwar, A.S. Helical intermediate formation and its role in amyloids of an amphibian antimicrobial peptide. Phys. Chem. Chem. Phys. 2023, 25, 12134–12147. [Google Scholar] [CrossRef]
- Makowska, M.; Kosikowska-Adamus, P.; Zdrowowicz, M.; Wyrzykowski, D.; Prahl, A.; Sikorska, E. Lipidation of Naturally Occurring α-Helical Antimicrobial Peptides as a Promising Strategy for Drug Design. Int. J. Mol. Sci. 2023, 24, 3951. [Google Scholar] [CrossRef] [PubMed]
- Niu, M.; Gu, X.; Yang, J.; Cui, H.; Hou, X.; Ma, Y.; Wang, C.; Wei, G. Dual-Mechanism Glycolipidpeptide with High Antimicrobial Activity, Immunomodulatory Activity, and Potential Application for Combined Antibacterial Therapy. ACS Nano 2023, 17, 6292–6316. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zeng, S.; Wang, Y.; Yin, X.; Zhang, B.; Zhang, B.; Xu, S.; Zhang, Y.; Zheng, J.; Fan, J.; et al. Metal coordinating-induced self-assembly of cyclic lipopeptides into high-performance antimicrobial supramolecules. Food Chem. 2023, 422, 136203. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Ji, J.; Wang, H.; Nangia, S.; Wang, H.; Libera, M. Self-Defensive Antimicrobial Surfaces Using Polymyxin-Loaded Poly(styrene sulfonate) Microgels. ACS Biomater. Sci. Eng. 2022, 8, 4827–4837. [Google Scholar] [CrossRef] [PubMed]
- Thankappan, B.; Thomas, A.; Sakthivadivel, A.; Shanmuganathan, N.; Angayarkanni, J. In vitro and in vivo antimicrobial activity of self-assembled melittin nanoparticles: A comparative study with melittin peptide. Colloids Surf. B Biointerfaces 2023, 226, 113331. [Google Scholar] [CrossRef]
- Yadav, N.; Kumar, U.; Chauhan, V.S. Conformationally restricted, dipeptide-based, self-assembled nanoparticles for efficient vancomycin delivery. Nanomedicine 2022, 17, 2023–2035. [Google Scholar] [CrossRef]
- Yu, W.; Sun, Y.; Li, W.; Guo, X.; Liu, X.; Wu, W.; Yu, W.; Wang, J.; Shan, A. Self-Assembly of Antimicrobial Peptide-Based Micelles Breaks the Limitation of Trypsin. ACS Appl. Mater. Interfaces 2023, 15, 494–510. [Google Scholar] [CrossRef]
- Rashid, R.; Nair, Z.J.; Chia, D.M.H.; Chong, K.K.L.; Cazenave Gassiot, A.; Morley, S.A.; Allen, D.K.; Chen, S.L.; Chng, S.S.; Wenk, M.R.; et al. Depleting Cationic Lipids Involved in Antimicrobial Resistance Drives Adaptive Lipid Remodeling in Enterococcus faecalis. mBio 2023, 14, e0307322. [Google Scholar] [CrossRef]
- Romero-Calle, D.X.; Pedrosa-Silva, F.; Ribeiro Tomé, L.M.; Fonseca, V.; Guimarães Benevides, R.; de Oliveira Santos, L.T.S.; de Oliveira, T.; da Costa, M.M.; Alcantara, L.C.J.; de Carvalho Azevedo, V.A.; et al. Molecular Characterization of Salmonella Phage Wara Isolated from River Water in Brazil. Microorganisms 2023, 11, 1837. [Google Scholar] [CrossRef] [PubMed]
- Sousa, A.; Borøy, V.; Bæverud, A.; Julin, K.; Bayer, A.; Strøm, M.; Johannessen, M.; Škalko-Basnet, N.; Obuobi, S. Polymyxin B stabilized DNA micelles for sustained antibacterial and antibiofilm activity against P. aeruginosa. J. Mater. Chem. B. 2023, 11, 7972–7985. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, Y.; Zhou, Z.; Liu, S.; Mao, S.; Li, G. Co-delivery of EGCG and melittin with self-assembled fluoro-nanoparticles for enhanced cancer therapy. Aging 2023, 15, 4875–4888. [Google Scholar] [CrossRef] [PubMed]
- Manioglu, S.; Modaresi, S.M.; Ritzmann, N.; Thoma, J.; Overall, S.A.; Harms, A.; Upert, G.; Luther, A.; Barnes, A.B.; Obrecht, D.; et al. Antibiotic polymyxin arranges lipopolysaccharide into crystalline structures to solidify the bacterial membrane. Nat. Commun. 2022, 13, 6195. [Google Scholar] [CrossRef]
- Stachurski, O.; Neubauer, D.; Walewska, A.; Iłowska, E.; Bauer, M.; Bartoszewska, S.; Sikora, K.; Hać, A.; Wyrzykowski, D.; Prahl, A.; et al. Understanding the Role of Self-Assembly and Interaction with Biological Membranes of Short Cationic Lipopeptides in the Effective Design of New Antibiotics. Antibiotics 2022, 11, 1491. [Google Scholar] [CrossRef]
- Zhan, W.; Gao, G.; Liu, Z.; Liu, X.; Xu, L.; Wang, M.; Xu, H.D.; Tang, R.; Cao, J.; Sun, X.; et al. Enzymatic Self-Assembly of Adamantane-Peptide Conjugate for Combating Staphylococcus aureus Infection. Adv. Healthc. Mater. 2023, 12, e2203283. [Google Scholar] [CrossRef]
- Dong, L.; Huang, C.; Zhao, B.; Hu, G.; Huang, Y.; Zhang, X.; Hu, X.; Wang, Y.; Sun, X.; Qian, W.; et al. A pH/enzyme dual responsive PMB spatiotemporal release hydrogel promoting chronic wound repair. J. Nanobiotechnol. 2023, 21, 213. [Google Scholar] [CrossRef]
- Zhao, H.; Zhong, L.L.; Yang, C.; Tang, N.; He, Y.; He, W.; Zhao, Z.; Wu, C.; Yuan, P.; Yang, Y.Y.; et al. Antibiotic-Polymer Self-Assembled Nanocomplex to Reverse Phenotypic Resistance of Bacteria toward Last-Resort Antibiotic Colistin. ACS Nano 2023, 17, 15411–15423. [Google Scholar] [CrossRef]
- Zhang, P.; Ouyang, Q.; Zhai, T.; Sun, J.; Wu, J.; Qin, F.; Zhang, N.; Yue, S.; Yang, X.; Zhang, H.; et al. An inflammation-targeted nanoparticle with bacteria forced release of polymyxin B for pneumonia therapy. Nanoscale 2022, 14, 15291–15304. [Google Scholar] [CrossRef]
- Coya, J.M.; Fraile-Ágreda, V.; de Tapia, L.; García-Fojeda, B.; Sáenz, A.; Bengoechea, J.A.; Kronqvist, N.; Johansson, J.; Casals, C. Cooperative action of SP-A and its trimeric recombinant fragment with polymyxins against Gram-negative respiratory bacteria. Front. Immunol. 2022, 13, 927017. [Google Scholar] [CrossRef]
Figure 1.
Approved AMPs for use in clinical settings. STD—sexually transmitted diseases. Reproduced from [4].
Figure 1.
Approved AMPs for use in clinical settings. STD—sexually transmitted diseases. Reproduced from [4].

