
Increased Production of Taxoids in Suspension Cultures of Taxus globosa after Elicitation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Conditions and Treatments
2.2. Inoculation and Elicitation
2.3. HPLC Analysis of Taxoids
2.4. Statistical Analysis
3. Results
3.1. Cell Culture
3.2. Cell Viability
3.3. Biomass Accumulation
3.4. pH Profiles
3.5. Effect of Elicitors on the Production of 10-Diacetylbaccatin
3.6. Effect of Elicitors on the Production of Cephalomannine
3.7. Effect of Elicitors on Taxol Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chun, F.; Long, J.; Li, Q. Simultaneous identification and determination of major taxoids from extracts of Taxus chinensis cell cultures. J. Chem. Sci. 2007, 62, 1–10. [Google Scholar]
- Ojima, I.; Kumar, K.; Awasthi, D.; Vineberg, J.G. Drug discovery targeting cell division proteins, microtubules and FtsZ. Bioorganic Med. Chem. 2014, 22, 5060–5077. [Google Scholar] [CrossRef]
- Wordeman, L.; Vicente, J.J. Microtubule Targeting Agents in Disease: Classic Drugs, Novel Roles. Cancers 2021, 13, 5650. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, J.; Kohli, K.; Mir, S. Development and validation of RP-HPLC method for analysis of novel self-emulsifying paclitaxel formulation. J. Pharm. Biomed. Anal. 2013, 2, 17–27. [Google Scholar]
- Su, J.; Zhang, Z.; Deng, J. Study on the taxol content in Taxus yunnanensis of different age and different provenance. Eur. J. Res. 2005, 18, 369–374. [Google Scholar]
- Hussain, A.; Ahmed, Q.; Nazir, H. In vitro callogenesis and organogenesis in Taxus wallichiana Zucc. The himalayan yew. Pak. J. Bot. 2013, 45, 1755–1759. [Google Scholar]
- Escrich, A.; Almagro, L.; Moyano, E.; Cusido, R.M.; Bonfill, M.; Hosseini, B.; Palazon, J. Improved biotechnological production of paclitaxel in Taxus media cell cultures by the combined of corona tine and calixarenes. Plant Phys. Biochem. 2021, 163, 68–75. [Google Scholar] [CrossRef]
- Liao, Z.; Chen, M.; Sun, X. Micropropagation of endangered plant species. Methods Mol. Biol. 2006, 318, 179–185. [Google Scholar]
- Khosroushahi, A.; Valizadeh, M.; Ghasempour, A. Improved taxol production by combination of inducing factors in suspension cell culture of Taxus baccata. Cell Biol. Int. 2006, 30, 262–269. [Google Scholar] [CrossRef]
- Linden, J.; Phisalaphong, M. Oligosaccharides potentiate methyl jasmonate-induced production of paclitaxel in Taxus canadensis. Plant Sci. 2000, 158, 41–51. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M. Jasmonic acid signaling pathway in plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed]