Figure 2.
Peptide amphiphiles (PAs) synthesized as molecules containing a hydrocarbon chain attached to a peptide segment self-assemble into cylindrical nanostructures capable of enhanced antimicrobial performance. Reprinted with permission from [20]. Copyright 2019 American Chemical Society.
Figure 2.
Peptide amphiphiles (PAs) synthesized as molecules containing a hydrocarbon chain attached to a peptide segment self-assemble into cylindrical nanostructures capable of enhanced antimicrobial performance. Reprinted with permission from [20]. Copyright 2019 American Chemical Society.

Figure 3.
Scheme of antimicrobial coatings on glass based on casting and drying gramicidin (Gr) nanoparticles (NPs) [21] and self–assembled NPs of DODAB bilayer fragments (BF)/carboxymethylcellulose (CMC)/poly (diallyldimethylammonium chloride) (PDDA) [43,44]. The layer–by–layer discoid assemblies of DODAB BF were covered by consecutive layers of CMC and PDDA, yielding the DODAB BF/CMC/PDDA disks in water dispersion. Reproduced from [42].
Figure 3.
Scheme of antimicrobial coatings on glass based on casting and drying gramicidin (Gr) nanoparticles (NPs) [21] and self–assembled NPs of DODAB bilayer fragments (BF)/carboxymethylcellulose (CMC)/poly (diallyldimethylammonium chloride) (PDDA) [43,44]. The layer–by–layer discoid assemblies of DODAB BF were covered by consecutive layers of CMC and PDDA, yielding the DODAB BF/CMC/PDDA disks in water dispersion. Reproduced from [42].