- Okada, K.; Abe, H.; Gen-Ichuro, A. Jasmonates induce both defense responses and communication in monocotyledonous and dicotyledonous plantas. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Guo, H.; Zhang, J. Hydrogen peroxide is involved in salicylic acid-elicited rosmarinic acid production in Salvia miltiorrhiza cell cultures. Sci. World J. 2014, 2014, 843764. [Google Scholar] [CrossRef] [PubMed]
- Ransy, C.; Vaz, C.; Lombés, A.; Bouillaud, F. Use of H2O2 to cause oxidative stress, the catalase issue. Int. J. Mol. Sci. 2020, 21, 9149. [Google Scholar] [CrossRef]
- Narayani, M.; Srivastava, S. Elicitation: A stimulation of stress in in vitro plant cell/tissue cultures for enhancement of secondary metabolite production. Phytochem. Rev. 2017, 16, 1227–1252. [Google Scholar] [CrossRef]
- Sarmadi, M.; Karimi, N.; Palazón, J.; Ghassempour, A.; Mirjalili, M. The effects of salicylic acid and glucose on biochemical traits and taxane production in a Taxus baccata callus culture. Plant Physiol. Biochem. 2018, 132, 271–280. [Google Scholar] [CrossRef]
- Nakagawara, S.; Nakamura, N.; Guo, Z. Enhanced formation of constitutive sesquiterpenoid in cultured cells of Liverwort. Plant Cell Physiol. 1992, 34, 421–429. [Google Scholar]
- Ghafoori, R.; Bernard, F.; Abolmaali, S. Improved effect of glutathione on the induction and growth of Taxus baccata L. callus. Ann. Biol. Res. 2012, 4, 1726–1730. [Google Scholar]
- Halliwell, B. Reflections of an aging free radical. Free. Rad. Biol. Med. 2020, 161, 234–245. [Google Scholar] [CrossRef]
- Yukimune, Y.; Tabata, H.; Higashi, Y. Methyl jasmonate induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat. Biotechnol. 1996, 14, 1129–1132. [Google Scholar] [CrossRef]
- Gamborg, O.; Millar, R.; Djma, A. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Farkya, S.; Srivastava, A. Bioprocess considerations for production of secondary metabolites by plant cell suspension cultures. Biotechnol. Bioprocess Eng. 2002, 7, 138–149. [Google Scholar] [CrossRef]
- Fett-Neto, A.; Melanson, S.; Nicholson, S. Improved taxol yield by aromatic carboxylic acid and amino acid feeding to cell cultures of Taxus cuspidate. Biotechnol. Bioeng. 1994, 44, 966–971. [Google Scholar] [CrossRef] [PubMed]
- Wickremesinhe, E.; Arteca, R. Taxus callus cultures: Optimizing growth and production of taxol. J. Plant Physiol. 1994, 144, 183–188. [Google Scholar] [CrossRef]
- Ateacha, D.N.; Koch, U.; Engelhard, C. Direct analysis of alkaloids in natural Cinchona bark and commercial extracts using time-of-flight secondary ion mass spectrometry. Anal. Methods 2018, 10, 950–958. [Google Scholar] [CrossRef]