Figure 4.
Issues and solutions required to bring AMPs to the clinic. Reproduced from [60] published by Frontiers.
Figure 4.
Issues and solutions required to bring AMPs to the clinic. Reproduced from [60] published by Frontiers.

Figure 5.
Chemical structure of some natural AMPs in clinical use. Polymyxin B (a); bacitracin A (b); vancomycin (c); gramicidin A (d) as a peptide dimer traversing the membrane and anchoring to the membrane interface by its four Trp residues. Adapted with permission from [64] Elsevier, copyright 2005. Nisin (e) with its unusual amino acids such as dehydroalanine (DHA), dehydrobutyrine (DHB), and several intramolecular thio-ether bridges linking β-methyllanthionine (ABU) to an alanine residue. Reprinted from [65].
Figure 5.
Chemical structure of some natural AMPs in clinical use. Polymyxin B (a); bacitracin A (b); vancomycin (c); gramicidin A (d) as a peptide dimer traversing the membrane and anchoring to the membrane interface by its four Trp residues. Adapted with permission from [64] Elsevier, copyright 2005. Nisin (e) with its unusual amino acids such as dehydroalanine (DHA), dehydrobutyrine (DHB), and several intramolecular thio-ether bridges linking β-methyllanthionine (ABU) to an alanine residue. Reprinted from [65].

Figure 6.
AMPs can act directly by killing microbes and/or by modulating immune responses. Reproduced from Ulm and co-workers [103].
Figure 6.
AMPs can act directly by killing microbes and/or by modulating immune responses. Reproduced from Ulm and co-workers [103].

Figure 7.
Gramicidin A molecules acquire a helix conformation in bilayer membranes (A); Gr dimers function as channels that allow passive diffusion of cations such as Na+ and K+ in accordance with their concentration gradients; thereby, Na+ influx and K+ efflux take place (B). Reproduced from [109].
Figure 7.
Gramicidin A molecules acquire a helix conformation in bilayer membranes (A); Gr dimers function as channels that allow passive diffusion of cations such as Na+ and K+ in accordance with their concentration gradients; thereby, Na+ influx and K+ efflux take place (B). Reproduced from [109].

Figure 8.
Scanning electron micrograph of gramicidin (Gr) dispersions at 0.05 mM Gr in pure water and the effect of gramicidin (Gr) concentration on the physical properties of Gr dispersions in water and 1% trifluoroethanol. Measurements were performed 30 min after preparation of the Gr dispersions. Physical properties of the dispersions were the macroscopic aspect seen in photos, the mean z–average diameter (Dz), the zeta–potential (ζ), and the polydispersity (P). All measurements represent a mean value ± the mean standard deviation. Reproduced from [21].
Figure 8.
Scanning electron micrograph of gramicidin (Gr) dispersions at 0.05 mM Gr in pure water and the effect of gramicidin (Gr) concentration on the physical properties of Gr dispersions in water and 1% trifluoroethanol. Measurements were performed 30 min after preparation of the Gr dispersions. Physical properties of the dispersions were the macroscopic aspect seen in photos, the mean z–average diameter (Dz), the zeta–potential (ζ), and the polydispersity (P). All measurements represent a mean value ± the mean standard deviation. Reproduced from [21].


Figure 9.
Gramicidin D dimers in large bilayer vesicles or bilayer fragments made of dioctadecyldimethylammonium bromide (DODAB) and dipalmitoylphosphatidylcholine (DPPC) 1:1. Reprinted from [122], copyright (2012), with permission from Elsevier.
Figure 9.
Gramicidin D dimers in large bilayer vesicles or bilayer fragments made of dioctadecyldimethylammonium bromide (DODAB) and dipalmitoylphosphatidylcholine (DPPC) 1:1. Reprinted from [122], copyright (2012), with permission from Elsevier.