- Maheshwari, P.; Garg, S.; Kumar, A. Taxoids: Biosynthesis and in vitro production. Biotechnol. Mol. Biol. Rev. 2008, 3, 71–87. [Google Scholar]
- Abbasi, K.; Moghim, S.; Reza, M. Optimization of the basal medium for improving production and secretion of taxanes from suspension cell culture of Taxus baccata L. J. Pharm. Sci. 2012, 20, 54–59. [Google Scholar]
- Yao, T.; Asayama, Y. Animal-cell culture media: History, characteristics, and current issues. Reprod. Med. Biol. 2017, 16, 99–117. [Google Scholar] [CrossRef]
- Isah, T. Stress and defense responses in plant secondary metabolites production. Biol. Res. 2019, 25. [Google Scholar] [CrossRef]
- Gull, A.; Lone, A.; Ul, N.; Wani, I. Biotic and abiotic stresses in plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019; pp. 1–6. [Google Scholar]
- Marsik, P.; Langhansova, L.; Dvorakova, M. Increased ginsenosides production by elicitation of In vitro cultivated Panax Ginseng adventitious roots. J. Med. Aromat. Plants 2014, 3, 39. [Google Scholar]
- Mirjalili, N.; Linden, J. Methyl jasmonate induced production of taxol in suspension cultures of Taxus cuspidata: Ethylene interaction and induction models. Biotechnol. Prog. 1996, 12, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Guo, Z. Effect of methyl jasmonic acid on baccatin III biosynthesis. Tsinghua Sci. Technol. 2006, 11, 363–367. [Google Scholar] [CrossRef]
- Luo, J.; Mu, Q.; Gu, Y. Protoplast culture and paclitaxel production by Taxus yunnanensis. Plant Cell Tiss. Org. 1999, 59, 25–29. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Wu, G.; Fang, Y.; Yang, S. Glutathione metabolism and its implications for health. J. Nutr. 2004, 134, 489–492. [Google Scholar] [CrossRef]
- Lewis-Wambi, J.; Kim, H.; Wambi, C. Buthionine sulphoximine sensitizes antihormone-resistant human breast cancer cells to estrogen-induced apoptosis. Breast Cancer Res. 2008, 10, R104. [Google Scholar] [CrossRef]
- Kim, J.; Yun, J.; Hwang, Y. Production of taxol and related taxanes in Taxus brevifolia cell cultures: Effect of sugar. Biotechnol. Lett. 1995, 17, 101–106. [Google Scholar] [CrossRef]
- Parc, G.; Canaguier, A.; Landre, P. Production of taxane with biological activity by plants and callus culture from selected Taxus genotypes. Phytochemistry 2002, 59, 725–730. [Google Scholar] [CrossRef]
- Yamamoto, S.; Taura, K.; Hayashi, S. Effect of lauryl alcohol on production of taxanes in a suspension callus culture. Solvent Extr. Res. Dev. 2014, 21, 95–101. [Google Scholar] [CrossRef]
- Yamamoto, S.; Ogawa, K.; Hayashi, S. Effect of increased volume fraction of organic solvents on callus growth and taxol production in simultaneous suspension callus culture of Taxus baccata and in situ extraction. Solvent Extr. Res. Dev. 2007, 14, 71–77. [Google Scholar]
- Dornenburg, H.; Knorr, D. Strategies for the improvement of secondary metabolite production in plant cell cultures. Enzym. Microb. Technol. 1995, 17, 674–684. [Google Scholar] [CrossRef]
- Kang, K.; Park, S.; Natsagdorj, U. Methanol is an endogenous elicitor molecule for the synthesis of tryptophan and tryptophan-derived secondary metabolites upon senescence of detached rice leaves. Plant J. 2011, 66, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Fall, R.; Benson, A. Leaf methanol-the simplest natural product from plants. Trends Plant Sci. 1996, 1, 296–301. [Google Scholar] [CrossRef]



{kind=link}
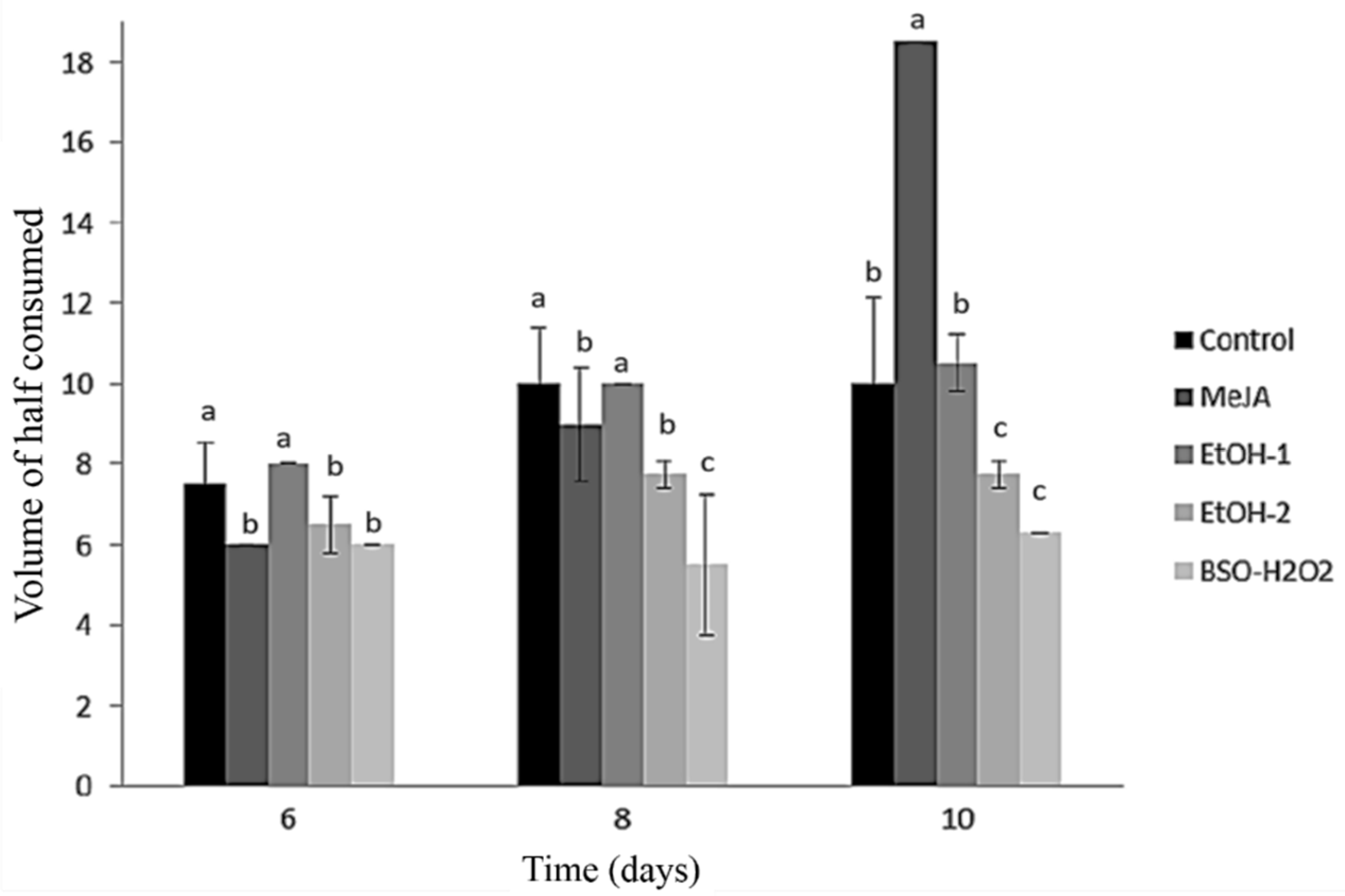{kind=link}
| pH at Six Day | pH at Eight Day | pH at Ten Day | |
|---|---|---|---|
| Control | 5.3 | 6.0 | 5.6 |
| MeJA | 5.7 | 5.8 | 5.8 |
| EtOH-1 | 5.2 | 5.3 | 5.3 |
| EtOH-2 | 5.2 | 5.2 | 5.2 |
| BSO + H2O2 | 6.0 | 5.4 | 6.3 |
| Molecule | Taxoid | Treatment | Day | Production (µg·g−1 of Dry Weight) |
|---|---|---|---|---|
 | 10-deacetylbaccatin | BSO 1 + H2O2 | 6 | 1662.15 a |
| 10-deacetylbaccatin | BSO + H2O2 | 10 | 309.12 b | |
| 10-deacetylbaccatin: C29H36NO10; 544.59 g/mol | 10-deacetylbaccatin | EtOH-1 | 10 | 193.50 b |
 | ||||
| Cephalomannine: C45H53NO14; 831.90 g/mol | Cephalomannine | BSO + H2O2 | 8 | 334.32 a |
 | Cephalomannine | BSO + H2O2 | 10 | 217.69 b |
| Taxol: C47H51NO14; 853.906 g/mol | Taxol | BSO + H2O2 | 6 | 86.78 b |
| Taxol | BSO + H2O2 | 8 | 84.2 b | |
| Taxol | BSO + H2O2 | 10 | 157.0 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barrales-Cureño, H.J.; Ramos Valdivia, A.C.; Soto Hernández, M. Increased Production of Taxoids in Suspension Cultures of Taxus globosa after Elicitation. Future Pharmacol. 2022, 2, 45-54. https://doi.org/10.3390/futurepharmacol2010004
Barrales-Cureño HJ, Ramos Valdivia AC, Soto Hernández M. Increased Production of Taxoids in Suspension Cultures of Taxus globosa after Elicitation. Future Pharmacology. 2022; 2(1):45-54. https://doi.org/10.3390/futurepharmacol2010004
Chicago/Turabian StyleBarrales-Cureño, Hebert Jair, Ana C. Ramos Valdivia, and Marcos Soto Hernández. 2022. "Increased Production of Taxoids in Suspension Cultures of Taxus globosa after Elicitation" Future Pharmacology 2, no. 1: 45-54. https://doi.org/10.3390/futurepharmacol2010004