Figure 10.
Oil-in-water nanoemulsions stabilized by a variety of surfactants evaluated the effect of the surfactant nature and hydrophobic–hydrophilic balance on the permeation across the mucus and the epithelial layers of intestinal mucosa; there was a superior permeation across the intestinal cellular layer for the zwitterionic and polyglycerol surfactants in comparison to polyethylene-glycol based ones (polysorbates 40 and 60). Stabilizers used for the nanoemulsions were phosphatidylcholine, lyso-phosphatidylcholine, polyglycerols, and polyethylene-glycols, whereas the oil phase was caprylic acid. Reprinted from [158].
Figure 10.
Oil-in-water nanoemulsions stabilized by a variety of surfactants evaluated the effect of the surfactant nature and hydrophobic–hydrophilic balance on the permeation across the mucus and the epithelial layers of intestinal mucosa; there was a superior permeation across the intestinal cellular layer for the zwitterionic and polyglycerol surfactants in comparison to polyethylene-glycol based ones (polysorbates 40 and 60). Stabilizers used for the nanoemulsions were phosphatidylcholine, lyso-phosphatidylcholine, polyglycerols, and polyethylene-glycols, whereas the oil phase was caprylic acid. Reprinted from [158].

Figure 11.
Staphylococcus aureus (107–108 CFU/mL) viability, in CFU/mL, after interacting for 1 h with gramicidin (Gr) nanoparticles or other Gr formulations in dioctadecyl dimethylammonium bromide (DODAB) bilayers. Cell viability in the presence of Gr in DODAB–supported bilayers on silica (SiO2/DODAB/Gr) [163]. Gr in DODAB–supported bilayers on polystyrene sulfate (PSS) nanoparticles (PSS/DODAB/Gr) [137] and DODAB bilayer fragments (DODAB BF) [164] or DODAB BF/Gr [135]. In the DODAB BF dispersions, Gr dimers in the channel conformation have been previously described [135,137]. The SiO2/DODAB/Gr stock dispersion was prepared with 2 mg/mL silica, 0.5 mM DODAB, and 0.05 mM gramicidin, yielding Dz = 280 ± 5 nm, p = 0.20 ± 0.02, and ζ = 45 ± 4.
Figure 11.
Staphylococcus aureus (107–108 CFU/mL) viability, in CFU/mL, after interacting for 1 h with gramicidin (Gr) nanoparticles or other Gr formulations in dioctadecyl dimethylammonium bromide (DODAB) bilayers. Cell viability in the presence of Gr in DODAB–supported bilayers on silica (SiO2/DODAB/Gr) [163]. Gr in DODAB–supported bilayers on polystyrene sulfate (PSS) nanoparticles (PSS/DODAB/Gr) [137] and DODAB bilayer fragments (DODAB BF) [164] or DODAB BF/Gr [135]. In the DODAB BF dispersions, Gr dimers in the channel conformation have been previously described [135,137]. The SiO2/DODAB/Gr stock dispersion was prepared with 2 mg/mL silica, 0.5 mM DODAB, and 0.05 mM gramicidin, yielding Dz = 280 ± 5 nm, p = 0.20 ± 0.02, and ζ = 45 ± 4.

Figure 12.
Scheme for a degradable nanogel based on a polyester polymer functionalized with reactive sites convertible into either cross-linkers (green) or carboxylic acid moieties (blue); the charged and cross-linked cores are surrounded by a PEG corona (grey); the polyester network is degradable at physiological pH and ionic strength (right). Reprinted from [178], copyright (2023), with permission from Elsevier.
Figure 12.
Scheme for a degradable nanogel based on a polyester polymer functionalized with reactive sites convertible into either cross-linkers (green) or carboxylic acid moieties (blue); the charged and cross-linked cores are surrounded by a PEG corona (grey); the polyester network is degradable at physiological pH and ionic strength (right). Reprinted from [178], copyright (2023), with permission from Elsevier.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carmona-Ribeiro, A.M. Antimicrobial Peptides and Their Assemblies. Future Pharmacol. 2023, 3, 763-788. https://doi.org/10.3390/futurepharmacol3040047
AMA Style
Carmona-Ribeiro AM. Antimicrobial Peptides and Their Assemblies. Future Pharmacology. 2023; 3(4):763-788. https://doi.org/10.3390/futurepharmacol3040047
Chicago/Turabian StyleCarmona-Ribeiro, Ana Maria. 2023. "Antimicrobial Peptides and Their Assemblies" Future Pharmacology 3, no. 4: 763-788. https://doi.org/10.3390/futurepharmacol3040047